

Abstract
Histones H3 and H4 have a well defined structural role in the nucleosome and an established role in the regulation of transcription. We have made use of a microinjection strategy using Xenopus embryos to define the minimal structural components of H3 and H4 necessary for nucleosome assembly into metazoan chromosomes in vivo. We find that both the N-terminal tail of H4, including all sites of acetylation, and the C-terminal α-helix of the H4 histone fold domain are dispensable for chromatin assembly. The N-terminal tail and an N-terminal α-helix of H3 are also dispensable for chromatin assembly. However, the remainder of the H3 and H4 histone folds are essential for incorporation of these proteins into chromatin. We suggest that elements of the histone fold domain maintain both nucleosomal integrity and have distinct functions essential for cell viability.
Histones H3 and H4 are highly conserved through eukaryotic evolution (1, 2), implying essential structural and functional roles. These histones exist in a precise stoichiometry within chromatin (3, 4) and wrap DNA to provide an essential architectural framework for the nucleosome (5). Histone H4 heterodimerizes with histone H3 via interaction of their C-terminal histone fold domains (6, 7). Each histone fold consists of three α-helices, the juxtaposition of which within the heterodimer creates three binding sites for the DNA wrapped within the nucleosome (6–8). The histone fold has recently been recognized as a feature of several transcriptional regulatory proteins, including components of TFIID (9, 10) and NF-Y (11). Like the histones, these regulatory proteins make use of the histone fold as a dimerization motif and then have additional domains with distinct structural and regulatory functions (12, 13).
Lying toward the outside of the nucleosome are the N-terminal tails of histone H3 and H4. The N-terminal tails are as conserved through evolution as the C-terminal histone fold domains and have been proposed to be involved in the regulation of transcription (14, 15). The N-terminal tails of histones H3 and H4 are individually dispensable for growth in Saccharomyces cerevisiae and have redundant functions in nucleosome assembly (16, 17). Point mutations or deletions of basic amino acids within the N-terminal tail of H4, including lysine 16, a site for acetylation, influence gene expression, replication, and nuclear division (18–20). Removal of the N-terminal tail domains from H3 and H4 leads to an increase or decrease, respectively, in the transcription of a specific gene, GAL1 (21). A nonbasic domain (residues 21–29) within the H4 tail is also required for repression of the silent mating loci (22). Both the H4 and H3 tails interact with proteins such as SIR3, which are components of heterochromatin in yeast (23, 24). However, the exact roles of the H3 and H4 tails in nucleosome and heterochromatin assembly have not been defined in yeast or any other system.
Modification of the N-terminal tail of H4 by acetylation has been correlated with both transcription (25, 26) and nucleosome assembly (27, 28). Direct evidence for a causal role for H4 acetylation in these events is, however, yet to be established. Tryptic removal of the N-terminal tail of histone H4 from the nucleosome or acetylation of the tail does not influence the helical periodicity of DNA in the nucleosome but might influence the exact path taken by the double helix as it wraps around the histone fold domains of the core histones (29, 30). Thus, the H4 histone tail is not essential for nucleosomal integrity. However, these experiments do not address which portions of histone H4 are necessary for the chaperone-mediated assembly of chromatin known to occur in vivo (31, 32).
We have now determined the sequences of histones H3 and H4 necessary for efficient assembly into the chromatin of metazoan chromosomes in vivo. Our strategy is to microinject mRNA encoding mutant H3 or H4 molecules containing an epitope tag into fertilized Xenopus eggs. Three hours later, following the subsequent development of the embryo to the morula stage, we isolate chromatin and nucleosomes from the embryonic nuclei. Histone containing the epitope tag, synthesized in the embryo, is examined for incorporation into nucleosomes. This requires the exogenous newly synthesized tagged histone to compete effectively for assembly into chromatin with the large stores of endogenous histone stored in the Xenopus egg and the newly synthesized H4 derived from stores of maternal mRNA (33). Thus, we define the requirements for the stable assembly of histone H3 and H4 into chromatin in vivo.
MATERIALS AND METHODS
Plasmids.
All plasmid constructs were cloned by PCR from plasmids pBSXlh3.EA and pBSX1h4.EA (derived from plasmids Xlh.3 and Xlh.4; ref. 34). All 5′ primers were designed with a HindIII site (preceded by an 11-nt sequence to enhance restriction digestion for cloning), an initiating AUG preceded by the sequence AAAG to enhance translation efficiency, and a 24-nt sequence coding for an eight-amino acid FLAG epitope (IBI); following this “common” sequence were histone H3 or H4 gene sequences designed to yield wild-type or 5′ deletion mutants after PCR. All 3′ primers included an XbaI site (again preceded by a sequence to enhance restriction digestion), followed by a stop codon and histone H3 or H4 sequences designed to yield wild-type or 3′ deletion mutants after PCR. After PCR, fragments were cloned into the HindIII and XbaI sites of pSP64pA (Promega). Point mutations were introduced by PCR. Microinjection of embryos followed by in vitro transcription of EcoRI linearized plasmids with Sp6 RNA polymerase (35).
Microinjection.
Microinjection of embryos was as described (35), except that 32 nl of solution containing approximately 50 ng of RNA, [α-32P]dCTP, [3H]arginine, and [3H]lysine were injected; embryos were maintained in 1 × MMR (100 mM NaCl/2 mM KCl/2 mM CaCl2/1 mM MgCl2/5 mM Hepes, pH 7.4)/5% Ficoll and were not transferred to water after injection. Forty-five to fifty embryos were harvested 3 hr after injection and stored at −70°C until analysis for incorporation of histone mutants into chromatin.
Chromatin Purification.
For chromatin analysis, embryos were homogenized by pipetting in micrococcal nuclease digestion buffer (80 mM NaCl/10 mM Tris·HCl, pH 7.5/2 mM CaCl2/25% glycerol) using 14 μl of buffer per embryo. Micrococcal nuclease (Worthington) (0.5–0.8 μl per embryo of a 10 units/μl stock) was added, and digestion was allowed to proceed at room temperature for 10–20 min. EDTA and phenylmethylsulfonyl fluoride were then added to 20 and 1 mM, respectively. Samples were dialyzed against 1 mM EDTA for 3 hr at 4°C and then microcentrifuged for 15 min at 4°C. The supernatant was applied to a 5–20% sucrose gradient and centrifuged at 38,000 rpm in an SW41 rotor for 16 hr at 4°C. After fractionation, 50-μl aliquots of alternate fractions were trichloroacetic acid-precipitated onto glass microfiber filters (Whatman) and Cerenkof-counted to identify the [32P]DNA peak corresponding to nucleosome-containing fractions. Pooled fractions were dialyzed 2 hr against 1 mM EDTA at 4°C. An aliquot (one-fourth to one-third of the total) was removed to confirm by DNA gel electrophoresis that the pooled fractions contained only nucleosome-sized DNA. To the remainder, BSA was added as carrier. Protein was precipitated with 25% trichloroacetic acid overnight at 4°C and collected by microcentrifuging for 10 min at 4°C. Pellets were washed once with acetone/0.2% HCl and once with neat acetone and were then dissolved in 21 μl of 8 M urea. Histone synthesis was quantitated relative to wild-type histone by radiolabeling and immunoblotting (ECL detection) for known loadings of protein mass onto 18% Tris–glycine–SDS gels for electrophoresis.
RESULTS
Mutations of the H4 N-Terminal Tail.
Initially, we examined the synthesis of epitope-tagged, wild-type histone H4 and N-terminal deletions by radiolabeling proteins in Xenopus embryos. Preliminary radiolabeling experiments indicated that epitope-tagged, wild-type histone H4 was incorporated into nucleosomes with an identical efficiency to newly synthesized histone H4 lacking the epitope tag (Table 1). In addition, in vitro reconstitution with epitope-tagged histones H3 and H4 demonstrated that the presence of the N-terminal extensions does not disturb the structure of the nucleosome core (H.K. and A.P.W., data not shown). Purification of nucleosomal material (Fig. 1) followed by electrophoresis of the proteins in these fractions demonstrated that significant levels of newly synthesized tagged exogenous H4 (Fig. 2A, lanes 2–5) were incorporated into chromatin in approximate stoichiometry with the other newly synthesized endogenous core histones (Fig. 2A, lanes 6–9, arrowheads). That these proteins contained the N-terminal epitope tag was confirmed by immunoblotting (Fig. 2A, lanes 10–14). We conclude that deletion of the N terminus, including amino acid 28, does not prevent the efficient incorporation of histone H4 into chromatin (see quantitation in Table 1). This deletion includes all of the sites of histone acetylation (15). Thus, an acetylated N-terminal tail on histone H4 is not essential for incorporation into a nucleosome during chromatin assembly in metazoans.
Table 1.
Assembly of mutant histones into chromatin
| Histones | Expression*, arbitrary unit | % assembled† |
|---|---|---|
| Histone H4 | ||
| WT | 1 | 100 |
| N16 | 0.66 | 77 |
| N28 | 0.43 | 55 |
| N32 | 2.75 | 43 |
| N34 | 3.95 | 22 |
| N36 | 3.1 | 12 |
| N45 | 0.65 | 0 |
| C70 | 1.8 | 24 |
| C75 | 1.2 | 42 |
| C80 | 1.0 | 84 |
| 60-63G | 0.78 | 0 |
| 35,36A | 2.9 | 85 |
| Histone H3 | ||
| WT | 1 | 100 |
| N63 | 1.1 | 36 |
| C121 | 0.33 | 0 |
| 65-68E | 1.4 | 63 |
| 79,83A | 2.2 | 65 |
| 97-100G | 1.6 | 48 |
| 109-112G | 3.2 | 11 |
There is an approximate error of ± 5% on these determinations. These results represent the average values of two experiments.
Expression relative to histone wild type = expression of mutant tagged histone in embryo/expression of wild type tagged histone in embryo.
% assembled into chromatin = (expression of mutant tagged histone in chromatin/expression of wild type tagged histone in chromatin)/(expression of mutant tagged histone in embryo/expression of wild type tagged histone in embryo).
Figure 1.
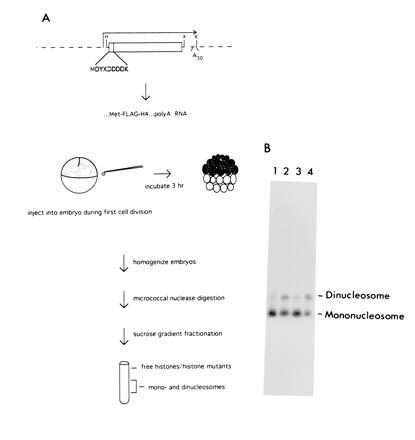
Strategy for determining histone domains required for assembly into replicating chromatin. (A) Wild-type or mutant histone H3 or H4 genes containing N-terminal FLAG epitope sequences (outlined), preceded by an initiating methionine, are subcloned into the HindIII (H)/XbaI (X) sites of transcription vector pSP64pA. These constructs contain an Sp6 promoter upstream and a stretch of 30 A residues (A30) downstream of the inserts. Linearization with EcoRI (R), followed by transcription with Sp6 RNA polymerase, yields a polyadenylylated transcript (mRNA) coding for an epitope-tagged wild-type or mutant H4 protein. This mRNA is injected into Xenopus embryos during the first cell division and incubated for 3 hr. Injected transcripts are vigorously translated into wild-type or mutant histones during this time. To test whether these histones are incorporated into replicating chromatin during this phase of intense cell division, embryos are homogenized and digested with micrococcal nuclease to release nucleosomes. Mono- and dinucleosomes are purified away from free histones by sucrose gradient centrifugation. (B) An aliquot is removed for DNA analysis to confirm the presence of nucleosomes; a typical autoradiogram is shown. The protein composition of the nucleosome fractions is then analyzed for the presence or absence of epitope-tagged histones.
Figure 2.
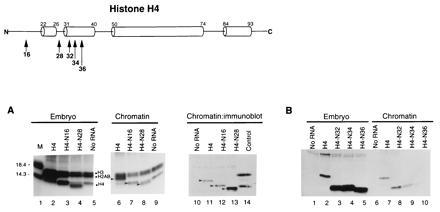
The amino terminal tail of histone H4 is dispensable for assembly into replicating chromatin, but the N-terminal α-helix is important. (A) mRNA encoding epitope-tagged, wild-type (H4; lanes 2, 6, and 11) or N-terminal deletion mutants 1–16 (N16; lanes 3, 7, and 12) and 1–28 (N28; lanes 4, 8, and 13) or no RNA (lanes 5, 9, and 10) were injected into embryos, along with tritiated arginine and lysine to radiolabel proteins and [α-32P]dCTP to label DNA. Embryos were assayed for levels of total newly synthesized radiolabeled histone (lanes 2–5) and for levels of newly synthesized radiolabeled histone incorporated into chromatin (lanes 6–9) by gel electrophoresis followed by fluorography. Filled squares indicate positions of labeled endogenous core histones; arrows indicate positions of labeled epitope-tagged, wild type and mutant histones. The level of epitope-tagged, wild-type and mutant histones were determined by immunoblotting (lanes 11–14). Lane 1 is a molecular weight marker from BRL (Mr 14,300; 18.4 kDa); lane 14 includes radiolabeled wild-type histones H3 and H4. (B) mRNA encoding epitope-tagged, wild-type (H4; lanes 2 and 7) or N-terminal deletion mutants NΔ1-32 (N32; lanes 3 and 8), NΔ1-34 (N36; lanes 4 and 9), NΔ1-36 (N36; lanes 5 and 10), or no RNA (lanes 1 and 6) were injected as above. Levels of epitope-tagged, wild-type and mutant histones in embryos (lanes 1–5) and chromatin (lanes 6–10) were determined by immunoblotting. The immunoreactive bands above those due to the epitope-tagged H4 correspond to non-histone proteins that crossreact with the FLAG epitope that copurify with nucleosomes.
Our next experiments continued the deletion of the N-terminal tail of histone H4 into the first α-helix of the histone fold domain (6–8). This α-helix is part of a DNA binding surface within the nucleosome, the “paired ends of α-helices” (7), and is proposed to be involved in protein-protein interactions with histone H3 (6). We find that deletion of the amino acids comprising this α-helix leads to a progressive decline in the efficiency with which histone H4 is assembled into the nucleosome (Fig. 2B; Table 1). These results suggest that the inviability of yeast strains in which amino acids 4–34 of histone H4 have been deleted (16) is due to the failure of the mutant protein to be stably incorporated into the nucleosome.
Mutations of the H4 Histone Fold Domain.
Grunstein and colleagues (16) found that a series of amino acid deletions from the C-terminal α-helix of the histone fold domain and flanking regions of histone H4 (amino acids 83–102) rendered yeast inviable. Surprisingly, our next experiments determined that a mutant histone H4 entirely lacking this C-terminal α-helix was efficiently incorporated into chromatin (Fig. 3, lanes 3 and 7; Table 1). As a control, we show that an N-terminal deletion of amino acids 1–45 is synthesized in embryos but not incorporated into chromatin (Fig. 3, lanes 4 and 8). Therefore, the C-terminal α-helix of histone H4 is dispensable for nucleosome assembly, and we propose that it has a distinct structural or functional role within the nucleosome subsequent to the assembly process.
Figure 3.
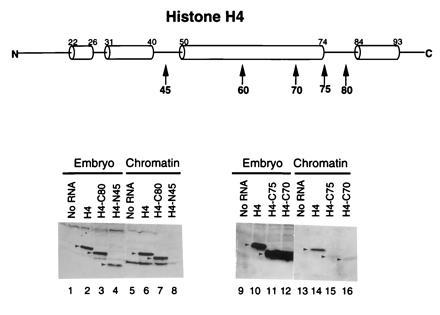
The C-terminal α-helix of histone H4 is dispensable for nucleosome assembly. As in Fig. 2B, Lanes 1–4 and 9–12, protein levels in the unfractionated embryo; lanes 5–8 and 13–16, protein levels in chromatin. Lanes 1, 5, 9, and 13, no RNA injected; lanes 2, 6, 10, and 14, wild-type H4 RNA; lanes 3 and 7, C-terminal deletion mutant CΔ80-102; lanes 4 and 8, N-terminal deletion mutant NΔ1-45; lanes 11 and 15, CΔ75-102; lanes 12 and 16, CΔ70-102.
Amino acids 75–83 in histone H4 lie between the two C-terminal α-helices of the histone fold domain (6–8). This positively charged region of the protein forms a β-structure and provides a DNA interaction surface in the H3-H4 heterodimer (6–8). Removal of amino acids 75–102 greatly reduces the efficiency of histone H4 assembly into the nucleosome (Fig. 3, lanes 11 and 15; Table 1) compared with the wild-type protein (Fig. 3, lanes 10 and 14). This DNA binding site, therefore, appears to have an important role in nucleosome assembly. Further deletion of C-terminal amino acids 70–102 does not significantly influence the low level of assembly of the mutant H4 protein into the nucleosome (Fig. 3, lanes 12 and 16). More extensive deletions from the C terminus (amino acids 60–102) or N terminus (amino acids 1–75) do not lead to the stable accumulation of the mutant H4 proteins in embryos (data not shown). Thus, it is not possible to ascertain their incorporation into chromatin.
We wished to assess the importance of substitution mutations rather than gross deletions of histone H4. The long central α-helix of the H4 histone fold domain extends from amino acids 50 to 74 (6–8). It is proposed to have an essential role in heterodimerization with H3. We substituted amino acids 60–63 (VFLE) with glycine residues to break the integrity of the helix. This mutant is synthesized in embryos (Fig. 4, lane 3) but is not incorporated into chromatin (Fig. 4, lane 8), consistent with an essential role. We next examined whether amino acids 35 and 36, a pair of arginine residues within the essential N-terminal α-helix of the histone fold were essential for the assembly of histone H4 into chromatin. Our results indicate that a mutant histone H4 containing alanine at amino acids 35 and 36 is synthesized in embryos (Fig. 4, lane 5) and is efficiently assembled into chromatin (Fig. 4, lane 10). Thus, the arginines in this α-helix are not essential for chromatin assembly.
Figure 4.
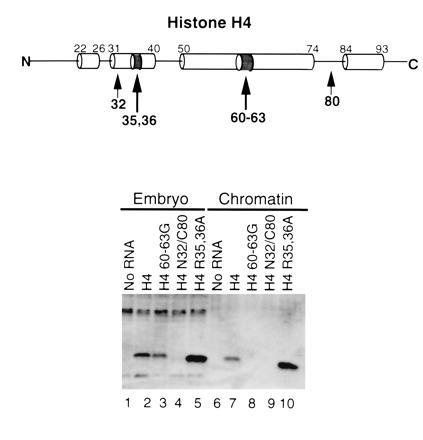
C-terminal DNA contacts and the long central α-helix of H4 are important for nucleosome assembly. Lanes 1–5, protein levels in the unfractionated embryo; lanes 6–10, protein levels in chromatin. Lanes 1 and 6, no RNA; lanes 2 and 7, H4; lanes 3 and 8, residues 60–63 mutated to glycine; lanes 4 and 9, epitope-tagged amino acids 32–80; lanes 5 and 10, arginines 35 and 36 mutated to alanine. The bar indicates the position of tagged H4 protein.
Mutagenesis of Histone H3.
We wished to determine the role of the N terminus of histone H3 in chromatin assembly. Deletion of 63 amino acids, including the entire N-terminal tail together with the N-terminal α-helix between amino acids 45 and 55 (6–8), reduces but does not prevent the stable assembly of histone H3 into the nucleosome (Fig. 5, lanes 7–10; Table 1). This deletion includes all of the sites of histone acetylation (15). Thus, an acetylated N-terminal tail on histone H3 is not essential for incorporation into a nucleosome during chromatin assembly in metazoans.
Figure 5.
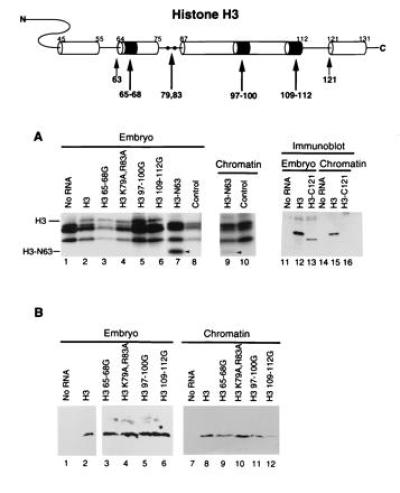
Domains of histone H3 essential and dispensable for nucleosome assembly. (A) Newly synthesized proteins in the embryo are indicated (lanes 1–8, 11–13), as in their incorporation into chromatin (lanes 9, 10, and 14–16). Detection is by radiolabeling (lanes 1–10) or by immunoblotting (lanes 11–16). The arrowhead indicates the N63 deletion. Note that endogenous histone synthesis is visualized in lanes 1, 8, and 10 in the absence of any expression of mutant H3 since no mRNA is injected (No RNA). (B) Indicated proteins synthesized in the embryo (lanes 1–6) are shown incorporated into chromatin (lanes 7–12). Detection is by immunoblotting.
Our next experiments examined the consequences of mutagenizing the histone fold domain in histone H3. Surprisingly, we find that the substitution of amino acids 65–68 (LPFQ) with glycine residues to break the integrity of the α-helix between amino acids 64 and 75 reduces but does not eliminate the assembly of H3 into chromatin (Fig. 5 A and B, lane 3, and Fig. 5B, lane 9; Table 1). Comparable mutagenesis of the long central α-helix at amino acids 97–100 (EAYL) and 109–112 (LCAI) reduces but does not eliminate assembly (Fig. 5 A and B, lanes 5 and 6, and Fig. 5B, lanes 11 and 12; Table 1). We suggest that there are redundant contacts necessary for dimerization in the “handshake” of the histone fold domains that mediate nucleosome assembly.
We examined the importance of two other sites in the H3 molecule. Replacement of arginine 83 and lysine 79 with alanine within a putative DNA binding site of H3 (6–8) is without effect on the incorporation of H3 into the nucleosome (Fig. 5A, lane 4, and Fig. 5B, lanes 4 and 10; Table 1). However, deletion of the putative C-terminal α-helix of H3 at amino acid 121 prevents the assembly of H3 into the nucleosome (Fig. 5A, lanes 11–16). This domain has been proposed to function as a dimerization surface for H3 (6–8), facilitating the assembly of the (H3/H4)2 tetramer.
DISCUSSION
In this work, we describe a useful general strategy for following the assembly of a mutant protein into a metazoan chromosome in vivo. The incorporation of a histone, or any other protein, into chromatin will depend on interactions with other structural proteins and DNA. Incorporation will also depend on interaction with molecular chaperones involved in assembly of nucleoprotein structures, nuclear import, and potential targeting to sites of DNA synthesis and chromatin assembly. Importantly, newly synthesized histones will have to compete with the large stores of histone sequestered in the Xenopus egg for assembly into chromatin (35, 36). Since any one of these variables could interfere with nucleosome assembly, our system imposes severe constraints on the assembly of histones into chromatin in vivo. However, there are no requirements for the histones to contribute to transcriptional regulation in the maintenance of the organism through early Xenopus development in our experiments (37). This is because transcription is not activated until the 4000-cell stage, a developmental transition that would occur after the stage at which nucleosomes are isolated in our work. Thus, constraints on chromatin assembly in the Xenopus embryo may differ from those in S. cerevisiae.
Our results lend considerable insight into earlier work on histones H3 and H4. Although acetylation of the N-terminal tail is conserved among metazoans (28) and correlates with the assembly of histone H4 into chromatin, it is not an essential feature of the assembly process. As can be inferred from work in yeast (16, 17), amino acids 1–28 with H4 are dispensable for nucleosome assembly. Because the H4 tail is highly conserved through eukaryotic evolution, our observations imply an essential role in other processes, such as gene regulation in metazoans, as has been clearly demonstrated in S. cerevisiae (20–24). Similar considerations apply to histone H3, where deletion of the N-terminal 63 amino acids does not interfere with the incorporation of the protein into nucleosomes. It is probable that the absence of a specific requirement for either the H3 or H4 N-terminal tail reflects the assembly of these proteins into nucleosomes as a heterodimer (17). Thus, it might be anticipated that mutations in the histone fold domains of H3 and H4 would have the most deleterious impact on chromatin assembly.
Our results provide insight into the requirements of the histone fold domain for incorporation into chromatin (6–8) and potentially into other nucleoprotein complexes (9–11). The essential and nonessential domains of the proteins are summarized in Fig. 6 for H4 (Fig. 6A) and for H3 (Fig. 6B). Remarkably, a region of histone H4 dispensable for nucleosome assembly is a portion of the histone fold domain: the C terminus from amino acids 80 to 102. This region of H4 is absolutely conserved from Tetrahymena to humans and contains only two substitutions from the S. cerevisiae sequence (1, 2). The integrity of this sequence is essential for viability in yeast (16), yet is not required for the chromatin assembly process in our experiments. Included in this sequence is the C-terminal α-helix of histone H4 from amino acids 84 to 94. This α-helix does not contain any DNA binding surface or interface with histone H3 (Fig. 6). Potentially, it might interact with histone H2B (6–8, 14). This latter interaction might be an important site for regulatory phenomena, since displacement of the H2A/H2B heterodimer from the nucleosome might accompany transcription (38, 39). The essential domains of histone H4 for nucleosome assembly appear to be the regions potentially required for interaction with histone H3. This is consistent with the critical role of the (H3/H4)2 tetramer in nucleating assembly of the nucleosome (5, 40). This conclusion is further strengthened by the deleterious effect on assembly of H3 into chromatin of deleting the C-terminal α-helix. This region is believed to be essential for interaction of the two H3 molecules in the assembly of the (H3/H4)2 tetramer (6–8).
Figure 6.
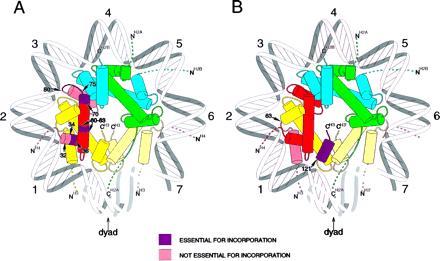
Summary of role of domains of H3 and H4 in nucleosome assembly. (A) Histone H4; (B) histone H3. Histone H4 is shown in red, H3 in yellow, H2B in blue, and H2A in green. Mutated or deleted regions are as indicated. The view shown is down the superhelical axis of the DNA in the nucleosome. For simplicity, the DNA is shown as a uniform superhelix. The helical turns are numbered relative to the dyad axis (0). Only one complete heterotypic tetramer of H2A, H2B, H3, and H4 is shown. The second H3 molecule is indicated in a pale yellow to indicate the site of dimerization across the dyad axis. N and C termini of the histones are indicated; the dashed lines indicate the N-terminal tails, the exact path of which is not known at this time.
Acknowledgments
We thank Dr. Dmitry Pruss for help with Fig. 6 and Ms. Thuy Vo for manuscript preparation.
References
- 1.De Lange R J, Farnbrough D M, Smith E L, Bonner J. J Biol Chem. 1969;244:5669–5679. [PubMed] [Google Scholar]
- 2.Wells D, Brown D. Nucleic Acids Res. 1991;19:2173–2188. doi: 10.1093/nar/19.suppl.2173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kornberg R. Science. 1974;184:868–871. doi: 10.1126/science.184.4139.868. [DOI] [PubMed] [Google Scholar]
- 4.Kornberg R, Thomas J O. Science. 1974;184:865–868. doi: 10.1126/science.184.4139.865. [DOI] [PubMed] [Google Scholar]
- 5.Camerini-Otero R D, Sollner-Webb B, Felsenfeld G. Cell. 1976;8:333–347. doi: 10.1016/0092-8674(76)90145-8. [DOI] [PubMed] [Google Scholar]
- 6.Arents G, Burlingame R W, Wang B C, Love W E, Moudrianakis E N. Proc Natl Acad Sci USA. 1991;88:10148–10152. doi: 10.1073/pnas.88.22.10148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Arents G, Moudrianakis E N. Proc Natl Acad Sci USA. 1993;90:10489–10493. doi: 10.1073/pnas.90.22.10489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Richmond T J, Rechsteiner T, Luger K. Cold Spring Harbor Symp Quant Biol. 1993;58:265–276. doi: 10.1101/sqb.1993.058.01.031. [DOI] [PubMed] [Google Scholar]
- 9.Xie X, Kokubo T, Cohen S L, Mirza U A, Hoffmann A, Chait B T, Roeder R G, Nakatani Y, Burley S K. Nature (London) 1996;380:316–322. doi: 10.1038/380316a0. [DOI] [PubMed] [Google Scholar]
- 10.Hoffmann A, Chiang C-M, Oelgeschlager T, Xie X, Burley S K, Nakatani Y, Roeder R G. Nature (London) 1996;380:356–359. doi: 10.1038/380356a0. [DOI] [PubMed] [Google Scholar]
- 11.Sinha S, Maity S N, Lu J, de Crombrugghe B. Proc Natl Acad Sci USA. 1995;92:1624–1628. doi: 10.1073/pnas.92.5.1624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Baxevanis A D, Arents G, Moudrianakis E N, Landsman D. Nucleic Acids Res. 1995;23:2685–2691. doi: 10.1093/nar/23.14.2685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ebralidse K K, Grachev S A, Mirzabekov A D. Nature (London) 1988;331:365–367. doi: 10.1038/331365a0. [DOI] [PubMed] [Google Scholar]
- 14.Pruss D, Hayes J J, Wolffe A P. BioEssays. 1995;17:161–170. doi: 10.1002/bies.950170211. [DOI] [PubMed] [Google Scholar]
- 15.Turner B M. Cell. 1993;75:5–8. [PubMed] [Google Scholar]
- 16.Kayne P S, Kim U J, Han M, Mullen J R, Yoshizaki F, Grunstein M. Cell. 1988;55:27–39. doi: 10.1016/0092-8674(88)90006-2. [DOI] [PubMed] [Google Scholar]
- 17.Ling X, Harkness T A A, Schultz M C, Fisher-Adams G, Grunstein M. Genes Dev. 1996;10:686–699. doi: 10.1101/gad.10.6.686. [DOI] [PubMed] [Google Scholar]
- 18.Megee P C, Morgan B A, Mittman B A, Smith M M. Science. 1990;247:841–845. doi: 10.1126/science.2106160. [DOI] [PubMed] [Google Scholar]
- 19.Park E C, Szostak J W. Mol Cell Biol. 1990;10:4932–4934. doi: 10.1128/mcb.10.9.4932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Durrin L K, Mann R K, Kayne P S, Grunstein M. Cell. 1991;65:1023–1031. doi: 10.1016/0092-8674(91)90554-c. [DOI] [PubMed] [Google Scholar]
- 21.Wan J, Mann R K, Grunstein M. Proc Natl Acad Sci USA. 1995;92:5664–5668. doi: 10.1073/pnas.92.12.5664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Johnson L M, Fisher-Adams G, Grunstein M. EMBO J. 1992;11:2201–2209. doi: 10.1002/j.1460-2075.1992.tb05279.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Johnson L M, Kayne P S, Kahn E S, Grunstein M. Proc Natl Acad Sci USA. 1990;87:6286–6290. doi: 10.1073/pnas.87.16.6286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hecht A, Laroche T, Strahl-Bolsinger S, Gasser S M, Grunstein S. Cell. 1995;80:583–592. doi: 10.1016/0092-8674(95)90512-x. [DOI] [PubMed] [Google Scholar]
- 25.Hebbes T R, Thorne A W, Crane-Robinson C. EMBO J. 1988;7:1395–1402. doi: 10.1002/j.1460-2075.1988.tb02956.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hebbes T R, Clayton A L, Thorne A W, Crane-Robinson C. EMBO J. 1994;13:1823–1830. doi: 10.1002/j.1460-2075.1994.tb06451.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Allis C D, Chicoine L G, Richman R, Schulman I G. Proc Natl Acad Sci USA. 1985;82:8048–8052. doi: 10.1073/pnas.82.23.8048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sobel R E, Cook R G, Perry C A, Annunziaton A T, Allis C D. Proc Natl Acad Sci USA. 1995;92:1237–1241. doi: 10.1073/pnas.92.4.1237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hayes J J, Clark D J, Wolffe A P. Proc Natl Acad Sci USA. 1991;88:6829–6833. doi: 10.1073/pnas.88.15.6829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bauer W R, Hayes J J, White J H, Wolffe A P. J Mol Biol. 1994;236:685–690. doi: 10.1006/jmbi.1994.1180. [DOI] [PubMed] [Google Scholar]
- 31.Smith S, Stillman B. Cell. 1989;58:15–25. doi: 10.1016/0092-8674(89)90398-x. [DOI] [PubMed] [Google Scholar]
- 32.Smith S, Stillman B. EMBO J. 1991;10:971–980. doi: 10.1002/j.1460-2075.1991.tb08031.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Woodland H R, Adamson E D. Dev Biol. 1977;57:118–132. doi: 10.1016/0012-1606(77)90359-1. [DOI] [PubMed] [Google Scholar]
- 34.Perry M, Thomsen G H, Roeder R G. J Mol Biol. 1985;185:479–499. doi: 10.1016/0022-2836(85)90065-8. [DOI] [PubMed] [Google Scholar]
- 35.Bouvet P, Dimitrov S, Wolffe A P. Genes Dev. 1994;8:1147–1159. doi: 10.1101/gad.8.10.1147. [DOI] [PubMed] [Google Scholar]
- 36.Dimitrov S, Dasso M, Wolffe A P. J Cell Biol. 1994;126:591–601. doi: 10.1083/jcb.126.3.591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Davidson E H. Gene Activity in Early Development. San Diego: Academic; 1986. [Google Scholar]
- 38.Hansen J C, Wolffe A P. Proc Natl Acad Sci USA. 1994;91:2339–2342. doi: 10.1073/pnas.91.6.2339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hayes J J, Wolffe A P. Proc Natl Acad Sci USA. 1994;89:1229–1332. [Google Scholar]
- 40.Eickbusch T H, Moudrianakis E N. Biochemistry. 1978;17:4955. doi: 10.1021/bi00616a016. [DOI] [PubMed] [Google Scholar]