

Abstract
Okawa, Y., Hideshima, T., Ikeda, H., Raje, N., Vallet, S., Kiziltepe,T., Yasui, H., Enatsu, S., Pozzi, S., Breitkreutz, I., Cirstea, D., Santo, L.,Richardson, P. & Anderson, K.C. (2008) Fatty acid synthase is a novel therapeutic target in multiple myeloma. British Journal of Haematology, 141, 659–671.
The above article, published in the online issue, 141·5 on 7 May 2008 in Blackwell Synergy (http://www.blackwell‐synergy.com), has been retracted by agreement between the authors, the Journal Editor‐in‐Chief and Blackwell Publishing Ltd.
Finbarr Cotter
Editor‐in‐Chief
British Journal of Haematology
Summary
This study investigated the biological significance of the inhibition of fatty acid synthase (FAS) in multiple myeloma (MM) using the small molecule inhibitor Cerulenin. Cerulenin triggered growth inhibition in both MM cell lines and MM patient cells, and overcame the survival and growth advantages conferred by interleukin‐6, insulin‐like growth factor‐1, and bone marrow stromal cells. It induced apoptosis in MM cell lines with only modest activation of caspase ‐8, ‐9, ‐3 and PARP; moreover, the pan‐caspase inhibitor Z‐VAD‐FMK did not inhibit Cerulenin‐induced apoptosis and cell death. In addition, treatment of MM cells with Cerulenin primarily up‐regulated apoptosis‐inducing factor/endonuclease G, mediators of caspase‐independent apoptosis. Importantly, Cerulenin induced endoplasmic reticulum stress response via up‐regulation of the Grp78/IRE1α/JNK pathway. Although the C‐Jun‐NH2‐terminal kinase (JNK) inhibitor SP600215 blocked Cerulenin‐induced cytotoxicity, it did not inhibit apoptosis and caspase cleavage. Furthermore, Cerulenin showed synergistic cytotoxic effects with various agents including Bortezomib, Melphalan and Doxorubicin. Our results therefore indicate that inhibition of FAS by Cerulenin primarily triggered caspase‐independent apoptosis and JNK‐dependent cytotoxicity in MM cells. This report demonstrated that inhibition of FAS has anti‐tumour activity against MM cells, suggesting that it represents a novel therapeutic target in MM.
Keywords: multiple myeloma, fatty acid synthase, apoptosis, JNK
Multiple myeloma (MM) is characterized by the clonal proliferation of malignant plasma cells in the bone marrow (BM) associated with bone lesion, renal failure and immunodeficiency. Despite conventional therapies including high‐dose chemotherapy with stem cell transplantation, it remains incurable due to both intrinsic and acquired drug resistance. Novel biologically‐based treatments targeting MM cells as well as the BM microenvironment, including thalidomide, lenalidomide and Bortezomib, offer great promise to improve patient outcome (Kyle & Rajkumar, 2004; Richardson et al, 2004; Hideshima et al, 2007).
Fatty acid synthesis occurs in all living organism and provides for essential components of both biological membranes and energy storage. Fatty acid synthase (FAS) is a multifunctional enzyme that catalyzes the terminal steps in the synthesis of the 16‐carbon fatty acid palmitate in cells (Wakil, 1989; Asturias et al, 2005). In normal cells, FAS expression levels are relatively low, since fatty acid is generally supplied by dietary fatty acid. In contrast, FAS is expressed at significantly higher levels in a variety of human cancers including breast, colon, ovary, lung, prostate and others (Milgraum et al, 1997; Kuhajda, 2000, 2006; Pizer et al, 2001; Baron et al, 2004; Menendez & Lupu, 2007). Moreover, several reports have shown that FAS expression levels correlate with tumour progression, aggressiveness and metastasis (Kuhajda, 2000, 2006; Rossi et al, 2003). For example, FAS expression levels are predictive of poor prognosis in breast and prostate cancer (Kuhajda, 2006). As FAS has been strongly linked to tumour cell proliferation (Kuhajda, 2000) and is preferentially expressed in cancer cells, it may represents an attractive target for novel anticancer therapy (Kuhajda, 2006; Menendez & Lupu, 2007). Importantly, Cerulenin, a specific inhibitor of FAS derived from a natural product of Cephalosporium caerulens, inhibits tumour growth and induces apoptosis in a variety of cancer cell lines (1996a, 1996b; Heiligtag et al, 2002). It irreversibly inhibits FAS by binding covalently to cysteine at the active site of the β keto acyl synthase moiety, which catalyzes the condensation reaction between the elonging fatty acid chain and each successive acetyl or malonyl residue. Other FAS inhibitors, such as C75 and orlistat, also show antitumour activity in vitro and in vivo (Kuhajda et al, 2000; Gabrielson et al, 2001; Kridel et al, 2004). The effect of FAS inhibitor on cancer is further verified by RNA interference to downregulate FAS in prostate and breast cancer cells, which induces apoptosis (De Schrijver et al, 2003; Bandyopadhyay et al, 2006).
This study determined the biological significance of targeting FAS in MM using the small molecule inhibitor Cerulenin. Cerulenin triggers growth inhibition in both MM cell lines and patient MM cells, as well as overcomes the survival and growth advantages conferred by interleukin‐6 (IL‐6), insulin‐like growth factor‐1 (IGF‐1) and bone marrow stromal cells (BMSC). It induces apoptosis in MM cell lines with only modest activation of caspase ‐8, ‐9, ‐3 and Poly (ADP‐ribose) polymerase (PARP); moreover, the pan‐caspase inhibitor Z‐VAD‐FMK does not inhibit Cerulenin‐induced apoptosis and cell death. During Cerulenin‐induced apoptosis, both apoptosis‐inducing factor (AIF) and endonuclease G (Endo G) are released from the mitochondria to cytosol, suggesting that Cerulenin triggers apoptosis, at least in part, via a caspase‐independent, AIF/Endo G pathway. Importantly, Cerulenin also induces endoplasmic reticulum (ER) stress response via up‐regulation of Grp78/IRE1α/JNK pathway. Although the C‐Jun‐NH2‐terminal kinase (JNK) inhibitor, SP600215, blocks Cerulenin‐induced cytotoxicity, it does not inhibit apoptosis and caspase cleavage triggered by Cerulenin. Importantly, Cerulenin triggers synergistic cytotoxicity with various agents including Bortezomib, Melphalan and Doxorubicin, which induce apoptosis via activation of caspases. Our results therefore indicate that inhibition of FAS by Cerulenin triggers primarily caspase‐independent apoptosis and JNK‐dependent cell death in MM, confirming that FAS represents a novel therapeutic target in MM.
Materials and methods
Reagents
The FAS inhibitor, Cerulenin, was provided by EMD chemicals (San Diego, CA, USA). Cerulenin, (2R, 3S)‐2,3‐epoxy‐4‐oxo‐7,10‐trans, trans‐dodecadienamide (C12H17NO3), a natural product of Cephalosporium caerulens, is a specific inhibitor of FAS enzyme across a broad phylogenetic spectrum (Vance et al, 1972; Omura, 1976; Moche et al, 1999). It was dissolved in dimethyl sulphoxide (DMSO) at 50 mmol/l stock solution and stored at −20°C. Recombinant human IL‐6, IGF‐1 (R&D Systems, Minneapolis, MN, USA) were reconstituted with sterile phosphate‐buffered saline (PBS) containing 0·1% bovine serum albumin (BSA). Pan‐caspase inhibitor Z‐VAD‐FMK (Bachem, Bubendorf, Switzerland) was dissolved in methanol, stored at −20°C, and used at 25 μmol/l. JNK inhibitor SP600215 (Calbiochem, San Diego, CA, USA) was stored at −20°C, and used at 5–10 μmol/l. Tunicamycin (Sigma Chemical, St Louis, MO, USA) was dissolved in DMSO at 10 mg/ml stock solution and stored at 4°C, and used at 5 μg/ml. Melphalan and Doxorubicin were purchased from Sigma Chemical. Bortezomib was obtained from Millenium Pharmaceuticals Inc., Cambridge, MA, USA.
Cells
Dex‐sensitive (MM.1S) and resistant (MM.1R) human MM cell lines were kindly provided by Dr Steven Rosen (Northwestern University, Chicago, IL, USA). RPMI‐8226 and U266 human MM cell lines were obtained from American Type Culture Collection (Manassas, VA, USA). RPMI 8226‐Dox40 (Doxorubicin‐resistant) and RPMI 8226‐LR5 (Mel‐resistant) human MM cells were kindly provided by Dr William Dalton (Lee Moffitt Cancer Center, Tampa, FL, USA). OPM1 and OPM2 were provided from by Dr P. Leif Bergsagel (Mayo Clinic, Tucson, AZ, USA). Each of these cell lines were cultured in RPMI‐1640 containing 10% fetal bovine serum (FBS; Sigma Chemical), 2 μmol/l l‐glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin (GIBCO, Grand Island, NY, USA). Fresh peripheral blood mononuclear cells (PBMNC) obtained from three healthy volunteers were separated from heparinized peripheral blood by Ficoll‐Hypaque (Pharmacia, Piscataway, NJ, USA) density sedimentation.
Primary MM cells and BMSCs from MM patients, and plasma cells from healthy volunteers
Tumour cells freshly isolated from MM patients and plasma cells from healthy volunteers (>90% CD138+) were purified by CD138 positive selection using CD138 (Syndecan‐1) Micro Beads and the Auto MACS magnetic cell sorter (Miltenyi Biotec Inc., Auburn, CA, USA). BM mononuclear cells separated by Ficoll‐Hypaque were cultured in Dulbecco’s modified Eagles medium (Sigma Chemical) supplemented with 15% heat‐inactivated FBS, 2 μmol/l l‐glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin for 3–6 weeks to generate BMSCs. Approval for these studies was obtained from the Dana‐Farber Cancer Institute Institutional Review Board. Informed consent was obtained from all patients in accordance with the Declaration of Helsinki protocol.
Growth inhibition assay
The growth inhibitory effect of Cerulenin in MM cell lines, primary MM cells, PBMNC and normal plasma cells was assessed by measuring 3‐(4,5‐dimethylthiazol‐2‐yl)‐2,5‐diphenyl tetrasodium bromide (MTT; Sigma) dye absorbance. Cells were plated in 96‐well microtitre plates at a density of 2–3 × 104 cells per well, and each plate was incubated for 24 and/or 48 h, with MTT added to each well for the last 4 h. The absorbance of each well was measured at 570/630 nm using a spectrophotometer (Molecular Devices, Sunnyvale, CA, USA).
To measure proliferation of MM cells and BMSC, the rate of DNA synthesis was measured, as described previously (Hideshima et al, 2000). MM cells were incubated in 96‐well culture plates (Costar, Cambridge, MA, USA) in the presence of Cerulenin and/or recombinant cytokines IL‐6 or IGF‐1 for 48 h at 37°C. Cells were pulsed with 18·5 kBq/well of [3H]‐thymidine ([3H]‐TdR; Perkin Elmer, Boston, MA, USA) during the last 8 h of culture, harvested onto glass filters with an automatic cell harvester (Cambridge Technology, Cambridge, MA, USA), and counted using the LKB Betaplate scintillation counter (Wallac, Gaithersburg, MD, USA). MTT and 3H uptake analysis were performed in triplicate or quadruplicate cultures.
Immunoblotting
The MM cells were harvested, washed twice with PBS, and lysed using lysis buffer: 50 mmol/l Tris–HCl (pH 7·4), 150 mmol/l NaCl, 1% Nonidet P‐40, 10 mmol/l sodium pyrophosphate, 5 mmol/l EDTA, 1 mmol/l EGTA, 2 mmol/l Na3VO4, 5 mmol/l NaF, 1 mmol/l phenylmethylesulphonyl fluoride (PMSF), 5 μg/ml leupeptin and 5 μg/ml aprotinin, as described previously (Hayashi et al, 2002). Subcellular proteins were extracted from 2 × 107 viable cells using a mitochondria isolation kit (Pierce, Rockford, IL, USA). The whole cell lysates or fractionated proteins (20 μg per lane) were subjected to sodium dodecyl sulphate–polyacrylamide gel electrophoresis, transferred to nitrocellulose membranes (Bio‐Rad Laboratories, Hercules, CA, USA), and immunoblotted with anti‐PARP, caspase‐8, caspase‐3, caspase‐9, AIF, voltage‐dependent anion channel (VDAC), ASK1, GRP78/Bip, IRE1α, p‐JNK, JNK, p‐ATF‐2, p‐c‐Jun and glyceraldehyde‐3‐phosphate dehydrogenase (GAPDH) (Cell Signaling, Beverly, MA, USA); as well as with anti‐α‐tubulin antibodies (Abs) (Santa Cruz Biotechnology, Inc. Santa Cruz, CA, USA), anti‐Fatty Acid Synthase Abs (BD Bioscience, San Jose, CA, USA), anti‐Endo G (ProSci Inc., Poway, CA, USA) and anti‐XBP‐1 (Abcam, Inc. Cambridge, MA, USA) Abs.
Flow cytometric analysis
For detection of apoptotic cells, mitochondrial protein staining was performed with pycoerythrin‐labeled anti‐Apo2·7 antibody (Immunotec, Marseille, France). Cells were treated with various concentrations of Cerulenin for 24 h, washed with PBS, and then incubated with Apo2·7 for 30 min. Apo2·7 staining was analysed using the RXP cytomics software on an Epics flow cytometer (Coulter Immunology, Hialeah, FL, USA).
Immunocytochemistry
Cytospins of cells were prepared on glass slides and fixed with 50% methanol/50% acetone at 4°C. The slides were blocked in PBS with 5% FBS at room temperature (RT), and then incubated with primary antibody overnight at 4°C. Fluorescein isothiocyanate (FITC)‐labeled secondary antibody reaction for 30 min was then performed. Coverslips were mounted on the glass slides with VectaShield antifade/4′,6‐diamidino‐2‐phenylindole (DAPI), which were analysed by Nikon E800 fluorescence microscopy (Nikon Inc., Melville, NY, USA).
Statistical analysis
Statistical significance of differences observed in drug‐treated compared with control cultures was determined using the Mann–Whitney test. The minimal level of significance was P < 0·05. All statistical analyses were determined using GraphPad Prism software (GraphPad Software, Inc. San Diego, CA, USA).
Isobologram analysis
The interaction between Cerulenin and Bortezomib, Melphalan, and Doxorubicin was analysed using CalcuSyn software program (Biosoft, Ferguson, MO, USA) to determine whether the combination was additive or synergistic, as described previously (Chou & Talalay, 1984; Raje et al, 2004). This program is based upon the Chou‐Talalay method, which calculates a combination index (CI). Analysis is performed based on the following equation: CI = (D)1/(Dx)1 + (D)2/(Dx)2 + (D)1(D)2/(Dx)1(Dx)2, where (D)1 and (D)2 are the doses of drug 1 and drug 2 that have x effect when used in combination; whereas (Dx)1 and (Dx)2 are the doses of drug 1 and drug 2 that have the same x effect when used alone. A CI c. 1·1 represents the conservation isobologram and indicates additive effects, whereas CI c. <0·9 indicates synergism.
Results
FAS expression in various cells
We first examined baseline expression of FAS in various cells. FAS protein was expressed in all MM cell lines (Fig 1A and B; lane 5), as well as in primary tumour cells from MM patients (Fig 1B; lane 4, Fig 1C). Importantly, FAS expression was higher in both MM cell lines and primary tumour cells than in normal plasma cells, as assessed by Western blotting (Fig 1B) and confirmed by immunocytochemical analysis (Fig 1D).
Figure 1.
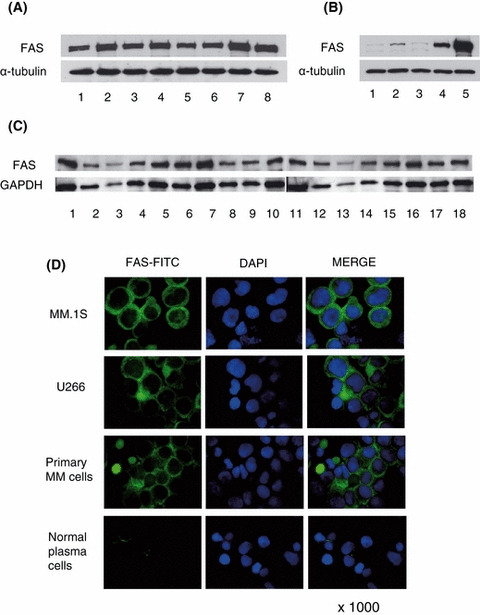
FAS expression in various cells. Cell lysates (20 μg) of MM cell lines (A), normal cells and MM cells (B), and patient cells (B; lane 4, C) were immunoblotted with anti‐FAS antibody. (A) FAS expression was detected in all MM cell lines: lane 1, U266; lane 2, MM.1S; lane 3, MM.1R; lane 4, RPMI8226; lane 5, RPMI Dox40; lane 6, RPMI LR5; lane 7, OPM1; and lane 8, OPM2. (B) FAS expression level was compared in plasma cells and MM cells (lane 1–3, normal plasma cells; lane 4, primary MM cells; lane 5, MM.1S). FAS protein was more highly expressed in MM.1S and primary MM cells than in plasma cells. (C) FAS protein was expressed in all (18/18) primary MM cells. (D) FAS expression in MM cell lines, primary MM cells and normal plasma cells was analysed by immunocytochemistry. FITC‐labeled FAS, nuclear staining by DAPI, and combined staining (Merge) were evaluated by fluorescence microscopy (×1000). Green and blue signal show FAS‐FITC and DAPI respectively. FAS protein in MM cells is most abundant in the cytoplasm with only weak non‐specific of nuclear membrane staining.
Cerulenin inhibits growth of MM cells
We next examined the effect of FAS inhibition by Cerulenin (C12H17NO3; Fig 2A) on growth of MM cells and normal cells, including PBMNC and normal plasma cells, using the MTT assay. Cerulenin significantly inhibited the growth of drug‐sensitive MM.1S, U266, RPMI8226, OPM1 and OPM2 MM cell lines, with a 50% inhibitory concentration (IC50) at 24 h of 24·16, 22·7, 24·03, 37·03 and 21·53 μmol/l, respectively, and IC50 at 48 h of 12·59, 11·12, 17·08, 11·45 and 9·71 μmol/l respectively (Fig 2B and C). Cerulenin also inhibited growth of Dex‐resistant MM.1R, Mel‐resistant RPMI‐LR5, Dox‐resistant RPMI‐Dox40 MM cell lines, with IC50 at 24 h of 22·59, 86·21 and 33·29 μmol/l, and IC50 at 48 h of 10·52, 22·73 and 16·52 μmol/l respectively (Fig 2B and C). However, Cerulenin did not induce cytotoxicity in PBMNC and normal plasma cells from three healthy volunteers (Fig 2E and F). Importantly, Cerulenin induced dose‐dependent cytotoxicity against CD138 positive MM cells (IC50 at 24 h of 27·37 μmol/l) isolated from three patients whose disease was refractory to Dexamethasone, Melphalan, Thalidomide, or Bortezomib therapy (Fig 2D). These results indicate that FAS inhibition by Cerulenin selectively and potently induces cytotoxicity in MM cell lines as well as primary MM cells, even those resistant to conventional and novel therapy.
Figure 2.
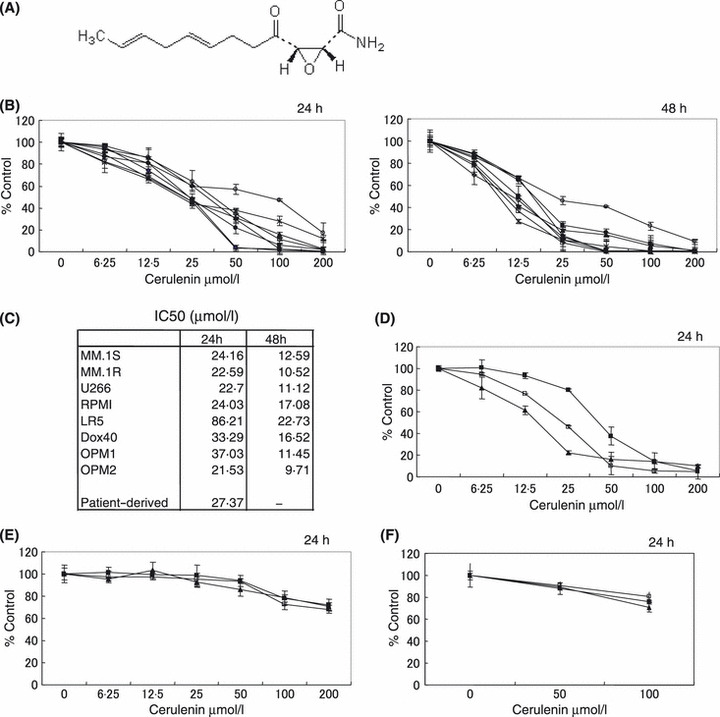
Cerulenin inhibits MM cell growth. (A) Chemical structure of Cerulenin. (B) MM.1S (), MM.1R (□), U266 (), RPMI8226. (•), RPMI LR5 (○), RPMI Dox40 (◆), OPM1 (⋄) and OPM2 (*) cells were cultured for 24 or 48 h in the presence of Cerulenin. (C) IC50 of MM cell lines treated with Cerulenin for 24 or 48 h. (D) Cerulenin inhibits growth of primary MM patient cells [#1 (), #2 (□), #3 (), n = 3]. (E) Cerulenin does not affect viability of PBMNC derived from three healthy volunteers [#1 (), #2 (□), #3 (), n = 3]. (F) Cerulenin does not affect viability of plasma cells derived from three healthy volunteers [#1 (), #2 (□), #3 (), n = 3]. Cell growth was assessed by MTT assay (B, C, D, E, F), and data represent mean ± SD of triplicate cultures.
Cerulenin overcomes the growth stimulatory effects of cytokines, and inhibits MM cell growth in culture with BMSCs
It has been demonstrated that IL‐6 and IGF‐1 induce both growth and inhibition of apoptosis in MM cells (Jourdan et al, 2000; 2002, 2004), therefore, we examined whether FAS inhibition can overcome the effects of exogenous IL‐6 and IGF‐1. Both IL‐6 (10 ng/ml) and IGF‐1 (50 ng/ml) triggered increased growth in MM.1S and U266 cells, which was inhibited by Cerulenin (P < 0·05) (Fig 3A and B). As we previously have shown that the BM microenvironment confered growth and drug resistance in MM cells (Hideshima et al, 2004), we next examined the effect of FAS inhibition on MM cell growth in the BM milieu. MM.1S and U266 cells were cultured for 48 h with or without BMSC, in the presence or absence of Cerulenin. MM cell adherence to BMSC enhanced [3H]‐thymidine uptake in MM cells, which was inhibited by Cerulenin (P < 0·05) (Fig 3C). These results indicate that Cerulenin also blocks the growth stimulatory effect of the BM microenvironment on MM cells.
Figure 3.
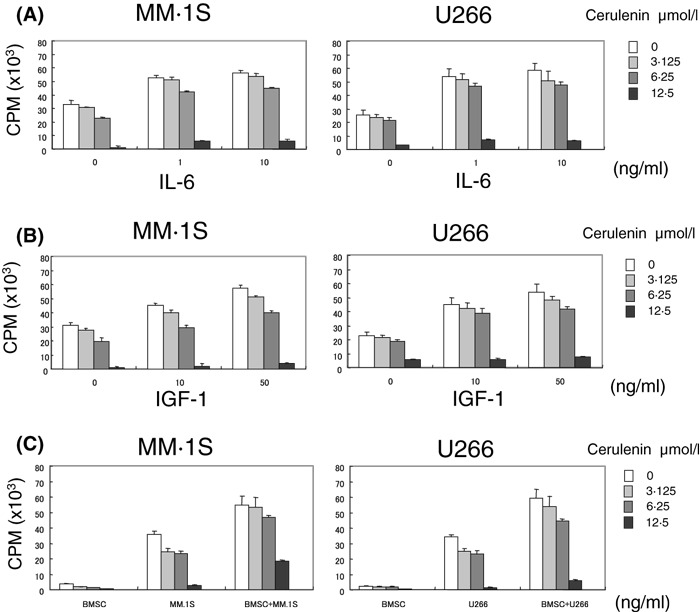
Cerulenin overcomes the protective effects of IL‐6, IGF‐1 and BMSCs on MM cell growth. (A, B, C) MM.1S and U266 cells were cultured for 48 h with the indicated concentrations [0 μmol/l (□), 3·15 μmol/l ( ) 6·25 μmol/l (
) 6·25 μmol/l ( ), 12·5 μmol/l (
), 12·5 μmol/l ( )] of Cerulenin, in the presence or absence of IL‐6 (1 or 10 ng/ml: A), IGF‐1 (10 or 50 ng/ml: B), or BMSC (C). Cell growth was assessed by [3H]‐thymidine uptake. Cerulenin inhibits MM cell growth and overcomes the stimulating effect of IL‐6 (A) or IGF‐1 (B) (P < 0·05), and BMSC (P < 0·05) (C). Values represent mean ± SD of quadruplicate cultures.
)] of Cerulenin, in the presence or absence of IL‐6 (1 or 10 ng/ml: A), IGF‐1 (10 or 50 ng/ml: B), or BMSC (C). Cell growth was assessed by [3H]‐thymidine uptake. Cerulenin inhibits MM cell growth and overcomes the stimulating effect of IL‐6 (A) or IGF‐1 (B) (P < 0·05), and BMSC (P < 0·05) (C). Values represent mean ± SD of quadruplicate cultures.
Cerulenin induces apoptosis via activation of caspase‐independent pathway
To analyse the mechanism of cytotoxicity triggered by Cerulenin in MM cells, we next performed Apo2·7 analysis and immunoblotting in MM.1S and U266 cells. Cerulenin induced apoptosis in a dose‐dependent manner, evidenced by Apo2·7 analysis (Fig 4A, Fig S1C). Cerulenin also triggers caspase ‐8, ‐9, ‐3, and PARP cleavage in MM.1S and U266 (Fig 4B, Fig S1A). Although the pan‐caspase inhibitor Z‐VAD‐FMK blocked Cerulenin‐induced caspase and PARP cleavage in both MM.1S and U266 cells (Fig 4B, Fig S1A), it did not inhibit Cerulenin‐triggered apoptosis (Fig 4D, Fig S1C) and cell death (Fig 4C, Fig S1B). During Cerulenin‐induced apoptosis in MM cells, both AIF and Endo G were significantly released from the mitochondria to cytosol in a time‐dependent manner (Fig 4E). These results suggest that Cerulenin triggers apoptosis, at least in part, via a caspase‐independent, AIF/Endo G pathway.
Figure 4.
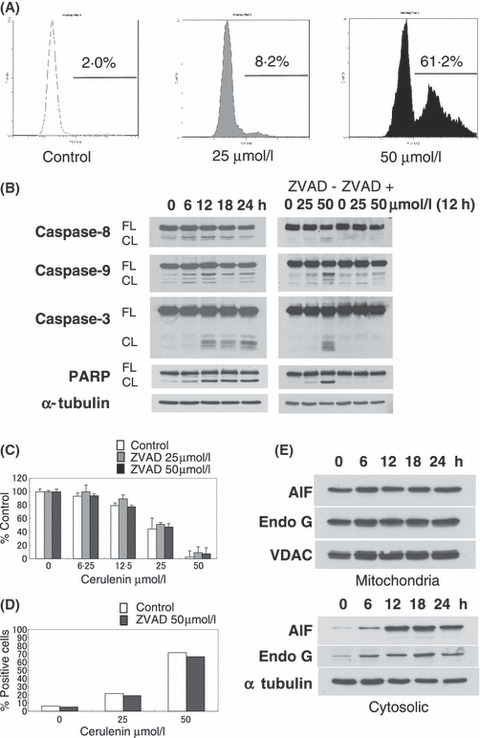
Cerulenin induces apoptosis via activation of caspase‐independent pathway. (A) MM.1S cells were cultured for 24 h with Cerulenin at the indicated doses. Induction of apoptosis by Cerulenin was determined by Apo2·7 staining and flow‐cytometric analysis. (B) MM.1S cells were cultured with Cerulenin (50 μmol/l) for the indicated times (left panel), and preincubated with or without Z‐VAD‐FMK (50 μmol/l) for 1 h prior to treatment with Cerulenin for 12 h at the indicated doses (right panel). Total cell lysates (20 μg /lane) were subjected to Western blotting using anti‐caspase ‐8, ‐9, ‐3, PARP, and α‐tubulin Abs. FL, CF indicate the full length and cleaved form, respectively. (C, D) MM.1S cells were treated with the indicated dose of Cerulenin for 24 h, with or without Z‐VAD‐FMK (25 μmol/l or 50 μmol/l) 1 h pretreatment. Cytotoxicity was determined by MTT assay (C). Values represent mean ± SD of quadruplicate cultures. The percentage of apoptotic cells was determined by flow‐cytometric analysis for APO2·7 staining (D). (E) Mitochondrial proteins AIF and Endo G were released into the cytosolic fraction from mitochondria after Cerulenin (50 μmol/l) treatment in MM.1S cells. Total cell lysates (20 μg/lane) were subjected to Western blotting using anti‐AIF, Endo G, VDAC and α‐tubulin Abs.
Cerulenin induces ER stress response via up‐regulation of GRP78, IRE1α, JNK pathway
As some reports have demonstrated that FAS inhibitor induced ER stress response in tumour cells (Little et al, 2007), we next examined whether FAS inhibition by Cerulenin triggers activation of GRP78, IRE1α, JNK pathway as an ER stress response in MM cells. The ER stress inducer Tunicamycin significantly up‐regulated Grp78, IRE1α, ASK1, JNK and XBP‐1 in MM.1S cells (Fig 5A). In addition, Cerulenin treatment also induced up‐regulation of Grp78, IRE1α and p‐JNK, as well as its downstream molecules p‐ATF‐2 and p‐c‐Jun (Fig 5A). Importantly, JNK inhibitor SP600215 significantly blocks Cerulenin‐induced cytotoxicity in MM.1S and U266 cells, (P < 0·05) (Fig 5B, Fig S2A). We also confirmed that SP600215 markedly inhibited JNK signaling pathway in MM.1S cells by Western blotting (Fig 5E); however, it did not inhibit apoptosis and caspase cleavage triggered by Cerulenin in both MM.1S and U266 cells (Fig 5C and D, Fig S2B and C). In addition, combination pretreatment with JNK inhibitor and ZVAD‐FMK did not enhance the inhibitory effect of JNK inhibitor alone, indicating that JNK activation is the major pathway mediating Cerulenin‐induced cytotoxicity in MM cells (Fig S3). These results strongly indicate that targeting FAS by Cerulenin induces ER stress response via up‐regulation of Grp78/IRE1α/JNK pathway, associated with JNK‐dependent, non‐apoptotic cell death in MM cells.
Figure 5.
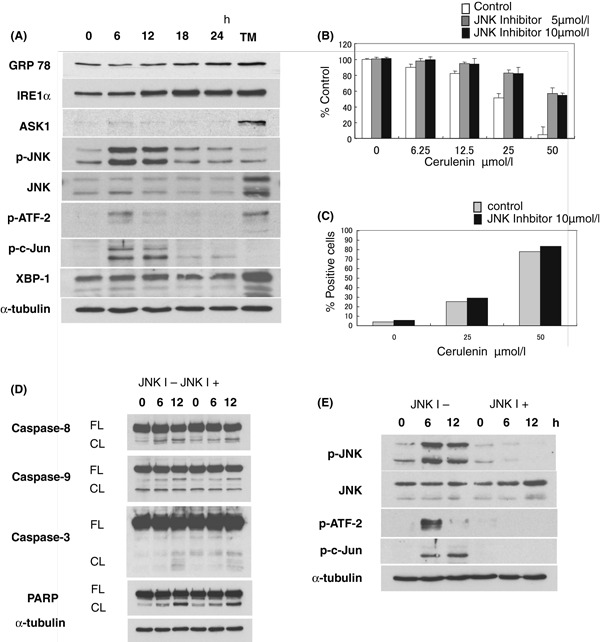
Cerulenin triggers ER stress response and JNK activation. (A) MM.1S cells were cultured with Cerulenin (50 μmol/l) for indicated times. Induction of ER stress response by Cerulenin was examined by Western blotting. MM.1S cells treated with 5 μg/ml Tunicamycin (TM) for 24 h were used as a positive control of ER stress induction. Total cell lysates (20 μg protein/lane) were analysed by anti‐GRP78, IRE1α, ASK1, p‐JNK, JNK, p‐ATF‐2, p‐c‐Jun, XBP‐1 and α‐tubulin Abs. (B, C) MM.1S cells were treated with the indicated dose of Cerulenin for 24 h, with or without JNK inhibitor SP600215 (5 μmol/l or 10 μmol/l) pretreatment for 1 h. Cytotoxicity was determined by MTT assay (B). Values represent mean ± SD of quadruplicate cultures. JNK inhibitor significantly blocks Cerulenin‐induced cytotoxicity (P < 0·05) (B). The percentage of apoptotic cells was determined by flow‐cytometric analysis for APO2·7 staining (C). (D, E) MM.1S cells were cultured with Cerulenin (50 μmol/l) for the indicated times, with or without SP600215 (10 μmol/l) pretreatment for 1 h. Total cell lysates (20 μg/lane) were subjected to Western blotting using anti‐caspase ‐8, ‐9, and ‐3, PARP, p‐JNK, JNK, p‐ATF‐2, p‐c‐Jun and α‐tubulin Abs. FL, CF indicate the full length and cleaved form respectively.
Cerulenin combined with Bortezomib, Melphalan, or Doxorubicin induces synergistic cytotoxicity against MM cells
We further examined whether Cerulenin enhances the growth inhibitory effect of other conventional agents in MM cells. MM.1S, U266, and primary MM cells were cultured for 24 h with Bortezomib (0–3·75 nmol/l), Melphalan (0–10 μmol/l) or Doxorubicin (0–400 nmol/l) combined with Cerulenin (0–25 μmol/l). Cerulenin enhanced growth inhibition triggered by Bortezomib, Melphalan and Doxorubicin, as analysed by MTT assay (Fig 6, Fig S4). Next, we analysed the interaction between Cerulenin and these agents by isobologram analysis using CalcuSyn software program (Raje et al, 2004). Based upon the Chou‐Talalay method to calculate the CI, we showed synergistic cytotoxic effects between Cerulenin and Bortezomib, Melphalan, and Doxorubicin, with a maximum CI of 0·567, 0·188 and 0·529, respectively, in MM.1S cells; 0·63, 0·404 and 0·079, respectively, in primary MM cells; and 0·879, 0·591, and 0·301, respectively, in U266 cells. These data demonstrate that these combination therapies enhance Cerulenin‐ induced cytotoxicity synergistically in MM cells, due to the fact that these agents induce caspase‐dependent apoptosis whereas Cerulenin triggers primarily caspase‐independent apoptosis.
Figure 6.
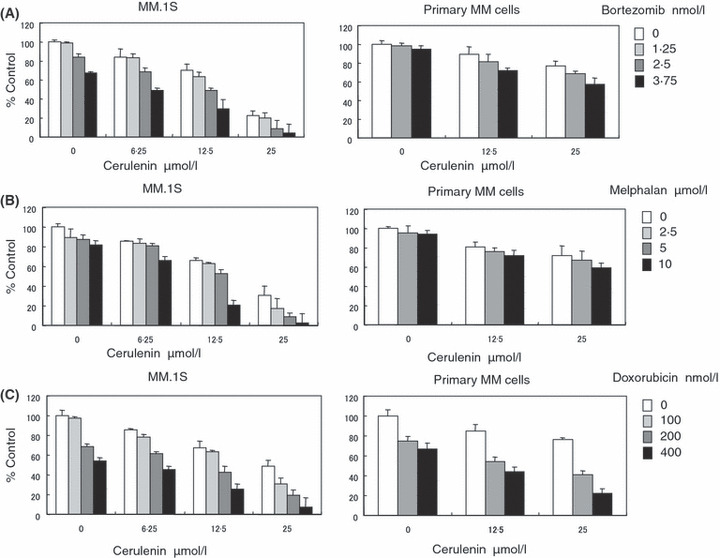
Cerulenin combined with Bortezomib, Melphalan, or Doxorubicin induces synergistic cytotoxicity against MM cells. MM.1S and primary MM cells were cultured for 24 h with the indicated concentrations of Bortezomib (A), Melphalan (B), and Doxorubicin (C) [control media (□), 1·25 nmol/l Bortezomib ( ), 2·5 nmol/l Bortezomib (
), 2·5 nmol/l Bortezomib ( ), 3·75 nmol/l Bortezomib (
), 3·75 nmol/l Bortezomib ( ); control media (□), 2·5 μmol/l Melphalan (
); control media (□), 2·5 μmol/l Melphalan ( ), 5 μmol/l Melphalan (
), 5 μmol/l Melphalan ( ), 10 μmol/l Melphalan (
), 10 μmol/l Melphalan ( ); control media (□), 100 nmol/l Doxorubicin (
); control media (□), 100 nmol/l Doxorubicin ( ), 200 nmol/l Doxorubicin (
), 200 nmol/l Doxorubicin ( ), 400 nmol/l Doxorubicin (
), 400 nmol/l Doxorubicin ( )], in the presence or absence of Cerulenin (0–25 μmol/l). Cell growth was assessed by MTT assays. Data represent mean (±SD) of triplicate cultures. The combination index (CI) of Cerulenin and these agents was analysed using CalcuSyn software program.
)], in the presence or absence of Cerulenin (0–25 μmol/l). Cell growth was assessed by MTT assays. Data represent mean (±SD) of triplicate cultures. The combination index (CI) of Cerulenin and these agents was analysed using CalcuSyn software program.
Discussion
This report demonstrated that FAS inhibition by Cerulenin induced cytotoxicity in drug‐sensitive and drug‐resistant MM cell lines and in primary patient MM cells, with an IC50 at 24 h of 21·53–86·21 μmol/l. In contrast, we observed no cytotoxicity against PBMNC and normal plasma cells, indicating selective and potent cytotoxicity against MM tumour cells. Immunoblot and immunocytochemical analysis revealed that FAS protein was more highly expressed in all MM cell lines and primary MM cells than in normal plasma cells, consistent with prior reports of FAS overexpression in cancer cells (Omura, 1976; Milgraum et al, 1997; Kuhajda, 2000, 2006; Pizer et al, 2001; Baron et al, 2004; Menendez & Lupu, 2007) and further suggesting that it represents a novel therapeutic target in MM.
The BM microenvironment confers drug resistance in MM cells via at least two different mechanisms: (i) cell adhesion mediated drug resistance (CAM‐DR) through adhesion of MM cells to fibronectin and BMSC; and (ii) activation of phosphatidylinositol 3‐kinase (PI3‐K)/Akt and/or Janus kinase 2 (JAK2)/signal transducers and activators of transcription 3 (STAT3) signaling due to high levels of cytokines in the BM milieu (such as IL‐6 and IGF‐1) (Damiano et al, 1999; Hazlehurst et al, 2000; 2001, 2004). Importantly, treatments targeting the BM microenvironment as well as the MM cell can overcome drug resistance in both preclinical and clinical studies (Mitsiades et al, 2004; Hideshima et al, 2007). Our data indicate that Cerulenin overcomes the survival and growth advantages conferred by IL‐6, IGF‐1 and BMSC, strongly suggesting its utility to overcome conventional drug resistance in MM.
Although caspases are the key executors of apoptosis, recent studies suggest that caspase activation is not the sole pathway for inducing apoptosis or necrosis (Jaattela & Tschopp, 2003; Abraham & Shaham, 2004; Lockshin & Zakeri, 2004). For example, caspase‐independent cell death in MM cells is triggered by AS2O3 and inosine monophosphate dehydrogenase inhibitor VX‐944 treatment (McCafferty‐Grad et al, 2003; Ishitsuka et al, 2005). The AIF/Endo G pathway is a major caspase‐independent apoptotic cascade (Daugas et al, 2000; Ahn et al, 2004; Cande et al, 2004; Cregan et al, 2004). In this pathway, enhanced Bcl‐2 family expression induces mitochondrial membrane permeability, thereby releasing AIF and Endo G from the mitochondria to the cytosol and nucleus, with subsequent chromatin condensation and cell death. The present study showed that MM cell apoptosis triggered by Cerulenin was primarily mediated via a caspase‐independent pathway. Although cleavage of caspase ‐8,‐9,‐3 and PARP was induced by Cerulenin treatment, Z‐VAD‐FMK did not block completely Cerulenin‐induced apoptosis and cell death. Moreover, both AIF and Endo G were released from the mitochondria to cytosol during Cerulenin‐induced apoptosis. These results suggest that Cerulenin triggers apoptosis, at least in part, via a caspase‐independent AIF/Endo G pathway, with only modest activation of caspase ‐8,‐9,‐3 and PARP. As most conventional and novel drugs induce apoptosis in MM cells via activation of caspases, we next examined the effect of combining Cerulenin and other drugs. Cerulenin combined with Bortezomib, Melphalan or Doxorubicin triggered synergistic MM cell cytotoxicity, indicating that it may be useful to sensitize or overcome drug resistance.
The ER stress response is a choreographed series of cellular events activated by specific insults that result in altered ER function (Kaufman, 2004; Xu et al, 2005). The combined effect of this response is the activation of genes associated with an adaptation response. Upon prolonged stress, the adaptation mechanism of the ER stress response is saturated, resulting in cell death. Several studies have demonstrated important associations between lipid synthesis pathways and the ER stress response (Cox et al, 1997; van der Sanden et al, 2004; Sriburi et al, 2004; Tessitore et al, 2004). Inhibition of phospholipid synthesis, especially that of phosphatidylcholine, induces ER stress‐related pathways (van der Sanden et al, 2004). Fatty acid synthesis in general, and FAS activity in particular, occurs in the ER (Swinnen et al, 2003). Importantly, because of the direct connection between FAS activity and phospholipid synthesis, previous reports demonstrated that pharmacologic blockade of FAS activity induced ER stress in tumour cells via activation of PERK/eIF2α and IRE1α/XBP‐1 pathways (Swinnen et al, 2003; Little et al, 2007). In addition, Nakamura et al (2006) reported that activation of the ER stress pathway was associated with survival of MM cells, as they contained a well‐developed ER due to M‐protein production. Our data and other reports indicated that increased FAS expression in tumour cells is important for ER function; and conversely, suggest a role for ER stress in the antitumour effects of FAS inhibitors. The present study showed that Cerulenin treatment induced upregulation of Grp78, IRE1α and p‐JNK, as well as its downstream molecules p‐ATF‐2 and p‐c‐Jun. IRE1α can activate the ASK1/JNK mitogen‐activated protein kinase pathway (Urano et al, 2000; Xu et al, 2005; Li et al, 2006), and activation of JNK is a common response to many forms of stress known to influence the cell‐death machinery (Davis, 2000). Although studies show that multiple pathways can contribute to ER‐stress‐induced apoptosis (Li et al, 2006), the mechanisms by which unresolved ER stress causes cell death are not fully characterized (Szegezdi et al, 2006; Zhao & Ackerman, 2006).
Bandyopadhyay et al (2006) proposed a model for apoptotic signaling in which inhibition of FAS leads to accumulation of malonyl‐CoA, which in turn inhibits carnitine palmitoyltransferase‐1 (CPT‐1) and results in up‐regulation of ceramide, followed by induction of the proapoptotic genes BNIP3, tumour necrosis factor (TNF)‐related apoptosis‐inducing ligand (TNFSF10), and death‐associated protein kinase 2 (DAPK2) . BNIP3 not only induces apoptotic cell death, but also is implicated in necrosis and autophagy (Vande Velde et al, 2000; Daido et al, 2004). TNFSF10 has been shown to induce apoptosis in a wide range of transformed cells (Kelley & Ashkenazi, 2004). In addition, a human single‐chain fragment variable (HW1) antibody against TRAIL receptor 2 induced autophagic cell death predominantly via the JNK pathway in a caspase‐independent manner (Park et al, 2007). DAPK2 is associated with ceramide‐mediated apoptosis (Pelled et al, 2002; Yamamoto et al, 2002), but also induces another cell death pathway mediating autophagic or type II cell death (Inbal et al, 2002; Bialik & Kimchi, 2004) without caspase activation (Inbal et al, 2002). Our data and others suggest that inhibition of FAS by Cerulenin activates multiple death signal pathways in MM cells, including caspase‐independent apoptosis and JNK dependent non‐apoptotic cell death. JNK and its downstream molecules ATF‐2 and c‐Jun induce apoptosis in various tumours (Bossy‐Wetzel et al, 1997; Podar et al, 2007). Moreover, caspase‐independent JNK activation promotes autophagic cell death induced by TNF‐α (Jia et al, 2006) and other chemicals (Borsello et al, 2003; Yu et al, 2004; Park et al, 2007). JNK can activate the mitochondrial pathway including AIF, Endo G, cytochrome C and Smac (Weston & Davis, 2007). However, the exact role of c‐Jun in mediating cell differentiation, growth, survival and apoptosis is not fully understood (Jochum et al, 2001; Vogt, 2001).
In summary, our data showed that inhibition of FAS by Cerulenin induced primarily caspase‐independent apoptosis via upregulation of AIF/Endo G pathway, and JNK‐dependent cell death in MM cells. This report demonstrated that inhibition of FAS had anti‐tumour activity against MM cells, identifying FAS as a novel therapeutic target. Taken together, our data provide the preclinical rationale for clinical trials targeting FAS to improve patient outcome in MM.
Conflict of interest disclosure
The authors declare no competing financial interests.
Author’s contributions
YO designed, performed, analysed research and wrote the paper; TH and NR designed, analysed research and wrote the paper; HI, SV, TK, HY performed and designed research; SE performed research and analysed data; SP, IB, LS, PR performed research; KCA participated in design, coordination and performance of study, assisted in writing the paper and funded the study.
Supporting information
Fig S1. Cerulenin induces apoptosis in U266 cells. (A) U266 cells were cultured with Cerulenin (50 &mgr;mol/l) for the indicated times, with or without Z‐VAD‐FMK (50 &mgr;mol/l) for 1 h pretreatment. Total cell lysates (20 &mgr;g/lane) were subjected to Western blotting using anti‐caspase ‐8, ‐9, ‐3, PARP and &alpha‐tubulin. FL, CF indicated as full length and cleaved form, respectively. (B, C) U266 cells were treated with indicated dose of Cerulenin for 24 h, with or without Z‐VAD‐FMK (25 or 50 mmol/l) 1 h pretreatment. Cytotoxicity was determined by MTT assay (B). Values represent mean ± SD of quadruplicate cultures. The percentage apoptotic cells was determined by flow‐cytometric analysis for APO2.7 staining (C).
Fig S2. JNK inhibitor blocks Cerulenin‐induced cytotoxicity in U266 cells. (A, B) U266 cells were cultured with indicated dose of Cerulenin for 24 h, with or without JNK inhibitor SP600215 (5 or 10 &mgr;mol/l) pretreatment for 1 h. Cytotoxicity was determined by MTT assay (A). Values represent mean ± SD of quadruplicate cultures. JNK inhibitor significantly blocks Cerulenin‐induced cytotoxicity (P < 0.05). The percentage apoptotic cells was determined by flow‐cytometric analysis for APO2.7 staining (B). (C) Total cell lysates (20 mg/lane) were subjected to Western blotting using anti‐caspase ‐8, ‐9, ‐3, PARP and α‐tubulin. FL, CF indicated as full length and cleaved form, respectively.
Fig S3. Analysis of inhibition by JNK inhibitor SP600215 and/or ZVAD‐FMK in MM cells. (A, B) MM.1S and U266 cells were cultured with indicated dose of Cerulenin for 24 h, with or without JNK inhibitor SP600215 (10 &mgr;mol/l) and/or ZVAD‐FMK (50 &mgr;mol/l) pretreatment for 1 h. Cytotoxicity was determined by MTT assay. Values represent mean ± SD of quadruplicate cultures. Although ZVAD‐FMK does not affect cytotoxicity by Cerulenin, JNK inhibitor significantly blocks Cerulenin‐induced cytotoxicity (P < 0.05).
Fig S4. Cerulenin enhances Bortezomib, Melphalan, and Doxorubicin‐induced cytotoxicity in U266 cells. U266 cells were cultured for 24 h with indicated concentrations of Bortezomib (A), Melphalan (B), and Doxorubicin (C) [control media (□), 1.25 nmol/l Bortezomib (&bsl00001;), 2.5 nmol/l Bortezomib (&bsl00001;), 3.75 nmol/l Bortezomib (&bsl00001;); control media (□), 2.5 &mgr;mol/l Melphalan (&bsl00001;), 5 &mgr;mol/l Melphalan (&bsl00001;), 10 &mgr;mol/l Melphalan (&bsl00001;); control media (□), 100 nmol/l Doxorubicin (&bsl00001;), 200 nmol/l Doxorubicin (&bsl00001;), 400 nmol/l Doxorubicin (&bsl00001;)], in the presence or absence of Cerulenin (0–25 &mgr;mol/l). Cell growth was assessed by MTT assays. Data represent mean (±SD) of triplicate cultures. Combination index (CI) of Cerulenin and these agents was analysed using CalcuSyn software.
Please note: Blackwell Publishing are not responsible for the content or functionality of any supplementary materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
Supporting info item
Supporting info item
Supporting info item
Supporting info item
Acknowledgements
This study was supported by National Institutes of Health Specialized Programs of Research Excellence (SPORE) IP50 CA10070‐01, PO‐1 CA78378, and RO‐1 CA50947 grants; the Doris Duke Distinguished Clinical Research Scientist Award (KCA); the Multiple Myeloma Research Foundation (TH, DC, KP, TY); and the LeBow Fund to Cure Myeloma Fund (KCA).
Re‐use of this article is permitted in accordance with the Creative Commons Deed, Attribution 2.5, which does not permit commercial exploitation.
References
- Abraham, M.C. & Shaham, S. (2004) Death without caspases, caspases without death. Trends in Cell Biology, 14, 184–193. [DOI] [PubMed] [Google Scholar]
- Ahn, E.Y. , Lim, S.T. , Cook, W.J. & McDonald, J.M. (2004) Calmodulin binding to the Fas death domain. Regulation by Fas activation. Journal of Biological Chemistry, 279, 5661–5666. [DOI] [PubMed] [Google Scholar]
- Asturias, F.J. , Chadick, J.Z. , Cheung, I.K. , Stark, H. , Witkowski, A. , Joshi, A.K. & Smith, S. (2005) Structure and molecular organization of mammalian fatty acid synthase. Nature Structural & Molecular Biology, 12, 225–232. [DOI] [PubMed] [Google Scholar]
- Bandyopadhyay, S. , Zhan, R. , Wang, Y. , Pai, S.K. , Hirota, S. , Hosobe, S. , Takano, Y. , Saito, K. , Furuta, E. , Iiizumi, M. , Mohinta, S. , Watabe, M. , Chalfant, C. & Watabe, K. (2006) Mechanism of apoptosis induced by the inhibition of fatty acid synthase in breast cancer cells. Cancer Research, 66, 5934–5940. [DOI] [PubMed] [Google Scholar]
- Baron, A. , Migita, T. , Tang, D. & Loda, M. (2004) Fatty acid synthase: a metabolic oncogene in prostate cancer? Journal of Cellular Biochemistry, 91, 47–53. [DOI] [PubMed] [Google Scholar]
- Bialik, S. & Kimchi, A. (2004) DAP‐kinase as a target for drug design in cancer and diseases associated with accelerated cell death. Seminars in Cancer Biology, 14, 283–294. [DOI] [PubMed] [Google Scholar]
- Borsello, T. , Croquelois, K. , Hornung, J.P. & Clarke, P.G. (2003) N‐methyl‐d‐aspartate‐triggered neuronal death in organotypic hippocampal cultures is endocytic, autophagic and mediated by the c‐Jun N‐terminal kinase pathway. European Journal of Neuroscience, 18, 473–485. [DOI] [PubMed] [Google Scholar]
- Bossy‐Wetzel, E. , Bakiri, L. & Yaniv, M. (1997) Induction of apoptosis by the transcription factor c‐Jun. EMBO Journal, 16, 1695–1709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cande, C. , Vahsen, N. , Garrido, C. & Kroemer, G. (2004) Apoptosis‐inducing factor (AIF): caspase‐independent after all. Cell Death and Differentiation, 11, 591–595. [DOI] [PubMed] [Google Scholar]
- Chou, T.C. & Talalay, P. (1984) Quantitative analysis of dose‐effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Advances in Enzyme Regulation, 22, 27–55. [DOI] [PubMed] [Google Scholar]
- Cox, J.S. , Chapman, R.E. & Walter, P. (1997) The unfolded protein response coordinates the production of endoplasmic reticulum protein and endoplasmic reticulum membrane. Molecular Biology of the Cell, 8, 1805–1814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cregan, S.P. , Dawson, V.L. & Slack, R.S. (2004) Role of AIF in caspase‐dependent and caspase‐independent cell death. Oncogene, 23, 2785–2796. [DOI] [PubMed] [Google Scholar]
- Daido, S. , Kanzawa, T. , Yamamoto, A. , Takeuchi, H. , Kondo, Y. & Kondo, S. (2004) Pivotal role of the cell death factor BNIP3 in ceramide‐induced autophagic cell death in malignant glioma cells. Cancer Research, 64, 4286–4293. [DOI] [PubMed] [Google Scholar]
- Damiano, J.S. , Cress, A.E. , Hazlehurst, L.A. , Shtil, A.A. & Dalton, W.S. (1999) Cell adhesion mediated drug resistance (CAM‐DR): role of integrins and resistance to apoptosis in human myeloma cell lines. Blood, 93, 1658–1667. [PMC free article] [PubMed] [Google Scholar]
- Daugas, E. , Susin, S.A. , Zamzami, N. , Ferri, K.F. , Irinopoulou, T. , Larochette, N. , Prevost, M.C. , Leber, B. , Andrews, D. , Penninger, J. & Kroemer, G. (2000) Mitochondrio‐nuclear translocation of AIF in apoptosis and necrosis. FASEB Journal, 14, 729–739. [PubMed] [Google Scholar]
- Davis, R.J. (2000) Signal transduction by the JNK group of MAP kinases. Cell, 103, 239–252. [DOI] [PubMed] [Google Scholar]
- De Schrijver, E. , Brusselmans, K. , Heyns, W. , Verhoeven, G. & Swinnen, J.V. (2003) RNA interference‐mediated silencing of the fatty acid synthase gene attenuates growth and induces morphological changes and apoptosis of LNCaP prostate cancer cells. Cancer Research, 63, 3799–3804. [PubMed] [Google Scholar]
- Gabrielson, E.W. , Pinn, M.L. , Testa, J.R. & Kuhajda, F.P. (2001) Increased fatty acid synthase is a therapeutic target in mesothelioma. Clinical Cancer Research, 7, 153–157. [PubMed] [Google Scholar]
- Hayashi, T. , Hideshima, T. , Akiyama, M. , Richardson, P. , Schlossman, R.L. , Chauhan, D. , Munshi, N.C. , Waxman, S. & Anderson, K.C. (2002) Arsenic trioxide inhibits growth of human multiple myeloma cells in the bone marrow microenvironment. Molecular Cancer Therapeutics, 1, 851–860. [PubMed] [Google Scholar]
- Hazlehurst, L.A. , Damiano, J.S. , Buyuksal, I. , Pledger, W.J. & Dalton, W.S. (2000) Adhesion to fibronectin via beta1 integrins regulates p27kip1 levels and contributes to cell adhesion mediated drug resistance (CAM‐DR). Oncogene, 19, 4319–4327. [DOI] [PubMed] [Google Scholar]
- Heiligtag, S.J. , Bredehorst, R. & David, K.A. (2002) Key role of mitochondria in cerulenin‐mediated apoptosis. Cell Death and Differentiation, 9, 1017–1025. [DOI] [PubMed] [Google Scholar]
- Hideshima, T. , Chauhan, D. , Shima, Y. , Raje, N. , Davies, F.E. , Tai, Y.T. , Treon, S.P. , Lin, B. , Schlossman, R.L. , Richardson, P. , Muller, G. , Stirling, D.I. & Anderson, K.C. (2000) Thalidomide and its analogs overcome drug resistance of human multiple myeloma cells to conventional therapy. Blood, 96, 2943–2950. [PubMed] [Google Scholar]
- Hideshima, T. , Nakamura, N. , Chauhan, D. & Anderson, K.C. (2001) Biologic sequelae of interleukin‐6 induced PI3‐K/Akt signaling in multiple myeloma. Oncogene, 20, 5991–6000. [DOI] [PubMed] [Google Scholar]
- Hideshima, T. , Bergsagel, P.L. , Kuehl, W.M. & Anderson, K.C. (2004) Advances in biology of multiple myeloma: clinical applications. Blood, 104, 607–618. [DOI] [PubMed] [Google Scholar]
- Hideshima, T. , Mitsiades, C. , Tonon, G. , Richardson, P.G. & Anderson, K.C. (2007) Understanding multiple myeloma pathogenesis in the bone marrow to identify new therapeutic targets. Nature Reviews Cancer, 7, 585–598. [DOI] [PubMed] [Google Scholar]
- Inbal, B. , Bialik, S. , Sabanay, I. , Shani, G. & Kimchi, A. (2002) DAP kinase and DRP‐1 mediate membrane blebbing and the formation of autophagic vesicles during programmed cell death. Journal of Cell Biology, 157, 455–468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishitsuka, K. , Hideshima, T. , Hamasaki, M. , Raje, N. , Kumar, S. , Podar, K. , Le Gouill, S. , Shiraishi, N. , Yasui, H. , Roccaro, A.M. , Tai, Y.Z. , Chauhan, D. , Fram, R. , Tamura, K. , Jain, J. & Anderson, K.C. (2005) Novel inosine monophosphate dehydrogenase inhibitor VX‐944 induces apoptosis in multiple myeloma cells primarily via caspase‐independent AIF/Endo G pathway. Oncogene, 24, 5888–5896. [DOI] [PubMed] [Google Scholar]
- Jaattela, M. & Tschopp, J. (2003) Caspase‐independent cell death in T lymphocytes. Nature Immunology, 4, 416–423. [DOI] [PubMed] [Google Scholar]
- Jia, G. , Cheng, G. , Gangahar, D.M. & Agrawal, D.K. (2006) Insulin‐like growth factor‐1 and TNF‐alpha regulate autophagy through c‐jun N‐terminal kinase and Akt pathways in human atherosclerotic vascular smooth cells. Immunology and Cell Biology, 84, 448–454. [DOI] [PubMed] [Google Scholar]
- Jochum, W. , Passegue, E. & Wagner, E.F. (2001) AP‐1 in mouse development and tumorigenesis. Oncogene, 20, 2401–2412. [DOI] [PubMed] [Google Scholar]
- Jourdan, M. , De Vos, J. , Mechti, N. & Klein, B. (2000) Regulation of Bcl‐2‐family proteins in myeloma cells by three myeloma survival factors: interleukin‐6, interferon‐alpha and insulin‐like growth factor 1. Cell Death and Differentiation, 7, 1244–1252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufman, R.J. (2004) Regulation of mRNA translation by protein folding in the endoplasmic reticulum. Trends in Biochemical Sciences, 29, 152–158. [DOI] [PubMed] [Google Scholar]
- Kelley, S.K. & Ashkenazi, A. (2004) Targeting death receptors in cancer with Apo2L/TRAIL. Current Opinion in Pharmacology, 4, 333–339. [DOI] [PubMed] [Google Scholar]
- Kridel, S.J. , Axelrod, F. , Rozenkrantz, N. & Smith, J.W. (2004) Orlistat is a novel inhibitor of fatty acid synthase with antitumor activity. Cancer Research, 64, 2070–2075. [DOI] [PubMed] [Google Scholar]
- Kuhajda, F.P. (2000) Fatty‐acid synthase and human cancer: new perspectives on its role in tumor biology. Nutrition, 16, 202–208. [DOI] [PubMed] [Google Scholar]
- Kuhajda, F.P. (2006) Fatty acid synthase and cancer: new application of an old pathway. Cancer Research, 66, 5977–5980. [DOI] [PubMed] [Google Scholar]
- Kuhajda, F.P. , Pizer, E.S. , Li, J.N. , Mani, N.S. , Frehywot, G.L. & Townsend, C.A. (2000) Synthesis and antitumor activity of an inhibitor of fatty acid synthase. Proceedings of the National Academy of Sciences of the United States of America, 97, 3450–3454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyle, R.A. & Rajkumar, S.V. (2004) Multiple myeloma. New England Journal of Medicine, 351, 1860–1873. [DOI] [PubMed] [Google Scholar]
- Li, J. , Lee, B. & Lee, A.S. (2006) Endoplasmic reticulum stress‐induced apoptosis: multiple pathways and activation of p53‐up‐regulated modulator of apoptosis (PUMA) and NOXA by p53. Journal of Biological Chemistry, 281, 7260–7270. [DOI] [PubMed] [Google Scholar]
- Little, J.L. , Wheeler, F.B. , Fels, D.R. , Koumenis, C. & Kridel, S.J. (2007) Inhibition of fatty acid synthase induces endoplasmic reticulum stress in tumor cells. Cancer Research, 67, 1262–1269. [DOI] [PubMed] [Google Scholar]
- Lockshin, R.A. & Zakeri, Z. (2004) Caspase‐independent cell death? Oncogene, 23, 2766–2773. [DOI] [PubMed] [Google Scholar]
- McCafferty‐Grad, J. , Bahlis, N.J. , Krett, N. , Aguilar, T.M. , Reis, I. , Lee, K.P. & Boise, L.H. (2003) Arsenic trioxide uses caspase‐dependent and caspase‐independent death pathways in myeloma cells. Molecular Cancer Therapeutics, 2, 1155–1164. [PubMed] [Google Scholar]
- Menendez, J.A. & Lupu, R. (2007) Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nature Reviews Cancer, 7, 763–777. [DOI] [PubMed] [Google Scholar]
- Milgraum, L.Z. , Witters, L.A. , Pasternack, G.R. & Kuhajda, F.P. (1997) Enzymes of the fatty acid synthesis pathway are highly expressed in in situ breast carcinoma. Clinical Cancer Research, 3, 2115–2120. [PubMed] [Google Scholar]
- Mitsiades, C.S. , Mitsiades, N. , Poulaki, V. , Schlossman, R. , Akiyama, M. , Chauhan, D. , Hideshima, T. , Treon, S.P. , Munshi, N.C. , Richardson, P.G. & Anderson, K.C. (2002) Activation of NF‐kappaB and upregulation of intracellular anti‐apoptotic proteins via the IGF‐1/Akt signaling in human multiple myeloma cells: therapeutic implications. Oncogene, 21, 5673–5683. [DOI] [PubMed] [Google Scholar]
- Mitsiades, C.S. , Mitsiades, N.S. , McMullan, C.J. , Poulaki, V. , Shringarpure, R. , Akiyama, M. , Hideshima, T. , Chauhan, D. , Joseph, M. , Libermann, T.A. , Garcia‐Echeverria, C. , Pearson, M.A. , Hofmann, F. , Anderson, K.C. & Kung, A.L. (2004) Inhibition of the insulin‐like growth factor receptor‐1 tyrosine kinase activity as a therapeutic strategy for multiple myeloma, other hematologic malignancies, and solid tumors. Cancer Cell, 5, 221–230. [DOI] [PubMed] [Google Scholar]
- Moche, M. , Schneider, G. , Edwards, P. , Dehesh, K. & Lindqvist, Y. (1999) Structure of the complex between the antibiotic cerulenin and its target, beta‐ketoacyl‐acyl carrier protein synthase. Journal of Biological Chemistry, 274, 6031–6034. [DOI] [PubMed] [Google Scholar]
- Nakamura, M. , Gotoh, T. , Okuno, Y. , Tatetsu, H. , Sonoki, T. , Uneda, S. , Mori, M. , Mitsuya, H. & Hata, H. (2006) Activation of the endoplasmic reticulum stress pathway is associated with survival of myeloma cells. Leukemia & Lymphoma, 47, 531–539. [DOI] [PubMed] [Google Scholar]
- Omura, S. (1976) The antibiotic cerulenin, a novel tool for biochemistry as an inhibitor of fatty acid synthesis. Bacteriological Reviews, 40, 681–697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park, K.J. , Lee, S.H. , Kim, T.I. , Lee, H.W. , Lee, C.H. , Kim, E.H. , Jang, J.Y. , Choi, K.S. , Kwon, M.H. & Kim, Y.S. (2007) A human scFv antibody against TRAIL receptor 2 induces autophagic cell death in both TRAIL‐sensitive and TRAIL‐resistant cancer cells. Cancer Research, 67, 7327–7334. [DOI] [PubMed] [Google Scholar]
- Pelled, D. , Raveh, T. , Riebeling, C. , Fridkin, M. , Berissi, H. , Futerman, A.H. & Kimchi, A. (2002) Death‐associated protein (DAP) kinase plays a central role in ceramide‐induced apoptosis in cultured hippocampal neurons. Journal of Biological Chemistry, 277, 1957–1961. [DOI] [PubMed] [Google Scholar]
- Pizer, E.S. , Jackisch, C. , Wood, F.D. , Pasternack, G.R. , Davidson, N.E. & Kuhajda, F.P. (1996a) Inhibition of fatty acid synthesis induces programmed cell death in human breast cancer cells. Cancer Research, 56, 2745–2747. [PubMed] [Google Scholar]
- Pizer, E.S. , Wood, F.D. , Pasternack, G.R. & Kuhajda, F.P. (1996b) Fatty acid synthase (FAS): a target for cytotoxic antimetabolites in HL60 promyelocytic leukemia cells. Cancer Research, 56, 745–751. [PubMed] [Google Scholar]
- Pizer, E.S. , Pflug, B.R. , Bova, G.S. , Han, W.F. , Udan, M.S. & Nelson, J.B. (2001) Increased fatty acid synthase as a therapeutic target in androgen‐independent prostate cancer progression. Prostate, 47, 102–110. [DOI] [PubMed] [Google Scholar]
- Podar, K. , Raab, M.S. , Tonon, G. , Sattler, M. , Barila, D. , Zhang, J. , Tai, Y.T. , Yasui, H. , Raje, N. , DePinho, R.A. , Hideshima, T. , Chauhan, D. & Anderson, K.C. (2007) Up‐regulation of c‐Jun inhibits proliferation and induces apoptosis via caspase‐triggered c‐Abl cleavage in human multiple myeloma. Cancer Research, 67, 1680–1688. [DOI] [PubMed] [Google Scholar]
- Raje, N. , Kumar, S. , Hideshima, T. , Ishitsuka, K. , Chauhan, D. , Mitsiades, C. , Podar, K. , Le Gouill, S. , Richardson, P. , Munshi, N.C. , Stirling, D.I. , Antin, J.H. & Anderson, K.C. (2004) Combination of the mTOR inhibitor rapamycin and CC‐5013 has synergistic activity in multiple myeloma. Blood, 104, 4188–4193. [DOI] [PubMed] [Google Scholar]
- Richardson, P. , Hideshima, T. & Anderson, K.C. (2004) An update of novel therapeutic approaches for multiple myeloma. Current Treatment Options in Oncology, 5, 227–238. [DOI] [PubMed] [Google Scholar]
- Rossi, S. , Graner, E. , Febbo, P. , Weinstein, L. , Bhattacharya, N. , Onody, T. , Bubley, G. , Balk, S. & Loda, M. (2003) Fatty acid synthase expression defines distinct molecular signatures in prostate cancer. Molecular Cancer Research, 1, 707–715. [PubMed] [Google Scholar]
- Van Der Sanden, M.H. , Meems, H. , Houweling, M. , Helms, J.B. & Vaandrager, A.B. (2004) Induction of CCAAT/enhancer‐binding protein (C/EBP)‐homologous protein/growth arrest and DNA damage‐inducible protein 153 expression during inhibition of phosphatidylcholine synthesis is mediated via activation of a C/EBP‐activating transcription factor‐responsive element. Journal of Biological Chemistry, 279, 52007–52015. [DOI] [PubMed] [Google Scholar]
- Sriburi, R. , Jackowski, S. , Mori, K. & Brewer, J.W. (2004) XBP1: a link between the unfolded protein response, lipid biosynthesis, and biogenesis of the endoplasmic reticulum. Journal of Cell Biology, 167, 35–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swinnen, J.V. , Van Veldhoven, P.P. , Timmermans, L. , De Schrijver, E. , Brusselmans, K. , Vanderhoydonc, F. , Van de Sande, T. , Heemers, H. , Heyns, W. & Verhoeven, G. (2003) Fatty acid synthase drives the synthesis of phospholipids partitioning into detergent‐resistant membrane microdomains. Biochemical and Biophysical Research Communications, 302, 898–903. [DOI] [PubMed] [Google Scholar]
- Szegezdi, E. , Logue, S.E. , Gorman, A.M. & Samali, A. (2006) Mediators of endoplasmic reticulum stress‐induced apoptosis. EMBO Reports, 7, 880–885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tessitore, A. , Del, P.M.M. , Sano, R. , Ma, Y. , Mann, L. , Ingrassia, A. , Laywell, E.D. , Steindler, D.A. , Hendershot, L.M. & D’Azzo, A. (2004) GM1‐ganglioside‐mediated activation of the unfolded protein response causes neuronal death in a neurodegenerative gangliosidosis. Molecular Cell, 15, 753–766. [DOI] [PubMed] [Google Scholar]
- Urano, F. , Wang, X. , Bertolotti, A. , Zhang, Y. , Chung, P. , Harding, H.P. & Ron, D. (2000) Coupling of stress in the ER to activation of JNK protein kinases by transmembrane protein kinase IRE1. Science, 287, 664–666. [DOI] [PubMed] [Google Scholar]
- Vance, D. , Goldberg, I. , Mitsuhashi, O. & Bloch, K. (1972) Inhibition of fatty acid synthetases by the antibiotic cerulenin. Biochemical and Biophysical Research Communications, 48, 649–656. [DOI] [PubMed] [Google Scholar]
- Vande Velde, C. , Cizeau, J. , Dubik, D. , Alimonti, J. , Brown, T. , Israels, S. , Hakem, R. & Greenberg, A.H. (2000) BNIP3 and genetic control of necrosis‐like cell death through the mitochondrial permeability transition pore. Molecular and Cellular Biology, 20, 5454–5468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogt, P.K. (2001) Jun, the oncoprotein. Oncogene, 20, 2365–2377. [DOI] [PubMed] [Google Scholar]
- Wakil, S.J. (1989) Fatty acid synthase, a proficient multifunctional enzyme. Biochemistry, 28, 4523–4530. [DOI] [PubMed] [Google Scholar]
- Weston, C.R. & Davis, R.J. (2007) The JNK signal transduction pathway. Current Opinion in Cell Biology, 19, 142–149. [DOI] [PubMed] [Google Scholar]
- Xu, C. , Bailly‐Maitre, B. & Reed, J.C. (2005) Endoplasmic reticulum stress: cell life and death decisions. Journal of Clinical Investigation, 115, 2656–2664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto, M. , Hioki, T. , Ishii, T. , Nakajima‐Iijima, S. & Uchino, S. (2002) DAP kinase activity is critical for C(2)‐ceramide‐induced apoptosis in PC12 cells. European Journal of Biochemistry, 269, 139–147. [DOI] [PubMed] [Google Scholar]
- Yu, L. , Alva, A. , Su, H. , Dutt, P. , Freundt, E. , Welsh, S. , Baehrecke, E.H. & Lenardo, M.J. (2004) Regulation of an ATG7‐beclin 1 program of autophagic cell death by caspase‐8. Science, 304, 1500–1502. [DOI] [PubMed] [Google Scholar]
- Zhao, L. & Ackerman, S.L. (2006) Endoplasmic reticulum stress in health and disease. Current Opinion in Cell Biology, 18, 444–452. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig S1. Cerulenin induces apoptosis in U266 cells. (A) U266 cells were cultured with Cerulenin (50 &mgr;mol/l) for the indicated times, with or without Z‐VAD‐FMK (50 &mgr;mol/l) for 1 h pretreatment. Total cell lysates (20 &mgr;g/lane) were subjected to Western blotting using anti‐caspase ‐8, ‐9, ‐3, PARP and &alpha‐tubulin. FL, CF indicated as full length and cleaved form, respectively. (B, C) U266 cells were treated with indicated dose of Cerulenin for 24 h, with or without Z‐VAD‐FMK (25 or 50 mmol/l) 1 h pretreatment. Cytotoxicity was determined by MTT assay (B). Values represent mean ± SD of quadruplicate cultures. The percentage apoptotic cells was determined by flow‐cytometric analysis for APO2.7 staining (C).
Fig S2. JNK inhibitor blocks Cerulenin‐induced cytotoxicity in U266 cells. (A, B) U266 cells were cultured with indicated dose of Cerulenin for 24 h, with or without JNK inhibitor SP600215 (5 or 10 &mgr;mol/l) pretreatment for 1 h. Cytotoxicity was determined by MTT assay (A). Values represent mean ± SD of quadruplicate cultures. JNK inhibitor significantly blocks Cerulenin‐induced cytotoxicity (P < 0.05). The percentage apoptotic cells was determined by flow‐cytometric analysis for APO2.7 staining (B). (C) Total cell lysates (20 mg/lane) were subjected to Western blotting using anti‐caspase ‐8, ‐9, ‐3, PARP and α‐tubulin. FL, CF indicated as full length and cleaved form, respectively.
Fig S3. Analysis of inhibition by JNK inhibitor SP600215 and/or ZVAD‐FMK in MM cells. (A, B) MM.1S and U266 cells were cultured with indicated dose of Cerulenin for 24 h, with or without JNK inhibitor SP600215 (10 &mgr;mol/l) and/or ZVAD‐FMK (50 &mgr;mol/l) pretreatment for 1 h. Cytotoxicity was determined by MTT assay. Values represent mean ± SD of quadruplicate cultures. Although ZVAD‐FMK does not affect cytotoxicity by Cerulenin, JNK inhibitor significantly blocks Cerulenin‐induced cytotoxicity (P < 0.05).
Fig S4. Cerulenin enhances Bortezomib, Melphalan, and Doxorubicin‐induced cytotoxicity in U266 cells. U266 cells were cultured for 24 h with indicated concentrations of Bortezomib (A), Melphalan (B), and Doxorubicin (C) [control media (□), 1.25 nmol/l Bortezomib (&bsl00001;), 2.5 nmol/l Bortezomib (&bsl00001;), 3.75 nmol/l Bortezomib (&bsl00001;); control media (□), 2.5 &mgr;mol/l Melphalan (&bsl00001;), 5 &mgr;mol/l Melphalan (&bsl00001;), 10 &mgr;mol/l Melphalan (&bsl00001;); control media (□), 100 nmol/l Doxorubicin (&bsl00001;), 200 nmol/l Doxorubicin (&bsl00001;), 400 nmol/l Doxorubicin (&bsl00001;)], in the presence or absence of Cerulenin (0–25 &mgr;mol/l). Cell growth was assessed by MTT assays. Data represent mean (±SD) of triplicate cultures. Combination index (CI) of Cerulenin and these agents was analysed using CalcuSyn software.
Please note: Blackwell Publishing are not responsible for the content or functionality of any supplementary materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
Supporting info item
Supporting info item
Supporting info item
Supporting info item