

Abstract
Focally evoked calcium waves in astrocyte cultures have been thought to propagate by gap-junction-mediated intercellular passage of chemical signal(s). In contrast to this mechanism we observed isolated astrocytes, which had no physical contact with other astrocytes in the culture, participating in a calcium wave. This observation requires an extracellular route of astrocyte signaling. To directly test for extracellular signaling we made cell-free lanes 10–300 μm wide in confluent cultures by deleting astrocytes with a glass pipette. After 4–8 hr of recovery, regions of confluent astrocytes separated by lanes devoid of cells were easily located. Electrical stimulation was used to initiate calcium waves. Waves crossed narrow (<120 μm) cell-free lanes in 15 of 36 cases, but failed to cross lanes wider than 120 μm in eight of eight cases. The probability of crossing narrow lanes was not correlated with the distance from the stimulation site, suggesting that cells along the path of the calcium wave release the extracellular messenger(s). Calculated velocity across the acellular lanes was not significantly different from velocity through regions of confluent astrocytes. Focal superfusion altered both the extent and the direction of calcium waves in confluent regions. These data indicate that extracellular signals may play a role in astrocyte–astrocyte communication in situ.
Keywords: glia, astrocytes, calcium waves, extracellular signal
In recent years, widespread communication between astrocytes has been demonstrated. Most prominent has been the propagation of transient rises in cytosolic free Ca2+ or “calcium waves” (1–3). Calcium waves have been observed to travel for millimeters at velocities of about 0.1 mm/s, considerably slower than propagation of neuronal action potentials. In addition, it is now clear that activity in neurons can evoke calcium waves in astrocytes (4). Even more strikingly, astrocytes participating in calcium waves can directly excite calcium transients in neurons and neuronal circuits (5–7). The large domains of astrocytic activity evidenced by rises in calcium may affect central nervous system information processing both physiologically and pathologically. Several different mechanisms may contribute to the communication between astrocytes and between astrocytes and neurons. Glia and neurons can each initiate calcium rises in the other cell type by release of glutamate (5–7). Communication via gap junctions has also been suggested to mediate propagation of calcium waves from glia to neurons (6), and the use of gap junction blockers indicates a role for gap junctions in propagation of calcium waves between astrocytes as well (2).
Astrocytes are known to release and to respond to a number of bioactive compounds (1, 8, 9). The possibility that astrocytes may communicate with other astrocytes via an extracellular pathway has not been investigated, although extracellular signaling pathways have been implicated in several other nonneuronal cell types (10, 11), as well as neurons.
The present study reinvestigates the mechanism of astroyte–astrocyte signaling in the propagation of calcium waves, and demonstrates that an extracellular factor can diffuse across acellular areas and evoke calcium elevations in astrocytes physically isolated from the releasing cells. Furthermore, this extracellular signaling pathway is necessary for maximal propagation of astrocyte calcium waves in confluent cultures.
METHODS
Cell Culture.
Glia from postnatal day 1–5 mouse pups were obtained using the methods described for rat pups (5). Briefly, the cortical hemispheres were removed, cleaned, and dissociated by trypsinization (0.2%, GIBCO) and mechanical trituration. Cells were plated on poly-l-lysine coated glass coverslips. Cells were plated at 50,000 cells per cm2 and maintained in growth medium [MEM (GIBCO)/10% fetal calf serum (HyClone)/penicillin/streptomycin (GIBCO)/essential amino acids (GIBCO)/nonessential amino acids (GIBCO)] with 1–2 media changes per week. Cultures were used at 5–15 days in vitro.
Extracellular signaling was investigated using both confluent cultures and cultures in which artificial lanes were created. Cell-free lanes were created, in otherwise confluent cultures, using a sharp glass pipette or razor blade. The lanes varied in width from 10 to several hundred μm. These cultures were used after a 3- to 8-hr recovery period. No data were used from areas where astrocytes had regrown across the lane within 200 μm of the area of observation. Additionally, data from waves that gave any indication of crossing the lane outside of the area of observation were discarded.
For experiments employing glutamate antagonists, agents were added to observation saline (135 mM NaCl/5 mM KCl/1 mM MgCl2/3 mM CaCl2/10 mM Hepes/25 mM glucose, pH 7.25) at 2× final concentration; the antagonist was added in a 1:1 dilution with the observation saline in the culture dish. Two different antagonist solutions were used: the general glutamate receptor antagonist gamma-d-glutamyl-glycine (1 mM, Sigma) alone; or a mixture of the following: the N-methyl-d-aspartate receptor antagonist DL-2-amino-5-phosphonovaleric acid (100 μM, Sigma), the non-N-methyl-d-aspartate receptor antagonist 6,7-dinitroquinoxaline-2,3-dione (50 μM, Sigma), and the metabotropic receptor antagonist l-2-amino-3-phosphonopropionic acid (200 μM, Research Biochemicals, Natick, MA).
Astrocyte Calcium Wave Stimulation.
Waves of elevated glial calcium were evoked electrically by extracellular stimulation using a saline-filled glass pipette with a 5–10 μm bore. The electrode was placed within 10 μm of a target cell. Stimulation consisted of bipolar voltage pulses 2 msec in total duration, delivered at 17 Hz from a Grass Instruments (Quincy, MA) SD9 stimulator over a period of 3–8 sec. The adequate stimulus voltage for a given stimulation pipette was determined, at a test site remote from the experimental sites in the same culture dish, as the voltage that reproducibly evoked astrocytic calcium waves. This voltage was used for stimulation of all fields in that culture dish. With this method we obtained up to 10 successive responses from the same initiation site. The calcium wave was initiated during the stimulation period; a fixed stimulation period was used for each data set. Electrical stimulation seldom resulted in any observable damage to the stimulated cell. Only fields where the stimulated cell returned to near baseline fluorescence values were analyzed. For the data in this paper requiring multiple stimulation of the same field, a 10-min interval was maintained between successive stimuli in the same field to avoid any refractory period or desensitization from prior stimuli. For all other data, a given field was only stimulated once.
Superfusion of Specific Culture Areas.
Selected regions of cultures were superfused by using a strategically placed small bore glass pipette (10–20 μm). The perfusion pipette was always placed in the upper left corner of the field for perfusion experiments; stimulation occurred in the center of the visual field at a distance of 350 μm from the perfusion pipette. A microstream of observation medium was directed at stimulation sites and the strength was adjusted by raising and lowering the perfusion reservoir. In five experiments the perfusion was reversed and the pipette acted to pull medium from the culture dish. For these experiments, the pipette was placed within 50 μm of the electrical stimulation site.
Imaging and Data Analysis.
Intracellular calcium was monitored using the fluorescent indicator Fluo-3 AM (ref. 12; Molecular Probes). Cultures were loaded with 10 μM Fluo-3 AM for 50–60 min at 37°C. Cultures were then rinsed three times in observation saline solution and allowed to de-esterify 30 min before use. Fluorescent images (100 msec exposures) were acquired using a 10×, 0.5 numerical aperture or a 20×, 0.75 numerical aperture objective, and a Photometrics cooled CCD camera. A mercury light source, with a computer-controlled shutter and attenuated by 0.5 or 1.0 neutral density filters, was used for illumination. Fluorescence intensity was analyzed with iplab software (Signal Analytics, Vienna, VA) on a Macintosh Power PC computer. Experiments were performed at room temperature (21–24°C).
Sequences of images were acquired at 1- to 3-sec intervals. Control images (5–20 images) were collected prior to electrical stimulation. Changes in calcium concentrations within the astrocytes were estimated by changes in the fluorescence intensity relative to the control images. A positive astrocytic response was recorded if relative fluorescence intensity exceeded 1.15. Astrocytes participating in the calcium wave were identified on the basis of both temporal and spatial correlation with respect to both the stimulation site and other astrocytes participating in the calcium wave. For analysis of wave size, nuclei of astrocytes participating in the calcium wave were counted. For analysis of wave directional bias during local perfusion, a line was drawn on the image through the site of stimulation, perpendicular to the direction of the perfusion stream. Nuclei of participating astrocytes were counted separately on the side of this line toward the perfusion pipette and on the side away from the perfusion pipette.
Analysis of calcium wave propagation was performed on image series captured at 1-sec intervals. Distances were measured from the site of stimulation. The arrival of the wave front was defined as a rise of the relative fluorescent intensity above 1.15. The first three images following initiation of the calcium wave, which frequently included images taken during the stimulation period, were not included. The distance was least-squares fitted to an exponential equation of the form: A + B × (1 − exp(−t/τ)) that described the slowing of the calcium wave over time. In this equation, B represents the extrapolated maximal extent of the calcium wave, and A represents the distance at which measurements were begun.
RESULTS
These experiments were prompted by a series of observations using mechanical touch to activate a glial calcium wave, as employed by a number of investigators (3, 5–7). We noted that astrocytes on isolated islands—i.e., not in contact with the population of glial cells in which the wave was propagating—could also respond with a transient elevation in calcium. This observation suggested the presence of an extracellular signal, not previously described in astrocytic calcium waves.
Evoked Calcium Waves Propagate Across Cell-Free Regions.
To test the hypothesis that an extracellular signal participated in propagation of a calcium wave between astrocytes, we made two alterations in methods previously employed for studying such events. (i) We used electrical stimulation that enabled repeated activation of waves from the same initiation site. This method proved far more reproducible than touch stimulation. (ii) We made cell-free regions of defined widths between regions of confluent astrocytes. These lanes were made by drawing a fine micropipette or razor blade through a confluent plate of astrocytes and then allowing 3–8 hr for recovery. At the time of experimentation, the cell-free lanes were bordered by healthy cells, which loaded with fluo-3 and displayed normal baseline fluorescence values as compared with controls. Careful phase contrast and fluorescence microscopy examination was used to ensure that there were no cellular processes spanning the lanes employed in this study. These two changes in technique have allowed us to demonstrate the existence of an extracellular signal.
Fig. 1 illustrates an electrically evoked calcium wave as it approached and encountered an cell-free lane between regions of confluent cells. It also illustrates that the wave continued across the lane into astrocytes that were physically isolated from the initially stimulated group. There are many variations on the responses observed in these experiments, ranging from no response of the astrocytes on the other side of the lane to a local response involving several astrocytes, to a response propagating vigorously out of the camera field of view.
Figure 1.

An electrically-evoked astrocytic calcium wave traversing a cell-free lane. (A) The phase-contrast image of a bed of astrocytes in which a cell-free lane had been created 6 hr earlier. Also in view is the extracellular stimulating electrode ≈10 μm above the astrocyte upper surface. The red square delineates the field of fluorescent imaging of fluo-3 signals. (B) Cellular outlines showing the astrocytic bed, the cell-free lane, and all of the astrocytes participating in the calcium wave during the observation time depicted on the left. (C) Pseudocolor images of the change in fluorescent signal (F/Fo) in the astrocytes at selected time points. The images were collected at (from the top): 3, 16, 22, and 36 sec after stimulation. Note that in the third frame from the top, the first astrocytes on the distal side of the calcium wave show a response, which subsequently spreads to additional astrocytes. While the magnitude of spread, amplitude of response, and number of participating cells can vary in each experiment, most preparations with lanes of less than 100 μm show responses similar to that shown in this figure. (Bar = 50 μm.)
An astrocytic calcium wave traveling in a confluent monolayer of astrocytes can display varying behaviors. Waves can travel in circuitous routes, skip over individual cells while continuing to travel through adjacent astrocytes, pause between cells, or travel with changing velocity. The representative curves in Fig. 2A demonstrate wave propagation in confluent glial cultures. Waves generally slowed over time. Fig. 2B shows three representative curves for calcium waves that encountered and crossed experimentally produced acellular lanes. In each case, there was a clear delay between when the wave reached the proximal lane border, and when it appeared on the other side. Then, the curve rose abruptly as the signal began on the other side of the lane and propagated further. The curves in Fig. 2B are similar to those in Fig. 2A, except for the pauses at the proximal edge of the acellular lanes. Fitting exponential curves (ignoring the lane-related pauses), showed that the calcium waves were similar in confluent regions and across cell-free lanes: neither B nor τ were significantly different between the calcium waves in confluent regions and the waves crossing lanes (P > 0.4 for each parameter, Student’s t test). Velocities measured at the midpoint of the exponential curves fit to the cell-free lane data (as in Fig. 2B, n = 9) did not differ significantly from the velocities at the corresponding points on the exponential curves fit to waves through confluent regions in the same fields (P > 0.35, paired Student’s t test). The constancy of velocity suggests that diffusion is rate limiting in propagation.
Figure 2.

Wave progression in confluent astrocytic monolayers and across cell-free lanes. The distance of the wave front (F/Fo>1.15) from the point of stimulation was measured in images taken at 1-sec intervals. The symbols indicate the measured distance; the solid curves are the least-squares best fit to an exponential function. The traces have been shifted vertically for clarity. The first three time points following initiation of the calcium wave were not included; the waves continued beyond the microscope field in each case. The tick marks on the y axis represent 100 μm distances. (A) Three representative traces of waves in confluent monolayers are shown. Although the wave velocity tends to decline gradually with distance, each wave showed occasional, brief pauses at cell boundaries. (B) Three representative traces of waves crossing cell-free lanes are shown. The rises in calcium progressed at a relatively constant rate until they reached the lane, in which no calcium signal could be seen. The wave reappeared on the far side of the lane after an interval that increased with lane width. (Lane widths and locations are indicated on the traces.) Since the waves reappeared a short distance beyond the edge of the lane, the jumps in distance are larger than the lane widths.
The probability of crossing diminished with increasing lane width [P < 0.03, n = 43, point-biserial correlation (13)] (Fig. 3A). Thus, waves crossed in 16 of 35 cases when lanes were less than 120 μm in width; in contrast, waves never crossed in eight cases where lanes were greater than 120 μm in width. There was no correlation of probability of crossing and distance from the stimulated cell to the far side of the lane (P > 0.85, n = 43, point-biserial correlation) (Fig. 3B). Furthermore, the stimulation sites for 12 of the 16 waves that did cross were more than 300 μm from the far side of the lane, a distance substantially greater than the widest lane crossed. These observations indicate that release occurs from cells along the path of the wave, and not solely from the electrically stimulated cell. In confirmation of this mechanisms, on seven occasions, the glial wave traveled an indirect route, arriving at a cell at the edge of the lane that was at a distance from the edge cell closest to the stimulation site. In these cases, the wave crossed the lane at a position closest to where the wave reached the lane, not at the closest linear distance from the stimulated cell. In three of these cases, a second wave was evoked from a different region of the field. In all three, this second stimulation demonstrated that a wave traveling along a different path could cross the lane at the site closest to the original stimulation site. Finally, in one case a spontaneous wave appeared to cross a cell-free lane. Taken together, these data indicate that the extracellular signal released at the site of stimulation evokes further release from astrocytes along the path of the calcium wave.
Figure 3.

Probability of crossing a lane depends on lane width and not on distance from the stimulation site. The correlation coefficient and probability (point-biserial correlation) are indicated on the histogram. The number of fields examined is indicated in parentheses above each bar. Data for both histograms are taken from the same experiments. (A) The probability of a calcium wave crossing a cell-free lane decreased with increasing width of the lane. (B) The probability of a calcium wave crossing a cell-free lane was not correlated with the distance from the stimulation site to the far side of the lane.
Because our earlier results implicated glutamate release from astrocytes during calcium waves in activation of calcium transients in neurons (5), we investigated whether glutamate was involved in the extracellular signaling between astrocytes. During nine astrocyte calcium waves in the presence of either the nonselective glutamate antagonist γ-d-glutamyl-glycine (1 mM) or a mixture of: dl-2-amino-5-phosphonovaleric acid (100 μM), 6,7-dinitroquinoxaline-2,3-dione (50 μM), and l-2-amino-3-phosphonoproprionic acid (200 μM), communication across the lane was observed seven times. In most instances these treatments blocked propagation of calcium waves from astrocytes to neurons (5). These data suggest that if glutamate is released, it is not necessary for the propagation of astrocyte calcium waves.
Experimentally Altering the Extracellular Signaling Pathway Alters the Calcium Wave Propagation.
By moving the medium directly over the cells, it should be possible to alter the diffusion of an extracellular signal. Using this rationale in two different sets of experiments, we employed a glass pipette perfusion system and positive or negative pressure to perturb the medium in the vicinity of the stimulated cell. The results of these experiments demonstrate that the presence of an extracellular signal is necessary for the full propagation of a calcium wave.
Direct perfusion with positive pressure applied to the perfusion pipette could be adjusted to provide a variety of responses. Strong perfusion limited calcium waves to a few cells close to the stimulation site, while gentle perfusion altered their shape. An example of the asymmetry produced is seen in Fig. 4. In this case (and in seven of eight of such experiments) passage of medium out of the pipette strongly biased wave propagation away from the perfusion pipette (Fig. 4B). In contrast, the control profile showed a radially propagating calcium wave (Fig. 4A). During perfusion, the number of cells participating in the calcium wave toward the perfusion pipette was significantly less than the number of cells away from the pipette (7 ± 4 toward the pipette, versus 19 ± 14 away from the pipette; P < 0.02, paired Student’s t test); control waves showed no such bias (32 ± 21 toward the pipette versus 28 ± 18 away from the pipette; P > 0.50, paired Student’s t test). In addition, the number of astrocytes participating in the calcium wave decreased during perfusion from 60 ± 36 to 26 ± 18 (P < 0.03, unpaired Student’s t test). Perfusion alone had no effect on the baseline fluorescence of the astrocytes. Additionally, the order of control (pressure off) and experimental (pressure on) stimulation did not influence the results. These data indicate that perfusion distorted the flow of extracellular signal, and accordingly shaped the direction of calcium wave propagation.
Figure 4.

The effect of gentle perfusion on the shape and magnitude of an astrocyte calcium wave. The + indicates the point of stimulation. (Bar = 100 μm.) (A) Astrocytes participating in a control calcium wave are radially distributed around the stimulation site. (B) The same field of astrocytes participating in a calcium wave during perfusion with positive pressure on the pipette. Participating cells are biased away from the perfusion pipette, and the number of participating cells is reduced.
Direct perfusion with negative pressure applied to the pipette produced a similar effect. Fig. 5 shows a typical case in which three waves were evoked sequentially from the same site. Fig. 5A is a control field, showing typical wave propagation. When negative pressure was applied, removing medium from the immediate vicinity of the stimulated astrocyte (Fig. 5B), there was a significant reduction in the area of calcium rise. On termination of perfusion, the stimulated wave returned to control values (Fig. 5C). In five such experiments, the number of cells participating in the calcium wave decreased from 82 ± 44 to 26 ± 11 (P < 0.05, paired Student’s t test). No directional bias was observed in these experiments; however, lower perfusion velocities were not tested. Again, the perfusion itself had no affect on calcium levels. These observations further indicate that perfusion affected an agent released from these astrocytes and thereby altered the calcium wave.
Figure 5.
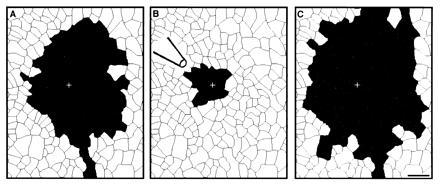
The effect of gentle suction through a pipette close to the stimulation site on the shape and magnitude of astrocytic calcium waves. The + indicates the point of stimulation. (Bar = 100 μm.) (A) Astrocytes participating in a calcium wave in the absence of suction are radially distributed around the stimulation site. (B) The same astrocytes during a calcium wave while negative pressure was applied to the perfusion pipette. The number of participating astrocytes was markedly reduced. (C) Restimulation in the same area with no movement of extracellular fluid showing recovery of the number of participating astrocytes.
DISCUSSION
The data presented here clearly demonstrate that an extracellular signal released from astrocytes during a calcium wave plays an important role in its propagation. The extracellular signal is able to initiate a calcium rise in physically isolated astrocytes, eliminating the possibility of gap-junction-mediated events. Furthermore, the effects of local perfusion demonstrate that the extracellular signal is an integral component of calcium wave propagation within confluent cultures. Finally, the observation that probability of crossing a cell-free lane is independent of distance from the stimulation site indicates that astrocytes in the path of a calcium wave are releasing the signal. The signal is not yet identified and may have multiple components. Although glutamate is an extracellular messenger in glial-neuronal communication, application of several glutamate blockers indicates that it has little role in propagation of calcium waves between astrocytes.
The basic observation of communication between astrocytes that are not in direct physical contact proves that extracellular signaling is sufficient for initiation of a calcium wave. Careful examination of the experimentally generated cell-free lanes, with both phase and fluorescence microscopy, showed that there were no cellular contacts between astrocytes on opposing sides of the lanes even though the astrocyte calcium wave propagated across the lane. The fact that the wave velocity is similar through confluent astrocytes and across an acellular lane also suggests that the diffusion of the extracellular signal is rate limiting in the normal propagation velocity.
Perfusion experiments indicate that the extracellular signal is necessary for the normal propagation of the calcium wave in confluent astrocytes. Perfusion of the stimulus site under positive pressure altered the wave shape and/or decreased the number of participating cells. The potential importance of extracellular signaling is highlighted by the conclusion that it is both necessary and sufficient for maximal glial wave propagation in our cultures.
Previous work has shown activation of glial calcium waves by glutamate (1) and neuronal calcium transients by glutamate released from astrocytes (5, 7). Our experiments using glutamate receptor antagonists clearly indicate that glutamate is not required for calcium wave propagation in our cultures. Blockers of other messengers might show that glutamate is sufficient for propagation. Astrocytes are known to release and respond to a number of agents (8), and the identification of the signaling molecules awaits further study.
While the identity of the extracellular signal remains unknown, it appears to be regenerated by release from astrocytes throughout the path of the wave, and not to come solely from the stimulated cell. If the probability of a calcium wave crossing a lane depended on the distance from the stimulated cell and not on lane width, one would have to conclude that the stimulated cell was releasing most or all of the extracellular messenger. In fact, the probability of crossing was correlated with the lane width and not the distance from the stimulation site, leading to the conclusion that the cells adjacent to the lane were releasing the extracellular signal. Moreover, the stimulation sites for waves that crossed cell-free lanes were usually more distant than the widest lane that was crossed. Thus it appears extracellular signals can be released by cells along the path of the wave.
A Working Model for Astrocyte–Astrocyte Communication.
Other workers have reported evidence that conduction of astrocytic calcium waves is mediated by gap-junctions. Studies utilizing Cx43 knockout mice and blockers of gap-junctional conductance show reduced propagation of calcium waves (8, 14), while transfection of communication-defective cells with connexins increases calcium wave propagation (15). In other experiments calcium waves were observed in the presence of superfusion (2). It seems reasonable that under some circumstances gap junctions may be the sole or at least the primary pathway of calcium wave propagation between glia. However, Charles and Sanderson (unpublished data as reported in ref. 16) did state that perfusion could alter the “pattern of propagation of mechanically induced intercellular calcium waves.” While no supporting data were presented, this observation is consistent with the experimental results presented here.
In our system, which is dominated by an extracellular pathway, many, if not all, of the astrocytes release a signal into the extracellular medium and display elevated intracellular calcium. In principle, if each astrocyte released such a signal in a sequential fashion and stimulated the adjacent astrocyte, one would have a form of saltatory conduction of calcium waves. Each astrocyte could also stimulate itself in an autocrine fashion, if it releases a signal while it is still capable of responding to the signal. This extracellular communication pathway would allow populations of astrocytes not coupled by gap junctions to communicate.
A more general model includes both mechanisms discussed above (Fig. 6). The task remaining then, is to resolve the relative contribution of the gap-junction-mediated and extracellular-mediated pathways in each system. Indeed, the relative contributions of the two pathways may differ in different regions of the brain. The strength of gap-junctional coupling between astrocytes shows regional differences (17). In addition, astrocytes release and respond to a wide range of substances (8) but the ability to respond to a given substance varies between astrocytes. Even sibling astrocytes, derived from a single parent cell by a single mitosis, can respond differentially to a given substance (18). These variations in the strength of gap-junction coupling and the release specificity and responsitivity to an extracellular signal could underlie the variability in calcium wave propagation seen in confluent cultures. In addition, communication between astrocytes might vary within the same cell population over time either developmentally or with physiological state. The communication between astrocytes that underlies calcium waves might be mediated largely by gap-junctions at one time, by extracellular messengers at another, and by a combination of both mechanisms at yet another. Given the additional condition that extracellular space is known to vary substantially as in pathological conditions, the relative contribution of extracellular signaling in vitro might be highly context dependent.
Figure 6.

A model for the pathways mediating an astrocytic calcium wave. The calcium wave is propagated both through an intercellular, gap-junction pathway (2, 15) and through the extracellular pathway defined in this study. Autocrine stimulation is suggested by the dashed arrow, shown only for the leftmost cell. The contribution of each of these pathways to the communication between astrocytes in situ may vary developmentally, regionally, and with physiological state.
The growing body of evidence that neuronal and glial ensembles interact more substantively that previously recognized highlights the importance of defining the communication paths they employ. The prominent extracellular signaling identified here adds a new component to the repertoire of mechanisms available for linking activities of diverse components of the central nervous system.
References
- 1.Cornell-Bell A H, Finkbeiner S M, Cooper M S, Smith S J. Science. 1990;247:470–473. doi: 10.1126/science.1967852. [DOI] [PubMed] [Google Scholar]
- 2.Finkbeiner S. Neuron. 1992;8:1101–1108. doi: 10.1016/0896-6273(92)90131-v. [DOI] [PubMed] [Google Scholar]
- 3.Charles A C, Merrill J E, Dirksen E R, Sanderson M J. Neuron. 1991;6:983–992. doi: 10.1016/0896-6273(91)90238-u. [DOI] [PubMed] [Google Scholar]
- 4.Dani J W, Chernjavsky A, Smith S J. Neuron. 1992;8:429–440. doi: 10.1016/0896-6273(92)90271-e. [DOI] [PubMed] [Google Scholar]
- 5.Hassinger T D, Atkinson P B, Strecker G J, Whalen L R, Dudek F E, Kossel A H, Kater S B. J Neurobiol. 1995;28:159–170. doi: 10.1002/neu.480280204. [DOI] [PubMed] [Google Scholar]
- 6.Nedergaard M. Science. 1994;263:1768–1771. doi: 10.1126/science.8134839. [DOI] [PubMed] [Google Scholar]
- 7.Parpura V, Basarsky T, Liu F, Jeftinija K, Jeftinija S, Haydon P. Nature (London) 1994;369:744–747. doi: 10.1038/369744a0. [DOI] [PubMed] [Google Scholar]
- 8.Martin D L. Glia. 1992;5:81–94. doi: 10.1002/glia.440050202. [DOI] [PubMed] [Google Scholar]
- 9.Cornell-Bell A H, Finkbeiner S M. Cell Calcium. 1991;12:185–204. doi: 10.1016/0143-4160(91)90020-f. [DOI] [PubMed] [Google Scholar]
- 10.Hansen M, Boitano S, Dirksen E R, Sanderson M J. J Cell Sci. 1993;106:995–1004. doi: 10.1242/jcs.106.4.995. [DOI] [PubMed] [Google Scholar]
- 11.Osipchuk Y, Cahalan M. Nature (London) 1992;359:241–244. doi: 10.1038/359241a0. [DOI] [PubMed] [Google Scholar]
- 12.Kao J P Y, Harootunian A T, Tsien R Y. J Biol Chem. 1989;264:8179–8184. [PubMed] [Google Scholar]
- 13.Bruning J L, Kintz B L. Computational Handbook of Statistics. Foresman, Glenview, IL: Scott; 1968. pp. 163–166. [Google Scholar]
- 14.Spray D C, Viera D, El-Sabban M E, Gao Y, Bennett M V L. Soc Neurosci Abstr. 1995;21:563. [Google Scholar]
- 15.Charles A. J Cell Biol. 1992;118:195–201. doi: 10.1083/jcb.118.1.195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Charles A. Dev Neurosci. 1994;16:196–206. doi: 10.1159/000112107. [DOI] [PubMed] [Google Scholar]
- 17.Lee S H, Kim W T, Cornell-Bell A H, Sontheimer H. Glia. 1994;11:315–325. doi: 10.1002/glia.440110404. [DOI] [PubMed] [Google Scholar]
- 18.Shao Y, Porter J T, McCarthy K D. Perspect Dev Neurobiol. 1994;2:205–215. [PubMed] [Google Scholar]