

Abstract
From pharmacological studies using histamine antagonists and agonists, it has been demonstrated that histamine modulates many physiological functions of the hypothalamus, such as arousal state, locomotor activity, feeding, and drinking. Three kinds of receptors (H1, H2, and H3) mediate these actions. To define the contribution of the histamine H1 receptors (H1R) to behavior, mutant mice lacking the H1R were generated by homologous recombination. In brains of homozygous mutant mice, no specific binding of [3H]pyrilamine was seen. [3H]Doxepin has two saturable binding sites with higher and lower affinities in brains of wild-type mice, but H1R-deficient mice showed only the weak labeling of [3H]doxepin that corresponds to lower-affinity binding sites. Mutant mice develop normally, but absence of H1R significantly increased the ratio of ambulation during the light period to the total ambulation for 24 hr in an accustomed environment. In addition, mutant mice significantly reduced exploratory behavior of ambulation and rearings in a new environment. These results indicate that through H1R, histamine is involved in circadian rhythm of locomotor activity and exploratory behavior as a neurotransmitter.
Biological activity of histamine has been characterized mainly from pharmacological methods using its antagonists and agonists. These studies have demonstrated that histamine plays an important role in various physiological reactions such as inflammation and gastric acid secretion. In the mammalian central nervous system (CNS), histamine is thought to modulate many functions of the hypothalamus that exhibit circadian and other rhythms, such as arousal state, locomotor activity, feeding, and drinking (1–4). The histaminergic neuron system arises from the tuberomammillary nucleus of the posterior hypothalamus receiving inputs mainly from the limbic system and projects efferent nerve fibers to almost all parts of the brain (5–9). Autoradiographic binding studies show that three types of receptors are widely expressed in brain, not only on neurons but also on astrocytes and blood vessels (10, 11). However, much remains to be cleared about its roles in the CNS. To study the function of histamine neuron system, histamine H1 receptor (H1R)-deficient mice were generated by homologous recombination. Considering the unique distribution pattern of the histamine neuron system, circadian rhythm, and exploratory behavior in a new environment were studied here in mice lacking H1R.
MATERIALS AND METHODS
Construction of Targeting Vector.
The H1R gene was isolated from a mouse 129/ola-embryonic stem (ES)/EMBLE3 genomic library. A BglII–EcoRI 6.4-kb fragment containing DNA from mostly 3′ of the H1R gene was subcloned into PUC19 at the sites of BamHI and EcoRI. A 0.8-kb ApaI–EcoRI fragment containing 5′ upstream sequences was subcloned into the BamHI site of pMC1 poly(A) (Stratagene) in blunted end ligation. The 1.9-kb fragment with 5′ upstream sequence and Neo-poly(A) cassette including promoter (excised with SalI and XhoI from the vector) was integrated at the site of SalI into the PUC19 with 6.4-kb fragment.
Gene Targeting and Generation of Mutant Mice.
The vector was linearized at a unique SalI site of the plasmid. Linearized targeting vector was electroporated in E14 ES cells, and neor colonies were selected as previously described (12). Homologous recombinants were screened at first by PCR with a primer located upstream of the short arm of the homology (5′-GAAGTATCTGGCTCTGAGTG-3′) and a primer complementary to the neor gene sequence (5′-GCGTTGGCTACCCGTGATAT-3′) and verified by Southern blotting. Genomic DNA from PCR positive clones was digested with EcoRI and hybridized with probe A, a neo probe (probe B), and probe D. ES cells carrying the H1R mutation were injected into blastocysts from C57BL/6 mice and the resulting male chimeras were mated with C57BL/6 mice as described (12). Agouti offsprings were analyzed by Southern blotting of genomic DNAs from tail biopsies for the presence of the H1R mutant allele.
Autoradiography and Ligand Binding Assay.
Wild-type (wt), heterozygous, and homozygous mice for the mutant H1R gene were decapitated after ether anesthesia. These brains were rapidly removed from skulls. Cryostat sections (20 μm thick) were prepared and incubated in 50 mM Na/K phosphate (pH 7.5) buffer containing 1.0 nM [3H]pyrilamine as described (13). Analysis was performed after 7 days exposure by using tritium-sensitive imaging plates (Fujix Bioimaging analyzer BAS3000; Fuji). H1R ligand-binding assays were carried out for forebrain tissue homogenized with a polytron in ice-cold 50 mM Na/K phosphate buffer, pH 7.5. The homogenates were centrifuged twice at 50,000 × g for 20 min at 4°C, and then 25 μg of forebrain membrane was used for ligand-binding assay with [3H]pyrilamine (25°C) (14) and [3H]doxepin (37°C) (15–17). Concentrations of [3H]pyrilamine and [3H]doxepin of 0.025–9.5 nM were used for Scatchard analysis. [3H]Doxepin was synthesized by the reaction of [3H]methyliodide (specific activity, 51 Ci nmol−1; 1 Ci = 37 GBq; Amersham) with the desmethylated precursor of doxepin [(E)-isomer, 99.7%] as described (18). Nonspecific binding was determined in the presence of 2 μM triprolidine, and binding data were analyzed with ligand program.
Analysis of Locomotor Activity in an Accustomed Environment.
Spontaneous locomotor activity of mice was measured every hour for 2 days using an infrared ray passive sensor system (Neuroscience, Tokyo). An apparatus with the infrared beam-sensor was set on the top of conventional polypropylene cage and number of movements was counted and relayed by an interface to a computer. Inexperienced mice of 9–11 weeks old were used for the experiments. They were housed in standard cages and kept on a 12-hr light/12-hr dark cycle with light onset at 8:00 a.m. Data obtained for last 24 hr of each experiment were analyzed by the Mann–Whitney U test for statistical significance between groups (wt: male = 4, female = 4; mutant mice: male = 5, female = 3; data between males and females in each group were not significantly different.)
Analysis of Exploratory Behavior.
H1R-deficient mice or wt mice were separately reared in groups of 6–8 male mice at least for 3 weeks. Mice 12–15 weeks of age were used for the experiments. Each mouse was placed in the open field for 30 min from 10:00 a.m. to 3:00 p.m. Total distance of ambulation and time of rearings for 3 min were measured in a square area (60 cm × 60 cm) for 30 min by a photo-beam apparatus (BTA-1, Muromachikikai, Tokyo) that monitored movements linked to a computer (n = 23 and 30 for wt and homozygous mice, respectively). Distance was calculated from the trace of animal by the computer. Statistical analysis was performed by the Mann–Whitney U test.
RESULTS
Establishment of H1R-Deficient Mice.
To disrupt the H1R gene that is intronless, the H1R targeting vector was constructed as shown in Fig. 1a. The EcoRI–BglII 0.8-kb fragment containing the initial translation codon was replaced with the neor gene in reverse orientation. This vector was transfected into E14 ES cells by electroporation and four clones containing the desired homologous recombination were identified among 542 G418-resistant clones by Southern blot hybridization. These were injected into blastocysts to produce chimeric animals. Chimeric males were mated to C57 BL/6 females. Some chimeric mice from two independent clones transmitted the mutation through the germ line. Heterozygote progenies were intercrossed, and homozygous (−/−), heterozygous (+/−), and wt (+/+) mice were produced (Fig. 1b). They were derived from totally independent several homozygous crossings. The H1R gene disrupted mice appeared to develop normally and be apparently healthy and were fertile, indicating that there were little or no effects of H1R on development of mice. Histochemical analysis of H1R mutant mice did not reveal any gross anatomical abnormalities in brain, heart, lung, liver, spleen, and kidney (data not shown).
Figure 1.
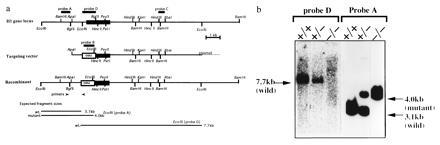
Disruption of the H1R gene in mouse ES cells and generation of mutant mice. (a) Schematic diagram of the strategy used to target the H1R gene. Top line, genomic structure of the H1R gene in wt. The closed box represents the entire coding region of H1R gene. Middle line represents the targeting plasmid vector linearized at a unique SalI site. Neomycin-resistant (neor) gene with polyadenylylation signals driven by thymidine kinase promoter is shown by open box. The transcriptional orientation of the neor gene is opposite to the H1R gene. Bottom line represents the structure of the inactivated H1R gene after a correct homologous recombination. (b) Southern blots of tail DNA from the offspring of germ-line chimeras generated by blastocyst injection with the targeted ES cells. Tail DNA was extracted and digested with EcoRI and hybridized with probe A. Three genotypes, wt (+/+), heterozygote (+/−), and homozygote (−/−), are shown. Rehybridization of the same filter with a probe D confirmed that the 5′ half of H1R gene was absent from the genomic DNA of homozygous mice.
Reevaluation of Receptor-Binding Studies Using [3H]Pyrilamine and [3H]Doxepin in the Brain.
To verify the absence of H1R, autoradiography of the brains from wt, heterozygous, and homozygous mutant mice was performed with 3H-labeled antihistamine, [3H]pyrilamine (or mepyramine). In wt mice, the binding of [3H]pyrilamine was observed in hypothalamus, cerebral cortex, amygdala, thalamus, hippocampus, and cerebellum of the brain section (Fig. 2 a and b), whereas no specific binding was seen in homozygous mutant mice (Fig. 2 e and f). The level of H1R-binding in homozygous mutant mice is almost equal to that of nonspecific binding defined by 2 μM triploridine. The mice homozygous for H1R gene were identified as lacking H1R completely. In receptor binding assay with [3H]pyrilamine using homogenates of brains from heterozygotes, the amounts of H1R protein were found to be about half as much as that of wt mice as shown in Fig. 2g [H1R-binding = 294.9 ± 22.1 (+/+), 146.7 ± 11.2 (+/−), and 7.8 ± 6.9 (−/−) fmol/mg protein/nM]. The Scatchard analysis of the saturation isotherm for [3H]pyrilamine binding to brain of wt and mutant mice showed that pyrilamine binds specifically to the H1R with a single binding site and that H1R-binding sites were completely absent in the homozygous mice (Fig. 2h). This indicates that the H1R is the only binding protein for pyrilamine in brain, although the possibility of existence of subclass in H1R was suggested previously (19). Doxepin, a tricyclic antidepressant, is a potent histamine antagonist and an alternative choice as a ligand (15–17). [3H]doxepin has two saturable binding sites with higher and lower affinities in brains of wt mice (Fig. 2i). Only the weak labeling of [3H]doxepin was observed in the H1R-deficient homozygotes, which corresponds to lower-affinity binding sites (Fig. 2i). These data clearly demonstrate that the high affinity component of doxepin binding is associated with H1R and that the low-affinity component is distinct from that of H1R.
Figure 2.
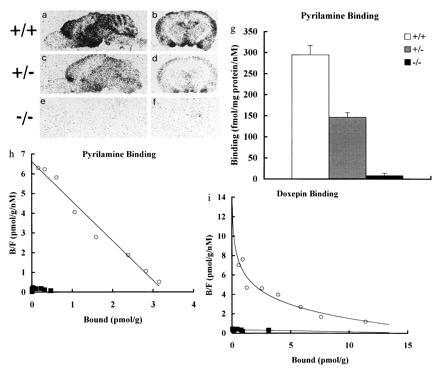
Localization of H1R in brain of wt, heterozygous and homozygous mice. Autoradiographic localization of [3H]pyrilamine (specific activity: 27 Ci mmol−1) binding in the mouse brain. [3H]Pyrilamine, a specific H1R antagonist, was used to label H1R in vitro on saggital (a, c, and e) and coronal sections (b, d, and f) of mouse brain. (a and b) wt mice (+/+); (c and d) heterozygous mice (+/−); (e and f) homozygous mice (−/−); and (g) H1R binding with 1.0 nM [3H]pyrilamine on homogenates of forebrain. Data are expressed as means ± SD of 6–12 mice in each group. (h and i) Scatchard analysis of the saturation isotherm for [3H]pyrilamine (h) and [3H]doxepin (i) binding to forebrain membranes from +/+ (○), and −/− (▪). The results shown are representative of three independent experiments each conducted in duplicate. The maximum binding capacity for pyrilamine were 3.3 ± 0.6 pmol/g of brain tissue for +/+ (○), and inestimable for −/− (▪). The dissociation constants for pyrilamine were 0.65 ± 0.25 nM for +/+ (○), and inestimable for −/− (▪). Scatchard analysis of doxepin binding in wt mice, Kd (high) = 0.15 ± 0.05 nM and Bmax (high) = 2.0 ± 0.5 pmol/g; Kd (low) = 33 ± 13 nM and Bmax (low) = 25 ± 8.0 pmol/g: Scatchard analysis in homozygous mice, Kd = 58 ± 24 nM and Bmax = 31 ± 24 pmol/g.
Role of H1R in the Control of Circadian Rhythm.
To determine the role of H1R in circadian rhythm of ambulation, locomotor activity was measured in an accustomed environment every hr for 24 hr using a photosensor system. During the experiment, mice were housed alone in an accustomed environment with a 12-hr light:12-hr dark cycle. Measurement of locomotion revealed that the absence of H1R disturbed the circadian rhythm of locomotor activity (Fig. 3a). The H1R-deficient mice showed increased locomotor activity especially during the early portion of the light phase and decreased ambulation during the dark period. Total ambulation during the 24 hr was not significantly different between wt and homozygous mutant mice, but mutant mice showed significant increase of locomotor activity during the light period (P < 0.05) and decreased movement during the dark period. Ratio of ambulation during the light period to total ambulation during 24 hr and to the dark period were remarkably high in mutant mice (P < 0.0001 for both cases) (Fig. 3b).
Figure 3.
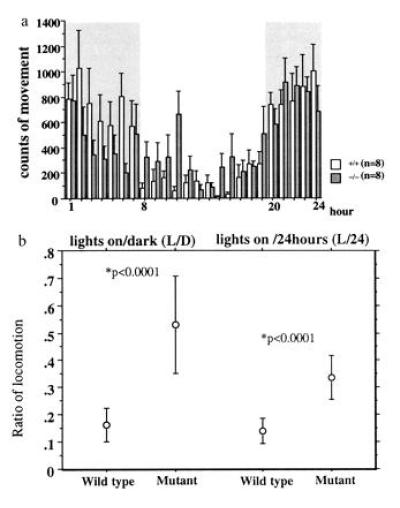
Locomotor activity of H1R mutant mice. (a) Ambulation of the mouse was monitored every an hr for two days in a cage under a 12 hr light-dark cycle. Locomotor activity of last 24 hr was shown in every hr. Shaded area represents the dark period. Data are expressed as means ± SEM from eight mice. H1R mutant were significantly more active in the light period, and less active in the dark period of the light/dark cycle. (b) The ratio of ambulation during the light period to the locomotor activity during the 24 hr (L/24) and the dark period (L/D) are shown. Data are expressed as means ± SD from eight mice.
Impaired Exploratory Behavior in H1R-Deficient Mice.
Under stressful conditions, such as the passive avoidance test using electrical shock, the H1R-deficient mice showed more frequent urination and defecation, and decreased movements in comparison with wt mice, but showed no difference in latency of passive avoidance between the two groups (data not shown). Since histaminergic neurons receive inputs mainly from the limbic system (9), the exploratory behavior in a new environment was examined to clarify whether the absence of H1R in mice affects actions related to their emotion. The distance of ambulation and the total time of rearings were measured in an open-field by a photosensor apparatus (20). Although motor coordination and grasp reflex were normal in H1R-deficient mice, the mutant mice significantly decreased ambulation and time of rearings for the first 30 min in a new place (Fig. 4 a and b). This strongly indicates that through H1R, the histaminergic neuron system plays an important role in locomotor activities related to emotion.
Figure 4.
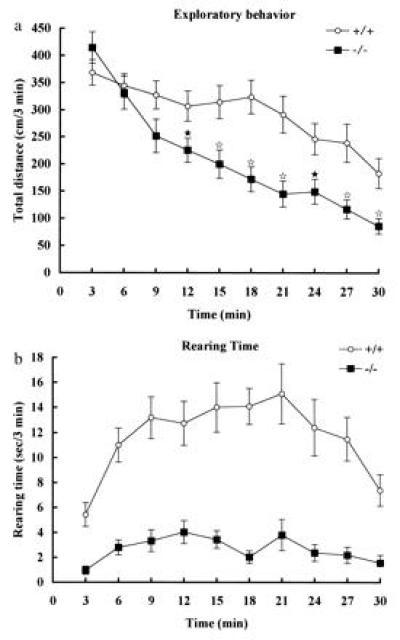
Exploratory behavior of H1R-deficient (−/−) and wt (+/+) mice in a new environment. (a) Locomotor activity. (b) Time of rearings. Data are expressed as means ± SEM from 23 wild-type and 30 mutant mice. ★, P < 0.05; ⋆, P < 0.01. Rearing was significantly decreased in H1R-deficient mice at all time points (P < 0.001).
DISCUSSION
Histamine is an endogenous compound that is synthesized, stored, and released primarily by mast cells and after release exerts profound effects on many tissues and organs. It is one of the cellular mediators of the immediate hypersensitivity reaction and the acute inflammatory response, as well as a primary stimulant of gastric acid secretion. A central neurotransmitter role for histamine has been recently demonstrated (1–5). Because the actions of histamine have considerable species variation in the periphery, it would not be easy to extrapolate the action of histamine obtained from studies on experimental animals to humans. The median lethal dose for histamine is estimated to be more than 500 mg/kg in mice whereas that in guinea pigs is less than 0.3 mg/kg. Actually, in our studies, there was no significant difference of passive cutaneous reaction between the homozygous and wt mice (data not shown). On the contrary, the central actions of histamine have been ubiquitously observed among the species.
The function of histamine in the CNS has been investigated mainly by intraperitoneal or intracerebral injection of its antagonists, agonists, and synthesis inhibitor. The precise physiological role of histamine in the CNS is still unclear. Since these drugs are metabolized, have other pharmacological effects, and may require a stressful delivery procedure, the results must be considered with care. Mutant mice lacking H1R, generated here by homologous recombination, are superior in this regard.
Binding studies were used to confirm the absence of H1R in mutant mice. Though existence of subclasses of H1R was suggested in some reports, these subclasses have not been isolated so far. The results of [3H]pyrilamine binding study in mutant mice indicate that H1R is the only binding protein of pyrilamine in brain. In addition, the loss of the high affinity component of doxepin binding in mutant mice confirmed that the high affinity binding site is related to H1R. This result was essentially the same as the previous studies (16, 17).
Morphologically, the histaminergic neuron system arises from the tuberomammillary nucleus of the hypothalamus and projects efferent fibers of varicosity to almost all brain (5–9). It has been shown that the suprachiasmatic nucleus and secondary circadian pacemakers, such as the ventromedial hypothalamus, receive a rich histaminergic innervation (5–9). The autoradiography of [3H]pyrilamine showed the existence of many H1R around these areas (11, 21) (Fig. 2). It was also reported that histamine release from the hypothalamus in rats showed a clear circadian rhythm, being high in the dark period and low in the light period (22). Intracerebroventricular injection of histaminergic and antihistaminergic drugs modulates ambulation and circadian rhythm of locomotor activity and sleep-waking cycles (23, 24). These suggest the possibility of histamine playing a crucial role for circadian rhythm in some activities.
Pharmacological studies have demonstrated that the central histaminergic neurons may be involved in circadian rhythm and act as an awaking amine. It is well known that many antihistamines cause drowsiness and impaired performance in humans, which is thought to be due mainly to their blockade of central H1R (25, 26). Our mutant mice, lacking H1R, show decreased ambulation in the dark period and increased locomotor activity in the light period, but no difference in total ambulation compared with wt mice. The distorted locomotor activity in the dark period observed in H1R mutant mice is probably due to the blockade of its neurotransmission. It is well known that intracerebroventricular administration of histamine or H1 agonists induce behavioral arousal (23). However, it is not clear why the locomotion in the light period was augmented. Though this result appears to be contrary to some previous pharmacological studies, the long-term studies on locomotor activity induced by the blockade of H1R neurotransmission have observed conflicting changes. For example, the intraperitoneal administration of (S)-α-fluoromethylhistidine, a specific inhibitor of histidine decarboxylase, decreased the short-term exploratory behavior of mice but increased the long-term locomotor activity in association with the depletion of brain histamine (20, 27). And according to another report, intracerebroventricular injection of (S)-α-fluoromethylhistidine in rat decreased the total ambulation and increased L/24 ratio of ambulation (28). In mutant mice, the congenital defect of H1R might affect other factors.
Information concerning light-dark cycle is transmitted via the eyes and the retinohypothalamic tract to the suprachiasmatic nucleus (29). Histamine concentrations in the retina and iris-ciliary body are reported to change significantly upon the change from the dark to light (30). The distribution of histaminergic neuron system and the findings in the present study suggest that histamine might play a certain role in the circadian rhythm in response to light. But locomotor activity might be affected not only by light but also by noise, smell, and other unknown factors in experiment conditions. Other possibilities cannot be also excluded completely.
Mutant mice also demonstrated decreased exploratory behavior in new environments. This result was almost consistent with the several previous works that classical H1 antagonists showed a potent depressant effect on locomotor activity (31). This result is also convincing for the fact that histamine is involved in the regulation of emotional behaviors (2). Through histamine and the H1R, the histamine neuron system may function as a regulatory center in the CNS, in relation to rhythm and emotion (1). This animal model should be a useful tool to clarify functions of histaminergic nervous system in the CNS.
Acknowledgments
We thank H. Riele and M. Hooper for E14 ES cells, Y. Horio and H. Fukui for information of H1R gene, A. Miyake and K. Nakao for technical assistance, and M. Katsuki for valuable comments. This work was supported in part by grant from the Ministry of Health and Welfare of Japan, and by grant from Japan Research Foundation for Clinical Pharmacology.
Footnotes
Abbreviations: H1R, histamine H1 receptors; wt, wild type; ES, embryonic stem; CNS, central nervous system.
References
- 1.Wada H, Inagaki N, Yamatodani A, Watanabe T. Trends Neurosci. 1991;14:415–418. doi: 10.1016/0166-2236(91)90034-r. [DOI] [PubMed] [Google Scholar]
- 2.Schwartz J C, Arrang J M, Garbarg M, Pollard H, Ruat M. Physiol Rev. 1991;71:1–51. doi: 10.1152/physrev.1991.71.1.1. [DOI] [PubMed] [Google Scholar]
- 3.Hill S J. Pharmacol Rev. 1990;42:45–83. [PubMed] [Google Scholar]
- 4.Tuomisto L. In: Histaminergic Neurons: Morphology and Function. Watanabe T, Wada H, editors. Boca Raton, FL: CRC; 1991. pp. 283–295. [Google Scholar]
- 5.Watanabe T, Taguchi Y, Shiosaka S, Tanaka J, Kubota H, Terano Y, Tohyama M, Wada H. Brain Res. 1984;295:13–25. doi: 10.1016/0006-8993(84)90811-4. [DOI] [PubMed] [Google Scholar]
- 6.Panula P, Yang H Y, Costa E. Proc Natl Acad Sci USA. 1984;81:2572–2576. doi: 10.1073/pnas.81.8.2572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Airaksinen M S, Panula P. J Comp Neurol. 1988;273:163–186. doi: 10.1002/cne.902730204. [DOI] [PubMed] [Google Scholar]
- 8.Inagaki N, Yamatodani A, Ando-Yamamoto M, Tohyama M, Watanabe T, Wada H. J Comp Neurol. 1988;273:283–300. doi: 10.1002/cne.902730302. [DOI] [PubMed] [Google Scholar]
- 9.Ericson H, Blomqvist A, Kohler C. J Comp Neurol. 1991;311:45–64. doi: 10.1002/cne.903110105. [DOI] [PubMed] [Google Scholar]
- 10.Fukui H, Inagaki N, Ito S, Kubo A, Kondoh H, Yamatodani A, Wada H. Agents Actions, Suppl. 1991;33:363–379. doi: 10.1007/978-3-0348-7309-3_12. [DOI] [PubMed] [Google Scholar]
- 11.Palacios J M, Wamsley J K, Kuhar M J. Neuroscience. 1981;6:15–37. doi: 10.1016/0306-4522(81)90240-2. [DOI] [PubMed] [Google Scholar]
- 12.Taniuchi I, Kitamura D, Maekawa Y, Fukuda T, Kishi H, Watanabe T. EMBO J. 1995;14:3664–3678. doi: 10.1002/j.1460-2075.1995.tb00036.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yanai K, Ryu J H, Watanabe T, Iwata R, Ido T. NeuroReport. 1992;3:961–964. doi: 10.1097/00001756-199211000-00003. [DOI] [PubMed] [Google Scholar]
- 14.Chang R S, Tran V T, Snyder S H. J Neurochem. 1979;32:1653–1663. doi: 10.1111/j.1471-4159.1979.tb02276.x. [DOI] [PubMed] [Google Scholar]
- 15.Kanba S, Richelson E. Brain Res. 1984;304:1–7. doi: 10.1016/0006-8993(84)90856-4. [DOI] [PubMed] [Google Scholar]
- 16.Taylor J E, Richelson E. Eur J Pharmacol. 1982;78:279–285. doi: 10.1016/0014-2999(82)90029-2. [DOI] [PubMed] [Google Scholar]
- 17.Tran V T, Lebovitz R, Toll L, Snyder S H. Eur J Pharmacol. 1981;70:501–509. doi: 10.1016/0014-2999(81)90361-7. [DOI] [PubMed] [Google Scholar]
- 18.Yanai K, Watanabe T, Yokoyama H, Meguro K, Hatazawa J, Itoh M, Iwata R, Ishiwata K, Takahashi T, Ido T. Neurosci Lett. 1992;137:145–148. doi: 10.1016/0304-3940(92)90390-s. [DOI] [PubMed] [Google Scholar]
- 19.Yamashita M, Fukui H, Sugama K, Horio Y, Ito S, Mizuguchi H, Wada H. Proc Natl Acad Sci USA. 1991;88:11515–11519. doi: 10.1073/pnas.88.24.11515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sakai N, Onodera K, Maeyama K, Yanai K, Watanabe T. Life Sci. 1992;51:397–405. doi: 10.1016/0024-3205(92)90406-f. [DOI] [PubMed] [Google Scholar]
- 21.Bouthenet M L, Ruat M, Sales N, Garbarg M, Schwartz J C. Neuroscience. 1988;26:553–600. doi: 10.1016/0306-4522(88)90167-4. [DOI] [PubMed] [Google Scholar]
- 22.Mochizuki T, Yamatodani A, Okakura K, Horii A, Inagaki N, Wada H. Physiol Behav. 1992;51:391–394. doi: 10.1016/0031-9384(92)90157-w. [DOI] [PubMed] [Google Scholar]
- 23.Lin J-S, Sakai K, Jouvet M. Neuropharmacology. 1988;27:111–122. doi: 10.1016/0028-3908(88)90159-1. [DOI] [PubMed] [Google Scholar]
- 24.Itowi N, Yamatodani A, Mochizuki T, Wada H. Neurosci Lett. 1991;123:53–56. doi: 10.1016/0304-3940(91)90156-n. [DOI] [PubMed] [Google Scholar]
- 25.Nicholson A N, Pascoe P A, Turner C, Ganellin C R, Greengrass P M, Casy A F, Mercer A D. Br J Pharmacol. 1991;104:270–276. doi: 10.1111/j.1476-5381.1991.tb12418.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yanai K, Ryu J H, Watanabe T, Iwata R, Ido T, Sawai Y, Ito K, Itoh M. Br J Pharmacol. 1995;116:1649–1655. doi: 10.1111/j.1476-5381.1995.tb16386.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sakai N, Sakurai E, Onodera K, Sakurai E, Asada H, Miura Y, Watanabe T. Behav Brain Res. 1996;72:83–88. doi: 10.1016/0166-4328(96)00059-9. [DOI] [PubMed] [Google Scholar]
- 28.Doi T, Sakata T, Yoshimatsu H, Machidori H, Kurokawa M, Jayasekara L A, Niki N. Brain Res. 1994;641:311–318. doi: 10.1016/0006-8993(94)90160-0. [DOI] [PubMed] [Google Scholar]
- 29.Mosko S S, Moore R Y. Brain Res. 1979;164:17–38. doi: 10.1016/0006-8993(79)90003-9. [DOI] [PubMed] [Google Scholar]
- 30.Nowak J Z, Socko R, Uznanski P. Agents Actions. 1988;23:233–236. doi: 10.1007/BF02142550. [DOI] [PubMed] [Google Scholar]
- 31.Weidmann H, Petersen P V. J Pharmacol Exp Ther. 1953;108:201–216. [PubMed] [Google Scholar]