

Abstract
Cessation of transcription at specific terminator DNA sequences is used by viruses, bacteria, and eukaryotes to regulate the expression of downstream genes, but the mechanisms of transcription termination are poorly characterized. To elucidate the kinetic mechanism of termination at the intrinsic terminators of enteric bacteria, we observed, by using single-molecule light microscopy techniques, the behavior of surface-immobilized Escherichia coli RNA polymerase (RNAP) molecules in vitro. An RNAP molecule remains at a canonical intrinsic terminator for ≈64 s before releasing DNA, implying the formation of an elongation-incompetent (paused) intermediate by transcription complexes that terminate but not by those that read through the terminator. Analysis of pause lifetimes establishes a complete minimal mechanism of termination in which paused intermediate formation is both necessary and sufficient to induce release of RNAP at the terminator. The data suggest that intrinsic terminators function by a nonequilibrium process in which terminator effectiveness is determined by the relative rates of nucleotide addition and paused state entry by the transcription complex.
Significant regulation of DNA transcription, and hence of gene expression, occurs after the initiation of RNA synthesis during elongation of the RNA chain. A principal means by which elongation is regulated is through the action of transcription terminators, DNA sequences that trigger the release of the RNA transcript and DNA template from RNA polymerase (RNAP; refs. 1–4). In general, only a fraction of the RNAP molecules that reach a terminator stop transcription there; the remainder read through the terminator to transcribe downstream DNA sequences. This fraction, called the termination efficiency (TE), is affected by sequences in terminator and nonterminator segments of the DNA and by accessory proteins that bind to the transcription complex (TC). Modulation of TE is an essential genetic control mechanism in the HIV life-cycle and in organisms ranging from bacteria to humans (2). Understanding both termination itself and how regulatory mechanisms modulate termination signals requires knowing what reaction steps occur during termination and the rates of these steps.
At the intrinsic terminators of enteric bacteria, an RNA stem–loop structure and an ≈8-nt uridine-rich sequence that separates it from the 3′ end of the terminated RNA together destabilize the TC (1). The probability of termination is increased when transcription is slow (5, 6), possibly reflecting a kinetic competition between the transcript elongation reaction and processes leading to the dissociation of RNAP from DNA (7). Several studies of intrinsic bacterial terminators have suggested that the TC can enter an altered conformation that is either slowed or blocked in elongation at the terminator (8–10). We will refer to this form of the TC as a paused species, meaning that it is delayed in the process of elongation, the process of release, or both. Thus, termination could occur through direct dissociation of the elongating form of the TC, or it could require the initial formation of a paused species as a kinetically significant intermediate before TC dissociation. However, unambiguous detection of a paused species and demonstration that the species is an authentic intermediate in termination has not yet been achieved. Analysis of the kinetic mechanism of termination in conventional biochemical experiments is complicated by the rapid loss of synchronization in populations of TCs during RNA elongation (11). This problem can be circumvented through the use of techniques that directly observe the behavior of single, isolated RNAP molecules (11).
Methods
DNA Template Construction.
Transcription templates (Fig. 1a) were synthesized by using PCR protocols described previously (12). Templates 574 (template 5 of ref. 13), 572-1B-2, 572-1-2B (12), and 667-1B-2 were used as control templates. pRL667, the PCR template plasmid for synthesis of template 667-1B-2 was made by deletion between BamHI and Tth111I (both sites blunt-ended with the Klenow fragment of DNA polymerase I) in plasmid pRL664. pRL664 was obtained by a similar blunt-end deletion between BspEI and BstEII of plasmid pRL574 (13). The transcription template was prepared by PCR with primers 1B and 2 (12). pRL669, the PCR template plasmid for synthesis of the his template, was made by a BamHI to Tth111I blunt-end deletion of plasmid pRL666, which was made from plasmid pRL651 by blunt-end deletion between BspEI and BstEII. pRL651 was made by ligating the DNA segment 5′-GATCTGGATACCCTGATGCCACAGGATATGATCTCGCGTTCAATTTAAACACCACCATCATCACCATCATCCTGACTAGCTAGCACGCTAGAGAAAGCCCCCGGAAGATGCATCTTCCGGGGGCTTTTTTTTTGGCGCGCGATACAGACC- GGTTCG-3′ (4-nt 5′ overhang; 3′ blunt) between BclI and HpaI of pRL574. The his template was prepared by PCR with primers 1B (12) and 5′-GAAACAGCTATGACCATG-3′. It contains the sequence encoding the termination stem–loop (E:F) and the adjacent run of uracil residues from the Salmonella typhimurium his operon leader region (14), with the insertion of a G residue in the loop of the stem–loop. The DNA sequence encoding the original stem–loop and poly(U) tract triggers termination in vivo; the termination site has been mapped to be between the seventh and the eighth T after the base of the stem (15). The DNA sequence encoding the G-inserted stem–loop and the following poly(U) region is called the “his terminator” in this paper.
Figure 1.

(a) Schematic drawings of DNA templates and the predicted 3′ end secondary structure of a transcript terminated at the his terminator. Crosshatched DNA segments are sequences from the Escherichia coli rpoB gene (13). Control templates lack terminator sequences within 1,800 bp of the promoter. Shown are lengths of DNA segments between the upstream end, transcription start site (bent arrow), and last transcribed nucleotide. The his template contains the his terminator (solid box; see Methods). Shown are segment lengths between the upstream DNA end, transcription start site, primary termination site, and last transcribed nucleotide. Biotin (B) is incorporated at the upstream end of the his template and at either end of control templates. (b) The single-molecule termination experiment (not to scale). Surface-immobilized, stalled TCs are labeled with an avidin-coated bead attached to the biotin-labeled end of the template (12). After addition of nucleoside triphosphates, RNAP moves along the DNA template (elongation), progressively changing the length of the DNA segment between the polymerase and the bead (“tether length”). On reaching the terminator, the enzyme either releases the DNA template and the RNA transcript (termination) or continues elongation past the terminator (read through).
Transcription Reaction in Solution.
RNA products from a single round of transcription were electrophoresed through 4% denaturing polyacrylamide gels as described (9), located by autoradiography, excised, and quantified by counting Cerenkov radiation from the radiolabeled RNA. The macroscopic TE was calculated as
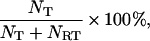 |
1 |
where NT represents the radioactivity in the band of the terminated transcript and NRT is the radioactivity in bands of RNA products from all the complexes that transcribed past the terminator. The reported TE is the average of two independent measurements that differed by <10% of the mean. All experiments were carried out in 100 μM ATP, GTP, CTP, and UTP.
Tethered Particle Motion (TPM) Measurements.
Nanometer-scale movement of single RNAP molecules along template DNAs was observed by using TPM methods as described (12, 13). The elongation rate of an individual TC was measured by linear regression of the tether-length vs. time trace in a window of duration 43–48 s placed at tether lengths ≤792 bp. Measurement of the rate at this short tether length was desirable, because the precision of tether-length measurements worsens with increasing tether length (12). In approximately one-third of the elongating complexes, the tether length exceeded 792 bp before 43 s of data collection; those complexes were excluded from the mean elongation rate measurement, but this exclusion does not affect the mean significantly (data not shown). The 91 complexes used in the elongation rate measurement included 12 complexes that had the tR2 terminator from phage λ ≈1,000 bp downstream of the promoter. Exclusion of these 12 complexes also did not affect the mean elongation rate significantly.
Pauses were taken to be occurring at the terminator if the tether length was between t − w and t + w, where t is the distance between the terminator and the biotin-labeled end of the template and w is the root-mean-square accuracy of the tether-length measurement when the measurement is carried out on a population of complexes with a nominal tether length of t (12). For the his template, this “terminator region” was 792–1,288 bp from the upstream end; the same range of tether lengths was examined in control templates. In 8 of the 105 elongating complexes studied, the bead prematurely released from the surface before the polymerase reached the terminator region; such complexes were excluded from further data analysis.
To determine the pause duration at the terminator, we measured the DNA tether length at 4.8-s intervals (12). The tether-length vs. time records were smoothed with a 5-point mean filter, and the instantaneous elongation rate was computed by numerical differentiation. The polymerase was considered to be pausing at the terminator whenever (i) its location deduced from the smoothed tether-length data was within the terminator region and (ii) the instantaneous elongation rate was less than 1.75 bp⋅s−1, approximately one standard deviation below the measured mean elongation rate. A binary digit called the “pausing index” (p) was assigned to each time point, with p = 1 indicating that the polymerase was pausing at the terminator and p = 0 indicating not pausing. In the record, long sequences in which p = 1 were sometimes interrupted by short sequences of p = 0, presumably because of experimental noise. To reduce underestimation of pause duration caused by these interruptions, p = 0 interruptions of ≤2 points were reassigned the value p = 1 (16). The duration of the longest continuous sequence of p = 1 in the revised record was taken as the “raw pause duration” (traw). traw still overestimates the length of short pauses and underestimates the length of long pauses because of the effects of experimental noise. To correct for this bias, we first measured it by computing traw for simulated data traces with various known pause durations (tknown). For each value of tknown, 100 simulated elongation traces were constructed with an elongation rate of 4.8 bp⋅s−1 and a pause at 1,048-bp tether length, which corresponds to the position of the primary termination site on the his template. This ideal tether-length time course was then averaged in 4.8-s blocks, and Gaussian-distributed noise of amplitude equal to the tether-length measurement precision (12) was added. traw was then computed for each simulated trace by the same algorithm used for the experimental data. A fourth-order polynomial fit to the relation between tknown and the corresponding mean traw was used subsequently to correct the traw values obtained from the experiments, with corrected pause durations constrained to be ≥0. Separate simulations and correction curves were computed for the termination and read-through cases.
The frequency distribution (S) of corrected experimental pause durations (t) was fit by the Levenberg–Marquardt algorithm to an exponential cumulative distribution function:
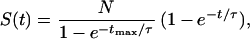 |
2 |
where N represents the total number of duration measurements; tmax = 720 s, the duration of the shortest data records; and τ is the time constant. Of 97 complexes, 6 stopped in the terminator region for >720 s or until the end of observation. These complexes may have lapsed into an arrested conformation, possibly because of backsliding of the polymerase along the DNA–RNA heteroduplex (17–19). Those uncharacteristically long pauses were excluded from the statistical analysis of pause-duration measurements.
RNA Release Measurements.
A linear DNA template containing the same his terminator used in TPM experiments but producing a terminated RNA of 121 nt and a run-off RNA of 212 nt was generated by PCR from pGF104 (20). Stalled A21 TCs (20) were formed by mixing 40 nM template; excess hexahistidine-tagged RNAP either in solution or on Ni2+-NTA agarose beads (Qiagen, Chatsworth, CA); 5 μM 5′-ApUpG-3′ trinucleotide; and 5 μM ATP, GTP, and CTP (Amersham Pharmacia) in transcription buffer (20 mM Tris⋅HCl, pH 7.9/125 mM KCl/1 mM MgCl2/14 mM β-mercaptoethanol/0.1 mM Na2EDTA) with [α-32P]CTP (DuPont; 3,000 Ci/mmol) and then incubating at 25°C for 15 min. Transcription to the terminator was then begun by addition of all four NTPs to 100 μM and heparin to 100 μg/ml.
To measure RNA release from TCs in solution, 30-μl samples were removed at the desired times after NTP addition and filtered through nitrocellulose BA85 filters (Schleicher & Schuell) at 25o. Filters were washed once with 300 μl of transcription buffer. The flow-through and wash fractions were combined, and the RNAs were ethanol precipitated with carrier tRNA and dissolved in 1× stop solution (9). To measure RNA release from immobilized RNAP, samples of supernatants (released RNA) and beads (bound RNA) were collected at the desired times. Supernatants were mixed with equal volumes of 2× stop solution; the beads were washed twice with 100-fold excess of cold transcription buffer and resuspended in 1× stop solution. Samples were analyzed on denaturing 10% polyacrylamide gels. Relative concentrations of RNA species were determined by using a PhosphorImager and imagequant software (Molecular Dynamics). Reported data were obtained from at least three independent experiments; control experiments did not detect measurable RNA release from TCs stalled at nonterminator positions either in solution or on beads.
Results and Discussion
To visualize directly the putative paused intermediate at an intrinsic terminator, we used light microscopy and the TPM method (12, 13) to observe nanometer-scale movement of single RNAP molecules as they transcribed a linear DNA fragment containing a canonical intrinsic terminator (the his terminator from the S. typhimurium his operon leader region; see Methods). In a TPM experiment, TCs are adsorbed to glass slides through RNAP, and transcription is observed by changes in the range of Brownian motion of a visible, submicrometer particle attached to the end of the DNA. Immobilizing the RNAP decreases the diffusion rate of the enzyme and therefore is expected to reduce the rates of the reaction steps in which the TC dissociates (21). Thus, if formation of a paused intermediate precedes DNA dissociation from the TC, TPM should enhance its lifetime and thus facilitate its detection. Further, if dissociation and elongation compete directly, the efficiency of termination should be increased by immobilization. By contrast, if a paused intermediate is no longer capable of elongation, immobilization is not expected to affect TE.
We first constructed biotin end-labeled DNA templates containing a strong E. coli RNAP promoter followed by variable lengths of DNA (Fig. 1a). One template contained the his terminator near the middle of the transcribed sequence, whereas the control templates lacked intrinsic terminators in this region. We then assembled in solution stalled ternary TCs, each of which contained a single RNAP molecule in association with a DNA template and a nascent RNA transcript (12, 13). When transcription by the stalled TCs was restarted in solution by addition of all four NTPs and the RNA products were examined by gel electrophoresis, the measured TE (i.e., the fraction of RNA transcripts that end at the his terminator) was 66%.
For TPM studies, stalled TCs were adsorbed onto the glass surface of a microscope flow cell, and avidin-coated polystyrene beads of 0.2-μm diameter were attached to the biotin-labeled end of the DNA template (Fig. 1b; ref. 12). The density of the TCs on the glass surface was controlled such that each bead had a probability ≤0.2 of being attached to more than one complex (12). On addition of NTPs, the range of Brownian motion of many beads was observed to increase progressively because of transcript elongation and concomitant translocation of DNA through the surface-immobilized enzyme. By calculating the length of the DNA segment between the bead and the immobilized RNAP (Fig. 2) from the measured bead Brownian motion (12), we determined the transcript elongation rate to be 4.8 ± 3.2 bp⋅s−1 (mean ± SD; n = 91) at 0.1 mM NTPs and 23°C, consistent with measurements in previous macroscopic and single-molecule experiments performed under similar conditions (12, 13).
Figure 2.

Example measurements of tether length during transcription of the his terminator template. Each trace represents data from a single TC. The tethered bead detached from the surface at the indicated times (arrows). Dashed lines show the limits of the his terminator region (see Methods). Some complexes read through the terminator region (traces 1 and 2); others terminate without (traces 3 and 4) or with (traces 5–7) a visible pause (brackets) between the cessation of elongation and template release.
During TPM experiments, tethered beads were sometimes observed to detach and diffuse away from the surface. These release events were likely due to release of DNA by RNAP, rather than detachment of RNAP from the glass or detachment of the bead from DNA, because they depended on the presence of the his terminator (see below) and because beads similarly linked to DNA molecules that were directly bound to glass rarely released in observation periods much longer than the experiments reported here (22). About two-thirds of the elongating complexes transcribing the his template released the bead when the tether-length measurement showed that RNAP was in the vicinity of the terminator (Fig. 2, traces 3–7). RNAP molecules that did not release at the terminator ordinarily continued transcription until the enzyme neared the end of the template (Fig. 2, traces 1–2). In contrast, only 4 of 29 complexes transcribing control templates released the bead during transcription of the corresponding region, showing that nearly all of the release events in the terminator region of the his template were caused by the terminator. This result is consistent with the near absence of premature release in previous studies of similar templates lacking intrinsic terminators (12). The TE measured in our TPM experiments, 65 ± 5% (SE; n = 91), agrees well with the 66% value obtained from macroscopic gel electrophoresis experiments. We observed similar agreement between macroscopic and microscopic TEs in experiments with transcription templates containing the phage λ tR2 terminator (data not shown). These results suggest that immobilization of RNAP affects neither its recognition of an intrinsic terminator nor the TE.
Some RNAP molecules were observed to stop DNA translocation when transcribing the his terminator (Fig. 2, traces 5–7). In principle, such a pause could end by the enzyme either releasing the template (termination) or resuming transcript elongation and moving past the terminator (read through). For each complex studied, we used the measured lengths of DNA tethers averaged over 4.8 s intervals (ref. 12; see Methods) to determine the duration of the longest pause in the terminator region. Separate pause-duration histograms were constructed for the groups of complexes that terminated (Fig. 3a) or read through (Fig. 3b). The former had significant pausing: the pause durations of terminating complexes are well fit by a single exponential distribution function with time constant of 67 ± 2 s (SE) and mean pause duration of 64 ± 11 s (Table 1; see refs. 16, 23). In contrast, RNAP molecules that read through the terminator never showed significant pausing; the mean duration and time constant of the pause-length distribution for these complexes were close to the experimental detection limit for pauses and were indistinguishable from those of molecules transcribing the corresponding regions of control (nonterminator) templates (Fig. 3 b and c; Table 1). The barely detectable short pauses in read-through and control complexes may arise from the many weak pause signals not associated with terminators that exist throughout natural gene sequences (24).
Figure 3.

Cumulative frequency distributions of pause durations in the his terminator region (or the equivalent region of control templates). Complexes that terminated in (a) or read through (b and c) the region were analyzed separately. Each point shows the number of observed pauses with durations less than or equal to the indicated duration t. The lines indicate least-squares fit of the data to an exponential distribution function (see Methods) with time constants of 67 s (a), 17 s (b), and 18 s (c).
Table 1.
Transcription termination and pausing at the his terminator
| Template and complex type | No. of observations | Mean pause duration, s* | Pause duration distribution time constant τ, s* |
|---|---|---|---|
| his template | |||
| Termination | 59 | 64 ± 11 | 67 ± 2 |
| Read through | 32 | 16 ± 7 | 17 ± 1 |
| Control template | |||
| Read through | 25 | 16 ± 7 | 18 ± 1 |
Values (± SE) are based on measurements of the longest pause in the vicinity of the terminator (see Methods).
The existence of a lengthy pause in complexes that terminate at the his terminator, but not in those that read through the terminator, implies that the TC can exist in two (or more) distinct chemical states at the terminator (23). Fig. 4 shows the minimal kinetic mechanism that is consistent with these data. In this scheme, RNAP arrives at the terminator in an elongation-competent state E-D-Rn* that can add nucleotide to reach state E-D-Rn+1*. From E-D-Rn*, however, the TC also can undergo a rearrangement to E-D-Rn, a paused state in which RNAP is associated with RNA and DNA but no longer is capable of nucleotide addition. From E-D-Rn, RNAP can in principle either relax back to E-D-Rn* with rate constant k−2 or dissociate with rate constant kt to E + D + Rn. The single-exponential pause distribution of the terminating subpopulation of complexes is consistent with a mechanism in which there is no appreciable template release directly from E-D-Rn*.
Figure 4.

Proposed kinetic mechanism for transcription termination at the his terminator. E, RNAP; D, DNA template; R, RNA transcript. Subscripts n−1, n, or n+1 indicate the length of the transcript in nucleotides, with the terminated transcript having length n. States marked with asterisks (*) are normal intermediates in the elongation reaction; E-D-Rn represents the paused state. For simplicity, the multiple step transcript elongation cycles (11) are shown as single steps with apparent first-order rate constants kn−1 and kn equal to 4.8 s−1. Reaction steps are shown as irreversible when at least one product produced by the step is at negligible concentration in the experiments.
To determine the rate constants in the minimal mechanism, we assume (3) that each cycle of the elongation reaction occurs at the average elongation rate (4.8 nt⋅s−1) under the reaction conditions used here (this value corresponds to kn−1 = kn = 4.8 s−1 in Fig. 4). The measured TE and pause lifetime distributions then determine the values of kt (0.016 s−1) and k2 (9 s−1) in the limiting case where k−2 = 0. With these values, the proposed mechanism predicts a termination pause lifetime distribution mathematically identical to the fit curve in Fig. 3a, with no detectable pause in the read-through population as observed (Fig. 3b). If, in reality, k−2 is greater than zero, k2 must be larger than the limiting case value of 9 s−1 to produce the measured TE. To determine the largest values of k2 and k−2 consistent with the data, we systematically varied these constants in Monte Carlo simulations of lifetime distributions produced by the mechanism. This analysis showed that k−2 must be <0.003 s−1 and k2 must be <11 s−1 (Fig. 4); larger values would produce experimentally detectable pauses in the read-through population.
If the elongation step at the terminator is unusually slow, kn may in reality be as small as 0.056 s−1, because terminator-associated pauses of duration up to ≈18 s are not reliably detected in our experiments (Fig. 3c). Such pausing on the elongation pathway before rearrangement into the termination intermediate (which has been proposed previously; see refs. 9, 10, and 25) would imply a smaller k2. However, it would not affect the basic conclusions that RNAP can exist in two states at a terminator and that the E-D-Rn* → E-D-Rn step is irreversible to the stated limit of detection. Thus, all or nearly all of the TCs that reach the paused state eventually terminate. Consequently, TE is determined in E-D-Rn* by simple competition between the addition of the next nucleotide (kn) and the transition to E-D-Rn (k2). Under the conditions studied, the rate-limiting step of the termination process is template release, which occurs only after the formation of the E-D-Rn paused state. E-D-Rn does not equilibrate with E-D-Rn* on the normal time scale of transcription.
The proposed termination mechanism is consistent with the observation that RNAP immobilization does not alter TE. However, immobilization is expected to slow TC dissociation reactions significantly (21). To confirm that immobilization significantly decreases kt (and thus more stringently tests the hypothesis that E-D-Rn does not equilibrate with E-D-Rn*), we made independent conventional biochemical measurements of the overall rate of RNA release at the his terminator from both immobilized and solution TCs. When TCs containing hexahistidine-tagged RNAP were immobilized on Ni2+-NTA agarose and then allowed to transcribe through the terminator, the rate of terminated RNA release from the agarose (and thus from RNAP) was 0.013 ± 0.001 s−1. However, when assayed from TCs in solution (by binding RNAP to nitrocellulose filters after it encountered the his terminator), RNA release was too fast to measure (>0.5 s−1). The results are consistent with the slowing of TC dissociation expected when the relative diffusion of RNAP and RNA is restricted by enzyme immobilization. (These experiments also imply that release-rate measurements made in vitro with TCs free in solution are of limited applicability in vivo, because the relative diffusion of TC components in living cells is likely to be significantly restricted, for example, by polyribosome decoration of nascent mRNAs.)
In light of the observed effect of immobilization on release rate, the identical TEs measured for free and immobilized TCs imply that RNAP release does not compete directly with transcript elongation at a terminator. Both the macroscopic and single-molecule results support the conclusion that intrinsic termination occurs via a two-step mechanism in which rearrangement of a terminating TC to a paused intermediate is followed by rate-limiting release of nucleic acids (8–10, 26, 27). The similarity between the overall rate of RNA release from immobilized TCs and the single-molecule DNA release rate (kt = 0.016 s−1) is consistent with a mechanism in which RNA and DNA releases from the intermediate occur nearly simultaneously (ref. 28; Fig. 4). A future experiment able to monitor detachment of both RNA and DNA from a single TC (11) could rigorously test this model.
Our results contradict a specific two-step model in which the elongation and paused forms of the complex (analogous to states E-D-Rn* and E-D-Rn in Fig. 4) are in rapid equilibrium, with TE determined by kinetic competition between the nucleotide addition and TC dissociation reactions (3). Rather, the data suggest that entry into the paused state is essentially irreversible at the terminator, with pause entry being the crucial reaction step that competes with nucleotide addition to control TE. A requirement for formation of a paused intermediate before release also is consistent with the observation that nascent RNA release induced by antisense oligonucleotides requires initial formation of a paused TC (25).
Further study will be required to determine the kinetic properties of other intrinsic terminators and the extent to which these properties are modulated by proteins that are bound to RNAP during transcription elongation in vivo (1–5, 9, 10). Full understanding of terminator function will also require elucidation of the structure of the paused intermediate. A variety of different TC structures have been proposed as hypothetical termination intermediates (10, 25, 29). However, the difficulty of distinguishing with conventional methods authentic termination intermediates from nonspecific complexes of RNAP with released RNA or DNA (see ref. 30) makes it unclear whether any of the proposed structures correspond to the irreversibly paused species described here. In particular, Gusarov and Nudler (10) recently isolated at low ionic strength a partially collapsed, “trapped” complex whose properties are consistent with the paused intermediate, but whether this complex lies on the termination pathway is not established. Future single-molecule experiments should contribute to fuller understanding of the mechanism of termination by determining the temporal relationship between RNA and DNA release and by investigating whether escape from the paused intermediate might be possible at other terminators (6, 9).
The mechanisms of transcription regulatory systems are most often analyzed simply by determining the energetics of the relevant protein–nucleic acid interactions. However, that approach is based on the assumption that the various intermediate species interconvert rapidly such that the relative amount of each is dictated only by its thermodynamic stability. The present study shows that this assumption is not necessarily correct even in a simple regulatory system. Termination at his is a nonequilibrium process; understanding the behavior of the system is thus achieved only by defining the individual steps in the termination and read-through pathways and measuring their rates. This study shows the power of single-molecule methods for studying transcription to accomplish such kinetic analysis.
Acknowledgments
We thank Edgar Young and OiKwan Wong for helpful discussions and Rachel Mooney, Chris Miller, and Melissa Moore for comments on the manuscript. This work was funded by grants from the National Institutes of Health to J.G. and R.L.
Abbreviations
- TE
termination efficiency
- TC
transcription complex
- RNAP
RNA polymerase
- TPM
tethered-particle motion
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Roberts J W. In: Regulation of Gene Expression in E. coli. Lin E, Lynch A, editors. Austin, TX: Landes; 1996. pp. 27–44. [Google Scholar]
- 2.Uptain S, Kane C, Chamberlin M. Annu Rev Biochem. 1997;66:117–172. doi: 10.1146/annurev.biochem.66.1.117. [DOI] [PubMed] [Google Scholar]
- 3.von Hippel P H, Yager T D. Science. 1992;255:809–812. doi: 10.1126/science.1536005. [DOI] [PubMed] [Google Scholar]
- 4.von Hippel P. Science. 1998;281:660–665. doi: 10.1126/science.281.5377.660. [DOI] [PubMed] [Google Scholar]
- 5.McDowell J C, Roberts J W, Jin D J, Gross C. Science. 1994;266:822–825. doi: 10.1126/science.7526463. [DOI] [PubMed] [Google Scholar]
- 6.Reynolds R, Bermúdez-Cruz R M, Chamberlin M J. J Mol Biol. 1992;224:31–51. doi: 10.1016/0022-2836(92)90574-4. [DOI] [PubMed] [Google Scholar]
- 7.von Hippel P H, Yager T D. Proc Natl Acad Sci USA. 1991;88:2307–2311. doi: 10.1073/pnas.88.6.2307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Farnham P J, Platt T. Cell. 1980;20:739–748. doi: 10.1016/0092-8674(80)90320-7. [DOI] [PubMed] [Google Scholar]
- 9.Chan C, Wang D, Landick R. J Mol Biol. 1997;268:54–68. doi: 10.1006/jmbi.1997.0935. [DOI] [PubMed] [Google Scholar]
- 10.Gusarov I, Nudler E. Mol Cell. 1999;3:495–504. doi: 10.1016/s1097-2765(00)80477-3. [DOI] [PubMed] [Google Scholar]
- 11.Gelles J, Landick R. Cell. 1998;93:13–16. doi: 10.1016/s0092-8674(00)81140-x. [DOI] [PubMed] [Google Scholar]
- 12.Yin H, Landick R, Gelles J. Biophys J. 1994;67:2468–2478. doi: 10.1016/S0006-3495(94)80735-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schafer D, Gelles J, Sheetz M, Landick R. Nature (London) 1991;352:444–448. doi: 10.1038/352444a0. [DOI] [PubMed] [Google Scholar]
- 14.Johnston H M, Roth J R. J Mol Biol. 1981;145:735–756. doi: 10.1016/0022-2836(81)90312-0. [DOI] [PubMed] [Google Scholar]
- 15.Freedman R, Schimmel P. J Biol Chem. 1981;256:10747–10750. [PubMed] [Google Scholar]
- 16.Colquhoun D, Sakmann B. Nature (London) 1981;294:464–466. doi: 10.1038/294464a0. [DOI] [PubMed] [Google Scholar]
- 17.Komissarova N, Kashlev M. Proc Natl Acad Sci USA. 1997;94:1755–1760. doi: 10.1073/pnas.94.5.1755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nudler E, Mustaev A, Lukhtanov E, Goldfarb A. Cell. 1997;89:33–41. doi: 10.1016/s0092-8674(00)80180-4. [DOI] [PubMed] [Google Scholar]
- 19.Wang M, Schnitzer M, Yin H, Landick R, Gelles J, Block S. Science. 1998;282:902–907. doi: 10.1126/science.282.5390.902. [DOI] [PubMed] [Google Scholar]
- 20.Feng G, Lee D N, Wang D, Chan C L, Landick R. J Biol Chem. 1994;269:22282–22294. [PubMed] [Google Scholar]
- 21.Eigen M, Hammes G G. Adv Enzymol. 1963;25:1–38. doi: 10.1002/9780470122709.ch1. [DOI] [PubMed] [Google Scholar]
- 22.Finzi L, Gelles J. Science. 1995;267:378–380. doi: 10.1126/science.7824935. [DOI] [PubMed] [Google Scholar]
- 23.Colquhoun D, Hawkes A G. In: Single Channel Recording. Sakmann B, Neher E, editors. New York: Plenum; 1983. pp. 135–175. [Google Scholar]
- 24.Levin J R, Chamberlin M J. J Mol Biol. 1987;196:61–84. doi: 10.1016/0022-2836(87)90511-0. [DOI] [PubMed] [Google Scholar]
- 25.Artsimovitch I, Landick R. Genes Dev. 1998;12:3110–3122. doi: 10.1101/gad.12.19.3110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yager T D, von Hippel P H. In: Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology. Neidhardt F C, Ingraham J L, Low K B, Magasanik B, Schaechter M, Umbarger H E, editors. Washington, DC: Am. Soc. Microbiol.; 1987. pp. 1241–1275. [Google Scholar]
- 27.Yager T D, von Hippel P H. Biochemistry. 1991;30:1097–1118. doi: 10.1021/bi00218a032. [DOI] [PubMed] [Google Scholar]
- 28.Arndt K M, Chamberlin M J. J Mol Biol. 1988;202:271–285. doi: 10.1016/0022-2836(88)90457-3. [DOI] [PubMed] [Google Scholar]
- 29.Yarnell W S, Roberts J W. Science. 1999;284:611–615. doi: 10.1126/science.284.5414.611. [DOI] [PubMed] [Google Scholar]
- 30.Berlin V, Yanofsky C. J Biol Chem. 1983;258:1714–1719. [PubMed] [Google Scholar]