

Abstract
An endogenous circadian biological clock controls the temporal aspects of life in most organisms, including rhythmic control of genes involved in clock output pathways. In the fungus Neurospora crassa, one pathway known to be under control of the clock is asexual spore (conidia) development. To understand more fully the processes that are regulated by the N. crassa circadian clock, systematic screens were carried out for genes that oscillate at the transcriptional level. Time-of-day-specific cDNA libraries were generated and used in differential screens to identify six new clock-controlled genes (ccgs). Transcripts specific for each of the ccgs preferentially accumulate during the late night to early morning, although they vary with respect to steady-state mRNA levels and amplitude of the rhythm. Sequencing of the ends of the new ccg cDNAs revealed that ccg-12 is identical to N. crassa cmt encoding copper metallothionein, providing the suggestion that not all clock-regulated genes in N. crassa are specifically involved in the development of conidia. This was supported by finding that half of the new ccgs, including cmt(ccg-12), are not transcriptionally induced by developmental or light signals. These data suggest a major role for the clock in the regulation of biological processes distinct from development.
Keywords: development, ccg, frq, light
Circadian rhythms, generated and controlled by an endogenous biological clock, are observed in a wide variety of organisms ranging from cyanobacteria to man (reviewed in ref. 1). These daily rhythms, which persist under constant environmental conditions, are observed at the biochemical, physiological, and behavioral levels. The essence of circadian rhythms involves the restriction of an activity to a particular time of day to coordinate biological processes with exogenous environmental cycles. This requires the regulation of genes involved in the output pathways by the circadian clock.
Lower eukaryotes provide powerful model organisms with which to study the molecular and biochemical bases of circadian rhythms. Included in this group are the fungi which harbor relatively simple circadian systems, where the clock can be studied in the absence of complex multicellular interactions. Owing to its long history of genetics and biochemistry, the circadian system of Neurospora crassa has been one of the most highly described (reviewed in refs. 2 and 3): genetic and molecular analyses have shown that frq encodes a central component of the N. crassa oscillator required for rhythmicity (reviewed in ref. 2). In N. crassa the clock controls the timing of asexual spore (conidia) development, where in a wild-type strain grown in constant darkness, conidiation is initiated once every 22 hr.
One useful approach for understanding clock control of cellular events involves the characterization of genes that are temporally regulated by the circadian clock. Systematic screens for clock-regulated genes have now been reported in N. crassa (4), Synechococcus (5), and Drosophila (6). In N. crassa, initial screens for clock-regulated genes were carried out by using subtractive hybridization of morning versus evening RNAs (4). Two morning-specific clock-controlled genes, ccg-1 [independently identified as a glucose-repressible gene (grg-1) (7) in N. crassa (4, 8)] and ccg-2 [later shown to be allelic to eas (9, 10)] were identified, and transcripts were shown to cycle with periods reflecting the genotypes of the strains analyzed. Nuclear run-on experiments demonstrated that rhythmic expression of ccg-1 and eas(ccg-2) results from clock control of transcription (11), implicating the involvement of cis-acting regulatory elements mediating temporal control. This was confirmed for the eas(ccg-2) gene through the localization of a critical positive-activating clock element that was shown to be both necessary and sufficient for clock-regulated expression (12). Additionally, the rhythm of conidiation is unaffected in both ccg-1 and eas(ccg-2) null strains, demonstrating that both genes are true output genes (as opposed to clock genes) that do not feedback on the oscillator (8, 9).
The commitment of N. crassa to development, a commitment regulated in part by the circadian clock, signals a substantial restructuring of the organism accompanied by major changes in the repertoire of expressed genes (13–16). In fact, all of the circadian clock-controlled genes heretofore described in N. crassa are induced during development and by light, and are believed to be involved with the conidial developmental pathway. These include eas(ccg-2), which encodes a fungal hydrophobin, a component of the hydrophobic rodlet layer of conidia important for spore dissemination (9, 10, 17), and ccg-1, which is highly regulated during development and by light (8, 18). External factors such as carbon source limitation, desiccation, and light, known to induce development (13), also induce al-3 (19) and the conidiation-associated genes con-6 and con-10, all three of which seem to be circadianly regulated (14, 16, 19). Where data are available for all five of these clock-regulated genes, regulation by light, development, or by the clock appears to be independently conferred by separate elements acting at the level of each gene (10, 12, 14, 16–19) rather than through global coordinated regulation of a single factor by all three processes. This unanticipated one-to-one correspondence between circadian clock regulation and light/developmental regulation has prompted the notion that the sole output of the N. crassa clock might be associated with the developmental pathway leading to conidiation.
While the isolation of only two ccgs in the initial screen (4) implied that few genes are regulated by the clock in N. crassa, the subsequent discovery that these genes were highly expressed suggested that clock regulation might be more prevalent. In addition, since only two phases of the circadian cycle were examined, and clock control may be more extensive at other times of the day, many potential rhythmic genes were likely missed (4). For these reasons we initiated a new search for clock-controlled genes to gain a more complete understanding of the cellular processes that are temporally regulated and to see if processes other than development might be clock regulated.
In this study, time-of-day-specific cDNA libraries were generated and used in differential screens to identify six new ccgs in N. crassa. All of the ccgs display maximal transcript accumulation in the late night to early morning, and differ in overall levels, amplitude and absolute peak of accumulation. As compared with ccg-1 and eas(ccg-2) which together represent ≈10% of poly(A)+ species, the relative mRNA expression levels of the new ccgs are low. In addition, the ccgs are found not to be tightly linked, ruling out coordinate regulation of gene expression at specific chromosomal locations by the clock. Preliminary sequencing of the ends of the ccgs revealed that ccg-12 is identical to the previously cloned copper metallothionein gene (cmt) of N. crassa, not known to be involved in development. Importantly, on further examination we found that half of the new ccgs, including cmt (ccg-12), show no regulation in response to changes in light or factors that trigger development. This establishes that the N. crassa circadian clock may have substantial involvement in aspects of the life of the organism exclusive of the regulation of development.
MATERIALS AND METHODS
Strains and Growth Conditions.
Strains of N. crassa used in this study include the frq+ strain 30-7 (bd; A), and the long period mutant 695-425 (bd; frq7; A); the band (bd) mutation enhances the circadian rhythm of conidiation (20) but does not affect the clock itself. Strains for restriction fragment length polymorphism (RFLP) mapping studies were obtained from the Fungal Genetics Stock Center (FGSC) (Kansas City). Neurospora media (Vogel’s or Fries) and vegetative growth conditions were as described (21, 22). Bacterial strain XLI-Blue (Stratagene) was used for all phage and plasmid manipulations as described (23).
Culture Harvesting Conditions.
For rhythmic RNA analyses, tissue was grown and synchronized by light to dark transfers as described (4). Light and developmental induction experiments were accomplished in strain 30-7 using published methods (12). Light treatments were carried out on cultures grown in the dark for 22 hr (CT12), a time of minimal expression for each of the ccgs.
cDNA Library Construction.
Poly(A)+ RNA (2 μg) isolated from N. crassa (24) was annealed to 0.6 μg of oligo(dT)-XbaI primer-adapter (Promega) by heating the reaction to 70°C for 3 min, and then cooling immediately on ice. First- and second-strand cDNA synthesis was carried out using the cDNA Synthesis System of BRL. Double-strand cDNA (ca. 250 ng) was ligated to 10 pM phosphorylated EcoRI adapters (Promega), phenol extracted, and then digested with 10 units XbaI. The cDNA was size selected using a Sephacryl S-400 spin column (Promega) and ligated to λ Zap II (Stratagene). The phage were then packaged in vitro using Promega’s Packagene extracts and plated on Escherichia coli XL1-Blue. Growth of the recombinant phage on media containing isopropyl β-d-thiogalactoside and 5-bromo-4-chloro-3-indolyl β-d-galactoside indicated that less than 1 × 104 plaque-forming units/μg of DNA were devoid of inserts.
Differential Screens and Plasmid Rescue.
Differential screens of the time-of-day-specific cDNA libraries were accomplished by plating a total of 5 × 104 phage on XL1-Blue in a top agar overlay, and duplicate plaque lifts (23) were hybridized to 5 × 105 cpm of 32P-labeled cDNA (specific activity, 5 × 107 cpm/μg) from the same and opposite time points. Following autoradiography, the signals from the duplicate lifts were compared by visual inspection.
In vivo excision of the pBluescript plasmid containing each ccg cDNA from the λ ZapII vector was accomplished by recircularization of the plasmid containing the cloned insert with M13 helper phage R408 as described in the Undigested Lambda ZapII Cloning Kit (Stratagene).
Nucleic Acid Isolation and Hybridization.
RNA was isolated using either a large scale method (25) or a miniprep method (26) and Northern blots were processed as described (12). RNA loading was normalized to rRNA as indicated, which remains at constant levels under the growth conditions used (4). Densitometric data was acquired and analyzed as described (12).
RFLP Mapping and Sequencing.
The multicent strains, comprising 38 individual progeny from a cross of Mauriceville and Oak-Ridge-derived strains were used in RFLP analysis. N. crassa genomic DNA was isolated from mycelia by the hexadecyltrimethylammonium bromide (CTAB) extraction method (27, 28). Restriction polymorphisms were detected by Southern blot analyses (23) of digested DNAs using the individual cDNAs as probes, with the exception of ccg-8. No polymorphisms were detected for the ccg-8 gene using its corresponding cDNA probe, so a cosmid containing the ccg-8 gene (X10:3G) was identified from an ordered cosmid library (Orbach and Sachs library, FGSC) and then used as a probe. The results were compared with published updates of RFLP maps in the Fungal Genetics Newsletter (29).
Dideoxy sequencing of the cDNA clones was initiated from the T7 and T3 sites in the in vivo excised pBluescript vector, using an Applied Biosystems Prism Dye-deoxy sequencing kit.
RESULTS
Generation of Time-of-Day-Specific cDNA Libraries.
To identify genes under control of the circadian biological clock in N. crassa, time-of-day-specific cDNA libraries were constructed for use in differential screens. To enrich for the selection of the desired clock-controlled genes instead of nutritionally and developmentally regulated genes (that could fluctuate in the liquid culture conditions used to synchronize the clock), a clock mutant strain, frq7 was used. Unlike the 22-hr period in a frq+ strain, the frq7 strain has a period of 29 hr, and this mutation [a single base pair change in the FRQ protein (30)] is specific to the circadian clock (31, 32). In liquid cultures of frq+ versus frq7 strains, the time of harvest can be chosen such that the same time in real hours in constant darkness (DD) represents different circadian times (CTs) in the two strains. (CT represents the normalization of biological time to 24 circadian hours per cycle in strains or organisms with varying periods. By convention, CT0 represents dawn and CT12 represents dusk in a 12:12 light/dark cycle.) Thus, clock-regulated transcripts, responding to CT, will have cycled out of phase (as shown for the ccg-1 gene at DD44 in Fig. 1A), whereas developmentally and nutritionally regulated transcripts, responding to sidereal time in culture, should remain at equivalent levels.
Figure 1.
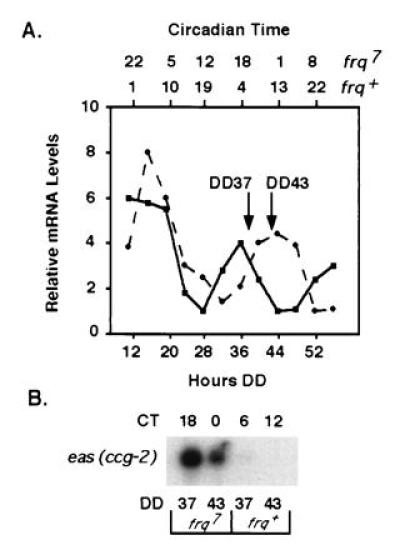
Isolation of RNA from frq+ and frq7 cultures representing four different times in the circadian day. (A) Densitometry of the clock-regulated ccg-1 mRNA isolated from N. crassa frq+ (solid line) and frq7 (dotted line) cultures grown in continuous dark and harvested every 4 hr (4). The time in the dark (Hours DD) is indicated on the bottom of the plot, and the corresponding CT is shown on the top for both strains. Arrows point to the time of harvest (DD37 and DD43) used to isolate mRNA for the generation of the time-of-day-specific cDNA libraries. (B) Northern blot hybridization of total RNA (10 μg) extracted from frq+ and frq7 strains at DD37 and DD43 representing approximate CT18, CT0, CT6, and CT12 as shown in A. Hybridization is to the clock-controlled eas(ccg-2) gene.
Poly(A)+ RNA was isolated from frq+ and frq7 cultures held for 37 hr [representing approximately CT6 (noon) in frq+ and approximately CT18 (midnight) in frq7] and 43 hr [representing approximately CT12 (dusk) in frq+ and approximately CT0 (dawn) in frq7] in constant darkness (Fig. 1A). To verify that the cultures were indeed rhythmic, RNA from each time point was hybridized to eas(ccg-2) and as expected, the message peaks in accumulation between CT18 and CT0 (Fig. 1B). Because the levels of eas(ccg-2) mRNA are not globally elevated in the frq7 strain (data not shown), the peaks at CT18 and CT0 are clearly due to clock control. The rhythmic mRNA from each of the four times was used to construct directional cDNA libraries in λ ZapII (see Materials and Methods). The total number of recombinant phage particles in each of the libraries exceeds 2.5 × 106, indicating that each gene has a greater than 99% chance of being represented in the library based on rough estimates of 2000 vegetative mRNA species in N. crassa (33).
As an internal control for the time-of-day-specific libraries, we analyzed the representation of both ccg-1 and eas(ccg-2) by hybridizing a minimum of 1000 plaques from each of the libraries with probes specific for both genes (Table 1). Compatible with our rhythmic Northern blot analyses (this study and ref. 4), the ccg-1 cDNA is most prevalent in the CT0 library, and the eas(ccg-2) cDNA is most prevalent in the CT18 and CT0 libraries. Additionally, we found ccg-1 and eas(ccg-2) to be abundantly represented in the libraries, comprising 8.5% of the CT0 cDNAs and 2.8% of the CT18 cDNAs, respectively.
Table 1.
Representation of ccg-1 and eas(ccg-2) in the time-of-day-specific libraries
| Gene | CT0, % | CT6, % | CT12, % | CT18, % |
|---|---|---|---|---|
| ccg-1 | 8.5 | 0.9 | 0 | 0.3 |
| eas(ccg-2) | 2.1 | 0 | 0.3 | 2.8 |
Percentages are derived from the number of positive hybridization signals/total number of plaques probed.
Identification of Clock-Controlled Genes by Differential Screens of the Time-of-Day-Specific Libraries.
To identify clock-regulated genes, differential screens were performed with the cDNA libraries representing opposite phases of the circadian day. The screens were designed to identify messages specific to early morning (CT0 versus CT12), midday (CT6 versus CT18), early evening (CT12 versus CT0), and midnight (CT18 versus CT6).
Approximately 5 × 104 phage from each library were hybridized to labeled cDNA from the same and opposite time points. Twenty eight plaques (out of ≈40,000 visualized signals) found to hybridize to cDNA of the same phase, but not the opposite phase, were chosen as candidate ccgs. Sixteen of these were shown by hybridization to be re-isolates of ccg-1 and eas(ccg-2) (providing a convincing internal control) while the other 12 clones (4-CT0, 1-CT6, 2-CT12, 5-CT18) represented candidate ccgs. When the candidate ccgs were hybridized to RNA from four different times of day (data not shown), hybridization signals were not detected for two clones, two did not display any apparent clock regulation, and one clone (ccg-6) was represented three times (as determined by preliminary sequencing of the ends of the remaining candidate cDNAs). In all, six new candidate ccgs emerged from this screen.
To confirm the circadian nature of the regulation of these genes, the six candidate ccgs were examined further by hybridization to RNA from both frq+ and frq7 cultures. Tissue was grown in constant darkness and harvested for RNA isolation every 4 hr over 2 consecutive days. A representative experiment is shown in Fig. 2 for the ccg-4 gene. In the frq+ strain (Fig. 2A), the mRNA cycles and accumulates to peak levels at DD12 (CT1) in the first cycle and DD32–DD36 (CT0-4) in the second cycle with a wild-type period of about 22 hr. A similar rhythm is observed in the frq7 strain where the period is ≈29 hr (Fig. 2B). As predicted for a clock-regulated gene, the message cycles in both frq+ and frq7, with a period that is consistent with the genotype of the strain. This is particularly evident at DD36, where the mRNA levels are 180° out of phase in the two strains (Fig. 2C). Similar analyses were conducted on the other candidate genes, and the results of hybridizations to RNA from the frq7 strain are shown in Fig. 3. In these experiments, the same RNA (with the exception of ccg-4) is probed with each of the ccgs, and eas(ccg-2) is used as an internal control. Although each gene hybridizes to an mRNA displaying an oscillation with a period of ≈29 hr, distinct differences in individual patterns of regulation are evident.
Figure 2.
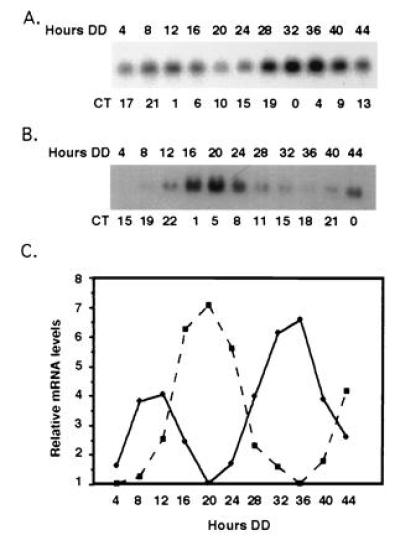
Criteria used to establish clock control of gene expression, as illustrated by the ccg-4 gene. The steady-state levels of ccg-4 mRNA were assayed by Northern blot analyses in frq+ (A) and frq7 (B) strains. Liquid cultures of mycelia were grown in constant darkness and harvested after the indicated times in the dark (Hours DD). The approximate CT at the time of harvests are shown below the autoradiograms. (C) Following autoradiography, ccg-4 mRNA was quantitated by densitometry, and plotted as relative band intensity versus time in the dark for both frq+ (solid line) and frq7 (dotted line). Equal loading of the RNA was verified by inspection of ethidium bromide stained rRNA on the gel (data not shown)
Figure 3.
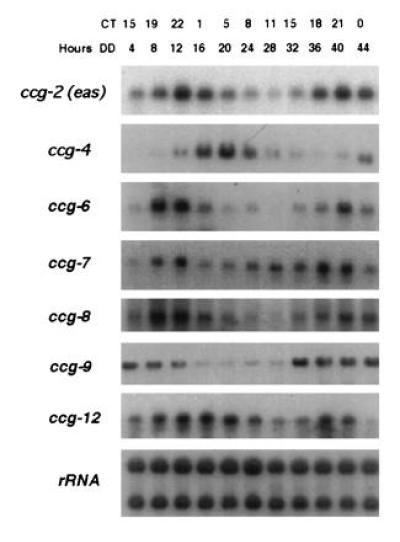
Rhythmic transcript accumulation of the ccgs in the frq7 (29-hr period) strain. RNA was isolated from the frq7 mutant strain at the indicated times (Hours DD) representing the approximate CTs shown at the top of the autoradiograms, and hybridized to RNA probes specific for each ccg (shown at left). Equivalent loading of RNA was verified by hybridization to rRNA. Exposure times of the autoradiograms are varied for visualization.
Transcript size, relative abundance, average peak of mRNA accumulation, and amplitude of the rhythm for each gene are summarized in Table 2. All of the genes show peak transcript accumulation around the late night (CT18-23) to early morning (CT0-5), ranging from CT18 (for ccg-12) to CT5 (for ccg-4); none peak in the late day (CT6-11) or early evening (CT12-17).
Table 2.
Summary of clock-controlled genes
| Measured transcript size,* kb | Relative transcript abundance† | Average peak‡ | Amplitude§ | |
|---|---|---|---|---|
| ccg-1 | 0.6 | +++ | CT3 | ×4 |
| eas(ccg-2) | 0.7 | +++ | CT22 | ×5 |
| ccg-4 | 1.1 | ++ | CT5 | ×11 |
| ccg-6 | 0.9 | + | CT19 | ×9 |
| ccg-7 | 1.0 | + | CT21 | ×4.5 |
| ccg-8 | 2.2 | − | CT20 | ×3.5 |
| ccg-9 | 2.0 | − | CT19 | ×4.5 |
| ccg-12 | 0.5 | + | CT18 | ×2 |
Transcript sizes were determined by comparison to known molecular weight markers.
Transcript abundance was roughly estimated from exposure times required to obtain a signal equivalent to the eas(ccg-2) transcript.
The average peak of transcript abundance was determined from densitometry of the RNA levels over the times analyzed from Fig. 3. The peak in message accumulation will vary slightly in different experiments.
The amplitude of the rhythm was calculated from the data in Fig. 3 using densitometry, and is reported as half the maximal distance between the inflection levels.
ccg-7, -8, and -12 Are Likely to be Involved in Clock-Output Pathways Distinct from Conidiation.
To begin to characterize the ccgs, the ends of the cDNA clones were sequenced. Comparisons of partial cDNA sequence for the ccg-12 gene (data not shown) in the data bases reveals that ccg-12 is identical to the previously cloned cmt gene of N. crassa (34) encoding copper metallothionein (CuMT). CuMT (26 aa) is multifunctional, having a role in copper storage, detoxification, and metal transfer to copper containing proteins. RNA abundance levels of the cmt gene are regulated not only by the clock (these studies), but expression is also induced by copper ions, with maximal accumulation occurring 1 hr after the addition of copper ions to the growth media (34). Based on all previous studies of N. crassa clock-regulated genes, we anticipated that the new ccgs would be directly involved in conidiation; however, the finding that ccg-12 encodes CuMT suggested otherwise, as no role for CuMT in conidiation has been reported. This prompted us to examine the transcripts from the ccgs for induction after conidiation is initiated (Fig. 4A).
Figure 4.
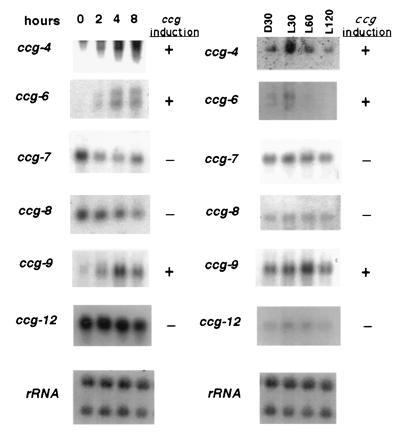
Developmental and light regulation of the ccgs. (A) RNA isolated from tissue 0, 2, 4, and 8 hr after induction of conidiation was hybridized to each new ccg (shown on the left) and to rRNA to verify equivalent loading. (B) RNA was isolated from cultures grown for 22 hr in the dark (CT12) and harvested after an additional 30 min in the dark (D30), or transferred to the light for 30, 60, and 120 min (L30, L60, and L120, respectively). The RNA was hybridized to probes as indicated above.
The expression patterns of ccg-4, -6, and -9 during conidial development are very similar to conidiation-specific genes (35), with each transcript accumulating to high levels about 4 hr after development is initiated by desiccation in constant light. In contrast, the abundance of the cmt(ccg-12) transcript remains unchanged, while both ccg-7 and ccg-8 show slightly higher levels of mRNA in undifferentiated vegetative hyphae (mycelia) (Fig. 4A; time 0).
Genes that are regulated during conidiation in N. crassa are in most, if not all, cases induced by light. Therefore, we also assayed the effects of light on transcript levels for the ccgs (Fig. 4B). Consistent with dual light and developmental control, the ccg-4 and ccg-6 mRNAs accumulate to higher levels after a light treatment at CT12 for 30 min (L30) as compared with the dark control (D30), and ccg-9 mRNA accumulates to maximal levels after a 1-hr light treatment (L60). However, transcripts from ccg-7, -8, or -12 do not display any light responsiveness at the times examined. In N. crassa, a light treatment at CT12 causes about a 2-hr phase delay (36); however, this delay cannot explain the observed light induction of the ccgs for two reasons. First, the response is rapid and not sustained over the short length of time examined. Second, the response to light varies for ccgs that peak at approximately the same time of day. In both experiments, the ccg-4 transcript is found to be present at very low levels as compared with previous experiments (Figs. 2 and 3), suggesting that ccg-4 expression may be sensitive to the differences in growth media or the developmental age in the different experimental protocols (see Materials and Methods). In addition, while only one transcript is observed for the ccg-6 gene in the rhythmic Northern blot (Fig. 3), two transcripts are observed here, and are shown to be similarly regulated during conidiation and after light treatment. These patterns are consistent with the complex regulation seen previously in the light, developmentally, and clock-regulated genes.
The Clock-Regulated Genes Are Not Chromosomally Clustered.
The genomic locations of the six new ccgs were determined by RFLP mapping. DNA from a set of 38 progeny from a single reference cross (N. crassa Oak-Ridge-derived strain FGSC 4411 × Mauriceville FGSC 2225) containing numerous polymorphisms (22) were digested with restriction enzymes previously found to generate distinctive band patterns in DNA isolated from the two parental strains (data not shown). Following hybridization of specific ccg probes to the restriction enzyme-digested DNA from the 38 progeny, the restriction patterns were compared with a table of known segregation markers (29), and the relative genetic locations of each ccg were determined. These map locations are shown in Fig. 5. Coordinate gene regulation by the clock based on chromosomal position is not indicated since the eight confirmed ccgs map to five of N. crassa’s seven chromosomes, with none of the genes being closely linked.
Figure 5.
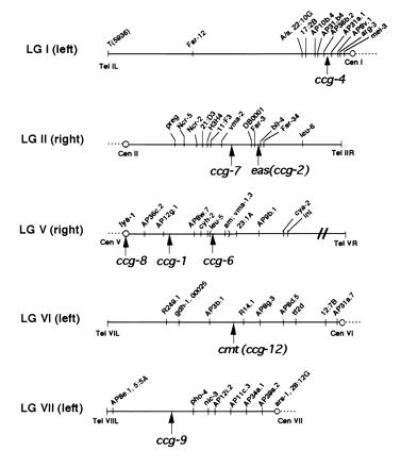
Clock-controlled genes map throughout the genome. Chromosomal map locations of N. crassa clock-controlled genes, as determined by RFLP analysis, are indicated by arrows on the appropriate linkage groups (this work and ref. 4). Individual linkage groups (LG) are designated with a Roman numeral, followed by left or right in parenthesis, indicating the specific arm of the chromosome represented. The map positions of other conventional markers are shown for reference, and the order of markers denoted in parenthesis are uncertain (37). The centromeres are shown as a circle. The map distances are rough approximations.
DISCUSSION
Six new circadian clock-controlled genes have been isolated using differential screens of time-of-day-specific cDNA libraries. Interestingly, all peak in the late night to early morning, although they vary in overall expression levels and amplitude of the rhythm (Table 2). In addition, we find that the ccgs map throughout the genome, suggesting that clock control of gene expression is not accomplished by clustering genes at specific chromosomal locations. Preliminary sequencing of the cDNAs reveals that ccg-12 is identical to N. crassa cmt, encoding CuMT. The lack of evidence for an involvement for cmt in conidiation prompted us to investigate the regulation of the new ccgs during asexual spore development. Three of the ccgs (ccg-4, -6, and -9) are shown to be induced during conidial development and by light; however, cmt(ccg-12) as well as ccg-7 and -8 are not. Thus, three of the six new ccgs identified here display similar yet distinctly novel regulation as compared with all of the other known ccgs.
While nothing in the design of our screen for ccgs precluded the selection of afternoon- or evening-specific genes, attempts to identify them were unsuccessful. This is interesting in view of the fact that conidiation is initiated in the late night to early morning, and our selection of late night- to morning-specific genes initially appeared consistent with the possibility that the sole purpose of the clock in N. crassa is to control the timing of conidiation. However, our finding that ccg-12 encodes CuMT, a protein not known to have a role in development, and the demonstration that not all of the ccgs are induced during conidiation or by light, points to control of other, as of yet undescribed, output pathways by the circadian clock. Although our screen did not approach saturation [as evidenced by the repeated isolation of only one of the new genes and the lack of selection of the inabundant, rhythmically expressed frq gene (38)], evening-specific genes appear to be either absent, in low abundance, or expressed at levels too low to be detected in this screen.
Interestingly, the time of maximal accumulation of the N. crassa clock-regulated transcripts coincides to some degree with peak expression of the clock gene frq (CT0-6) (38). Similar relationships were observed in Drosophila, where a majority of oscillating transcripts showed elevated levels after lights off (6), around the same time as the peak in cyclic clock proteins PER and TIM (39, 40). The overlapping phases might suggest the possibility of a direct involvement of the clock gene products in controlling rhythmic activity of the output genes, although transcriptional regulation by neither FRQ in N. crassa nor PER and TIM in Drosophila has yet been demonstrated directly. These findings might also suggest a common factor controlling rhythmicity of clock genes and clock-controlled genes. However, the phase of the rhythm (i.e., the time of peak expression) differs in the ccgs, so it is likely either that the transcripts have different half-lives, or are controlled by distinct regulatory factors or phasing factors that interact with a common element to manifest the rhythm. Together, these results suggest that diverse regulatory pathways exist in N. crassa to control rhythmic output from the clock.
The fact that the circadian clock can control the timing of conidiation implies that many clock-regulated genes in N. crassa will have a role in conidial development. This is clearly the case for eas(ccg-2) (9, 10), and is inferred for ccg-1 (8). Notably, we found that ccg-1 and eas(ccg-2) mRNA’s together represent about 10% of the total morning-specific poly(A)+ population. This level of expression was particularly of interest for ccg-1 as no detectable growth phenotype is observed in a ccg-1 null mutant strain (8). In addition, three other genes in N. crassa have in recent years been shown to be regulated during conidial development, by light, and likely by the circadian clock. These include al-3, encoding geranylgeranyl pyrophosphate synthase, essential for the biosynthesis of carotenoids (19), and two genes of unknown function, con-6 and con-10, both of which were isolated as a result of their increased expression during conidiation (14). Based on these findings, it is expected that three of the clock-regulated genes (ccg-4, -6, and -9) isolated here and found to be both developmentally regulated and photoinducible, will be involved in conidiation.
The extent of clock-control of gene expression in N. crassa appears to be limited to less than 10% of the genome based on this screen, previous screens (4), and by routine examination of genes in the laboratory (unpublished data). This is similar to what has been reported in Drosophila, where roughly 8% of 261 head-specific cDNA clones examined hybridized to an oscillating mRNA with a greater than 2-fold amplitude in a 12 hr light/dark cycle (6). Together, these findings are in dramatic contrast to the apparent global control of gene expression by the clock observed in the cyanobacterium Synechococcus (5), suggesting that in eukaryotes comprehensive regulation of gene expression by the oscillator is not a general phenomenon.
Studies of circadian-regulated gene expression, both at the transcriptional and translational levels, in diverse organisms are beginning to identify important metabolic pathways regulated by the circadian clock. In many organisms, control of gene expression by the clock includes both day- and night-specific regulation (reviewed in refs. 41–44). However, of these genes, only the clock-regulated genes of N. crassa and Drosophila have been identified in an organism where a component of the circadian oscillator has been well characterized, and thus, can ultimately be placed in a genetic pathway from the clock to the rhythmic output genes.
N. crassa, an organism that has been well defined both biochemically and genetically, now provides a rich source of eukaryotic genes known to be under control of the circadian clock. The systematic isolation and characterization of clock-controlled genes provides the framework for future studies aimed at detailing the pathways and molecular components involved in the conveyance of temporal information from the clock to rhythmic cellular processes. Together, these analyses will provide an entrée to understanding the mechanisms by which circadian regulation of gene expression is accomplished. In the long term, this will enable comparisons among the components necessary to achieve clock-mediated rhythmic gene expression in N. crassa and other eukaryotes.
Acknowledgments
We wish to thank Nam-Hai Chua for suggesting the use of frq7. We also thank N. Garceau, S. Brody, and D. Ebbole for comments on the manuscript, and both past and present members of our laboratories for useful discussions. This work was supported by grants from the National Institute of Mental Health (MH01186 to J.C.D and MH44651 to J.C.D and J.J.L.), the Air Force Office of Scientific Research (F49620-94-1-0260 to J.J.L.), the National Science Foundation (MCB-9307299 to J.J.L.), the Norris Cotton Cancer Center core grant at Dartmouth Medical School, and by a National Institutes of Health postdoctoral fellowship (GM15185 to D.B-P.).
Footnotes
Abbreviations: CT, circadian time; ccg, clock-controlled gene; RFLP, restriction fragment length polymorphism; FGSC, Fungal Genetics Stock Center; CuMT, copper metallothionein.
References
- 1.Edmunds L N. Cellular and Molecular Bases of Biological Clocks. New York: Springer; 1988. [Google Scholar]
- 2.Loros J J. Semin Neurosci. 1995;7:3–13. [Google Scholar]
- 3.Lakin-Thomas P L, Coté G G, Brody S. Crit Rev Microbiol. 1990;17:365–416. doi: 10.3109/10408419009114762. [DOI] [PubMed] [Google Scholar]
- 4.Loros J J, Denome S A, Dunlap J C. Science. 1989;243:385–388. doi: 10.1126/science.2563175. [DOI] [PubMed] [Google Scholar]
- 5.Liu Y, Tsinoremas N F, Johnson C H, Levedeva N V, Golden S S, Ishiura M, Kondo T. Genes Dev. 1995;9:1469–1478. doi: 10.1101/gad.9.12.1469. [DOI] [PubMed] [Google Scholar]
- 6.Van Gelder R N, Bae H, Palazzolo M J, Krasnow M A. Curr Biol. 1995;5:1424–1436. doi: 10.1016/s0960-9822(95)00280-6. [DOI] [PubMed] [Google Scholar]
- 7.McNally M T, Free S J. Curr Genet. 1988;14:545–551. doi: 10.1007/BF00434079. [DOI] [PubMed] [Google Scholar]
- 8.Lindgren K M. Ph.D. thesis. Hanover, NH: Dartmouth Medical School; 1994. [Google Scholar]
- 9.Bell-Pedersen D, Dunlap J C, Loros J J. Genes Dev. 1992;6:2382–2394. doi: 10.1101/gad.6.12a.2382. [DOI] [PubMed] [Google Scholar]
- 10.Lauter F-R, Russo V E, Yanofsky C. Genes Dev. 1992;6:2373–2381. doi: 10.1101/gad.6.12a.2373. [DOI] [PubMed] [Google Scholar]
- 11.Loros J J, Dunlap J C. Mol Cell Biol. 1991;11:558–563. doi: 10.1128/mcb.11.1.558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bell-Pedersen D, Dunlap J C, Loros J J. Mol Cell Biol. 1996;16:513–521. doi: 10.1128/mcb.16.2.513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Springer M L. BioEssays. 1993;15:365–374. doi: 10.1002/bies.950150602. [DOI] [PubMed] [Google Scholar]
- 14.Lauter F-R, Yanofsky C. Proc Natl Acad Sci USA. 1993;90:8249–8253. doi: 10.1073/pnas.90.17.8249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Schmidhauser T J, Lauter F-R, Schumacher M, Zhou W, Russo V E A, Yanofsky C. J Biol Chem. 1994;269:12060–12066. [PubMed] [Google Scholar]
- 16.Corrochano L M, Lauter F-R, Ebbole D J, Yanofsky C. Dev Biol. 1995;176:190–200. doi: 10.1006/dbio.1995.1016. [DOI] [PubMed] [Google Scholar]
- 17.Arpaia G, Loros J J, Dunlap J C, Morelli G, Macino G. Plant Physiol. 1993;102:1299–1305. doi: 10.1104/pp.102.4.1299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Arpaia G, Loros J J, Dunlap J C, Morelli G, Macino G. Mol Gen Genet. 1995;247:157–163. doi: 10.1007/BF00705645. [DOI] [PubMed] [Google Scholar]
- 19.Arpaia G, Carattoli A, Macino G. Dev Biol. 1995;170:626–635. doi: 10.1006/dbio.1995.1242. [DOI] [PubMed] [Google Scholar]
- 20.Sargent M L, Briggs W R, Woodward D O. Plant Physiol. 1966;41:1343–1349. doi: 10.1104/pp.41.8.1343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Davis R L, de Serres F J. Methods Enzymol. 1970;17:79–143. [Google Scholar]
- 22.Metzenberg R L, Stevens J N, Selker E U, Morzycka-Wroblewska E. Fungal Genet Newsl. 1984;31:35–39. [Google Scholar]
- 23.Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 24.Chirgwin J M, Przybyla A E, MacDonald R J, Rutter W J. Biochemistry. 1979;18:5294–5299. doi: 10.1021/bi00591a005. [DOI] [PubMed] [Google Scholar]
- 25.Reinert W R, Patel V B, Giles N H. Mol Cell Biol. 1981;1:829–835. doi: 10.1128/mcb.1.9.829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yarden O, Plamann M, Ebbole D J, Yanofsky C. EMBO J. 1992;11:2159–2166. doi: 10.1002/j.1460-2075.1992.tb05275.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zolan M E, Pukkila P J. Mol Cell Biol. 1986;6:195–200. doi: 10.1128/mcb.6.1.195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Taylor J W, Natvig D O. In: Zoosporic Fungi in Teaching and Research. Fuller M S, Jaworsky A, editors. Athens, GA: Southeastern; 1987. pp. 252–258. [Google Scholar]
- 29.Metzenberg R L, Grotelueschen J. Fungal Genet Newsl. 1993;40:130–138. [Google Scholar]
- 30.Aronson B D, Johnson K A, Dunlap J C. Proc Natl Acad Sci USA. 1994;91:7683–7687. doi: 10.1073/pnas.91.16.7683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Feldman J F. Annu Rev Plant Physiol. 1982;33:583–608. [Google Scholar]
- 32.Feldman J F, Dunlap J C. Photochem Photobiol Rev. 1983;7:319–368. [Google Scholar]
- 33.Wong L-J C, Marzluf G A. Biochim Biophys Acta. 1980;607:122–135. doi: 10.1016/0005-2787(80)90226-9. [DOI] [PubMed] [Google Scholar]
- 34.Munger K, Germann U A, Lerch K. Experientia Suppl (Basel) 1987;52:393–399. doi: 10.1007/978-3-0348-6784-9_37. [DOI] [PubMed] [Google Scholar]
- 35.Berlin V, Yanofsky C. Mol Cell Biol. 1985;5:849–855. doi: 10.1128/mcb.5.4.849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Dharmananda S. Ph.D thesis. Santa Cruz: Univ. of California; 1980. [Google Scholar]
- 37.Perkins D D. Fungal Genet Newsl. 1992;39:61–70. [Google Scholar]
- 38.Aronson B D, Johnson K A, Loros J J, Dunlap J C. Science. 1994;263:1578–1584. doi: 10.1126/science.8128244. [DOI] [PubMed] [Google Scholar]
- 39.Hardin P, Hall J, Rosbash M. In: Molecular Genetics of Biological Rhythms. Young M W, editor. New York: Dekker; 1993. pp. 155–169. [Google Scholar]
- 40.Sehgal A. Semin Neurosci. 1995;7:27–35. [Google Scholar]
- 41.Kay S A, Millar A J. In: Molecular Genetics of Biological Rhythms. Young M W, editor. New York: Dekker; 1993. pp. 73–89. [Google Scholar]
- 42.Cahill G M, Besharse J C. Prog Retinal Res. 1995;14:267–291. [Google Scholar]
- 43.Sassone-Corsi P. Cell. 1994;78:361–364. doi: 10.1016/0092-8674(94)90415-4. [DOI] [PubMed] [Google Scholar]
- 44.Mittag M, Lee D-H, Hastings J W. Proc Natl Acad Sci USA. 1994;91:5257–5261. doi: 10.1073/pnas.91.12.5257. [DOI] [PMC free article] [PubMed] [Google Scholar]