

Abstract
The comparison of malaria indicators among populations that have different genetic backgrounds and are uniformly exposed to the same parasite strains is one approach to the study of human heterogeneities in the response to the infection. We report the results of comparative surveys on three sympatric West African ethnic groups, Fulani, Mossi, and Rimaibé, living in the same conditions of hyperendemic transmission in a Sudan savanna area northeast of Ouagadougou, Burkina Faso. The Mossi and Rimaibé are Sudanese negroid populations with a long tradition of sedentary farming, while the Fulani are nomadic pastoralists, partly settled and characterized by non-negroid features of possible caucasoid origin. Parasitological, clinical, and immunological investigations showed consistent interethnic differences in Plasmodium falciparum infection rates, malaria morbidity, and prevalence and levels of antibodies to various P. falciparum antigens. The data point to a remarkably similar response to malaria in the Mossi and Rimaibé, while the Fulani are clearly less parasitized, less affected by the disease, and more responsive to all antigens tested. No difference in the use of malaria protective measures was demonstrated that could account for these findings, and sociocultural or environmental factors do not seem to be involved. Known genetic factors of resistance to malaria did not show higher frequencies in the Fulani. The differences in the immune response were not explained by the entomological observations, which indicated substantially uniform exposure to infective bites. The available data support the existence of unknown genetic factors, possibly related to humoral immune responses, determining interethnic differences in the susceptibility to malaria.
Keywords: Burkina Faso, Fulani, Mossi, Rimaibé
The ability to resist Plasmodium falciparum malaria is an important adaptive trait of human populations living in endemic areas. The detection of significant differences in the expression of this trait and the identification of the factors involved should improve the understanding of the host–parasite relationship and might lead to advances in control strategies (1). The importance of this research area is emphasized by the recent finding that particular HLA class I and class II alleles are associated with resistance to severe malaria (2) and by the complexities faced in the design of an effective vaccine against P. falciparum (3).
The study of human heterogeneities in the response to malaria can be approached at the population level by comparing either groups with similar genetic backgrounds exposed to different disease selective pressures (4)—e.g., the same population distributed over endemic and non-endemic zones—or groups with different genetic backgrounds exposed to similar parasite inoculation rates—e.g., different ethnic groups living together in an endemic zone. The interethnic comparison has been only rarely pursued (5–8), presumably deterred by the high number of possible confounding factors due to sociocultural differences. Following a preliminary survey carried out in 1993 (9), we considered this research approach feasible in the case of three sympatric West African ethnic groups, Fulani, Mossi, and Rimaibé, exposed to intense P. falciparum transmission in rural savanna areas near Ouagadougou, Burkina Faso.
The Fulani genetic/cultural background is well-differentiated from that of the Mossi and Rimaibé, since the former are nomadic pastoralists with non-negroid features of possible caucasoid origin (10), while the latter are Sudanese negroid populations with a long tradition of sedentary farming in sub-Saharan savannas (11). Closer to the Mossi in terms of ethnic origin, the Rimaibé have adopted most of the sociocultural habits of the Fulani, having been their slaves. The increasing tendency of the Fulani to settle and converge with the habits of sedentary populations determines sufficiently stable sympatric and even syntopic conditions that facilitate the interethnic comparison.
The results of our comparative study show consistent interethnic differences in P. falciparum infection rates, malaria morbidity, and prevalence and levels of antibodies to various P. falciparum antigens. The existence in the Fulani of an immunogenetically determined higher resistance to the parasite appears to be the most plausible interpretation of these findings.
MATERIALS AND METHODS
Study Area and Subjects.
Malaria surveys were carried out in two rural villages near the town of Ziniaré (35 km northeast of Ouagadougou) in a shrubby savanna of the Mossi plateau (≈300 m above sea level) belonging to the Sudan–Sahelian ecoclimatic zone (isohyets, 600–900 mm). Very intense P. falciparum transmission is recorded during the June–October rainy season, frequently reaching mean sporozoite inoculation rates well above one infective bite per person per night (12). The main malaria vectors are Anopheles gambiae (represented by the chromosomal forms Savanna and Mopti), Anopheles arabiensis, and Anopheles funestus (13). The great majority of the population belongs to the Mossi ethnic group, but the Fulani are widespread and usually more closely associated to the Rimaibé. The settled Fulani have converged with the habits of sedentary populations, living in similar houses. Their environment is still characterized by the presence of variable numbers of cows.
Oral informed consent for multiple immunoparasitological, clinical, and entomological surveys during 1994 and 1995 was obtained from a Fulani/Rimaibé community (≈600 inhabitants) living in the village of Barkoundouba and from a Mossi/Rimaibé community (≈1300 inhabitants) living ≈5 km apart in the village of Barkoumbilen (Fig. 1). Both communities have been settled in the area for >30 years. Information was collected for each ethnic group on movements out of the village, use of mosquito coils, bednets, or other mosquito protective methods, access to health care, and self-medication. Additional samples of the three ethnic groups, used for the serological tests, were from the village of Watinoma (≈30 km from the Barkoundouba area), which was the study site of the preliminary survey carried out in October 1993 (9).
Figure 1.
Sketch map of the study area showing the villages of Barkoumbilen and Barkoundouba. Capital letters indicate the position of the compounds of the ethnic groups. M, Mossi; R, Rimaibé; F, Fulani.
Entomological Observations.
Estimates of the entomological inoculation rate (which equals the number of potentially infective bites per person per night) were based on mosquito samples from indoor pyrethrum spray catches performed in 12 compounds for each of the three ethnic groups. Collections were made in the morning of 3 subsequent days in a total of 36 compounds (one room per compound and four rooms per day for each ethnic group). The number of compounds examined was further increased from 36 to 60 in the high transmission season of 1995 (July–October). The malaria vectors collected (A. gambiae, A. arabiensis, and A. funestus) were processed with immunoenzymatic methods (14) to estimate the P. falciparum circumsporozoite protein (CSP) positivity index and the human blood index. The entomological inoculation rates were calculated from the product of the human blood index and the CSP positivity index, multiplied by the number of blood-fed mosquitoes in the rooms, divided by the number of persons sleeping in the same rooms. We adopted this procedure in view of the marked endophilic behavior of the malaria vectors in the study area (M.C., unpublished work). Moreover, this allowed to refer to samples of human blood-fed vectors directly representative of the biting population on each ethnic group (actually, on the same individuals recruited for the immunoparasitological study) avoiding the pitfall of possible interethnic differences in attractiveness. Entomological surveys were performed in August and September 1994 and in March, July, and September 1995, ≈1 week before the corresponding immunoparasitological survey. Additional observations were carried out at the end of October and November 1994 and in August and October 1995, for a more effective comparison of the sporozoite inoculation rates during the main transmission seasons.
Blood Examination.
Thick and thin blood smears were prepared following the standard procedures, and 100 microscopic fields (≈20 leukocytes per field at ×1000 = ≈0.25 μl of blood) of the thick blood smear were examined. The Plasmodium species was identified on the thin blood smear.
Serological Tests.
Blood samples were collected in K3-EDTA sterile tubes. Within 3–4 hr after bleeding, the plasma was transferred and kept at −20°C until serological tests were done. We measured the humoral response against three P. falciparum antigens: CSP, thrombospondin-related anonymous protein (TRAP), and major surface antigen (MSA)-1. The CSP antibody response was evaluated with a Sclavo (Siera, Italy) ELISA Kit based on (NANP)40 antigen (14). The cut-off value for seropositivity was the mean plus three standard deviations of the values obtained on 30 negative Italian sera. CSP antibody levels were expressed as the log10 of arbitrary immunoenzymatic units, using a reference curve obtained by serial dilutions of a pool of human sera highly reactive to P. falciparum CSP or as the log10 of the absorbances. Anti-TRAP and anti-MSA-1 antibody responses were analyzed by ELISA based on recombinant segments corresponding to the 26–504 sequence of TRAP and to the MAD20 34–469 primary structure of MSA-1, respectively. Briefly, Polysorp immunoplates (Nunc) were coated with TRAP or MSA-1 at a concentration of 5 μg/ml. To prevent nonspecific binding, the wells were then incubated 2 hr at 37°C with 1% albumin in NaHCO3 (pH 9.2). Sera were diluted 1:200 and tested in duplicate. Sera were allowed to react for 1 hr at room temperature. For the detection of bound antibodies, goat anti-human Ig serum conjugated to alkaline phosphatase (Promega) was added at a dilution of 1:5000. Bound phosphatase activity was revealed by adding a solution of 1 mM 4-nitrophenylphosphate in 10% diethanolamine/10 mM MgCl2. After developing for 60 min, the optical density was measured at 405 nm (OD405) with a Titertek (Flow Laboratories, McLean, VA) spectrophotometer. The cut-off values for positive reactions were calculated as the mean OD405 plus three standard deviations of the Italian sera. TRAP and MSA-1 antibody levels were expressed as the log10 of the absorbances.
Active Case Detection.
The incidence of clinical episodes of malaria was compared in 496 children (0–10 years) of the three ethnic groups living in the Barkoundouba/Barkoumbilen area. Morbidity was monitored during 2 months of the 1995 high-transmission season (from August 21 to October 21) by means of active case detection, based on daily visits to the children at home. If the axillary temperature was >38.0°C, thick and thin blood films were prepared and an antimalaria treatment was given. The incidence of the probable malaria episodes was evaluated by the analysis of the fever cases associated with various thresholds of P. falciparum parasite density (see Table 2).
Table 2.
Clinical episodes of malaria
| Ethnic groups and comparisons | No. of Pf-positive fever cases | Distribution of Pf-positive fever
cases in different classes of Pf PD, %
|
Geometric mean of PD | Average of clinical episodes with different
Pf PD thresholds
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1–100 | 101–400 | 401–1600 | 1601–6400 | 6401–12,800 | 12,801–25,600 | >25,600 | >0 | >6400 | >12,800 | >25,600 | |||
| M | 95 | 7.4 | 11.6 | 15.8 | 22.1 | 9.5 | 16.8 | 16.8 | 3304 | 0.579 | 0.250 | 0.195 | 0.098 |
| RB | 92 | 7.6 | 10.9 | 15.2 | 17.4 | 16.3 | 20.0 | 13.0 | 3069 | 0.497 | 0.243 | 0.162 | 0.065 |
| RD | 21 | 0 | 14.3 | 14.3 | 28.6 | 9.5 | 14.3 | 19.0 | 4345 | 0.618 | 0.265 | 0.206 | 0.118 |
| NF | 208 | 6.7 | 11.5 | 15.4 | 20.7 | 12.5 | 17.8 | 15.4 | 3289 | 0.543 | 0.248 | 0.180 | 0.084 |
| F | 51 | 21.6 | 11.8 | 13.7 | 25.5 | 15.7 | 9.8 | 1.96 | 955 | 0.495 | 0.136 | 0.058 | 0.010 |
| M vs. RB | 0.83 | 0.94 | 0.93 | 0.53 | 0.24 | 0.77 | 0.60 | 0.85 | 0.32 | 0.76 | 0.25 | 0.25 | |
| M vs. RD | 0.35* | 0.72* | 1.0* | 0.57 | 1.0* | 1.0* | 0.76* | 0.68 | 0.72 | 0.80 | 0.73 | 0.57 | |
| RB vs. RD | 0.34* | 0.71* | 1.0* | 0.24 | 0.73 | 0.76* | 0.49* | 0.64 | 0.33 | 0.66 | 0.28 | 0.17 | |
| M vs. F | 0.026 | 0.91 | 0.93 | 0.80 | 0.24 | 0.36 | 0.016 | 0.004 | 0.38 | 0.09 | 0.005 | 0.009 | |
| RB vs. F | 0.032 | 0.81 | 0.96 | 0.35 | 0.89 | 0.20 | 0.061* | 0.004 | 0.97 | 0.15 | 0.052 | 0.06 | |
| RD vs. F | 0.027* | 1.0* | 1.0* | 0.98 | 0.71 | 0.68* | 0.023 | 0.021 | 0.36 | 0.16 | 0.011 | 0.004 | |
| NF vs. F | 0.003* | 0.85 | 0.94 | 0.58 | 0.71 | 0.24 | 0.019 | 0.001 | 0.56 | 0.08 | 0.009 | 0.016 | |
Clinical episodes of malaria were defined by temperature higher than 38.0°C and by different P. falciparum parasite densities recorded during 2 months of active case detection in children (0-10 years) of three ethnic groups. The size of the samples and their mean ages (in years) ± SD were as follows: 164 Mossi from Barkoumbilen, age = 5.0 ± 2.7; 185 Rimaibé from Barkoumbilen, age = 4.9 ± 2.8; 34 Rimaibé from Barkoundouba, age = 4.7 ± 3.0; 383 non-Fulani, age = 4.9 ± 2.8; 103 Fulani from Barkoundouba, age = 4.9 ± 2.7. The lower section of the table lists the various comparisons and the respective P values calculated by Yates-corrected χ2 test (or Fisher’s exact probability test when needed) in case of frequencies, and by Kruskal–Wallis test for means. Pf, P. falciparum; PD, parasite densities; M, Mossi; RB, Rimaibé from Barkoumbilen; RD, Rimaibé from Barkoundouba; NF, non-Fulani; F, Fulani. Ranges indicate number of parasites per μl of blood (PD).
Fisher exact test.
Statistical Methods.
Yates-corrected χ2 test (or Fisher’s exact probability test when needed) was used for the analysis of parasite rates, anti-CSP seroprevalences, and entomological indexes; Kruskal–Wallis test was used for the analysis of parasite and mosquito densities, antibody levels, and incidence of clinical malaria.
RESULTS
Population Movements and Antimalaria Measures.
All subjects included in the study can be considered permanently resident in the study area, since the reported movements were mostly daily and short range. No interethnic differences were recorded in the use of mosquito protective methods, access to health care, and self-medication. The use of bednets is exceptional in the study area, and the reported antimalaria self-medication was ≈10% during the 2 weeks before the high-transmission season surveys.
Inoculation Rates.
The results of the entomological surveys carried out during the 1994 and 1995 main transmission periods (August–November) and in the intervening dry season (March 1995), are shown in Fig. 2. The total number of malaria vectors processed is 14,334, of which 657 A. funestus and 13,677 A. gambiae sensu lato. The latter included both A. arabiensis and A. gambiae, mostly represented by the chromosomal form Mopti. The inoculation rates were found similar for the three ethnic groups. Higher mean vector densities were, in some cases, recorded in the Fulani rooms, but these differences were not reflected in the corresponding entomological inoculation rate values because they were generally counterbalanced by higher number of sleepers per room. CSP positivity index values ranged from 3.1% to 22.0% during the transmission season, while in March no CSP-positive specimen was found in a total sample of 214 mosquitoes. The intensity of transmission decreased in 1995, presumably influenced by the lower rainfall (and consequent vector densities) and by the impact of our parasitological surveys in terms of increased antimalaria treatments (and consequent lower frequency of gametocyte carriers). The mean human blood index was 93% (1626/1748), without significant differences between the samples from the compounds of the three ethnic groups.
Figure 2.
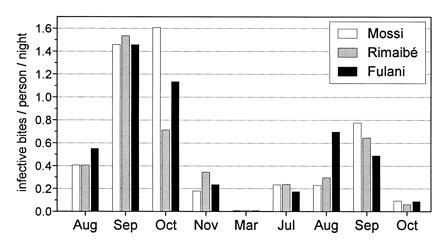
Entomological inoculation rates in the compounds of the three ethnic groups (Mossi, Rimaibé, and Fulani) in the Barkoundouba area from August 1994 to October 1995.
Parasite Rates and Density.
The parasitological data from the five cross-sectional surveys are summarized in Fig. 3. The lower P. falciparum prevalence in the Fulani was almost always highly significant in all age groups (Fig. 3A). The differences were particularly marked in the older age groups and in the period of lower transmission. Interethnic differences were also recorded in the positive parasite densities (Fig. 3B), showing lower values in the Fulani children <10 years of age. No intraethnic difference between sexes was found. The fifth survey showed, as expected, a general decrease of the parasite rates due to the chloroquine treatments of the fever cases detected by the daily follow-up of the children. The analysis of the negative-to-positive conversion rates in the age groups >10 years old further emphasized the difference between the Fulani, one-third of whom remained negative in the five surveys, and the Mossi and Rimaibé, who converted all but one to P. falciparum positivity (Fig. 4). The prevalences of P. malariae and P. ovale were generally much lower (data not shown), with few significant fluctuations of the parasite rates and densities and no evidence of coherent interethnic differences.
Figure 3.
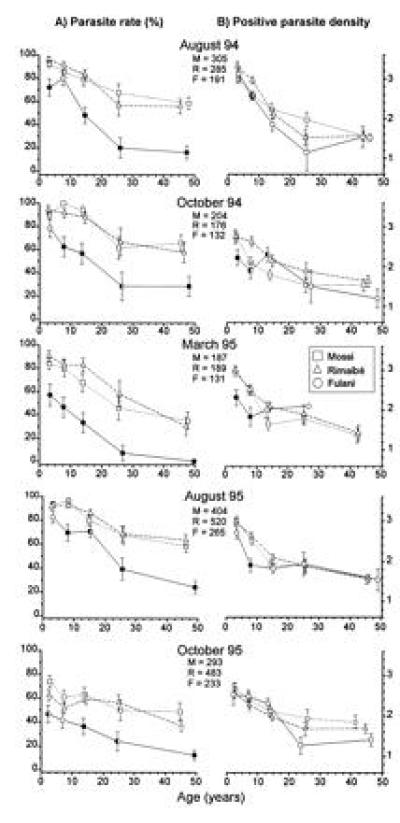
Values ± standard errors, by age and ethnic group, of P. falciparum parasite rate (A) and log10 of positive parasite density (B) in the Barkoundouba area, Burkina Faso. The positions of the symbols on the x-axis correspond to the mean age for each of the following age groups: 0–5, 6–10, 11–20, 21–30, and >30 years. Data refer to five cross-sectional surveys (August 1994, October 1994, March 1995, August 1995, and October 1995). To indicate a significant difference between the Fulani and one or both the other two groups, we used a half-filled or filled symbols, respectively. The sample sizes of each survey are shown under the respective month. M, Mossi; R, Rimaibé; and F, Fulani.
Figure 4.
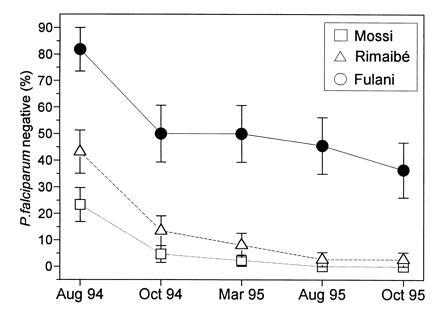
Comparative analysis in three sympatric ethnic groups of the percentage (± SE) of negative individuals at the first examination (August 1994) and rate of persistence of their negativity in the successive cross-sectional surveys. Only individuals more than 10 years old and examined in all five surveys of Fig. 1 are considered. No significant difference of mean age was observed among the three subsamples. The comparison does not include the 0–10 year age groups, since their negativity rates were too low for this type of analysis. All the Fulani/Mossi and Fulani/Rimaibé comparisons are statistically significant with P < 0.01.
Humoral Immune Response.
Anti-CSP antibody prevalences and levels in the first three surveys are summarized in Fig. 5. In all age groups, the Fulani had higher antibody levels than the other two ethnic groups. Antibody prevalences were also higher in the Fulani children, while the older age groups were 100% responsive in all samples tested. The results of additional serological tests are summarized in Table 1, where the Fulani are compared with syntopic Mossi and Rimaibé in Watinoma and Barkoundouba. The Fulani’s stronger humoral immune response is not limited to the CSP antigen but extends to TRAP and MSA-1 antigens. Moreover, preliminary data (not shown) clearly indicate a higher response of the Fulani also against the amino- and carboxy-terminal regions of the CSP antigen, respectively corresponding to the 22–125 and 289–390 amino acid sequence of the P. falciparum CSP, the Pf-155 ring-infected erythrocyte surface antigen (EENV)6, the Pf-332 (SVTEEIAEEDK)2, and Schistosoma mansoni schistosomula (D.M., unpublished data).
Figure 5.
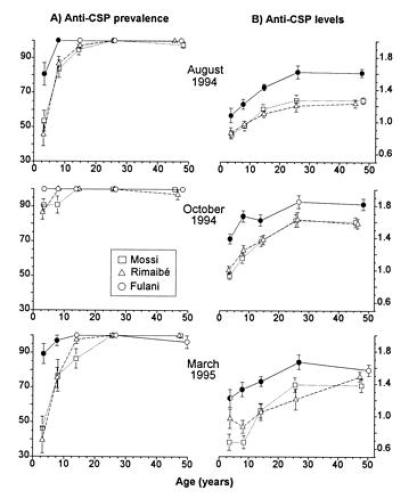
Anti-P. falciparum CSP antibody prevalences (A) and levels (B) by age and ethnic group. The data refer to the same samples examined in the first three surveys illustrated in Fig. 3 and the same symbols are used. The prevalences are shown as percent positive ± SE, and the levels are shown as log10 immunoenzymatic units ± SE.
Table 1.
Levels of anti-CSP, TRAP, and MSA-1 antibodies in syntopic ethnic groups of the villages of Watinoma and Barkoundouba
| Ethnic groups and comparisons | Log10
absorbances
|
|||||
|---|---|---|---|---|---|---|
| Watinoma (October
1993)
|
Barkoundouba (August 1995)
|
|||||
| CSP | TRAP | MSA-1 | CSP | TRAP | MSA-1 | |
| F | 2.900 | 2.658 | 3.083 | 2.964 | 2.531 | 3.105 |
| M | 2.645 | 2.596 | 2.730 | — | — | — |
| R | 2.627 | 2.488 | 2.546 | 2.628 | 2.376 | 2.848 |
| F vs. M | 0.002 | 0.470 | 0.002 | — | — | — |
| F vs. R | 0.000 | 0.079 | 0.000 | 0.000 | 0.002 | 0.000 |
| M vs. R | 0.770 | 0.260 | 0.160 | — | — | — |
Within each village, the samples analyzed had comparable ages. The Watinoma sample consisted of individuals more than 10 years old: 188 of them were tested for CSP (99 Mossi, 65 Rimaibé, and 24 Fulani), and 77 were tested for TRAP and MSA-1 (27 Mossi, 26 Rimaibé, and 24 Fulani). The Barkoundouba sample consisted of 90 individuals (61 Fulani and 29 Rimaibé) aged more than 30 years. P values in the bottom section of the table are from Kruskal–Wallis test. F, Fulani; M, Mossi; and R, Rimaibé.
Clinical Episodes of Malaria.
A total of 259 episodes of axillary temperature >38°C with P. falciparum parasitemia were recorded and are analyzed in Table 2. The geometric means of P. falciparum parasite densities were 955 parasites per μl in the 51 Fulani episodes and 3289 parasites per μl in the 208 Mossi and Rimaibé (Kruskal–Wallis test, P = 0.001). When defined by parasite density >12,800 parasites per μl, the clinical episodes of malaria were markedly fewer among the Fulani than in the Mossi and Rimaibé, who had very similar morbidity levels (Kruskal–Wallis test, P = 0.009).
DISCUSSION
The comparative analysis of the response to P. falciparum malaria in the West African ethnic groups Fulani, Rimaibé, and Mossi was greatly facilitated by their sympatric distribution in a hyperendemic area. Their expected similar exposure to high inoculation rates was clearly supported by the entomological data, and, given the information available on vector dispersion (15), it is also reasonable to assume a random challenge of the individuals compared with the same parasite variants. This condition and the absence of significant heterogeneities in the use of antimalaria drugs or of mosquito protection methods allow us to evaluate possible relationships between the response to malaria and ethnic group characteristics, either cultural or genetic.
The results demonstrated clear interethnic parasitological, clinical, and immunological differences in the response to P. falciparum, and the findings were equally consistent when comparing samples from neighboring villages or coresident in the same village (syntopic). The parasite prevalences in the Mossi and Rimaibé were very similar and within the range expected under the local conditions of hyperendemic transmission, while the Fulani showed much lower infection rates in all age groups and in five consecutive cross-sectional surveys. Constant P. falciparum negativity across the five surveys was observed in 36% of the Fulani older than 10 years, while the corresponding figure for the Mossi and Rimaibé was only 1%. These parasitological differences were fully coherent with the results of 2-month active case detection on the age group 0–10 years. The fever episodes with parasite-positive slides had a mean parasite density 3-fold lower in the Fulani, and an even greater difference was found in the average incidence of clinical episodes defined by fever >38°C and parasite counts over 12,800 parasites per μl of blood. Among the children with the highest parasitemias (>25,600), the Fulani were only exceptionally represented, which suggests their protection against severe malaria. Furthermore, the above parasitological and clinical evidence of a higher resistance of the Fulani to P. falciparum was found associated with their stronger humoral reactivity against all parasite antigens tested (namely CSP, TRAP, MSA-1, Pf155, and Pf332), without the evidence of more intense antigenic stimulations. The case of the repetitive NANP epitope of the CSP was particularly well-documented. This antigen elicits an antibody response that has been reported to reflect the degree of exposure to sporozoite-infected mosquitoes (12, 16, 17), and we actually recorded intraethnic variations fully consistent with the seasonal changes in the intensity of transmission. However, the interethnic comparison revealed anti-CSP responses constantly higher in the Fulani, while the entomological data support similar exposure levels of the three ethnic groups. This implies the capacity of the Fulani of mounting a stronger antibody response than the Mossi and Rimaibé.
Further interethnic comparative studies in other West African areas should verify to what extent the efficient Fulani response to P. falciparum malaria represents a general trait of the ethnic group. The available literature does not seem to provide specific support to our parasitological findings. Parasite prevalences lower than expected were reported in population samples mostly constituted by Fulani in Northern Nigeria (18) and The Gambia (19), but the authors were not in position to evaluate the role played by the ethnic variable. A few relevant references deal with immunological and clinical investigations. Riley et al. (20), comparing different ethnic groups in The Gambia, reported the higher antibody response of the Fulani to the 3′ repeat sequence (EENV)6 of the Pf155 ring-infected erythrocyte surface antigen (as shown in our samples) and pointed out the association of this immune reactivity with resistance to symptomatic malaria (20) and its correlation with one class II DQA–DQB combination (serological specificity DQw2) particularly common in the Fulani ethnic group (21). Other possible pertinent data refer to splenomegaly, which was reported to be more frequent in the Fulani from Nigeria (22) and The Gambia (23) than in the sympatric ethnic groups. These studies also noted higher IgM levels in the Fulani.
None of the cultural characteristics of the Fulani, including their dietary habits, may account for the parasitological, clinical, and immunological differences observed, particularly in view of their magnitude and stability. This is further supported by the Rimaibé response to malaria, which diverges from that of the Fulani in spite of important cultural similarities, and closely corresponds to that of the Mossi, in agreement with the anthropological relationships. The consequent working hypothesis is that we are dealing with the expression of genetic differences between the non-negroid Fulani and the negroid Rimaibé and Mossi.
Although the anthropological peculiarity of the Fulani among West African populations is well-established, intraethnic variations and genetic distances from other ethnic groups are poorly defined. Recent studies on GM polymorphism in Eastern Senegal (24) and on HLA class I (25) and II (26) in The Gambia indicate rather small genetic distances between the Fulani and their neighbors. However, preliminary analysis of our samples from Burkina Faso suggests a relatively higher degree of genetic divergence: the typing of 107 HLA class I alleles confirmed the close relationship between the Mossi and Rimaibé, while the Fulani were found clearly separated and less distant from caucasoids. There were 15 alleles with at least one interethnic statistically significant difference, and in 14 of them, the Mossi and Rimaibé frequencies turned out to be very similar, but different from those of Fulani.‖ Furthermore, the study of HbC frequencies (D.M. & R. Scozzari, unpublished) showed identical values in the Mossi and Rimaibé (0.117 ± 0.018 and 0.117 ± 0.020, respectively), which were different from that of the Fulani (0.059 ± 0.020).
Based on these data, the Fulani of our study area are distinct enough to hypothesize a degree of isolation of their gene pool compatible with the maintenance of a genetic diversity in the response to P. falciparum. Such a genetic differentiation appears unrelated to the sickle cell trait and to the known HLA class I factor of malaria protection. Actually, similar HbS gene frequencies were found in the three ethnic groups (Mossi, 0.024 ± 0.008; Rimaibé, 0.030 ± 0.011; and Fulani, 0.022 ± 0.013; D.M. & R. Scozzari, unpublished). Concerning the HLA B∗53, which has been associated with protection from severe malaria (2), a lower gene frequency was observed‖ among the Fulani (0.061 ± 0.024) as compared with the Mossi and the Rimaibé (0.189 ± 0.038 and 0.202 ± 0.041, respectively). Even if we cannot exclude at present the possible role of other “protective” erythrocytic factors, such as thalassemias, glucose-6-phosphate dehydrogenase-deficiencies, and ovalocytosis (1, 27), these are not probably involved, since their expressions would not easily explain our immunoparasitological findings.
The fact that the higher antibody response of the Fulani was observed for all antigens tested argues against an HLA class II-based explanation. The possible role of factors encoded by genes outside the HLA class II region in the regulation of humoral immune response against P. falciparum antigens has been suggested by the twins study by Sjöberg et al. (28). Possible alternative immunological mechanisms of protection may involve interethnic differences operating at levels such as B-cell function, antigen processing (29), and expression of aspecific factors such as interleukin 4, which is an important modulator for the differentiation of precursor T-helper cells to the T-helper 2 or T-helper 2-like cells that mediate humoral immunity and assist in antibody production (30–33). Moreover, Troye-Blomberg et al. (34) reported that the production of interleukin 4 by activated human cells is associated with elevated levels of antibodies to malaria antigens. The hypothesis of a stronger activation of the immune system in the Fulani is also suggested by the higher frequency in this ethnic group of the hyperreactive malarial splenomegaly syndrome (22, 23).
Acknowledgments
The study was based at the Centre National de Lutte contre le Paludisme (CNLP) of the Ministry of Health of Burkina Faso supported by the Programma di Assistenza Tecnica della Direzione Generale per la Cooperazione allo Sviluppo of the Italian Ministry of Foreign Affairs. We are particularly grateful to the villagers of Watinoma, Barkoundouba, and Barkoumbilen for their collaborative attitude throughout the investigation. We are deeply indebted to the personnel of the Immunoparasitology and Entomology laboratories of the CNLP and particularly to Mr. Albert Yameogo for his skillful technical assistance. Mrs. S. Bagalino and G. Croce of the Institute of Parasitology of the University of Rome “La Sapienza” are also acknowledged for the processing of part of the materials. Finally we thank Dr. L. Miller, Dr. L. Molineaux, Dr. B. Greenwood, and Prof. Guido Modiano for useful comments and Dr. A. Crisanti who provided MSA-1 and TRAP fragments. The study was partially supported by the World Health Organization, Division of Control of Tropical Diseases, the Fondazione Pasteur-Cenci Bolognetti of the University of Rome “La Sapienza,” and the Italian Ministry for Research.
Footnotes
Abbreviations: CSP, circumsporozoite protein; TRAP, thrombospondin-related anonymous protein; MSA, major surface antigen.
Modiano, D., Luoni, G., Petrarca, V., De Luca, M., Marsh, S. G. E., Coluzzi, M., Bodmer, G. J. & Modiano G., Proceedings of the 12th International Histocompatibility Workshop and Conference, June 9–12, 1996, Paris.
References
- 1.Miller L H. In: Malaria: Principles and Practice of Malariology. Wernsdorfer W H, McGregor I Sir, editors. Edinburgh: Churchill Livingstone; 1988. pp. 488–500. [Google Scholar]
- 2.Hill A V S, Allsopp C E M, Kwiatkowski D, Anstey N M, Twumasi P, Rowe P A, Bennett S, Brewster D, McMichael A J, Greenwood B M. Nature (London) 1991;352:595–600. doi: 10.1038/352595a0. [DOI] [PubMed] [Google Scholar]
- 3.Aidoo M, Lalvani A, Allsopp C E M. Lancet. 1995;345:1003–1006. doi: 10.1016/s0140-6736(95)90754-8. [DOI] [PubMed] [Google Scholar]
- 4.Piazza A, Belvedere M C, Bernoco D, Conighi C, Contu L, Curton E S, Mattiuz P L, Mayr W, Richiardi P, Scudeller G, Ceppellini R. In: Histocompatibility Testing: 1972. Dausset J, Colombani J, editors. Baltimore: Williams & Wilkins; 1973. pp. 73–84. [Google Scholar]
- 5.Armstrong J C. Trans R Soc Trop Med Hyg. 1978;72:342–344. doi: 10.1016/0035-9203(78)90123-2. [DOI] [PubMed] [Google Scholar]
- 6.Oomen J M V, Meuwissen J H E T, Gemert W. Trop Geogr Med. 1978;31:587–606. [PubMed] [Google Scholar]
- 7.Terrenato L, Shrestha S, Dixit K A, Luzzatto L, Modiano G, Morpurgo G, Arese P. Ann Trop Med Parasitol. 1988;82:1–11. doi: 10.1080/00034983.1988.11812202. [DOI] [PubMed] [Google Scholar]
- 8.Modiano G, Morpurgo G, Terrenato L, Novelletto A, Di Rienzo A, Colombo B, Purpura M, Mariani M, Santachiara-Benerecetti S, Brega A, Dixit K A, Shrestha S L, Lania A, Wanachiwanawin W, Luzzatto L. Am J Hum Genet. 1991;48:390–397. [PMC free article] [PubMed] [Google Scholar]
- 9.Modiano D, Petrarca V, Sirima B S, Bosman A, Nebié I, Lamizana L, Esposito F, Coluzzi M. Parassitologia (Rome) 1995;37:255–259. [PubMed] [Google Scholar]
- 10.Stenning D J. In: Peoples of Africa. Gibbs J L, editor. Rinehart and Winston, New York: Holt; 1965. pp. 363–401. [Google Scholar]
- 11.Skinner E P. The Mossi of the Upper Volta. Stanford, CA: Stanford Univ. Press; 1964. [Google Scholar]
- 12.Esposito F, Lombardi S, Modiano D, Zavala F, Reeme J, Lamizana L, Coluzzi M, Nussenzweig R S. Trans R Soc Trop Med Hyg. 1988;82:827–832. doi: 10.1016/0035-9203(88)90007-7. [DOI] [PubMed] [Google Scholar]
- 13.Petrarca V, Petrangeli G, Rossi P, Sabatinelli G. Parassitologia (Rome) 1986;28:41–61. [PubMed] [Google Scholar]
- 14.Habluetzel, A., Esposito, F. & Lombardi, S. (1989) Trans. R. Soc. Trop. Med. Hyg. 83, Suppl., 15–19. [DOI] [PubMed]
- 15.Costantini C, Song G L, Della Torre A, Sagnon N F, Coluzzi M, Taylor C. Med Vet Entomol. 1996;10:203–219. doi: 10.1111/j.1365-2915.1996.tb00733.x. [DOI] [PubMed] [Google Scholar]
- 16.Esposito F, Lombardi S, Modiano D, Zavala F, Reeme J, Lamizana L, Coluzzi M, Nussenzweig R S. Parassitologia (Rome) 1986;28:101–105. [PubMed] [Google Scholar]
- 17.Druilhe P, Pradier O, Marc I P, Miltgen F, Mazier D, Parent G. Infect Immun. 1986;53:393–397. doi: 10.1128/iai.53.2.393-397.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Molineaux L, Gramiccia G. The Garki Project: Research on the Epidemiology and Control of Malaria in the Sudan Savanna of West Africa. Geneva: W. H. O.; 1980. p. 311. [Google Scholar]
- 19.Riley E M, Allen S J, Bennett S, Thomas P J, O’Donnel A, Linsday S V, Good M F, Greenwood B M. Trans R Soc Trop Med Hyg. 1990;84:648–657. doi: 10.1016/0035-9203(90)90133-y. [DOI] [PubMed] [Google Scholar]
- 20.Riley E M, Allen S J, Troye-Blomberg M, Bennett S, Perlmann H, Andersson G, Smedman L, Perlmann P, Greenwood B M. Trans R Soc Trop Med Hyg. 1991;85:436–443. doi: 10.1016/0035-9203(91)90207-f. [DOI] [PubMed] [Google Scholar]
- 21.Riley E M, Olerup O, Bennett S, Rowe P, Allen S J, Blackman M J, Troye-Blomberg M, Holder A A, Greenwood B M. Int Immunol. 1992;4:1055–1063. doi: 10.1093/intimm/4.9.1055. [DOI] [PubMed] [Google Scholar]
- 22.Bryceson A D M, Fleming A F, Edington G M. Acta Trop. 1976;33:424–426. [PubMed] [Google Scholar]
- 23.Greenwood B M, Groenendaal F, Bradley A K, Greenwood A M, Shenton F, Tulloch S. Ann Trop Med Parasitol. 1987;81:345–354. doi: 10.1080/00034983.1987.11812130. [DOI] [PubMed] [Google Scholar]
- 24.Blanc M, Sanchez-Mazas A, Van Blyenburgh N H, Sevin A, Pison G, Langaney A. Am J Hum Genet. 1990;46:383–392. [PMC free article] [PubMed] [Google Scholar]
- 25.Allsopp C E, Harding R M, Taylor C, Bunce M, Kwiatkowski D, Anstey N, Brewster D, McMichael A J, Greenwood B M, Hill A V S. Am J Hum Genet. 1992;50:411–421. [PMC free article] [PubMed] [Google Scholar]
- 26.Olerup O, Troye-Blomberg M, Schreuder G M T, Riley E M. Proc Natl Acad Sci USA. 1991;88:8480–8484. doi: 10.1073/pnas.88.19.8480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Edington G M, Watson-Williams E J. In: Abnormal Haemoglobins in Africa. Jonxis J H P, editor. Oxford: Blackwell Scientific; 1965. p. 393. [Google Scholar]
- 28.Sjöberg K, Lepers J P, Raharimalala L, Larsson A, Olerup O, Marbiah N T, Troye-Blomberg M, Perlmann P. Proc Natl Acad Sci USA. 1992;89:2101–2104. doi: 10.1073/pnas.89.6.2101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Corradin G. Immunol Lett. 1990;25:11–13. doi: 10.1016/0165-2478(90)90082-2. [DOI] [PubMed] [Google Scholar]
- 30.Mosmann T R, Coffman R L. Annu Rev Immunol. 1989;7:145–173. doi: 10.1146/annurev.iy.07.040189.001045. [DOI] [PubMed] [Google Scholar]
- 31.Gollob K J, Coffman R L. J Immunol. 1994;152:5180–5188. [PubMed] [Google Scholar]
- 32.Erard F, Wild M T, Garcia-Sanz J A, Le Gros G. Science. 1993;260:1802–1805. doi: 10.1126/science.8511588. [DOI] [PubMed] [Google Scholar]
- 33.Kariv I, Hardy R R, Hayakawa K. Eur J Immunol. 1994;24:549–557. doi: 10.1002/eji.1830240309. [DOI] [PubMed] [Google Scholar]
- 34.Troye-Blomberg M, Riley E M, Kabilan L, Holmberg M, Perlmann H, Andersson U, Heusser C H, Perlmann P. Proc Natl Acad Sci USA. 1990;87:5484–5488. doi: 10.1073/pnas.87.14.5484. [DOI] [PMC free article] [PubMed] [Google Scholar]