

Abstract
Field observations showed that in its native Texas grasslands, dodder (Cuscuta attenuata) grows more vigorously in patches of mixed host species than in monospecific host patches. Field experiments with naturally occurring host individuals demonstrated that the parasite achieved greater volume when simultaneously infesting two hosts of differing species rather than two hosts of the same species, and that this effect depends on the order in which the parasite encounters those hosts. Sodium, acting as an osmoticum, is implicated as the factor responsible for experimentally produced as well as observed field patterns.
All but the smallest annual plant will encounter heterogeneity in minerals and nutrients as it grows (1–4); plants respond locally to changes in resource levels, allowing increased uptake in rich patches and suffering decreased investment in poor patches. For example, encounter with high nitrogen, and, to a lesser extent, high phosphorus levels has been shown to induce localized root proliferation in a number of species (5–8). Similarly, ramets (potentially independent subunits) of some clonal plant species may produce more stolons in “good” patches than in “poor” patches (9–11). Local enrichment with ammonium may physiologically enhance local uptake capacities for ammonium and potassium (12). In some few cases, localized responses to changes in resource levels have been correlated with fitness-related rewards, such as plant volume or seed set (13–16).
However plants may actively exploit resource heterogeneity. Indeed, it has been implied that the spatially disjunct distribution of resources in nature is a selective force behind the evolution of the clonal habit in higher plants (17). However, more than a decade of avid research has failed to reveal any such synergistic effect (e.g., refs. 10 and 18–22). Instead, and consistent with expectations of foraging theory derived from the study of animals, even the most careful experimental studies have shown only that the growth capacity of a plant simultaneously inhabiting more than one patch type (with more than one level or type of macronutrient) is no more than the growth capacity achievable in an average of the patches used by that individual (23).
Direct observation of naturally occurring plant resource uptake is made difficult by the cryptic nature of these generally soil-borne processes. Furthermore, because of naturally varying conditions, the destructiveness of sampling root systems, and the uncertainty of total recovery of the roots of an uncontained plant have prevented correlating plant response unequivocally with individual fitness. Hence, the causes and consequences of plant response to mineral and nutrient heterogeneity in nature have usually been extrapolated from plant growth under experimental manipulation in the laboratory, greenhouse, agricultural field (see, for example, studies cited in ref. 23), from theory (25), or in simulations (24). Thus, the field observations of undisturbed resource use and the field experiments reported here represent an example of data garnered from a natural system with unambiguous identification of resource use and plant growth response. To collect these exceptional data, we used as a model the stem parasite Cuscuta attenuata (Convolvulaceae), commonly known as dodder. For all species in this genus, the entire plant is aboveground, coiling about the stems and branches of its hosts. All points of resource uptake are visible; thus for any individual parasite, all of its resources can be identified, its investment (the length of dodder stem coiled about the host) can be measured, and its resulting growth can be assessed, all without disturbance to the individual. Because dodders are effectively nonphotosynthetic (26, 27), their responses to a resource are not confounded by carbohydrate production and thus are more easily correlated with the action of its resource.
In summary, we have shown that, under natural conditions, use of multiple resource types can enhance the growth of an individual plant beyond that expected from an averaging of the resource types; moreover, such enhancement relies upon a particular set of circumstances, specifically, the sequence of its encounters with resource types differing in sodium content. Furthermore, this response is fitness related, in that the seed set of an individual dodder is highly correlated with its size (13, 14). Our field experiments suggest that, within the parasite, sodium acts as an osmoticum in the extraction of water and possibly other nutrients as the plant grows between hosts that differ in sodium levels. Use of sodium as an osmoticum is not uncommon in nonparasitic plants (28); we propose that the process reported herein may function in any plant large enough to occupy more than one patch at a time, i.e., in the majority of angiosperms and many “lower” plants as well.
Methods and Results
A dodder may infest a host when a growing tip comes in contact with a host stem or leaf; with contact, the parasite stem can coil around the host, after which pegs of absorptive tissue (haustoria) grow into the host from the inner surface of the parasite coil. Through the haustoria a parasite can tap into the host’s vascular tissue and redirect its flow into the otherwise normal vascular system of the parasitic individual. In general, parasitic species maintain the physiological ground plan of their autotrophic ancestors; the parasite uses the haustorial connection to integrate itself into the host’s sink–source dynamics, drawing upon the productivity and reserves of the host in the same manner as that of the host’s own rapidly growing and/or transpiring tissue (29).
C. attenuata is native throughout the old fields of Oklahoma and northern Texas. In June 1996, the period of peak dodder growth in this region, it was observed that parasites infesting patches of mixed host species accumulated more biomass than those growing in monospecific host stands. In five 50-m transects across a 9-hectare field near Dallas, parasite stem density was significantly greater when infesting patches of interdigitated Aster ericoides and the architecturally similar Ambrosia artimisiifolia, than when infesting monocultures of either host (Fig. 1). These differences in biomass accumulation were not explained by differences in the value of an individual species as a host (Fig. 2) or by differences in physical resource availability (host–branch density) within patches (Fig. 1).
Figure 1.

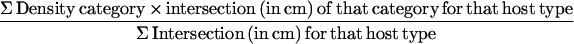 |
Figure 2.

Parasite volume after 10 days on single host individuals. Eighty dodder stems with growing tips were collected from a single parasite individual; all stems were derived from parasite tissue coiled on Aster ericoides. Each detached parasite stem was placed in a water-filled test tube taped to an individual of the target host species. Host individuals were selected for comparability of size, apparent vigor, insolation, and soil qualities. After the parasite stem length and width had been measured, the parasite stem was tied to actively growing host tissue with synthetic string, giving 40 replicates for each host species. Ten days after this transplant event, all surviving parasite stems were measured for length of established coil and length and width of dodder stem acropetal to the coil. There was no significant difference in parasite coil length between the two hosts (t = 1.33; P = 0.1875; n = 54), although parasite volume differed considerably (t = 3.22; P = 0.0025; n = 39).
To determine the possible bases for the observed differences in naturally occurring C. attenuata, detached parasite stems with growing tips were transplanted in the field, first onto one of two host species; after 10 days, the stems were trained onto the other host, of either the same or an alternate species. Successful establishment on a second host signified simultaneous connection to two hosts, a common occurrence under natural conditions. Results showed that parasite stems (approximately equal in length and diameter) attained significantly greater volume when sequentially infesting Ambrosia artimisiifolia followed by Aster ericoides than in any other host–pair sequence (Fig. 3). Stem extension is important in dodders; the two-dimensional, horizontal projection of an individual parasite stem is closely correlated with its number of hosts; both are positively associated with seed crop size (13).
Figure 3.

Parasite volume on two-host configurations. Amb to Ast, parasite stems first transplanted onto Ambrosia artimisiifolia and then trained onto Aster ericoides; Amb to Amb, parasite stems initiated then trained onto Ambrosia artimisiifolia; Ast to Amb, parasite stems initiated on Aster ericoides then trained onto Ambrosia artimisiifolia; Ast to Ast, parasite stems initiated and then trained onto Aster ericoides. These are the same parasite stems that are documented in Fig. 2. Parasite volume differed among treatments (F3, 29 = 4.83; P = 0.0075; an a posteriori Student–Neuman–Keuls test showed all treatments differing from one another at P < 0.05). Haustoria are functional within 4 to 5 days after contact (30). To calculate the rank order of treatments for the various hypotheses, parasite extraction of resources from hosts over the 20-day experimental period was assumed to begin 5 days after the initial transplant; three segments are defined: (i) 5 days immediately after functional establishment on the primary host, with the parasite drawing resources from that host; (ii) 5 days after the second transplant event, but before functional establishment on the second host, with the parasite still drawing resources only from the primary host; (iii) 5 days after establishment on the second host, with the parasite drawing resources from both host individuals. There were no significant differences in parasite coil lengths among the four treatments (F3, 31 = 1.69; P = 0.1891).
Although haustoria are functioning within 5 days of contact with host (30), the rank order of host–pair values is unexplained by the independent value of the hosts to the parasite measured as parasite volume (Figs. 2 and 3). However, comparison of dodder dry weights show no significant difference among treatments (F3, 31 = 1.45; P = 0.25), indicating that the observed differences in parasite volume are primarily a function of different levels of water uptake. Many parasitic plants appear to maintain rates of transpirational flow higher than do those of their hosts (31); dodders are aided in this uptake capacity by a markedly thin or nonexistent waxy cuticle (27). Nonetheless, a model of transpirationally induced flow from host to parasite (estimated as a function of parasite surface area) is not consistent with the observed pattern of treatment-induced parasite volume (Fig. 3).
The mineral contents of hosts and their parasites were therefore analyzed and found to differ significantly in the osmoregulatory ions Na+ and K+, but not in the major nutrients nitrogen or phosphorus (Student–Neuman–Keuls test, P > 0.05 in both cases; Fig. 4). Potassium participates in several roles in the plant, whereas the sole known positive function of sodium is in osmoregulation, in which case it may even substitute for potassium (8). The relative osmotic potential of hosts and parasites may be estimated by summing the concentrations of Na+ and K+ (32). The ratio of the host:parasite ion concentrations thus calculated indicates the flow rate from host to parasite, but does not produce the observed rank order of the treatment results (Fig. 4); using the difference of these host:parasite ion concentrations produces the same result.
Figure 4.

Mineral content of host and parasite. Whole plant tissue from hosts and tissue from parasites were collected from monospecific patches of Ambrosia artimisiifolia and Aster ericoides infested with C. attenuata (n = 4 for each). Samples comprising approximately 5 g of dry tissue from host stems and leaves, or 1 g of dry parasite tissue from each patch were subjected to wet ash analysis at the Texas A & M Extension Service Testing Laboratory. Samples were collected concurrently with the parasite transplant experiment described in Fig. 3.
When considered alone, Na+ differences between hosts and parasites can produce the observed pattern, which we surmise to function as follows. By using the ratio of host and parasite Na+ concentrations to estimate osmotically induced flow rates, the greatest overall uptake between hosts and parasite is predicted for dodder growing from Ambrosia artimisiifolia to Aster ericoides. Because of the high Na+ concentration derived from the primary host, the parasite would be expected to experience a large and immediate water flux from the secondary host. Second, the consistent differences between parasite and hosts when both hosts are the same species would lead to situations in which parasites infesting only Ambrosia artimisiifolia or only Aster ericoides would be second and fourth in volume accumulation, respectively. Last, although a parasite moving from Aster ericoides to the high-Na+ Ambrosia artimisiifolia may suffer an initial loss or decrease in flow from host to parasite, salt accumulators such as C. attenuata (33) are apparently able immediately to begin uptake of Na+ ions, such that sodium concentration may double within 24 hr and triple within 72 hr after exposure to a high-Na+ environment (34). Effective flow from the secondarily infested Ambrosia artimisiifolia host would rapidly begin, accompanied by enhanced flow from the original low-sodium Aster ericoides host, producing a total accumulated volume for this host sequence ranking third in the order of treatments, as was observed.
Discussion
Our results show that, under natural conditions, a plant is able actively to exploit resource patchiness; that is, it can achieve better growth under spatially heterogeneous than it can under homogeneous resource conditions. However, a model system has the capacity to show only what can occur. The value of a model system lies in pointing out the conditions necessary to the process of interest; any claim to generality must show that such conditions also occur in other situations (here, in other species). We note, then, that the implicated mechanism, that of sodium acting as an osmoticum to facilitate water uptake, has been shown to operate in a number of nonparasitic species (28, 35). Additionally, the scale at which resource types vary for dodder is comparable to the scale at which resources vary in the soil habitat (1–4). Thus, plant species beyond that used here, whether parasitic or not, may also possess the conditions necessary for the documented phenomenon.
In essence, the role of sodium in a synergistic response to resource heterogeneity is a subversion of normal cellular expansion processes; i.e., ion sequestration that creates a sink (36, 37) thereby enters into a process fundamental in the plant kingdom. Cellular expansion brought on by hydration is basic to growth and development in all plants, and water acquisition is of value for this reason alone. At the scale of the individual organism (as opposed to the cell), the sort of two-dimensional, horizontal size increase observed here has been shown by simulation to determine the probability of exploitation of any randomly distributed, patchy resource (38). Fieldwork has linked such dodder expansion to the acquisition of hosts (39), whereas in nonparasitic plants, space alone is a patchily distributed, two-dimensional resource (51).
We believe that we have been able to demonstrate a synergism between resource patches for several reasons. First, it is central to our results that we were able to work from observations of naturally occurring plants. The noncryptic nature of resource use in dodders allowed us to identify unequivocally the resources used by an individual and to correlate its responses with particular patterns of resource use. Thus, we could see directly the underlying complexities to which the parasitic plant reacts, integrating them into the simple, but fundamental response of growth, greater or lesser. Although a particular experimental treatment may produce a particular growth pattern in a laboratory environment, other treatment regimes may also produce that particular pattern, and none necessarily reflect that which actually occurs in nature. By deriving our experimental protocol from field observations, we avoided such ambiguity.
The synergism between “good” and “bad” patches that is reported here has been sought intensively in nonparasitic clonal plants for more than a decade, but without obvious success (for review, see refs. 23 and 40). Occupation of more than one nutrient patch is highly likely for any plant other than a small-bodied annual; the expectation of synergistic interaction between resource patches should be true for any plant, clonal or not, inhabiting simultaneously more than one patch. For clonal species, the focus on good vs. bad resource patches has been restricted to the availability of water, light, and macronutrients; sodium was included as only a factor in growth-inhibiting concentrations (41–43). Our findings show how sodium may induce a synergistic dynamic into lifetime patterns of plant resource acquisition. We wonder whether our success in demonstrating an interaction among patch types lies in the nature of the probable process involved. That is, in contrast to the effects of the macronutrients more often examined in the context of spatial heterogeneity (N, P, or K; e.g., see refs. 21, 23, 44), water uptake is relatively rapid and has a direct, unmediated impact on whole-plant expansion through its initiative role in cellular expansion. Whether the potential effect of sodium has to do with the immediacy of these effects relative to the slower, multistep processes of biosynthetic incorporation that the more commonly investigated nutrients experience was not within the scope of this project. We note, however, that rather than merely enhancing the efficiency or retention of resources otherwise acquired, the exploitation of sodium heterogeneity in the habitat has the potential to increase total resource extraction from any particular patch. This effect has the capacity to produce lifetime outcomes differing significantly from the consequences of balanced diets or stepwise limiting of nutrients (45, 46).
We emphasize that the effect depends on the order of uptake and the differential between and among the various levels of sodium in differing patch types. The effect of encounter order, combined with the lack of control a plant (or any organism with limited mobility) has over the order in which it encounters resources, introduces a previously unsuspected element of chance into the survival of competing individuals at early, more sensitive stages of their life cycle. Interactions between nutrient patches could mean that the greatest lifetime acquisition levels (as well as the correlated reproduction and survivorship) of a seed germinating in what might be a bad patch, would be, if followed by entrance of the plant body into a second patch type, transformed into a necessary part of the “best” resource schedule. Even so, the dodder maintains intermediate sodium levels when drawing resources from a large number of hosts encountered in no particular order, apparently acquiring sufficient resources at levels that result in greater growth if resources are mixed, rather than singular. Although treatments dealing with inter- and intraspecific competition have produced insight-yielding results by assuming that plant individuals are battling over the possession of a single patch of light (ref. 47; but see also ref. 48), our results suggest that the inclusion of multiple patch types in modeling the nutrient-based dynamics of competition between individuals large enough to inhabit more than a single patch may add additional depth to the discussion.
Last, these results also apply specifically to parasitic plants. Parasites such as dodders [and the even more notorious witchweed (Striga)] that can infest several hosts simultaneously are capable of considerable economic damage to agricultural crops (29). The relative sodium content of the parasitic plants, whether root or stem, holo- or hemiparasites (26, 31, 32, 49, 50), examined thus far is greater than that of their hosts, regardless of parasite taxonomic affinity. Parasitic plants, like other plants, spend most of any 24-hour period with closed stomata and no transpirational flux. However, the details of uptake dynamics as described here have not been largely considered. Our results suggest a means by which osmoregulatory dynamics may contribute significantly to extraction of resources from hosts. Understanding the function of relative salt content in the uptake from multiple hosts could provide an a priori criterion when choosing low-sodium intercrops (trap plants) with the potential to mitigate the destructiveness of the host–parasite relationship. Exploitation of the dynamic described here could minimize the economic and environmental costs entailed in the chemical and/or manual control of these parasites (29).
Acknowledgments
We thank F. I. Woodward, T. R. E. Southwood, and P. H. Harvey for comments. This research was supported by a National Science Foundation grant (DEB 9306179) to C.K.K.
References
- 1.Snaydon R W. J Ecol. 1962;50:133–143. [Google Scholar]
- 2.Jackson R B, Caldwell M M. Ecology. 1993;74:612–614. [Google Scholar]
- 3.Jackson R B, Caldwell M M. J Ecol. 1993;81:683–692. [Google Scholar]
- 4.Gross K L, Pregitzer K S, Burton A J. J Ecol. 1995;83:357–367. [Google Scholar]
- 5.Philipson J J, Coutts M P. J Exp Bot. 1977;28:864–871. [Google Scholar]
- 6.Drew M C, Saker L R. J Exp Bot. 1978;26:79–90. [Google Scholar]
- 7.Drew M C, Saker L R. J Exp Bot. 1978;29:435–451. [Google Scholar]
- 8.Marschner H. Mineral Nutrition of Higher Plants. London: Academic; 1993. [Google Scholar]
- 9.Salzman A G. Science. 1985;228:604–605. doi: 10.1126/science.3983647. [DOI] [PubMed] [Google Scholar]
- 10.Slade A J, Hutchings M J. J Ecol. 1987;75:95–112. [Google Scholar]
- 11.Kelly C K. Evol Ecol. 1994;8:459–470. [Google Scholar]
- 12.Jackson R B, Caldwell M M. Plant Soil. 1991;138:231–238. [Google Scholar]
- 13.Kelly C K. Host Use and Foraging in the Parasitic Plant Cuscuta subinclusa. Tucson: Univ. of Arizona; 1988. [Google Scholar]
- 14.Kelly C K. Ecology. 1990;71:1916–1925. [Google Scholar]
- 15.Kelly C K. Proc Natl Acad Sci USA. 1992;89:12194–12197. doi: 10.1073/pnas.89.24.12194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dudley S A, Schmitt J. Am Nat. 1996;147:445–465. [Google Scholar]
- 17.Pitelka L F, Ashmun J W. In: Population Biology of Clonal Organisms. Jackson J B C, Buss L W, Cook R E, editors. New Haven, CT: Yale Univ. Press; 1985. pp. 399–436. [Google Scholar]
- 18.Hutchings M J, deKroon H. Adv Ecol Res. 1994;25:159–238. [Google Scholar]
- 19.Alpert P, Mooney H A. Oecologia. 1986;70:227–233. doi: 10.1007/BF00379244. [DOI] [PubMed] [Google Scholar]
- 20.Friedman D, Alpert P. Oecologia. 1991. 76–80. [DOI] [PubMed] [Google Scholar]
- 21.Alpert P. Ecology. 1991;72:69–80. [Google Scholar]
- 22.Alpert P. J Ecol. 1996;84:395–406. [Google Scholar]
- 23.Lötscher M, Hay M J M. J Ecol. 1997;85:341–350. [Google Scholar]
- 24.Ryel R J, Caldwell M M. Ecology. 1998;79:2735–2744. [Google Scholar]
- 25.Caraco T, Kelly C K. Ecology. 1991;72:81–93. [Google Scholar]
- 26.Jeschke W D, Bäumel P, Räth N, Czygan F-C, Proksch P. J Exp Bot. 1994;45:801–812. [Google Scholar]
- 27.Kuijt J. The Biology of Parasitic Flowering Plants. Berkeley, CA: Univ. of California Press; 1969. [Google Scholar]
- 28.Brownell P F. Adv Bot Res. 1979;7:117–224. [Google Scholar]
- 29.Parker C, Riches C R. Parasitic Weeds of the World: Biology and Control. Wallingford, U.K.: CABI; 1993. [Google Scholar]
- 30.Tsivion Y. Bot Gaz. 1978;139:27–31. [Google Scholar]
- 31.Stewart G R, Press M C. Annu Rev Plant Physiol Mol Biol. 1990;41:127–151. [Google Scholar]
- 32.Popp M. In: Parasitic Flowering Plants, (Proceedings 4th Symp.), Weber H C, Forsteuter W, editors. Marburg, Germany: Philipps Universität; 1987. pp. 621–630. [Google Scholar]
- 33.Flowers T J, Hajibagheri M A, Clipson N J W. Q Rev Biol. 1986;61:313–337. [Google Scholar]
- 34.McNulty I B. Plant Physiol. 1985;78:100–103. doi: 10.1104/pp.78.1.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ungar I A. Ecophysiology of Vascular Halophytes. Boca Raton, FL: CRC Press; 1991. [Google Scholar]
- 36.Salisbury F B, Ross C W. Plant Physiology. Belmont, CA: Wadsworth; 1997. [Google Scholar]
- 37.Larcher W. Physiological Plant Ecology. London: Springer; 1995. [Google Scholar]
- 38.Rubin J A. Oecologia. 1987;72:46–51. doi: 10.1007/BF00385043. [DOI] [PubMed] [Google Scholar]
- 39.Kelly C K, Venable D L, Zimmerer K. Oikos. 1988;53:315–320. [Google Scholar]
- 40.Kelly C K. Evol Ecol. 1995;9:575–585. [Google Scholar]
- 41.Salzman A G, Parker M A. Oecologia. 1985;65:273–277. doi: 10.1007/BF00379229. [DOI] [PubMed] [Google Scholar]
- 42.Evans J P, Whitney S. Am J Bot. 1992;79:1344–1347. [Google Scholar]
- 43.Shumway S W. Ann Bot. 1995;76:225–233. [Google Scholar]
- 44.Jackson R B, Manwaring J H, Caldwell M M. Nature (London) 1990;344:58–60. doi: 10.1038/344058a0. [DOI] [PubMed] [Google Scholar]
- 45.Raubenheimer D, Simpson S J. Nutr Res Rev. 1997;10:1–29. doi: 10.1079/NRR19970009. [DOI] [PubMed] [Google Scholar]
- 46.Houston A, Clark C, McNamara J, Mangel M. Nature (London) 1988;332:29–34. [Google Scholar]
- 47.Pacala S W, Tilman D. Am Nat. 1994;143:222–257. [Google Scholar]
- 48.Schwinning S, Weiner J. Oecologia. 1998;113:447–455. doi: 10.1007/s004420050397. [DOI] [PubMed] [Google Scholar]
- 49.Ehleringer J R, Schulze E-D. Am J Bot. 1985;72:568–571. [Google Scholar]
- 50.Goldstein G, Rada F, Sternberg L, Burguera J L, Burguera M, Orozco A, Montilla M, Zabala O, Azocar A, Canales M J, et al. Oecologia. 1989;78:176–183. doi: 10.1007/BF00377153. [DOI] [PubMed] [Google Scholar]
- 51.Yodzis P. Nature (London) 1976;264:540–541. doi: 10.1038/264540a0. [DOI] [PubMed] [Google Scholar]