

Abstract
The Candida albicans genes, CST20 and HST7, were cloned by their ability to suppress the mating defects of Saccharomyces cerevisiae mutants in the ste20 and ste7 genes, which code for elements of the mating mitogen-activated protein (MAP) kinase pathway. These Candida genes are both structural and functional homologs of the cognate Saccharomyces genes. The pattern of suppression in Saccharomyces is related to their presumptive position in the MAP kinase cascade. Null alleles of these genes were constructed in Candida. The Candida homozygous null mutants are defective in hyphal formation on some media, but are still induced to form hyphae by serum, showing that serum induction of hyphae is independent of the MAP kinase cascade. The Candida heterozygotes CST20/cst20 and HST7/hst7 are also defective in hyphal formation. This lack of dominance of the wild-type allele suggests that gene dosage is important in Candida.
Candida albicans, the most frequently isolated fungal pathogen in humans, is dimorphic; it switches between growth as a budding yeast form and a filamentous hyphal or pseudohyphal form. This switch is induced by many different environmental cues including high temperature, neutral pH, and serum. Dimorphism is thought to contribute to Candida’s virulence (1). The direct isolation of mutations that prevent switching is difficult because C. albicans is diploid and has no known sexual cycle. To circumvent these problems, several studies have taken the “candidate gene” approach in which a gene is identified that is likely to be involved in dimorphism, and Candida strains homozygous for a null allele of that gene are constructed (2, 3). The gene is deemed to be required for dimorphism if the null mutant shows a defect in the induction of hyphae by one of the external cues.
One approach used to identify candidate genes is to clone Candida homologs of Saccharomyces cerevisiae genes known to regulate filamentous growth. Saccharomyces also switches from a yeast form to a filamentous pseudohyphal form, when the yeast cells are starved for nitrogen. This switch requires elements of the mating signal transduction pathway including the protein kinases Ste20, a p65PAK kinase homolog, Ste11 [mitogen-activated protein (MAP) kinase kinase kinase] and Ste7 (MAP kinase kinase), and the transcription factor Ste12 (4). The C. albicans homolog of STE12, CPH1, was cloned by its ability to enhance filamentous growth of Saccharomyces (5). Cph1 shows considerable amino acid identity with Ste12 and suppresses both the filamentation and mating defect of the Saccharomyces ste12 mutant (6). The cph1/cph1 null mutant is defective in hyphal formation under a number of conditions that induce wild-type Candida strains to filament. However, cph1/cph1 strains are still induced to form normal hyphae by serum. This result implicates CPH1 in Candida dimorphism, but suggests that serum induces hyphae by a Cph1-independent pathway.
The finding that the Candida CPH1 gene complemented both the filamentation and mating defect of the Saccharomyces ste12 mutant, suggested that other members of a Candida MAP kinase (MAPK) cascade could be isolated by suppression of the corresponding Saccharomyces mutant. In this report we describe the isolation of Candida homologs of STE20 and STE7, two other kinases of the Saccharomyces mating MAPK cascade. We constructed strains heterozygous and homozygous for null alleles of these genes in Candida and found that both heterozygotes and homozygotes show defects in hyphal formation. However, these mutant strains still form hyphae in response to serum.
MATERIALS AND METHODS
Isolation of C. albicans Genes That Suppress the Mating Defects of ste7 and ste20.
A C. albicans genomic library, prepared from strain 1006 on a URA3, 2 micron vector (5), was used to transform Saccharomyces strains JKY1(ste7) and JKY40(ste20) to Ura+ by a modified lithium acetate method (7). To identify clones with restored mating ability, 105 transformants were replica plated to the tester lawn JBY311 on yeast extract/peptone/dextrose (YPD), incubated for 20 h, and replica plated to minimal medium where only diploids could grow. Clones that mated were identified and the suppressing plasmids were isolated.
Plasmids.
pJK7 is an isolate from this library containing the HST7 open reading frame. The insert of pJK7 cloned into Bluescript SK+ (Stratagene), pJK19, was used for sequencing. The disruption construct, pJK41, was made by inserting a BglII and an SphI site by site-directed mutagenesis and replacing bases 205-1765 of the HST7 open reading frame with the hisG-URA3-hisG cassette on the BglII/SphI fragment of pMB7 (8). To complement the homozygous deletion (hst7/hst7) Candida strains with the intact HST7 gene, the polylinker BamHI and XbaI restriction sites were destroyed by blunting and religation, and a BamHI and XbaI site was introduced into pJK19 by site-directed mutagenesis between bases 268 and 312 downstream of the HST7 open reading frame. The Candida URA3 gene on the BamHI/XbaI fragment of pMB7 was cloned into these sites to make pJK58. The insert of pJK58, cut out by the polylinker sites KpnI and SacI, was used to replace one of the two disrupted copies of HST7 with an intact copy by means of a double crossover event.
The library isolate pJK34 contains the full-length CST20 open reading frame on a 7.2-kb insert. The isolate pJK25 contains a 295-bp 5′ truncated form of this gene on a 4.8-kb insert. The pJK25 insert, cloned into Bluescript to yield pJK42, was used for sequencing most of the gene. The first 0.3 kb were sequenced from the pJK34 insert cloned into Bluescript, pJK62. The gene disruption construct of CST20, pJK51, was made by replacing the BglII/PstI fragment of pJK42 with the hisG-URA3-hisG cassette from pMB7. To reintroduce an intact CST20 copy into the cst20/cst20 deletion strains, a complementing integrating construct was made containing CST20 with the URA3 selectable marker at its 3′ end, by cloning the URA3 gene on the BamHI/XbaI fragment of pMB7 into the HpaI site of pJK42 to make pJK60, whose insert was excised to induce a double crossover event with the polylinker sites KpnI and SacII. A second construct for this purpose, pJK61, was made by cloning the pJK25 insert into pRC2312, a vector containing the Candida URA3 and LEU2 genes, which can be linearized at the unique SalI site in LEU2 for integration at the chromosomal locus (9).
pHL14 contains the CPH1 gene (5), a Candida homolog of the S. cerevisiae gene STE12, and was used in the assays for complementation of Saccharomyces mating and pseudohyphal growth.
S. cerevisiae Strains.
For mating assays, all isogenic haploid strains were derived from JBY597 (10) MATa ura3-52 leu2-3 112 trp1Δ63 his3Δ200 or his3-11,15 ade2 kss1 lys2::FUS1::LacZ Gal+. JKY1 (ste7::LEU2) was constructed from this strain by one-step gene replacement with pDH 90 (from Don Higgins and Kelly Tatchell, North Carolina State University, Raleigh). JKY40 (ste20::TRP1), whose sterility phenotype is leaky, was constructed from JBY597 with pEL46-2 (11). Gene disruptions were confirmed by Southern analysis. JKY86 was derived from JBY837 (MATa ste11::URA3) and JKY87 was constructed from JBY841 (MATa ste12::URA3) (10), by disrupting URA3 with HIS3 using the plasmid B2388 (from Y. Kassir, Technion, Haifa, Israel). JBY311 (MATα lys9 ura3-52) (10) was used as a tester strain for mating. For assays of diploid pseudohyphal growth, we used isogenic strains in the Σ1278b background containing disruptions of STE20, STE11, STE7, and STE12 as described in ref. 4. The STE/STE wild-type strain was L5366.
S. cerevisiae Mating and Pseudohyphal Growth Assays.
To determine the effect of a cloned Candida gene on the behavior of a Saccharomyces mutant, we transformed Saccharomyces with a plasmid containing the Candida gene and assayed the behavior of two transformants for mating or pseudohyphal growth. Mating assays were done using the replica patch test (12), and pseudohyphal growth was assayed by constructing the appropriate diploids and observing colony morphology after 5 days growth on SLAD medium without histidine (13). For controls, the 2 micron URA3 vector used for the construction of the library, B2205, was used.
Candida Growth and Genetic Analysis.
C. albicans was routinely grown in YPD medium and at 37°C, unless otherwise noted. Sequential disruption of both alleles of the Candida genes was performed using the strategy developed for S. cerevisiae by Alani et al. (14), as adapted for Candida by Fonzi and Irwin (8). This strategy uses the Candida URA3 gene, flanked by two repeats derived from the Salmonella typhimurium hisG gene, as a selectable marker. Segregants that have lost the URA3 gene by recombination between the repeats can be selected on medium containing 0.1% (wt/vol) 5-fluoroorotic acid and 0.2 mM uridine, and the second allele of the gene can be disrupted reusing URA3 as the selectable marker.
The strains used for gene disruption were CAI4 (ura3::imm434/ura3::imm434) (8) and JKC18, which is derived from CAI4 and is cph1/cph1 ura3/ura3 (5). Transformation and selection for Ura− heterozygotes were performed as described (5). Transformants were screened by PCR for homologous recombination of the transforming construct. Homologous recombination was confirmed by Southern blot analysis. Integration of the intact wild-type gene for rescue of the hst7/hst7 and cst20/cst20 homozygous disruption strains was verified by Southern blot analysis, but the location was not determined. For the hst7/hst7 strains tranformed with an intact copy of HST7 on pJK58, expression of the gene was confirmed by Northern blot analysis. The wild-type strain used for assay of hyphal growth was a clinical isolate, sc5314 (15), the parent strain of CAI4. To analyze the effect of the introduced mutations on hyphal growth, mutants were assayed before loss of the URA3 selectable marker, because Ura− strains are impaired in hyphal formation. Hyphal growth of Ura3+ strains, whose URA3 gene was at the locus of the disrupted gene in the hisG-URA3-hisG cassette, was assayed at 37°C on Spider medium (5), on medium 199 buffered to pH 7 with Hepes (3) and solidified with 2% agar, on Lee’s medium (16) solidified with 2% agar, and on solid agar medium containing bovine calf serum (Sigma), made by adding serum at concentrations between 0.5 and 20% to a mixture of water and 2% agar. Hyphal growth of single cells was assayed at 37°C in liquid Lee’s medium (16) and in 20% calf serum.
RESULTS
Candida Homologs of S. cerevisiae STE7 and STE20.
Functional Candida homologs of S. cerevisiae STE7 and STE20 were isolated by complementation of the mating defect of Saccharomyces ste7 and ste20 mutants. Transformation of a ste7 mutant with a Candida library cloned in a high copy vector yielded CPH1, a homolog of Saccharomyces STE12, and HST7, which has been reported to be the structural and functional homolog of Saccharomyces Ste7 (17). Transformation of a ste20 mutant yielded HST7 and 10 distinct isolates of a DNA segment, which could encode a protein with homology to S. cerevisiae Ste20. The predicted 1230 amino acid protein is a member of the Ste20/p65PAK protein family based on the domains it shares with other members of this family. The carboxyl-terminal kinase domain of this protein is 78% identical (Fig. 1B) with the Ste20 kinase domain (11, 18). In addition, Cst20 is 69% identical with Ste20 in a putative Cdc42 binding domain (Fig. 1A), which it shares with other members of the Ste20/p65PAK kinase family like the Schizosaccharomyces pombe Ste20 homolog Pak1/Shk1, and the rat protein p65PAK (20–22). The amino-terminal portion of the Candida protein is longer than that of the Saccharomyces Ste20. Because of its functional and structural homology to the Saccharomyces STE20 gene, this Candida gene is called CST20.
Figure 1.

Cst20 and Ste20 share homologous Cdc42-binding domains and catalytic domains. Translation such that CTG codes for serine (19). (A) Cdc42 binding domains. (B) Kinase domains.
Candida Genes Function in Saccharomyces.
HST7 clones restore both mating and pseudohyphal growth (Fig. 2) to S. cerevisiae containing a ste7 disruption, as previously reported (17). Plasmids containing enough of the 5′ portion of the gene to express the putative Cdc42 binding domain of CST20 (pJK25 and pJK34) suppress both the mating and pseudohyphal defects of a Saccharomyces ste20 mutant. In contrast, plasmids with a 5′ truncation of CST20 lacking the presumptive Cdc42 binding domain (pJK21 and pJK29) suppress the mating defect, but not the filamentation defect (data not shown). These data suggest that there may be distinct domains of Ste20 that are devoted to each morphogenetic process. The Candida homologs of the Saccharomyces STE7 and STE12 genes bypass mutations in genes that act earlier in the MAPK cascade: CPH1 suppresses the pseudohyphal defect in ste20, ste11, ste7, and ste12 strains and partially suppresses the mating defect in ste20, ste11, and ste7 mutants. HST7 suppresses the pseudohyphal and mating defects in ste20, ste11, and ste7, but not ste12. CST20 fails to suppress the mating or pseudohyphal defect in any sterile mutants tested except for ste20.
Figure 2.
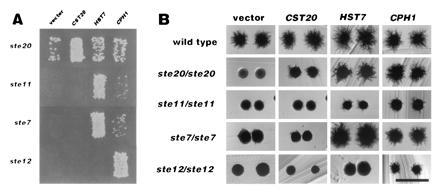
(A) Candida genes suppress the mating defects in haploid S. cerevisiae ste mutant strains. Saccharomyces strains were transformed with the 2-micron vector B2205 (column 1), and with plasmids based on this vector: pJK25 carrying the CST20 gene (column 2), pJK7 carrying HST7 (column 3), and pHL14 carrying CPH1 (column 4). Haploid Saccharomyces strains are JKY40 (ste20), JKY1 (ste7), and JKY 87 (ste12). Patches of transformants were grown on selective medium to maintain the plasmid, replica plated to YPD plates covered with lawns of the mating tester strain JBY311, incubated for 4 h, replica plated to minimal medium, and incubated for 2 days. (B) Candida genes complement pseudohyphal growth defects in diploid ste/ste S. cerevisiae strains containing mutations in genes encoding components of the mating MAPK cascade. Ste+ (L5366), ste20/ste20 (L5624), ste11/ste11 (L5625), ste7/ste7 (L5626), and ste12/ste12 (L5627) were transformed with Candida genes on 2-micron plasmids, as indicated above the columns. The same plasmids were used as in the mating patch assays. Transformants were streaked on SLAD medium and incubated for 5 days. (Bar = 1 mm.)
Candida Heterozygotes for Null Alleles of HST7 and CST20.
C. albicans heterozygotes made by gene disruption (HST7/hst7 or CST20/cst20) (Fig. 3) do not have the same hyphal morphology as the parent strain. Three independent single disruptions of HST7 were constructed. The three independent Ura+ HST7/hst7 heterozygotes all show reduced hyphal formation on Spider medium as compared with the HST7/HST7 parent (Fig. 4A). These heterozygotes show variability in the reduction of hyphal formation between strains and from colony to colony (Fig. 4). Overall, they show more peripheral hyphal growth than their hst7/hst7 derivatives. Two independent CST20/cst20 disruptants were constructed. Both of these heterozygotes also show a drastic reduction of peripheral hyphal growth on Spider plates as compared with the CST20/CST20 parent (Fig. 5B). The CST20/cst20 heterozygotes are more homogeneous than the HST7/hst7 heterozygotes and have a greater reduction in peripheral hyphal growth.
Figure 3.
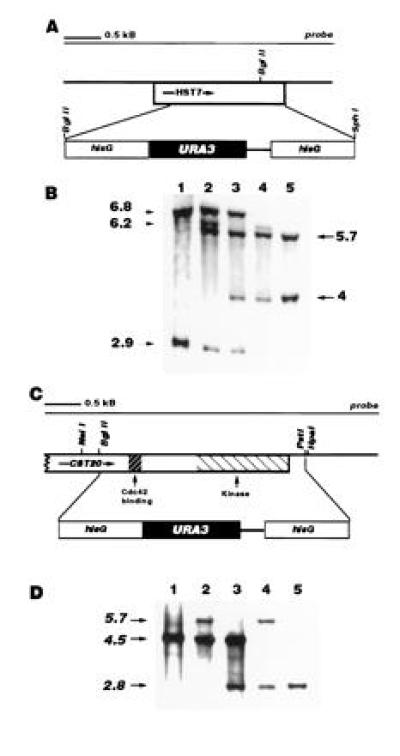
(A) HST7 deletion construct: map of pJK41. The BglII site, with which the hisG-URA3-hisG cassette was inserted, and the SphI site were created by site-directed mutagenesis and therefore not present in the wild-type gene. (B) Southern blot of HST7 deletion mutants: Candida genomic DNA digested with BglII. Lane 1, wild type. The 6.8-kb band results from digestion at the BglII site in the HST7 open reading frame and a site on the chromosome upstream of the gene, the 2.9-kb band results from digestion the BglII site in the open reading frame and a site on the chromosome downstream of it. Lane 2, HST7/hst7::hisG-URA3-hisG. Integration of the disruption construct at one of the two copies of HST7 results in loss of the BglII site in the open reading frame and gain of a new site in the construct, creating a 6.2-kb band between the new BglII site and the 3′ chromosomal BglII site, and a 5.7-kb band between the 5′ chromosomal site and the new BglII site. Lane 3, HST7/hst7::hisG. Eviction of URA3 and one hisG repeat results in a decrease in size of the fragment between the new BglII site and the 3′ site from 6.2 to 4 kb. Lane 4, hst7::hisG-URA3-hisG/hst7::hisG. Integration of the disruption construct into the remaining HST7 allele results in loss of the wild-type bands. Lane 5, hst7::hisG/hst7::hisG. Arrows indicate kilobases. Fragment sizes are approximate. (C) CST20 deletion construct: map of pJK51. The construct lacks the first 0.3 kb of the open reading frame. (D) Southern blot of CST20 mutants: Candida genomic DNA digested with NsiI. Lane 1, wild type. The band results from digestion at the NsiI site in the open reading frame and a chromosomal NsiI site 4.5 kb downstream of it. Lane 2. CST20/cst20::hisG-URA3-hisG. Lane 3, CST20/cst20::hisG. Lane 4, cst20::hisG-URA3-hisG/cst20::hisG. Lane 5, cst20::hisG/cst20::hisG. Arrows indicate kilobases. Fragment sizes are approximate.
Figure 4.
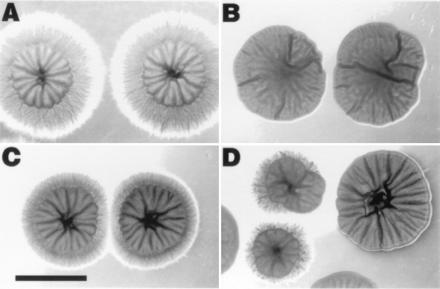
Filamentous growth defects of HST7/hst7 heterozygotes. The extent of filamentation varies among strains and from colony to colony. Incubation for 5 days on Spider medium. (A) Wild type SC5314. (B) JKC48. (C) JKC63. (D) JKC52. (Bar = 5 mm.)
Figure 5.
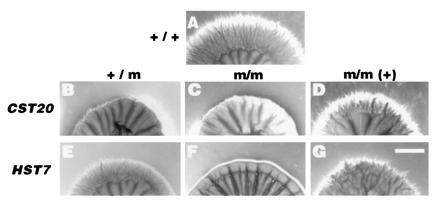
HST7 and CST20 are required for hyphal growth on Spider medium. (A) Wild-type strain SC5314 (CST20/CST20 HST7/HST7 URA3/URA3). (B–D) CST20 mutants. (E–G) HST7 mutants. (B) CST20/cst20::hisG-URA3-hisG (JKC91). (C) cst20::hisG/cst20::hisG-URA3-hisG (JKC97). (D) cst20::hisG/cst20::hisG/cst20-ura3 (JKC 178). (E) HST7/hst7::hisG-URA3-hisG (JKC63). (F) hst7::hisG/hst7::hisG-URA3-hisG (JKC131). (G) hst7::hisG/hst7::hisG/HST7-URA3 (JKC173). Candida strains were incubated for 5 days on Spider medium. (Bar = 2 mm.)
Candida Homozygotes for Null Alleles of HST7 and CST20.
Deletion of both alleles of the HST7 gene (hst7/hst7) by disruption of the remaining copy in any one of the three HST7/hst7 heterozygotes resulted in complete loss of hyphal formation at the periphery of colonies (Fig. 5F) on Spider medium, even after 3 weeks of incubation. Sparse short hyphae growing vertically into the agar are formed at the center of the colonies. Two cst20/cst20 disruption strains of Candida albicans, each derived from an independently constructed CST20/cst20 heterozygote have similar phenotypes (Fig. 5C). They have a reduction of peripheral hyphal growth as compared with both the heterozygote and CST20/CST20 parent; the hyphae are both shorter and more sparse. However, peripheral hyphal growth in the cst20/cst20 strains is not completely abolished on Spider medium, as it is in hst7/hst7 strains. On Lee’s medium and on medium 199 (pH 7) solidified with 2% agar, hyphal growth of the strains disrupted for HST7 and CST20 is also reduced compared with the wild-type parent, but the difference is less drastic than on Spider medium (data not shown).
To verify that hyphal phenotypes of the hst7/hst7 and cst20/cst20 homozygotes are a consequence of the deletions we constructed, we reintroduced a functional copy of the cognate genes and observed that hyphal formation was restored (Fig. 5 D and G). In neither case does reintroduction of a functional copy of the gene restore hyphal growth equivalent to that observed in the wild-type parent from which the strains were derived.
Despite the failure of the cst20/cst20 and the hst7/hst7 homozygotes to form significant hyphae on Spider medium, both strains form hyphae on agar plates containing serum. Germ tube formation—i.e., outgrowth of a hypha from a yeast-form cell—is also unaffected in cst20/cst20 and hst7/hst7 strains in 20% liquid serum and in liquid Lee’s medium. None of the strains carrying mutations in the known Candida homologs of the MAPK cascade (cst20/cst20, hst7/hst7, or cph1/cph1) blocks the serum induction of hyphae. One explanation for the failure of the mutants to block serum induction could be that these genes function in different, but redundant signaling pathways, either of which, when activated by serum, induces hyphal formation. To examine this question, we constructed hst7/hst7 cph1/cph1 and cst20/cst20 cph1/cph1 double mutants. The phenotypes of the double mutants on Spider medium resemble that of the single mutant with respect to hyphal formation: cst20/cst20 cph1/cph1 looked like cst20/cst20 and cph1/cph1 hst7/hst7 looked like hst7/hst7. However, both the cst20/cst20 cph1/cph1 and the hst7/hst7 cph1/cph1 strains still form hyphae in response to serum (data not shown).
DISCUSSION
Our results show that C. albicans has many of the elements of the filamentation MAPK cascade that has been described in the budding yeast S. cerevisiae. The Candida genes are both structural and functional homologs of the Saccharomyces genes. Previous reports have shown that CPH1 and HST7 are both structural and functional homologs of Saccharomyces STE12 and STE7, respectively (5, 6, 17). The results demonstrate that Candida CST20 like the HST7 and CPH1 genes functions in Saccharomyces to complement both the filamentation and mating defects of the cognate Saccharomyces mutants. High copy (2 micron) constructs of the Candida homologs expressed in Saccharomyces bypass upstream but not downstream mutant defects. For example, CST20 can bypass the defects of a ste20 mutant, but not ste11, ste7, or ste12, whereas HST7 bypasses ste20, ste11, and ste7.
Here we show that in Candida as in Saccharomyces these MAPK genes function to signal filamentation. cst20/cst20 and hst7/hst7 Candida strains have defects in filamentation on solid agar medium, forming only a few short hyphae instead of the florid filaments that emanate from the wild-type parent strain. Despite this defect, none of the null mutants blocks the Candida serum response—neither induction of filaments on solid medium nor germ tubes are affected. We examined the possibility that in Candida these genes are not in the same pathway, but are different elements of two parallel pathways, either of which can convey the inductive signal from serum. For example, CST20 might be in one pathway and CPH1 in another. If this hypothesis were correct then the double mutants (e.g., cst20/cst20 cph1/cph1) should have been defective in the serum response. However, the double mutants still respond to serum. Serum must activate hyphal formation via a distinct signal transduction pathway. Based on the phenotypes of the double mutants, it is likely that these Candida genes, CST20, HST7, and CPH1 like their Saccharomyces counterparts, are in the same pathway, though our experiments do not rule out more complicated models.
An unexpected feature of our analysis is the finding that all three heterozygotes (CST20/cst20, HST7/hst7, and CPH1/cph1) have obvious defects in filamentation, which suggests that the null alleles are not recessive. Furthermore, the defect in filamentous growth of the homozygote is not fully suppressed by introduction of a functional allele. By contrast, in Saccharomyces a single copy of a functional gene suffices to suppress the filamentation defect for any of the steps in the MAPK cascade. This phenomenon may be a reflection of the absence of a haploid phase in Candida. The existence of a haploid requires that a single copy of a gene provide sufficient activity to carry out its function. However, this requirement would not apply to obligate diploid organisms such as Candida, which has no known vegetative haploid stage. Thus, one interpretation of this data is that in Candida hyphal formation is sensitive to the dosage of the MAPK cascade components. The lack of dominance of the wild-type allele to the null allele has also been reported for the Candida drug transporter CaMDR and the Candida MKC1 gene, a homolog of the Saccharomyces MAPK SLT2/MPK1. The heterozygotes of null mutations of these genes are more sensitive to inhibitors than the wild type (23, 24).
This model, which predicts that +/+ > +/m > m/m, agrees reasonably well with the observed phenotypes of strains with these genetic constitutions. However, the transformants +/m/m are overall more filamentous than +/m, which does not fit the dosage model. An alternative possibility is that the partial dominance of the null mutation in the heterozygote is a reflection of a more exotic mechanism such as defective pairing between the normal and the deleted allele (25). Whatever the mechanism, the manifestation of mutant phenotypes in the heterozygote may facilitate the identification and isolation of Candida mutants.
Acknowledgments
We thank William Fonzi for the kind gift of Candida strain CAI4. We thank Don Higgins, Ekkehard Leberer, and Yona Kassir for kindly providing plasmids. We thank Haoping Liu, Jonathan Loeb, Bonnie Bartel, Judith Bender, and other members of the Fink laboratory for many fruitful discussions and Andrew Diener, Peter Hecht, Hiten Madhani, Rad Roberts, Steffen Rupp, and Eric Summers for helpful comments on the manuscript. J.R.K. thanks Ken McIntosh for support and encouragement. This work was supported by National Institutes of Health Grant GM40266 to G.R.F. J.R.K. was supported by National Research Service Award F32A109236. G.R.F. is an American Cancer Society Professor of Genetics.
Footnotes
References
- 1.Cutler J E. Annu Rev Microbiol. 1991;45:187–218. doi: 10.1146/annurev.mi.45.100191.001155. [DOI] [PubMed] [Google Scholar]
- 2.Birse C E, Irwin M Y, Fonzi W A, Sypherd P S. Infect Immun. 1993;61:3648–3655. doi: 10.1128/iai.61.9.3648-3655.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Saporito-Irwin S M, Birse C E, Sypherd P S, Fonzi W A. Mol Cell Biol. 1995;15:601–613. doi: 10.1128/mcb.15.2.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Liu H, Styles C A, Fink G R. Science. 1993;262:1741–1744. doi: 10.1126/science.8259520. [DOI] [PubMed] [Google Scholar]
- 5.Liu H, Köhler J, Fink G R. Science. 1994;266:1723–1726. doi: 10.1126/science.7992058. [DOI] [PubMed] [Google Scholar]
- 6.Malathi K, Ganesan K, Datta A. J Biol Chem. 1994;269:22945–22951. [PubMed] [Google Scholar]
- 7.Gietz D, Jean A S, Woods R A, Schiestl R H. Nucleic Acids Res. 1992;20:1425. doi: 10.1093/nar/20.6.1425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fonzi W A, Irwin M Y. Genetics. 1993;134:717–728. doi: 10.1093/genetics/134.3.717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cannon R D, Jenkinson H F, Shepherd M G. Mol Gen Genet. 1992;235:453–457. doi: 10.1007/BF00279393. [DOI] [PubMed] [Google Scholar]
- 10.Brill J A, Elion E A, Fink G R. Mol Biol Cell. 1994;5:297–312. doi: 10.1091/mbc.5.3.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Leberer E, Dignard D, Harcus D, Thomas D Y, Whiteway M. EMBO J. 1992;11:4815–4824. doi: 10.1002/j.1460-2075.1992.tb05587.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Trueheart J, Boeke J D, Fink G R. Mol Cell Biol. 1987;7:2316–2328. doi: 10.1128/mcb.7.7.2316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gimeno C J, Ljungdahl P O, Styles C A, Fink G R. Cell. 1992;68:1077–1090. doi: 10.1016/0092-8674(92)90079-r. [DOI] [PubMed] [Google Scholar]
- 14.Alani E, Cao L, Kleckner N. Genetics. 1987;116:541–545. doi: 10.1534/genetics.112.541.test. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gillum A M, Tsay E Y, Kirsch D R. Mol Gen Genet. 1984;198:179–182. doi: 10.1007/BF00328721. [DOI] [PubMed] [Google Scholar]
- 16.Lee K L, Buckley H R, Campbell C C. Sabouraudia. 1975;13:148–153. doi: 10.1080/00362177585190271. [DOI] [PubMed] [Google Scholar]
- 17.Clark K L, Feldman P J, Dignard D, Larocque R, Brown A J, Lee M G, Thomas D Y, Whiteway M. Mol Gen Genet. 1995;249:609–621. doi: 10.1007/BF00418030. [DOI] [PubMed] [Google Scholar]
- 18.Ramer S W, Davis R W. Proc Natl Acad Sci USA. 1993;90:452–456. doi: 10.1073/pnas.90.2.452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Santos M A, Tuite M F. Nucleic Acids Res. 1995;23:1481–1486. doi: 10.1093/nar/23.9.1481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ottilie S, Miller P J, Johnson D I, Creasy C L, Sells M A, Bagrodia S H, Forsburg S L, Chernoff J. EMBO J. 1995;14:5908–5919. doi: 10.1002/j.1460-2075.1995.tb00278.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Marcus S, Polverino A, Chang E, Robbins D, Cobb M H, Wigler M H. Proc Natl Acad Sci USA. 1995;92:6180–6184. doi: 10.1073/pnas.92.13.6180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Manser E, Leung T, Salihuddin H, Zhao Z S, Lim L. Nature (London) 1994;367:40–46. doi: 10.1038/367040a0. [DOI] [PubMed] [Google Scholar]
- 23.Goldway M, Teff D, Schmidt R, Oppenheim A B, Koltin Y. Antimicrob Agents Chemother. 1995;39:422– 426. doi: 10.1128/aac.39.2.422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Navarro-Garcia F, Sanchez M, Pla J, Nombela C. Mol Cell Biol. 1995;15:2197–2206. doi: 10.1128/mcb.15.4.2197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Aramayo R, Metzenberg R L. Cell. 1996;86:103–113. doi: 10.1016/s0092-8674(00)80081-1. [DOI] [PubMed] [Google Scholar]