

Abstract
We report on spectroscopic studies of the chiral structure in phospholipid tubules formed in mixtures of alcohol and water. Synthetic phospholipids containing diacetylenic moieties in the acyl chains self-assemble into hollow, cylindrical tubules in appropriate conditions. Circular dichroism provides a direct measure of chirality of the molecular structure. We find that the CD spectra of tubules formed in mixtures of alcohol and water depends strongly on the alcohol used and the lipid concentration. The relative spectral intensity of different circular dichroism bands correlates with the number of bilayers observed using microscopy. The results provide experimental evidence that tubule formation is based on chiral packing of the lipid molecules and that interbilayer interactions are important to the tubule structure.
Keywords: tubule, phospholipid, chirality, diacetylene
The self-assembly of biologically based lipids into unusual microstructures has been the subject of intense study in recent years, both for basic research and for potential applications in areas ranging from controlled release to electroactive composites (1, 2). Most long-chain phospholipids self-assemble into spherical bilayer aggregates, known as liposomes (3). However, certain synthetic phospholipids, with modified head groups or acyl chains, self-assemble into novel microstructures (4). One class of synthetic phospholipids, with photopolymerizable diacetylenic moieties in the acyl chains, was originally developed as an approach for increasing the durability of lipid bilayer aggregates (5–7). These lipids have been observed to self-assemble into hollow, cylindrical structures, known as tubules (8, 9). Similar cylindrical tubules have also been observed in other synthetic surfactants (10) and in bile (11). Such tubules appear to have potential for long-term release applications such as marine antifouling (12).
To explain the formation of tubules, several investigators have developed theories based on molecular chirality (11, 13–18). Although the details of these theories differ, they are all based on the principle that chiral interactions cause the molecules to pack at a nonzero angle with respect to their nearest neighbors. This chiral packing induces a twist in the bilayer, which results in the formation of a cylindrical structure. Electron microscopy of tubules decorated with metallic particles show helical markings that suggest chiral order in the bilayers (19, 20).
In an earlier study, we used circular dichroism to test the theoretical concept that the formation of tubules is driven by chiral molecular packing (20). CD, the difference in the absorption of right and left circularly polarized light, arises from the chirality of a molecular architecture. This chirality can arise from either the structure of individual molecules or from the chiral packing of molecules into larger aggregates (21, 22). Our experiments showed that diacetylenic lipid tubules have a very strong CD signal. By contrast, spherical liposomes of the same diacetylenic lipids have only a very weak CD signal, as do the lipid molecules dissolved in alcohol. These results provided experimental evidence that the packing of lipid molecules in tubules is chiral, while the packing of lipid molecules in spherical liposomes is not chiral.
In this paper, we present a detailed CD and UV absorption study of tubules formed in several different alcohol/water mixtures. This study supports the theoretical concept that tubule formation is driven by chiral packing in the lipid bilayer and it provides the first spectroscopic evidence for chiral interactions between bilayers. These interactions may be the basis for differences in wall thickness observed between tubules formed in different solvents. We find that there are two distinct peaks in the CD spectrum of tubules: a peak at 195 nm that can be associated with chiral packing of the diacetylene groups within a single bilayer and a peak at 202–205 nm that can be associated with chiral ordering of head groups between adjacent bilayers. By measuring the relative amplitudes of those two peaks, we investigate the chiral packing both within and between bilayers, and determine how it depends on the lipid concentration and the nature of the solvent. This work shows that spectroscopic techniques can provide new information about the chiral molecular architecture in lipid microstructures.
MATERIALS AND METHODS
Tubules were prepared in alcohol/water mixtures with a number of lipid concentrations and using the alcohols methanol, ethanol, 1-propanol, and isopropanol. The lipid 1,2 bis(tricosa-10, 12-diynoyl)-sn-glycero-3-phosphocholine (DC8,9PC) was purchased from Avanti Polar Lipids and recrystallized in acetone. The structure of this molecule is shown in Fig. 1a. The samples for UV absorption measurements were made by dissolving the lipid in spectroscopic grade solvents (Aldrich). For the CD studies, the tubules were prepared by dissolving DC8,9PC in the appropriate alcohol and mixing with water at 55°C. On slowly cooling the mixture through the lipid chain melting temperature (≈37°C), tubules are formed (22). The UV absorption measurements were performed using a Cary 2400 (Varian) dual-beam spectrometer operating at room temperature. The CD studies were performed on a Jasco J-720 spectropolarimeter operating between 175–700 nm. The samples were placed in water-jacketed quartz cells with path lengths of 0.1, 0.2, or 1.0 mm. Temperature control was provided by a water circulator that provided thermal stability of about 0.2°C. The spectrometer was calibrated with ammonium-d-camphorsulfonate (θ291 = 7910 deg cm2/dmol) and d-pantoyllactone (θ219 = −16,140 in water, θ223 = −12,420 in methanol) (24). Variations in the placement and orientation of the sample showed that the CD spectrum was independent of birefringent and scattering effects. Samples for electron microscopy were negative stained using uranyl acetate to enhance the contrast. Observations were made using a transmission electron microscope (Zeiss EM-010) operating at 60 kV.
Figure 1.
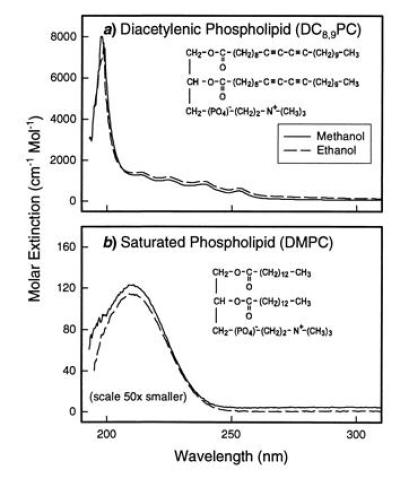
UV absorption spectra of DC8,9PC (a) and dimyrstoyl-phosphatidylcholine (b) dissolved in neat methanol and ethanol at a concentration of 5 mg/ml. The structure of these molecules is also shown.
RESULTS
The UV absorption spectra of the diacetylenic lipid and a similar saturated phospholipid are shown in Fig. 1. Fig. 1a shows the nearly identical spectra of DC8,9PC in methanol and ethanol that are composed of a large absorption at 198 nm and smaller peaks at 213, 225, 239, and 253 nm. Absorption spectra in 1-propanol and isopropanol are also similar. The molar extinction of the saturated lipid dimyrstoyl-phosphatidylcholine, with the same head group, but without the diacetylene group in the tails, is shown in Fig. 1b. This spectrum is characterized by a broad peak centered at 209 nm that arises from the ester chromophore in the glycerol backbone of the lipid head group.
The absorption spectrum of DC8,9PC is characteristic of a diacetylene group and the same as that observed in a diacetylenic fatty acid. The multiple absorption bands arise from mixing of the electronic states of the two conjugated acetylenic units. Theoretical studies on the electronic structure of a series of substituted conjugated diacetylenes showed the presence of an intense high energy band from the π-π* transition near 204 nm, the exact position depending on the substitution at the two ends of the acetylene group (25). The absorption from the head group seen in dimyrstoyl-phosphatidylcholine is also present in the DC8,9PC spectra, but obscured by the larger diacetylenic absorptions. Note that the scale in Fig. 1b is 50 times smaller than the scale in Fig. 1a. However, since CD is a differential measurement, weak absorption can give a large CD signal.
The CD spectra of DC8,9PC tubules prepared in different alcohol/water mixtures are compared in Fig. 2. Fig. 2a shows the molar ellipticity of tubules prepared with a 7:3 alcohol/water ratio and a lipid concentration of 1 mg/ml. The spectrum from methanol/water tubules is composed of three peaks, centered at 186, 195, and 205 nm, while that from ethanol/water tubules is dominated by a broad, asymmetric peak centered at 202 nm, with a smaller peak near 195 nm. Comparing the CD and absorption spectra, we associate the peak at 195 nm with absorption in the diacetylene group and the broader peak at 205 nm with the lipid head group. Experimental limitations due to solvent absorption prevent us from identifying the source of the CD peak at 186 nm. Fig. 2b shows the molar ellipticity of tubules prepared with a lipid concentration of 1 mg/ml in a 1:1 isopropanol/water solution and a 3:7 1-propanol/water solution. The spectrum of isopropanol/water tubules, with a broad peak at 201 nm, is nearly identical to that of ethanol/water tubules. The propanol/water tubules have a similar CD spectrum characterized by somewhat sharper peaks at 189 and 199 nm.
Figure 2.
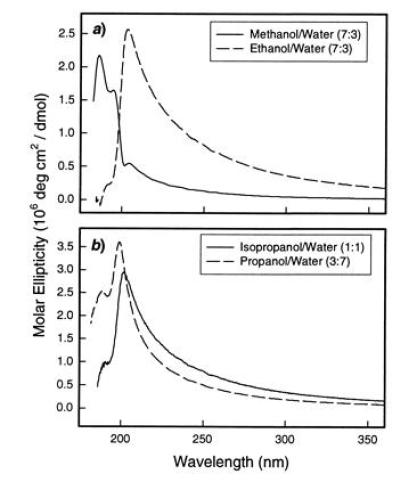
CD spectra of DC8,9PC tubules prepared in (a) methanol/water (7:3) and ethanol/water (7:3) and (b) isopropanol/water (1:1) and propanol/water (3:7) at a lipid concentration of 1 mg/ml.
In the ethanol/water system, we observe that the spectral intensity of the 202-nm band is much higher than that of the 195-nm band. Conversely, in the methanol/water tubules, the CD signal from the 195 nm peak is larger than for the 202-nm peak. This relative change in spectral intensities of the two bands indicates structural differences in the morphology of the tubules made in the two different solvents. Previous experiments found that for these lipid concentrations, the methanol/water tubules are composed of approximately 90% single bilayer walls, while ethanol/water tubules have an average of about seven bilayers (26). Because the CD peak at 202–205 nm is enhanced in multibilayer ethanol/water tubules, we infer that it is associated with chiral ordering between head groups in adjacent bilayers. Similarly, because the CD peak at 195 nm, associated with the diacetylene, is seen in single-bilayer methanol/water tubules, we presume that it is associated with chiral interactions between lipid molecules within a single bilayer.
The CD spectrum of DC8,9PC tubules in methanol/water (7:3) also shows an interesting concentration dependence. The ratio of the ellipticity at 205 nm to that at 195 nm changes with the lipid concentration as shown in Fig. 3. This figure shows a comparison of the CD spectra at 25°C of methanol/water tubules prepared with different lipid concentrations. The molar ellipticity of the 4 mg/ml DC8,9PC tubules is almost 50% greater than the 1 mg/ml sample at 205 nm, while the spectra are nearly identical below 200 nm. The ratio of the spectral intensity of the 205-nm to the 195-nm band as a function of the lipid concentration is shown in the inset of Fig. 3. Up to about 1 mg/ml the ratio is constant, while it begins to increase above this concentration. We attribute this to the tubules becoming more multilamellar as the concentration of lipid is increased. This was confirmed by electron microscopy as shown in Fig. 4. Such behavior has also been used to interpret heat capacity data on DC8,9PC tubules prepared in methanol/water (85:15), where the melting transition is observed to cross over from continuous to first-order near a concentration of 4 mg/ml (27). CD experiments on tubules made in 85:15 methanol/water indicate the 205- to 195-nm peak height ratio begins to increase at a higher lipid concentration than in 7:3 methanol/water. We expect that at higher lipid concentrations, the methanol/water tubule spectrum would begin to look like that of ethanol/water tubules. However, experimental limitations prevent us from studying concentrations higher than 5 mg/ml.
Figure 3.
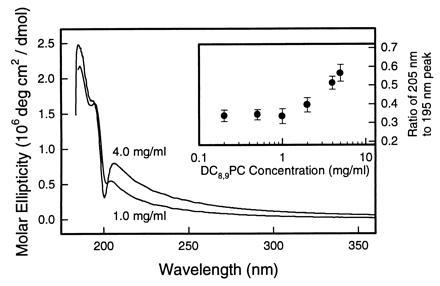
Concentration dependence of the CD spectra of DC8,9PC tubules in methanol/water (7:3) at 25°C. The ratio of the spectral intensities at 205 and 195 nm is shown in the Inset.
Figure 4.

Negative stained electron micrographs of DC8,9PC tubules in methanol/water (7:3) at lipid concentrations of 1 mg/ml (a and b) and 5 mg/ml (c and d). The 1 mg/ml sample (a and b) contains all single bilayer tubules, while the 5 mg/ml sample (c and d) is composed of approximately half single bilayer and half multiple bilayer tubules. The top-right corner of c shows several two bilayer tubules, as does d. Helical markings are seen on a tubule near the middle of c. (Bar = 1 μm.)
Fig. 4 compares negative-stained electron micrographs of methanol/water tubules at different lipid concentrations. Tubules prepared at a DC8,9PC concentration of 1 mg/ml are shown in Fig. 4 a and b, while 4 c and d show tubules at a lipid concentration of 5 mg/ml. The 1 mg/ml tubules all appear to be one bilayer thick, while approximately half of those prepared at 5 mg/ml contain two or more bilayers. The staining and drying process used in sample preparation results in nonuniform distribution of tubules on the sample grid. Fig. 4 a and c show concentrated regions of tubules. The difference in thickness is especially noticeable when the ends of the tubules are examined. Fig. 4b clearly shows the end of a single bilayer tubule, while Fig. 4d shows one that is two bilayers. These micrographs confirm that the number of bilayers in the methanol/water tubules increases as the lipid concentration is increased; this correlates with the relative increase in the 205-nm peak seen in the CD studies. In addition, helical markings are evident on a tubule near the middle of Fig. 4c.
DISCUSSION
As noted in the introduction, several investigators have developed theories of tubule formation based on molecular chirality (11, 13–18). The basic physical principle behind all of these theories is that chiral molecules do not tend to pack parallel to their neighbors, but rather at a slight angle with respect to their neighbors. For molecules constrained to a bilayer, this chiral packing induces a twist in the bilayer, which leads to the formation of a cylindrical structure. It may also induce a helical modulation in the direction of the molecular tilt on the cylinder, as well as slight helical ripples in the curvature. Although these theories predict the overall structure of tubules, they do not address certain important issues: they do not consider the effect of the solvent on tubule structure, and they do not consider the correlation between different bilayers of multilayer tubules.
Microscopy, particularly electron microscopy, has provided information that tends to support these theories (12, 19, 26). In contrast to the indirect evidence from microscopy, the CD studies reported in this paper provide direct evidence for a chiral molecular architecture in lipid tubules. The observation of several large CD peaks in tubules, which are not observed in spherical liposomes or in solutions of lipid molecules, provides clear evidence that the molecular packing in tubules is chiral. This chirality in molecular architecture occurs in both ethanol/water tubules, which show helical markings in electron micrographs, and single-bilayer methanol/water tubules, which do not. Thus, the CD results indicate a local, microscopic chirality in the molecular packing, in contrast with the more macroscopic helical structure visible in electron micrographs. The intralayer chiral order inferred from the CD results is consistent with the theoretical concept of chiral packing in the lipid membrane (11, 13–18).
In addition, the CD results provide the first clear spectroscopic evidence for differences between single-bilayer methanol/water tubules and multilayer tubules formed in other alcohols. Electron microscopy has shown that tubules prepared from methanol/water solutions are comprised mostly of single bilayer walls, while ethanol/water tubules have many bilayers (26). The relative intensity of the CD peak observed at about 205 nm compared with the one at 195 nm correlates with the existence of multiple bilayers. As discussed previously, the 205-nm peak appears to be associated with the head group of the lipids. In order for the intensity of a CD band to increase, there must be enhanced chiral ordering in the environment that affects the chromophore giving rise to the CD signal. Thus, the observed increase in CD at 205 nm suggests an increased chiral order between head groups of adjacent bilayers. This increased chiral order could result from a rotation in the molecular tilt direction from layer to layer, an increase in the magnitude of the tilt, or an increase in the biaxial order of the molecules. Since the distance between the bilayers can be as large as 1.5 nm (28), the interaction could be caused by hydrogen bonded clusters of water that are associated with the head groups on each of the layers. In this case, chemicals that either enhance or disrupt the structure of water could have a profound effect on the tubule structure. Further experiments are being carried out to investigate these ideas.
In conclusion, these CD studies provide new structural information about the molecular architecture of lipid tubules. The results support the theoretical concept of chiral molecular packing in tubules. The results also give new information about the effect of solvent on the chiral structure of tubules, which suggests an interlayer ordering in multilayer tubules. Experiments are now underway to determine if hydrogen bonding is important to the existence of the observed interlayer chiral ordering and if hydrogen bonding affects the number of bilayers.
Acknowledgments
We acknowledge valuable discussions with M. Markowitz, R. Price, B. Ratna, and R. Shashidhar. This research was supported by the Office of Naval Research. M. Spector is supported by a National Research Council–Naval Research Laboratory Research Associateship.
Footnotes
Abbreviation: DC8,9PC, 1,2 bis(tricosa-10, 12-diynoyl)-sn-glycero-3-phosphocholine.
References
- 1.Gaber B P, Schnur J M, Chapman D, editors. Biotechnological Applications of Lipid Microstructures. New York: Plenum; 1988. [Google Scholar]
- 2.Lasic D D. Liposomes: From Physics to Applications. Amsterdam: Elsevier; 1993. [Google Scholar]
- 3.Bangham A D, Hill M W, Miller N G A. Methods Membr Biol. 1973;1:1–68. [Google Scholar]
- 4.Schnur J M, Peckerar M, editors. Synthetic Microstructures in Biological Research. New York: Plenum; 1992. [Google Scholar]
- 5.Johnston D S, Sanghera S, Pons M, Chapman D. Biochim Biophys Acta. 1980;602:57–69. doi: 10.1016/0005-2736(80)90289-8. [DOI] [PubMed] [Google Scholar]
- 6.Hub H H, Hupfer B, Koch H, Ringsdorf H. Angew Chem Int Ed Engl. 1980;19:938–940. doi: 10.1002/anie.198009381. [DOI] [PubMed] [Google Scholar]
- 7.O’Brien D F, Whitesides T H, Klingbiel R T. J Polym Sci Polym Lett Ed. 1981;19:95–101. [Google Scholar]
- 8.Yager P, Schoen P E. Mol Cryst Liq Cryst. 1984;106:371–381. [Google Scholar]
- 9.Schnur J M. Science. 1993;262:1669–1676. doi: 10.1126/science.262.5140.1669. [DOI] [PubMed] [Google Scholar]
- 10.Nakashima N, Asakuma A, Kunitake T. J Am Chem Soc. 1985;107:509–510. [Google Scholar]
- 11.Chung D S, Benedek G B, Konikoff F M, Donovan J M. Proc Natl Acad Sci. 1993;90:11341–11345. doi: 10.1073/pnas.90.23.11341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schnur J M, Price R, Rudolph A S. J Controlled Release. 1994;28:3–13. [Google Scholar]
- 13.Helfrich W, Prost J. Phys Rev A. 1988;38:3065–3068. doi: 10.1103/physreva.38.3065. [DOI] [PubMed] [Google Scholar]
- 14.Zhong-can O-Y, Ji-xing L. Phys Rev A. 1991;43:6826–6836. [Google Scholar]
- 15.Yager P, Chappell J, Archibald D D. In: Biomembrane Structure and Function–The State of the Art. Gaber B P, Easwaran K R K, editors. Schenectady, NY: Adenine; 1992. [Google Scholar]
- 16.Nelson P, Powers T. J Phys II (Orsay, Fr) 1993;3:1535–1569. [Google Scholar]
- 17.Selinger J V, Schnur J M. Phys Rev Lett. 1993;71:4091–4094. doi: 10.1103/PhysRevLett.71.4091. [DOI] [PubMed] [Google Scholar]
- 18.Selinger J V, MacKintosh F C, Schnur J M. Phys Rev E. 1996;53:3804–3818. doi: 10.1103/physreve.53.3804. [DOI] [PubMed] [Google Scholar]
- 19.Burkett, S. L. & Mann, S. (1996) J. Chem. Soc. Chem. Commun. 321–322.
- 20.Schnur J M, Ratna B R, Selinger J V, Singh A, Jyothi G, Easwaran K R K. Science. 1994;264:945–947. doi: 10.1126/science.264.5161.945. [DOI] [PubMed] [Google Scholar]
- 21.Velluz L, Legrand M, Grosjean M. Optical Circular Dichroism. Principles, Measurements, and Applications. Weinheim, Germany: Verlag Chemie; 1965. [Google Scholar]
- 22.Woody R W. In: The Peptides. Hruby V J, editor. Vol. 7. New York: Academic; 1985. pp. 15–114. [Google Scholar]
- 23.Georger J H, Singh A, Price R R, Schnur J M, Yager P, Schoen P E. J Am Chem Soc. 1987;109:6169–6175. [Google Scholar]
- 24.Schippers P H, Dekkers H P J M. Anal Chem. 1981;53:778–782. [Google Scholar]
- 25.Jain M, Chandrasekhar J. J Phys Chem. 1993;97:4044–4049. [Google Scholar]
- 26.Ratna B R, Baral-Tosch S, Kahn B, Rudolph A S, Schnur J M. Chem Phys Lipids. 1992;63:47–53. doi: 10.1016/0009-3084(92)90021-g. [DOI] [PubMed] [Google Scholar]
- 27.Nounesis G, Ratna B R, Shin S, Flugel R S, Sprunt S N, Singh A, Litster J D, Shashidhar R, Kumar S. Phys Rev Lett. 1996;76:3650–3653. doi: 10.1103/PhysRevLett.76.3650. [DOI] [PubMed] [Google Scholar]
- 28.Caffrey M, Hogan J, Rudolph A S. Biochemistry. 1991;30:2134–2146. doi: 10.1021/bi00222a018. [DOI] [PubMed] [Google Scholar]