

Abstract
A K+ channel gene has been cloned from Drosophila melanogaster by complementation in Saccharomyces cerevisiae cells defective for K+ uptake. Naturally expressed in the neuromuscular tissues of adult flies, this gene confers K+ transport capacity on yeast cells when heterologously expressed. In Xenopus laevis oocytes, expression yields an ungated K+-selective current whose attributes resemble the “leak” conductance thought to mediate the resting potential of vertebrate myelinated neurons but whose molecular nature has long remained elusive. The predicted protein has two pore (P) domains and four membrane-spanning helices and is a member of a newly recognized K+ channel family. Expression of the channel in flies and yeast cells makes feasible studies of structure and in vivo function using genetic approaches that are not possible in higher animals.
Keywords: K+ channel, two P domains, yeast, muscles, nerves
Potassium channels comprise a large group of integral membrane proteins that determine the electrical properties of neurons and muscle fibers (1). They assume this central role in cellular physiology through regulation of action potential duration, firing patterns, and resting membrane potential. Distinguished individually by how they open and close and the magnitude of their single channel conductances, K+ channel subunits are unified by a shared pore-forming motif, the P domain (2, 3). Molecular cloning has revealed genes in Drosophila melanogaster and mammals encoding a superfamily of closely related K+ channels whose subunits contain a single P domain and six or more membrane-spanning segments (4–10). Another K+ channel superfamily, the inward rectifiers, is characterized by a single P domain and just two transmembrane segments (11, 12). The first K+ channel possessing two P domains in one contiguous polypeptide, Tok1, was cloned from the budding yeast Saccharomyces cerevisiae and has eight predicted membrane-spanning α-helices (13). More recently, a second family of K+ channels with two P domains and just four predicted transmembrane segments was expressed (14), confirming predictions based on data base analysis (13, 15, 16) .
Here, we describe the expression cloning from D. melanogaster of an example of this later type of two P domain K+ channel with four predicted transmembrane segments. The gene for the channel is located on chromosome 1 and gains wide expression in the excitable tissues of adult flies. Heterologous expression of the gene in yeast cells defective in K+ transport restores K+ uptake ability and allows cells to grow in medium containing micromolar concentrations of potassium. Gene expression in Xenopus laevis oocytes bathed in physiological solutions yields K+-selective currents that are instantaneous with changes in voltage and are outwardly rectifying. However, increased external K+ concentration ([K]o) significantly alters this current–voltage relationship; as [K]o rises, rectification is lost and K+ ions flow inward at hyperpolarized potentials. This sensitivity to [K]o appears intrinsic to the channel, since it approximates expectations from constant field theory for simple electrodiffusion through an open K+-selective pore (17, 18). That the channel directly underlies all these events is supported by the observation that K+ uptake, yeast cell growth, and oocyte currents are similarly sensitive to barium and resistant to tetraethylammonium (TEA).
While “leak conductances” have long been thought to be basic to neural function (19, 20), efforts to characterize them in dissected nerves have yielded inconsistent results (1). In function, the channel we describe here resembles the rectifying “leak” conductance postulated to underlie the resting potential of myelinated nerves (21) and is, to our knowledge, the first cloned channel to exhibit such behavior. Study of the channel’s influence on fly physiology in wild-type and mutant form may serve to clarify the role of leak conductances in neural function. Investigation of the channel in yeast cells allows application of genetic methods to assess the molecular basis for its function as well as high-throughput screens for isolation of channel modulators. The new gene and its channel product have been named ORK1, as an abbreviation for open rectifier K+ channel.
MATERIALS AND METHODS
Molecular Biology.
Cloning of ORK1 by complementation. Plasmid DNA was isolated from a λYES cDNA expression library made from third-instar larval mRNA (22). Library plasmids were introduced into S. cerevisiae strain CY162 by using the lithium acetate method, and 105 transformants were screened for complementation on low K+ as described (23). Plasmids isolated from colonies viable on low K+ were used to transform CY162 to confirm their complementing properties. One positive plasmid contained a complementing 2.4-kb XhoI fragment which was transferred to the XhoI sites of pYES2 (Invitrogen), forming pORK1, and to a variant of pRS426 (Stratagene) bearing the ADH1 promotor (24), forming pRAD-ORK1. Yeast media and methods were conventional; SC, complete synthetic medium, uracil-deficient with 3% galactose and ≤1 mM KCl; RPD, arginine phosphate dextrose medium, uracil-deficient and ≤1 mM KCl (25). S. cerevisiae strain CY162 (ura3 trk1 trk2) and the plasmid pKAT1 were obtained from Rick Gaber and have been described (23).
DNA sequence analysis.
Automated DNA sequence analysis was performed with Prism DNA sequencing kits (Applied Biosystems). Sequences were analyzed by using GeneWorks software (Intelligenetics). Quality scores were generated by the Gap Program of the Wisconsin Package (Version 8), Genetics Computer Group (26). A combined transcription–translation protocol (Promega) with [35S]methionine (NEN) was used. SDS/polyacrylamide gels were impregnated with soluble fluor (Amplify; NEN), and exposed to x-ray film (XAR-5, Kodak) after drying.
Rubidium uptake assay.
CY162 bearing pRAD-ORK1 or pRAD were grown overnight in RPD medium containing 100 mM KCl and no uracil, washed in distilled water, incubated in 10 mM calcium Hepes buffer (pH 6.8) for 1 hr, washed, and resuspended in buffer at 2 × 109 cells per ml. A 50-μl aliquot of the yeast suspension was mixed with 50 μl of buffer containing 5 kBq of 86RbCl (Amersham) and the indicated nonradioactive chloride salt. After 45 min, cells were concentrated on a 0.45-μm-pore HVLP Millipore filter and washed, and radioactivity was measured in a liquid scintillation counter. Rubidium uptake rates remained linear for over 60 min in calcium Hepes buffer containing 50 μM RbCl (not shown). Uptake by cells expressing pYES2-KAT1 was roughly 2-fold more efficient than in those expressing pRAD-ORK1 in buffer containing 50 μM RbCl (not shown).
Synthesis of ORK1 cRNA for microinjection.
Synthetic DNA duplexes were used to ligate the DNA sequence encoding ORK1 into the NcoI and BspeI sites of pSD (27), creating pSD-ORK1. Complementary RNA synthesis was catalyzed by T7 RNA polymerase in vitro using pSD-ORK1 as template after linearization with NotI. Transcript concentration was estimated spectrophotometrically and aliquots were stored at −80°C.
Electrophysiology.
Expression of ORK1 in X. laevis oocytes. Oocytes were isolated from X. laevis (Nasco, Atkinson, WI), defolliculated by collagenase treatment, and injected the next day with 46 nl of cRNA solution containing 1–5 ng of transcript. Whole cell currents were measured 1–3 days after cRNA injection by two-electrode voltage clamp using an Oocyte Clamp (Warner). Data were filtered at 1 kHz and sampled at 4 kHz. Electrodes contained 3 M KCl and had resistances of 0.3–1 MΩ. Recordings were performed under constant perfusion at room temperature. Bath solutions were 5 mM KCl solution (in mM: 95 NaCl, 5 KCl, 1 MgCl2, 0.3 CaCl2, 5 Hepes, pH 7.6); or 10, 20, 50, and 100 mM KCl solutions in which KCl is isotonically substituted for NaCl. For aspartate substitution, KCl and NaCl were replaced with potassium aspartate and sodium aspartate, respectively; for NMG substitution, NaCl was replaced with N-methylglucamine chloride.
RESULTS
Molecular Cloning of ORK1 by Functional Expression in S. cerevisiae.
In an effort to clone novel K+ channels, we employed a yeast complementation method used previously to isolate channels and transporters from plants (23, 28, 29). A yeast mutant, CY162, lacking high-affinity K+ uptake (23) was transformed with a D. melanogaster cDNA expression library made from third-instar larval mRNA (22). A single plasmid clone conferred upon these cells the ability to grow on low-K+ medium. The growth of yeast cells bearing this plasmid was comparable to that of control strains containing the inwardly-rectifying K+ channel of the plant Arabidopsis thaliana, KAT1 (23), and was much greater than that of CY162 cells containing vector alone (Fig. 1). Subcloning identified a 2422-bp XhoI fragment that conferred growth on low-K+ medium.
Figure 1.

Molecular cloning of ORK1 by complementation. ORK1 expression allows growth of yeast cells deficient in K+ transport; CY162 yeast cells were transformed with the designated plasmids, streaked on plates of uracil-deficient synthetic complete medium (SC), or SC containing 100 mM KCl and incubated for 40 hr at 30°C.
DNA sequencing of the 2422-bp fragment revealed a single long open reading frame (ORF) encoding a predicted protein of 618 amino acids (Fig. 2a). The start codon appears to be the first in-frame ATG, within the strong translation initiation sequence AGCATGT (31, 32). Within the proposed 3′ untranslated region is a consensus sequence for polyadenylylation, AATCAA (33), 122 bases downstream of the translational stop sequence. Hydrophobicity analysis of the predicted protein shows four stretches that could span the membrane in α-helical conformation (M1–M4) (Fig. 2b). Two motifs bearing close resemblance to the P domains of known K+ channels are in the ORF (Fig. 2c); the first (P1) is flanked by hydrophobic segments M1 and M2, the second (P2), by M3 and M4. The predicted mass of the protein (≈68 kDa) and a consensus site for N-glycosylation are consistent with products observed upon translation of ORK1 cRNA in vitro with and without microsomes (Fig. 2d). In the absence of a signal sequence, these data suggest a membrane topology in which the N and C termini are inside the cell and residue 58 (between M1 and M2) is glycosylated and thus external (Fig. 2e).
Figure 2.
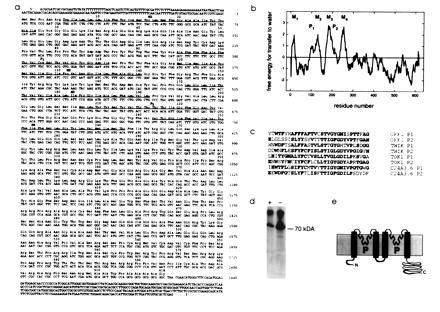
Nucleotide and deduced amino acid sequence of ORK1. (a) The 2.4-kb cDNA insert encoding ORK1 contains a single long ORF. Segments corresponding to putative pore-forming P domains and transmembrane segments (M1–M4) are underlined. The National Center for Biotechnology Information accession number for the nucleotide and amino acid sequence of ORK1 is U55321U55321. (b) Kyte–Doolittle hydrophilicity analysis of the ORK1 ORF with a window of 20 residues. (c) Alignment of the P domains of ORK1, Tok1, TWIK, and C24A3.6, a two P domain ORF from the Caenorhabditis elegans sequence data base (residues identical in 2 or more sequences are in boldface type); the probability that 8 random sequences would have the amino acid identities shown here is less than 10−50 as calculated by the method of Jan and Jan (30). While the overall similarity of ORK1 and Tok1 is low, the region extending from the first P domain to 20 residues past the second yields a 56% similarity and 23% identity with an quality score of 79 by the method of Needleman and Wunsch (26). Sequential randomizations and alignments of this region produce an average quality score of only 51 ± 3, suggesting that the channels share a single common ancestor; sequence data from other species are needed to establish this thesis. ORK1 and TWIK (14) show an overall amino acid identity of 21% and similarity of 48% with a quality score of 214. (d) In vitro translation of ORK1 with (+) or without (−) dog pancreatic microsomes resolved in an SDS/12.5% polyacrylamide gel and visualized by autoradiography. (e) Predicted membrane topology of ORK1.
Northern blot analysis using the 2422-bp XhoI fragment as the probe reveals a 2.8-kb species of poly(A)+ mRNA from D. melanogaster adults (Fig. 3a) that is also present at low levels in larvae and embryos (not shown). In situ hybridization to polytene chromosomal preparations localizes ORK1 to band 1-10A1-2 (Fig. 3b); several lethal mutations map to this band (34). In situ RNA·RNA hybridization using a 750-bp antisense fragment reveals that expression of ORK1 mRNA is widespread, with particular prominence in muscle, brain, and ovary of adult flies (Fig. 3c).
Figure 3.
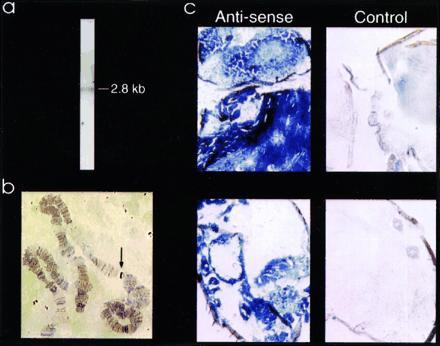
ORK1 is expressed in the excitable tissues of adult D. melanogaster. (a) Northern blot analysis of ORK1 mRNA. D. melanogaster mRNA (5 μg, adult, Clontech) was resolved in a formaldehyde/agarose gel, blotted to nitrocellulose, and probed with 32P-labeled 2.4-kb ORK1 XhoI fragment overnight at 65°C. The blot was exposed to x-ray film for 46 hr at −70°C with an intensifying screen. (b) In situ hybridization to polytene chromosomes identifies a band on 1–10A1-2 by the method of Todd Laverty using biotin-labeled probes produced by nick-translation and peroxidase-based visualization (Enzo Diagnostics, New York). (c) In situ hybridization in whole adult fly preparations reveals expression of ORK1 mRNA in excitable tissues in the head and thorax (Upper) and abdomen (Lower) with antisense but not control cRNA. Digoxigenin-labeled antisense RNA (to the terminal 300 nucleotides of the coding region and 400 nucleotides of the 3′ untranslated region) or a kit-supplied control RNA were hybridized to 18-μm sections of quick-frozen, OCT-embedded, D. melanogaster CS overnight at 65°C and visualized by an alkaline phosphatase-catalyzed reaction (Boehringer Mannheim).
ORK1 Mediates Barium-Sensitive Uptake of K+ When Expressed in Yeast Cells.
CY162 yeast cells grow poorly on low-K+ medium unless they express an exogenous protein that mediates K+ transport, such as the plant K+ channel KAT1 (Fig. 1) (23). Consistent with the idea that ORK1 is also an ion channel, CY162 cells constitutively expressing ORK1 grow on low-K+ medium (RPD containing 1 mM KCl) and their growth is inhibited by the common K+ channel blocker barium (Fig. 4a). Cell growth resulting from ORK1 expression differs in its pharmacology from that based on KAT1—only cells employing the latter are markedly sensitive to TEA and cesium (Fig. 4a). This difference suggests that the two proteins facilitate transport directly rather than by activating an endogenous transport protein.
Figure 4.
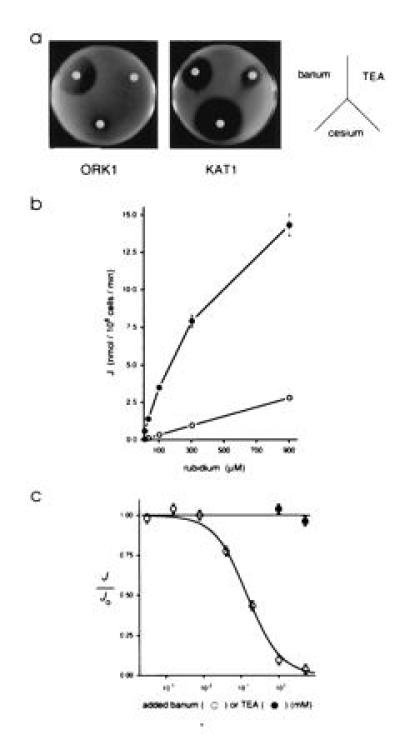
ORK1-dependent yeast cell growth and K+ uptake are inhibited barium. (a) Inhibition of ORK1 or KAT1-dependent yeast growth by three common K+ channel blockers. CY162 cells (105) expressing the designated channels were plated in RPD medium with 1 mM KCl. Sterile filter disks were placed on the surface of the agar and saturated with 20 μl of solutions containing chloride salts of barium (5 μmol), TEA (20 μmol), or cesium (2 μmol). The plates were incubated for 36 hr at 30°C. (b) Uptake of rubidium by CY162 cells carrying pRAD-ORK1 (•) or pRAD (○) at various concentrations of unlabeled rubidium chloride. (c) Inhibition of rubidium uptake by barium (○) or TEA (•) at 50 μM rubidium chloride. The solid curve is fitted to the data according to (1 + [blocker]/Ki)−1, where Ki is the equilibrium dissociation constant; under these conditions 0.15 mM external barium decreases flux ≈50%.
To directly assess ORK1-mediated ion flux into yeast cells, uptake of the K+ congener 86Rb+ was evaluated. Constitutive expression of ORK1 in CY162 cells results in 86Rb+ uptake rates comparable to those observed with KAT1 and much higher than those seen with vector alone (Fig. 4b). ORK1-mediated uptake of 86Rb+ is concentration dependent and, like cell growth, is sensitive to barium but not TEA (Fig. 4c).
ORK1 Mediates K+-Selective Currents When Expressed in X. laevis Oocytes.
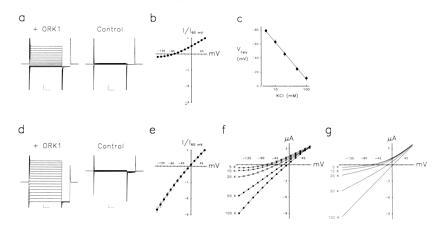
When 1 ng of ORK1 cRNA is injected into X. laevis oocytes a current is observed by two-electrode voltage clamp that is not present in control oocytes (Fig. 5a). In response to changes in transmembrane voltage, ORK1 currents rise to a maximum level with no apparent delay. Currents are steady with maintained voltage and show no evidence for inactivation. When external K+ is in the physiologic range (i.e., 5 mM KCl solution), currents are outwardly rectifying (Fig. 5b). Under these conditions, inward currents are seen but remain small even at very hyperpolarized voltages.
Figure 5.
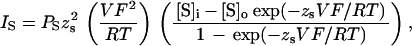 |
ORK1 currents result from movement of K+ (Fig. 5c). Oocytes expressing ORK1 and studied in solutions where KCl content is altered by isotonic substitution for NaCl exhibit reversal potentials in nearly perfect agreement with values predicted by the Nernst equation. This indicates that ORK1 currents are highly selective for K+ over both Na+ and Cl−. Indeed, substitution of aspartate for Cl− in the bath solution has no significant effect on currents (not shown).
ORK1 Current Rectification in Oocytes Is Altered by External K+ Concentration ([K]o).
The outwardly rectifying character of ORK1 currents changes in response to elevation of external K+ concentration (Fig. 5 d and e). Raising KCl by isotonic replacement for NaCl leads to large inward currents at hyperpolarized voltages that were previously electrically quiet as well as stable inward currents at the holding voltage (−80 mV). Current–voltage relationships in a variety of KCl solutions are shown for a representative oocyte expressing ORK1 (Fig. 5f). Increasing external KCl concentration leads to loss of rectification and altered reversal potential. These effects appear to be a direct reflection of the transmembrane concentration gradient of K+: such rectifying current–voltage relations conform to expectations of the Goldman–Hodgkin–Katz current relation in the case of unequal permeant ion concentrations (17, 18) (Fig. 5g), while shifts in reversal potential with [K]o were shown above to be Nernstian in character (Fig. 5c). These effects do not result from voltage-dependent Na+ blockade (and thus relief of block by substitution of KCl for NaCl), since substitution of N-methylglucamine for Na+ has no significant effect on ORK1 currents (not shown).
ORK1 Currents in Oocytes Are Barium Sensitive.
As predicted from growth inhibition assays (Fig. 4a) as well as 86Rb+ flux experiments (Fig. 4c), ORK1 currents are blocked by barium but not by TEA (Fig. 6a and b). Barium block is voltage dependent and does not change the instantaneous character of current development or lead to inactivation (Fig. 6c). This suggests that barium inhibits ORK1, as it does several well-characterized K+ channels (37), by direct, reversible, open-channel pore blockade. If barium does block by this mechanism, its binding energy will reflect the fraction of the electric field traversed to reach its pore site (36, 38); a determination for zδ of 0.80 ± 0.02 suggests that barium ions move through ≈40% of the potential drop across the membrane to bind (Fig. 6d).
Figure 6.
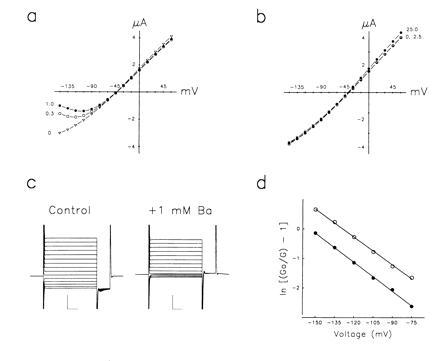
ORK1 currents in oocytes are blocked by barium but not TEA. (a) Current–voltage relationship with constant perfusion of 20 mM KCl solution containing 0 mM (▿), 0.3 mM (○), or 1 mM (•) barium chloride by the protocol in Fig. 4a (mean ± SEM, n = 3 cells). (b) Current–voltage relationship with constant perfusion of 20 mM KCl solution with 0 mM (▿), 2.5 mM (○), or 25 mM (•) TEA chloride isotonically substituted for NaCl (mean ± SEM, n = 3 cells). (c) ORK1-induced currents in an oocyte perfused with 20 mM KCl solution in the absence (control) or presence of 1 mM barium chloride by the protocol in Fig. 4a. Scale bars represent 2 μA and 15 ms. (d) Voltage dependence of barium block was assessed by measurements of conductance from −150 mV to −75 mV in the presence of 0.3 mM (○) or 1 mM (•) barium chloride by the protocol in Fig. 4a. The data shown are from one oocyte but are representative. Go and G are the cord conductances measured in the absence and presence of barium, respectively. The electrical distance was calculated from Ki(V) = Ki(0)exp(zδFV/RT), where Ki(0) is the zero voltage inhibition constant, z is the valence of the blocking ion, and δ is the fraction of the applied voltage drop experienced at the binding site (36).
DISCUSSION
The gene for a unique K+ channel containing two P domains in tandem has been cloned from D. melanogaster. Cloning was achieved by complementation using a mutant strain of S. cerevisiae defective in K+ transport (39) and a cDNA expression library produced from fly larvae (22). Channel expression in yeast cells confers uptake of K+ and allows cell growth in low-K+ medium; in X. laevis oocytes, expression yields instantaneous, K+-selective, and noninactivating currents whose rectification is altered by [K]o. The gene and its channel have been named ORK1.
Two motifs resembling the pore-forming P domains of known K+ channels are found in ORK1 channels (Fig. 2a). In single P domain K+ channels, a “signature sequence” within the P domain (TxxTxGxG, with lowercase letters indicating variable amino acid residues) is critical for K+ selectivity (2). The P1 sequence of ORK1 provides a variant of this sequence (TvcsTvGyG), while the P2 aligns well (TttTiGfG). As in other cloned K+ channels, each P domain is flanked by stretches of 20 or more hydrophobic amino acids (Fig. 2b, M1–M4) and ORK1 subunits are predicted to have four membrane-spanning segments (Fig. 2e). Because single P domain K+ channel subunits associate as tetramers to form functional complexes (40–42), it seems likely ORK1 channels, with two P domain subunits, will function as dimers. The hydrophilic C-terminal domain of ORK1 contains multiple consensus sites for protein kinase A and C phosphorylation and a region (residues 551–581) with strong similarity to the third intracellular loop of the somatostatin receptor subtype 3 that is thought to contribute to interaction with heterotrimeric G proteins (43). This suggests a regulatory function for this domain.
ORK1 shares its predicted two P domain, four membrane-spanning segment (2P/4TM) topology with TWIK, the product of a gene recently identified in the human expressed sequence tag data base using the blast alignment program (14); like ORK1, TWIK is predicted to influence membrane potential, since it yields a K+-selective pore with extremely weak inward rectification. This 2P/4TM topology is predicted for over a dozen ORFs in the C. elegans genome that may be K+ channels as well (15, 16). Characterization of additional 2P/4TM channels will be required to determine if they all produce open K+-selective channels and thus merit classification as a family based on function rather than their unique structure. While it is not yet possible to infer the evolutionary antecedents or progeny of the 2P/4TM channels, it is apparent that the P and near P segments of ORK1 are related to the human and C. elegans gene products as well as to Tok1 of S. cerevisiae, the founding member of the other two P domain K+ channel family, which has a proposed 2P/8TM topology (Fig. 2c). These alignments support the contention that a common ancestor of two P domain K+ channels was present in eukaryotes before differentiation of fungi and animals in the late Precambrian period (44). It is premature to assess the relationship between two P domain K+ channels and Na+ and Ca2+ channels, whose subunits carry four P domains in one contiguous polypeptide, which are thought to have arisen through gene duplication via two P domain channel intermediates (45).
The effects of extracellular K+ on ORK1 appear to result directly from unequal permeant ion concentrations across the membrane, as expected from constant field theory for an open K+ channel (17, 18). This type of instantaneous rectification through open K+ (and Na+) channels has been recognized in nerves and muscles previously (20, 46, 47) and differs from other known effects of [K]o on K+ channel gating (48–53) and blockade (54–57). The effects of [K]o on ORK1 are also distinct from those observed with Tok1, the archetype of the other two P domain K+ channel family. Tok1 allows only small inward currents in intact cells, even when extracellular K+ is high, but it does alter the voltage at which it passes outward currents in response to changes in [K]o (13). In contrast, ORK1 passes large inward K+ currents in elevated [K]o. Tok1 is thus categorized as an “outward rectifier” (13), in analogy to the inward rectifier K+ channels that shift the voltage at which they pass inward currents in response to changes in [K]o, and ORK1 is classified as an “open rectifier.” Of note, K+ conductances that are sensitive to [K]o have a recognized role in normal physiology (58) as well as the pathogenesis of epilepsy and postischemic cardiac arrhythmias (59, 60) but the channels that mediate these effects are undefined.
The attributes of ORK1 currents, (K+ selectivity, [K]o-sensitive rectification, voltage independence, and TEA insensitivity), are similar to those identified with the “residual leak” conductance hypothesized to mediate the resting potential of myelinated nerves dissected from vertebrates and, thus, indirectly argues for the feasibility of that model (21). Study of ORK1 in flies appears to offer a powerful strategy to explore the role of “leak” conductances in nervous function (61). ORK1 might be expected to have two effects in the excitable tissues of D. melanogaster. First, ORK1 yields a K+-selective current across all voltages studied which will shift resting membrane potential toward the equilibrium reversal for K+ (EK). Second, rectification of ORK1 currents is [K]o dependent, and this will influence the magnitude and duration of action potential afterhyperpolarizations and firing rates. Since action potential repolarization results from outward flux of K+ leading to transient elevation in [K]o at the external face of the membrane (62, 63), it follows that the size of inward currents through ORK1 channels will exhibit [K]o-dependent hysteresis with each wave of excitation and, so, dynamically influence the kinetics of changes in membrane potential. These effects would affect (and may control) overall membrane excitability. The potential of a genetic approach in flies is underscored by observation that mutations in voltage-gated ion channels have proven a rich source of D. melanogaster behavioral mutants (4–6, 64).
Successful application of the complementation cloning method that isolated ORK1 has previously been restricted to channels and transporters from plants (23, 65). In general, plants and fungi maintain resting membrane potentials more negative than −100 mV. Under these conditions, most voltage-gated K+ channels from animals are closed. Functional expression of ORK1 in yeast cells may result, first, from its ability to pass inward currents at negative potentials and, second, from its similarity to Tok1, an endogenous yeast two P domain channel that is processed and translocated to the cell membrane (66). Expression of ORK1 in yeast makes possible genetic approaches to ion channel structure and function not possible in other expression modalities (25, 67).
Acknowledgments
The authors are indebted to Glen Doughty, Robert Mann, Spyros Artavanis-Tsakonas, and Tian Xu for assistance with in situ studies; to Claude Grignon for advice on 86Rb flux assays; to Randal Numann and Jon Hayashi for oocyte expression; and to Nirpal Dhanjal and Ann Marie Quinn for sequence analysis. We are grateful for technical contributions of Kwok-Keung Tai and Marianne Buck. S.A.N.G. is supported by the National Institutes of Health, the Donaghue Foundation, and a grant from American Home Products (Wyeth-Ayerst) to the Molecular Cardiology Program of the Boyer Center for Molecular Medicine.
Footnotes
References
- 1.Hille B. Ionic Channels of Excitable Membranes. Sunderland, MA: Sinauer; 1992. [Google Scholar]
- 2.Heginbotham L, Lu Z, Abramson T, MacKinnon R. Biophys J. 1994;66:1061–1067. doi: 10.1016/S0006-3495(94)80887-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.MacKinnon R. Neuron. 1995;14:889–892. doi: 10.1016/0896-6273(95)90327-5. [DOI] [PubMed] [Google Scholar]
- 4.Kamb A, Iverson L E, Tanouye M A. Cell. 1987;50:405–413. doi: 10.1016/0092-8674(87)90494-6. [DOI] [PubMed] [Google Scholar]
- 5.Papazian D M, Schwarz T L, Tempel B L, Jan Y N, Jan L Y. Science. 1987;237:749–753. doi: 10.1126/science.2441470. [DOI] [PubMed] [Google Scholar]
- 6.Pongs O, Kecskemethy N, Muller R, Krah-Jentgens I, Baumann A, Kiltz H H, Canal I, Llamazares S, Ferrus A. EMBO J. 1988;7:1087–1096. doi: 10.1002/j.1460-2075.1988.tb02917.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Butler A, Wei A G, Baker K, Salkoff L. Science. 1989;243:943–947. doi: 10.1126/science.2493160. [DOI] [PubMed] [Google Scholar]
- 8.Wei A, Covarrubias M, Butler A, Baker K, Pak M, Salkoff L. Science. 1990;248:599–603. doi: 10.1126/science.2333511. [DOI] [PubMed] [Google Scholar]
- 9.Atkinson N S, Robertson G A, Ganetzky B. Science. 1991;253:551–555. doi: 10.1126/science.1857984. [DOI] [PubMed] [Google Scholar]
- 10.Bruggemann A, Pardo L A, Stuhmer W, Pongs O. Nature (London) 1993;365:445–448. doi: 10.1038/365445a0. [DOI] [PubMed] [Google Scholar]
- 11.Kubo Y, Reuveny E, Slesinger P A, Jan Y N, Jan L Y. Nature (London) 1993;364:802–806. doi: 10.1038/364802a0. [DOI] [PubMed] [Google Scholar]
- 12.Krapivinsky G, Gordon E A, Wickman K, Velimirovic B, Krapivinsky L, Clapham D E. Nature (London) 1995;374:135–141. doi: 10.1038/374135a0. [DOI] [PubMed] [Google Scholar]
- 13.Ketchum K A, Joiner W J, Sellers A J, Kaczmarek L K, Goldstein S A N. Nature (London) 1995;376:690–695. doi: 10.1038/376690a0. [DOI] [PubMed] [Google Scholar]
- 14.Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G, Barhanin J. EMBO J. 1996;15:1004–1011. [PMC free article] [PubMed] [Google Scholar]
- 15.Wilson R, Ainscough R, Anderson K, Baynes C, Berks M, et al. Nature (London) 1994;368:32–38. doi: 10.1038/368032a0. [DOI] [PubMed] [Google Scholar]
- 16.Salkoff L, Jegla T. Neuron. 1995;15:489–492. doi: 10.1016/0896-6273(95)90137-x. [DOI] [PubMed] [Google Scholar]
- 17.Goldman D E. J Gen Physiol. 1943;27:37–60. doi: 10.1085/jgp.27.1.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hodgkin A L, Katz B. J Physiol (London) 1949;108:37–77. doi: 10.1113/jphysiol.1949.sp004310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hodgkin A L, Huxely A F. J Physiol (London) 1952;117:500–544. doi: 10.1113/jphysiol.1952.sp004764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Frankenhaeuser B. J Physiol (London) 1963;169:424–430. doi: 10.1113/jphysiol.1963.sp007268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Baker M, Bostock H, Grafe P, Martius P. J Physiol (London) 1987;383:45–67. doi: 10.1113/jphysiol.1987.sp016395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Elledge S J, Mulligan J T, Ramer S W, Spottswood M, Davis R W. Proc Natl Acad Sci USA. 1991;88:1731–1735. doi: 10.1073/pnas.88.5.1731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Anderson J A, Huprikar S S, Kochian L V, Lucas W J, Gaber R F. Proc Natl Acad Sci USA. 1992;89:3736–3740. doi: 10.1073/pnas.89.9.3736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bennetzen J L, Hall B D. J Biol Chem. 1982;257:3018–3025. [PubMed] [Google Scholar]
- 25.Guthrie C, Fink G R. Guide to Yeast Genetics and Molecular Biology. New York: Academic; 1991. [Google Scholar]
- 26.Needleman S B, Wunsch C D. J Mol Biol. 1970;48:443–453. doi: 10.1016/0022-2836(70)90057-4. [DOI] [PubMed] [Google Scholar]
- 27.Wang K W, Goldstein S A N. Neuron. 1995;14:1303–1309. doi: 10.1016/0896-6273(95)90277-5. [DOI] [PubMed] [Google Scholar]
- 28.Sentenac H, Bonneaud N, Minet M, Lacroute F, Salmon J M, Gaymard F, Grignon C. Science. 1992;256:663–665. doi: 10.1126/science.1585180. [DOI] [PubMed] [Google Scholar]
- 29.Schachtman D P, Schroeder J I. Nature (London) 1994;370:655–658. doi: 10.1038/370655a0. [DOI] [PubMed] [Google Scholar]
- 30.Jan L Y, Jan Y N. Nature (London) 1994;371:119–122. doi: 10.1038/371119a0. [DOI] [PubMed] [Google Scholar]
- 31.Kozak M. J Cell Biol. 1991;115:887–903. doi: 10.1083/jcb.115.4.887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cavener D R, Ray S C. Nucleic Acids Res. 1991;19:3185–3192. doi: 10.1093/nar/19.12.3185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Manley J L. Biochim Biophys Acta. 1988;950:1–12. doi: 10.1016/0167-4781(88)90067-x. [DOI] [PubMed] [Google Scholar]
- 34.Lindsley D L, Zimm G G. The Genome of Drosophila melanogaster. San Diego: Academic; 1992. [Google Scholar]
- 35.Wang K-W, Tai K-K, Goldstein S A N. Neuron. 1996;16:571–577. doi: 10.1016/s0896-6273(00)80076-8. [DOI] [PubMed] [Google Scholar]
- 36.Woodhull A M. J Gen Physiol. 1973;61:687–708. doi: 10.1085/jgp.61.6.687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Neyton J, Miller C. J Gen Physiol. 1988;92:569–586. doi: 10.1085/jgp.92.5.569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Coronado R, Miller C. J Gen Physiol. 1982;79:529–547. doi: 10.1085/jgp.79.4.529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ko C H, Gaber R F. Mol Cell Biol. 1991;11:4266–4273. doi: 10.1128/mcb.11.8.4266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.MacKinnon R. Nature (London) 1991;350:232–235. doi: 10.1038/350232a0. [DOI] [PubMed] [Google Scholar]
- 41.Glowatzki E, Fakler G, Brandle U, Rexhausen U, Zenner H P, Ruppersberg J P, Fakler B. Proc R Soc London B. 1995;261:251–261. doi: 10.1098/rspb.1995.0145. [DOI] [PubMed] [Google Scholar]
- 42.Yang J, Jan Y N, Jan L Y. Neuron. 1995;15:1441–1447. doi: 10.1016/0896-6273(95)90021-7. [DOI] [PubMed] [Google Scholar]
- 43.Yasuda K, Rens-Domiano S, Breder C D, Law S F, Saper C B, Reisine T, Bell G I. J Biol Chem. 1992;267:20422–20428. [PubMed] [Google Scholar]
- 44.Knoll A H. Science. 1992;256:622–627. doi: 10.1126/science.1585174. [DOI] [PubMed] [Google Scholar]
- 45.Strong M, Chandy K G, Gutman G A. Mol Biol Evol. 1993;10:221–242. doi: 10.1093/oxfordjournals.molbev.a039986. [DOI] [PubMed] [Google Scholar]
- 46.Campbell D T, Hille B. J Gen Physiol. 1976;67:309–323. doi: 10.1085/jgp.67.3.309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chui S Y, Ritchie J M, Rogart R B, Stagg D. J Physiol (Lond) 1979;292:149–166. doi: 10.1113/jphysiol.1979.sp012843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Swenson R P, Jr, Armstrong C M. Nature (London) 1981;291:427–429. doi: 10.1038/291427a0. [DOI] [PubMed] [Google Scholar]
- 49.Shapiro M S, DeCoursey T E. J Gen Physiol. 1991;97:1251–1278. doi: 10.1085/jgp.97.6.1251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Pardo L A, Heinemann S H, Terlau H, Ludewig U, Lorra C, Pongs O, Stuhmer W. Proc Natl Acad Sci USA. 1992;89:2466–2470. doi: 10.1073/pnas.89.6.2466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lopez B J, Hoshi T, Heinemann S H, Aldrich R W. Recept Channels. 1993;1:61–71. [PubMed] [Google Scholar]
- 52.Silver M R, Shapiro M S, DeCoursey T E. J Gen Physiol. 1994;103:519–548. doi: 10.1085/jgp.103.4.519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Baukrowitz T, Yellen G. Neuron. 1995;15:951–960. doi: 10.1016/0896-6273(95)90185-x. [DOI] [PubMed] [Google Scholar]
- 54.Vandenberg C A. Proc Natl Acad Sci USA. 1987;84:2560–2564. doi: 10.1073/pnas.84.8.2560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lu Z, MacKinnon R. Nature (London) 1994;371:243–246. doi: 10.1038/371243a0. [DOI] [PubMed] [Google Scholar]
- 56.Lopatin A N, Makhina E N, Nichols C G. Nature (London) 1994;372:366–369. doi: 10.1038/372366a0. [DOI] [PubMed] [Google Scholar]
- 57.Fakler B, Brandle U, Glowatzki E, Weidmann S, Zenner H P, Ruppersberg J P. Cell. 1995;80:149–154. doi: 10.1016/0092-8674(95)90459-x. [DOI] [PubMed] [Google Scholar]
- 58.Carmeliet, E. (1989) Pflügers Arch. 414, Suppl., S88–S92. [DOI] [PubMed]
- 59.Fozzard H A, Makielski J C. Annu Rev Med. 1985;36:275–284. doi: 10.1146/annurev.me.36.020185.001423. [DOI] [PubMed] [Google Scholar]
- 60.Heinemann U, Konnerth A, Pumain R, Wadman W J. Adv Neurol. 1986;44:641–661. [PubMed] [Google Scholar]
- 61.Wolfner M F, Goldberg M L. In: Drosophila melanogaster: Practical Uses in Cell and Molecular Biology. Fyrberg E A, Goldstein L S B, editors. Vol. 44. San Diego: Academic; 1994. pp. 33–80. [Google Scholar]
- 62.Frankenhaeuser B, Hodgkin A L. J Physiol (London) 1956;131:341–376. doi: 10.1113/jphysiol.1956.sp005467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ritchie J M, Straub R W. J Physiol (London) 1957;136:80–97. doi: 10.1113/jphysiol.1957.sp005744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Loughney K, Kreber R, Ganetzky B. Cell. 1989;58:1143–1154. doi: 10.1016/0092-8674(89)90512-6. [DOI] [PubMed] [Google Scholar]
- 65.Schachtman D P, Schroeder J I. Nature (London) 1994;370:655–658. doi: 10.1038/370655a0. [DOI] [PubMed] [Google Scholar]
- 66.Zhou X L, Vaillant B, Loukin S H, Kung C, Saimi Y. FEBS Lett. 1995;373:170–176. doi: 10.1016/0014-5793(95)01035-d. [DOI] [PubMed] [Google Scholar]
- 67.Uozumi N, Gassmann W, Cao Y, Schroeder J I. J Biol Chem. 1995;270:24276–24281. doi: 10.1074/jbc.270.41.24276. [DOI] [PubMed] [Google Scholar]