

Abstract
Current therapies for inorganic mercury (Hg2+) intoxication include administration of a metal chelator, either 2,3-dimercaptopropane-1-sulfonic acid (DMPS) or meso-2,3-dimercaptosuccinic acid (DMSA). After exposure to either chelator, Hg2+ is rapidly eliminated from the kidneys and excreted in the urine, presumably as an S-conjugate of DMPS or DMSA. The multidrug resistance protein 2 (Mrp2) has been implicated in this process. We hypothesize that Mrp2 mediates the secretion of DMPS- or DMSA-S-conjugates of Hg2+ from proximal tubular cells. To test this hypothesis, the disposition of Hg2+ was examined in control and Mrp2-deficient TR− rats. Rats were injected i.v. with 0.5 μmol/kg HgCl2 containing 203Hg2+. Twenty-four and 28 h later, rats were injected with saline, DMPS, or DMSA. Tissues were harvested 48 h after HgCl2 exposure. The renal and hepatic burden of Hg2+ in the saline-injected TR− rats was greater than that of controls. In contrast, the amount of Hg2+ excreted in urine and feces of TR− rats was less than that of controls. DMPS, but not DMSA, significantly reduced the renal and hepatic content of Hg2+ in both groups of rats, with the greatest reduction in controls. A significant increase in urinary and fecal excretion of Hg2+, which was greater in the controls, was also observed following DMPS treatment. Experiments utilizing inside-out membrane vesicles expressing MRP2 support these observations by demonstrating that DMPS- and DMSA-S-conjugates of Hg2+ are transportable substrates of MRP2. Collectively, these data support a role for Mrp2 in the DMPS- and DMSA-mediated elimination of Hg2+ from the kidney.
2,3-Dimercaptopropane-1-sulfonic acid (DMPS) and meso-2,3-dimercaptosuccinic acid (DMSA) are dithiol metal chelators that have been demonstrated to reduce significantly and efficiently the body burden of mercury (Planas-Bohne, 1981; Aposhian, 1983; Aposhian et al., 1992; Zalups et al., 1992; Ruprecht, 1997). These chelators act mainly by promoting the unidirectional extraction of mercuric ions from within renal proximal tubular epithelial cells into the luminal compartment of the nephron. This extraction provides an effective route for the elimination of mercuric ions in the urine. Unlike other chelators, DMPS and DMSA are highly water soluble and are relatively nontoxic, which has contributed to their use as antidotes for mercury poisoning throughout the world. Surprisingly, only DMSA has been approved for use in humans by the Food and Drug Administration of the United States. Because of structural and chemical differences between DMPS and DMSA and their varied use throughout the world, it is important to understand the effect of each of these chelators on the disposition and handling of the various species of mercury.
After exposure to inorganic mercury (Hg2+), mercuric ions accumulate primarily in the renal proximal tubule (Zalups, 2000). Hg2+ gains access to proximal tubular cells primarily via amino acid transporters in the luminal plasma membrane and the organic anion transporter 1 in the basolateral plasma membrane (Zalups, 2000; Bridges and Zalups, 2005). The extraction of Hg2+ from proximal tubular cells via DMPS or DMSA appears to be unidirectional, where mercuric ions are moved from the intracellular compartment into the tubular lumen. Indeed, treatment of humans and rats with DMPS or DMSA has been shown to reduce the amount of Hg2+ within the kidneys and increase the urinary excretion of Hg2+ (Planas-Bohne, 1981; Aposhian, 1983; Aposhian and Aposhian, 1990; Aposhian et al., 1992; Zalups, 1993; Zalups et al., 1998). The extraction of Hg2+ from renal tubular cells does not appear to involve glomerular filtration but rather a direct secretory process whereby Hg2+ moves from peritubular blood into the tubular lumen (Diamond et al., 1988; Zalups et al., 1998). It is well documented that reduced and oxidized forms of DMPS (but not DMPS S-conjugates of Hg2+) are taken up by the organic anion transporter 1 in the basolateral membrane of proximal tubular cells (Zalups et al., 1998; Islinger et al., 2001; Bahn et al., 2002). DMSA, however, appears to be taken up by the sodium-dependent dicarboxylate transporter, which is also localized in the basolateral membrane of these cells (Burckhardt et al., 2002).
Given the current body of scientific knowledge, we hypothesize that after being taken up into cells, DMPS and DMSA bond to intracellular Hg2+, creating DMPS S- or DMSA S-conjugates of Hg2+. These conjugates are then exported across the luminal plasma membrane into the tubular lumen for elimination in the urine. Although it is well known that treatment with DMPS and/or DMSA facilitates a profound decrease in the renal burden of Hg2+ (Aposhian, 1983), the cellular mechanisms involved in this process have not been defined fully.
Because of data from rats implicating the multidrug resistance protein (Mrp) in the hepatobiliary transport of Hg2+ (Sugawara et al., 1998), we suggest that this carrier may also be involved in the export of DMPS S- and DMSA S-conjugates of Hg2+ from proximal tubular cells. Mrp2 is a member of the ATP-binding cassette transporter superfamily and is localized in the luminal membrane of proximal tubular cells (Schaub et al., 1997). This transporter has been hypothesized to play a role in the excretion of a number of metabolic wastes and xenobiotics (Leslie et al., 2005). The activity of human MRP2 appears to be dependent upon the cotransport of glutathione for the secretion of certain compounds (Leslie et al., 2005). Interestingly, Tanaka-Kagawa et al. (1993) suggest that the secretion of Hg2+ from proximal tubular cells also appears to require the cotransport of glutathione. These data suggest that Mrp2 may be responsible for at least part of the proximal tubular elimination of Hg2+.
Given the aforementioned data, we hypothesize that Mrp2 (and possibly additional members of the superfamily of ABC transporters) plays a role in the secretion of DMPS S- or DMSA S-conjugates of Hg2+ from within proximal tubular cells into the tubular lumen. We tested this hypothesis by examining the disposition of Hg2+ in Mrp2-deficient rats (TR−), before and after treatment with DMPS or DMSA. To our knowledge, the results of the current study provide the first substantive line of evidence indicating that DMPS S-conjugates of Hg2+ can be transported into the tubular lumen by Mrp2. Perhaps more importantly, our data also provide evidence suggesting that Mrp2 may be the primary transporter involved in the secretion of DMSA S-conjugates of Hg2+.
Materials and Methods
Chemicals
DMPS (sodium salt), DMSA, sodium pentobarbital, and HgCl2 were all obtained from Sigma-Aldrich (St. Louis, MO). Radioactive HgCl2 (203Hg2+) was produced by irradiation of mercuric oxide at the Missouri University Research Reactor facility as described previously (Belanger et al., 2001; Bridges et al., 2004). The specific activity of the 203Hg2+ ranged from 6 to 12 mCi/mmol.
Animals
Transport-deficient (TR−) rats possess a mutation in the mrp2 gene (Mayer et al., 1995; Paulusma et al., 1996); consequently, the Mrp2 protein is absent in the tissues and organs of these animals (Mayer et al., 1995). These rats represent a reliable model that has been used to study the hepatic and renal secretion of various Mrp2 substrates (de Vries et al., 1989; Masereeuw et al., 2003; Smeets et al., 2004).
Male TR− and normal (control) Wistar rats weighing 200 to 225 g were purchased from Harlan Laboratories (Indianapolis, IN). There were no significant differences between body weights of the animals used for these studies. All animals were provided a commercial laboratory diet (Tekland 6% rat diet; Harlan Laboratories) and water ad libitum throughout all aspects of animal experimentation. Animals were handled in accordance with the Institute of Laboratory Animal Resources (1996).
Intravenous Injections
Control and TR− rats were injected i.v. according to a previously published protocol (Zalups et al., 1992; Zalups, 1993). Each animal was anesthetized lightly with ether, and a small incision was made in the skin in the midventral region of the thigh to expose the femoral vein and artery. The fascia around the femoral vein was trimmed and a non-nephrotoxic dose of HgCl2 (0.5 μmol/kg in 2 ml of normal saline containing 1 μCi of 203Hg2+ per rat) was administered into the vein. The wound was closed using two 9-mm stainless steel wound clips.
Experimental Design
Eight control and eight TR− rats were injected i.v. with a 0.5 μmol/kg dose of HgCl2 as detailed above. Following the administration of HgCl2, control rats were divided randomly into two groups, with each group consisting of four rats. TR− rats were also divided randomly into two groups of four rats each. Animals were then placed in individual plastic metabolic cages, in which water and food were provided ad libitum.
Twenty-four hours after the injection of HgCl2, four control rats and four TR− rats were injected i.p. with a 100 mg/kg (0.39 μmol/kg) dose of DMPS (in 2 ml/kg normal saline). At the same time, another group of four control rats and a group of four TR− rats were injected i.p. with normal saline (2 ml/kg). Four hours later (28 h after injection of HgCl2), rats were administered a second i.p. dose of DMPS (100 mg/kg) or saline (2 ml/kg). Twenty-four hours after the initial dose of DMPS or saline (48 h after injection with HgCl2), rats were sacrificed by exsanguination, and blood, liver, kidneys, urine, and feces were collected for analysis of Hg2+ content. In a separate set of experiments, rats were treated with 100 mg/kg (0.55 μmol/kg) DMSA instead of DMPS. All other experimental details remained the same.
Collection of Tissues, Organs, Urine, and Feces
Forty-eight hours after the initial injection of Hg2+, rats were anesthetized with an i.p. overdose of sodium pentobarbital (100 mg/kg in 2 ml of saline). Two 1-ml samples of blood were obtained from the inferior vena cava with a 3-ml syringe and a 20-gauge needle. One of the samples was placed in a polystyrene tube for determination of 203Hg2+ content, whereas the other sample was placed in a Microtainer tube (Becton Dickinson and Co., Franklin Lakes, NJ), which was centrifuged at 21,000g for 90 s. Subsequently, the cellular and plasma fractions were removed and placed in separate polystyrene tubes for estimation of Hg content. Total blood volume was estimated to be 6% of body weight.
The right and left kidneys were also removed from each animal. Each kidney was weighed and cut in half along a transverse plain. A 3-mm transverse slice of the left kidney was utilized for separation of cortex, outer stripe of outer medulla, inner stripe of outer medulla, and inner medulla. Each zone of the kidney was weighed and placed in a polystyrene tube for estimation of 203Hg2+ content. The liver was then excised carefully, weighed, and a 1-g section was removed for determination of 203Hg2+ content.
Urine and feces were collected throughout the duration of each experiment. The urine excreted by each animal was collected 24 and 48 h after injection with HgCl2. Subsequently, the urine from each animal was mixed by vortexing, and a 1-ml sample was weighed and placed in a polystyrene tube for estimation of 203Hg2+ content. All of the feces excreted by each animal during each 24-h period were counted to determine accurately the total fecal content of 203Hg2+. The content of 203Hg2+ in each sample was determined by counting the samples in a Wallac Wizard 3 automatic gamma counter (PerkinElmer Life and Analytical Sciences, Boston, MA).
Membrane Vesicle Transport Assays
Membrane vesicle transport assays were performed using a rapid filtration method as described previously (Cui et al., 1999; Chancy et al., 2000). Inside-out membrane vesicles prepared from either Sf9 cells were transfected with human MRP2 or control SF9 cells obtained from Xenotech (Lenexa, KS). Before use, vesicles were centrifuged at 100,000g for 40 min at 4°C to remove the storage buffer. Vesicles were resuspended in ice-cold incubation buffer (250 mM sucrose, 10 mM Tris/HCl, 10 mM MgCl2, pH 7.4) by passing the suspension through a 27-gauge needle 25 times. Mercuric conjugates were formed by mixing 5 μM 203Hg2+ with 12.5 μM DMPS or DMSA in incubation buffer supplemented with 10 mM creatine phosphate and 100 μg/ml creatine phosphokinase in the presence or absence of 5 mM ATP. At the time of the experiment, membrane vesicles and mercuric conjugates were warmed to 37°C. Transport was initiated by the addition of 40 μl of membrane vesicle solution (20 μg) to 160 μl of DMPS-S-Hg-S-DMPS or DMSA-S-Hg-S-DMSA. Transport was allowed to proceed for 15 s, after which vesicles were collected on prewet HNWP nylon filter discs (pore size, 0.2 μm; Millipore, Billerica, MA). Filters were washed with two changes (8 ml each) of ice-cold incubation buffer containing 1 mM DMPS. Filters were placed in scintillation vials and the radioactivity present was determined by gamma counting in a Wallac gamma counter (PerkinElmer, Waltham, MA).
Data Analyses
Data for each rat experiment were analyzed first with the Kolmogorov-Smirnov test for normality and then with Levene's test for homogeneity of variances. Data were then analyzed using a 2 × 2 two-way analysis of variance to assess differences among the means. When statistically significant F values were obtained with analysis of variance, the data were analyzed using Tukey's post hoc multiple comparison test. A p value < 0.05 was considered statistically significant.
Data from membrane vesicle assays were analyzed by first calculating ATP-dependent transport. This calculation was performed by subtracting the amount of Hg2+ associated with the membrane vesicles in the absence of ATP from that associated with vesicles in the presence of ATP. The resulting data were analyzed by the Student's t test. A p value < 0.05 was considered statistically significant.
Results
Effect of DMPS on the Renal Burden of Hg2+ after Exposure to HgCl2
Figure 1 shows the renal burden of Hg2+ in control rats and TR− rats treated with either saline or DMPS 24 and 28 h after the injection of a 0.5 μmol/kg dose of Hg2+. The content of Hg2+ in the left kidney of TR− rats injected with saline (approximately 31% of dose) was significantly greater in than that of corresponding saline-injected controls (approximately 25% of dose). The total renal burden of Hg2+ was also greater in saline-injected TR− rats (approximately 61% of dose) than in matched controls (approximately 48% of dose). Treatment with DMPS caused a significant reduction in the renal content of Hg2+ in control and TR− rats. In controls, treatment with DMPS lowered the content of Hg2+ in the total renal mass by approximately 87%, whereas in the TR− rats, treatment with DMPS lowered the content of Hg2+ in the total renal mass by only approximately 38%. Even with these decreases in the renal burden of Hg2+, the content of Hg2+ in the total renal mass of TR− rats remained 6-fold greater than that of corresponding DMPS-treated controls.
Fig. 1.
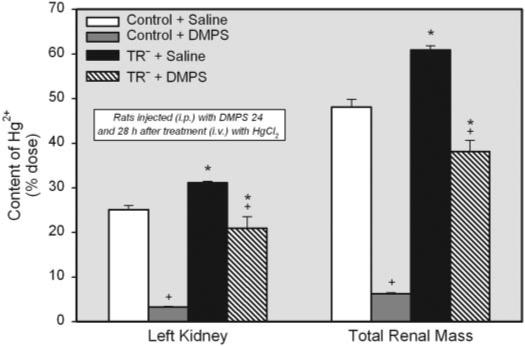
Content of Hg2+ in the left kidney and total renal mass (percentage of administered dose) of control and TR− rats injected i.v. with 0.5 μmol/kg HgCl2. Rats were injected i.p. with 100 mg/kg DMPS or 2 ml/kg saline 24 and 28 h after injection of HgCl2. Kidneys were harvested for determination of Hg2+ content after 48-h exposure to HgCl2. Data represent mean ± S.E. of four rats. *, significantly different (p < 0.05) from the corresponding mean for control rats treated in the same manner.
The effect of DMPS on the disposition of Hg2+ in the four zones of the kidney is shown in Fig. 2. The concentration of Hg2+ in the renal cortex was significantly greater in the saline-treated TR− rats than in the corresponding saline-treated control rats. However, the concentration of mercury in the outer stripe of the outer medulla was significantly greater in the saline-treated control rats than in the saline-treated TR− rats. In the DMPS-treated control rats, the concentration of Hg2+ was profoundly decreased in both the cortex and outer stripe of the outer medulla. Although treatment with DMPS induced significant reductions in the concentration of Hg2+ in the renal cortex and outer stripe of the outer medulla in the TR− rats, the reductions were not as great as that detected in the DMPS-treated control rats.
Fig. 2.
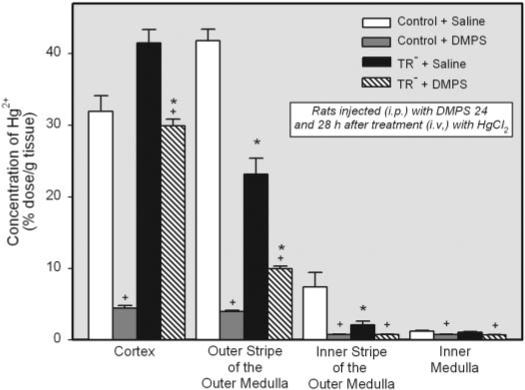
Concentration of Hg2+ (percentage of administered dose per gram of tissue) in the various zones of kidney of control and TR− rats injected i.v. with 0.5 μmol/kg HgCl2. Rats were injected i.p. with 100 mg/kg DMPS or 2 ml/kg saline 24 and 28 h after injection of HgCl2. Kidneys were harvested for determination of Hg2+ content after 48-h exposure to HgCl2. Data represent mean ± S.E. of four rats. *, significantly different (p < 0.05) from the corresponding mean for control rats treated in the same manner. +, significantly different (p < 0.05) from the corresponding mean for the same strain of rats treated with saline.
Nominal amounts of Hg2+ were detected in the inner stripe of the outer medulla and inner medulla. Although some differences in the disposition of Hg2+ were detected in these two zones among the four groups of rats studied, the differences are viewed as being due to slight differences in the separation of zones at the time of sacrifice.
Effect of DMPS on the Burden of Hg2+ in Liver and Blood after Exposure to HgCl2
The total amount of Hg2+ in liver of saline-injected and DMPS-treated TR− rats (approximately 9% of the dose in both groups) was significantly greater than that of corresponding saline-treated and DMPS-treated control rats (approximately 3% of dose in both groups) (Fig. 3). Interestingly, treatment with DMPS had no significant effect on the hepatic burden of Hg2+ in both control and TR− rats.
Fig. 3.
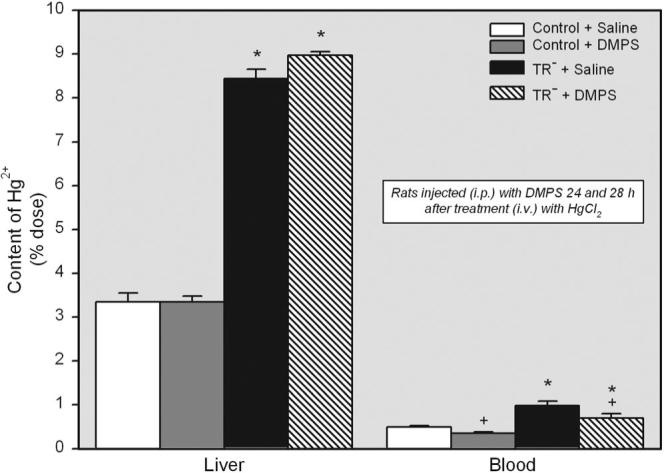
Content of Hg2+ in liver and blood (percentage of administered dose) of control and TR− rats injected i.v. with 0.5 μmol/kg HgCl2 followed 24 and 28 h later by treatment i.p. with 100 mg/kg DMPS or 2 ml/kg saline. Livers and blood were harvested 48 h after exposure to HgCl2. Data represent mean ± S.E. of four rats. *, significantly different (p < 0.05) from the corresponding mean for control rats treated in the same manner. +, significantly different (p < 0.05) from the corresponding mean for the same strain of rats treated with saline.
Hg2+ levels in blood were greater in saline-treated and DMPS-treated TR− rats than in corresponding control rats treated in the same manner. Moreover, administration of DMPS reduced the amount of Hg2+ in blood by approximately 30% in both control and TR− rats (Fig. 3).
Effect of DMPS on the Urinary and Fecal Excretion of Hg2+ after Exposure to HgCl2
The amount of Hg2+ excreted in the urine after treatment with saline was significantly greater in the control rats than in the corresponding TR− rats (Fig. 4A). Treatment with DMPS increased the urinary excretion of Hg2+ in control rats by approximately 13-fold and in TR− rats by nearly 40-fold. Nonetheless, the urinary excretion of Hg2+ by the TR− rats, following treatment with DMPS, remained significantly less than that of control rats.
Fig. 4.
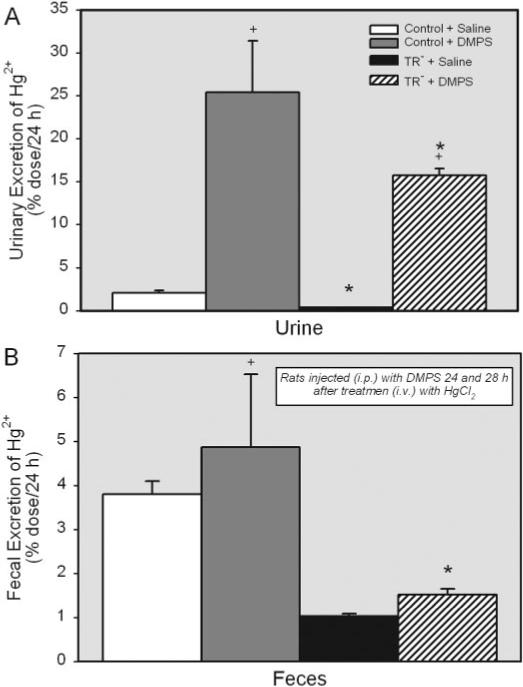
Amount of Hg2+ excreted in urine and feces (percentage of administered dose) of control and TR− rats injected i.v. with 0.5 μmol/kg HgCl2, followed 24 and 28 h later by injection with 100 mg/kg DMPS or saline (2 ml/kg). Data represent the amount of Hg2+ in urine and feces collected during the 24 h following treatment with DMPS. Data represent mean ± S.E. of four rats. *, significantly different (p < 0.05) from the corresponding mean for control rats treated in the same manner.
The fecal elimination of Hg2+ was also compared in control and TR− rats (Fig. 4B). The amount of Hg2+ excreted into the feces by the control rats treated with saline was approximately 25% greater than that excreted in the feces by the corresponding saline-injected TR− rats. In control rats, DMPS enhanced the fecal elimination of Hg2+ by approximately 2-fold. In contrast, treatment with DMPS did not alter significantly the fecal elimination of Hg2+ by TR− rats.
Effect of DMSA on the Renal Burden of Hg2+ after Exposure to HgCl2
Separate groups of control and TR− rats were treated with saline or DMSA 24 and 28 h after the i.v. injection of HgCl2. The disposition of Hg2+ in these animals was examined 48 h after the initial injection of Hg2+. The content of Hg2+ in the left kidney was significantly greater in TR− rats injected with saline (approximately 32% of dose) than in corresponding saline-treated controls (approximately 24% of dose) (Fig. 5). Likewise, the percentage of administered dose of Hg2+ in the total renal mass was greater in saline-injected TR− rats (approximately 66% of dose) than in corresponding saline-injected controls (approximately 48% of dose) (Fig. 5). Treatment with DMSA reduced the total renal content of Hg2+ in control rats by approximately 80%. Interestingly, DMSA did not alter the renal burden of Hg2+ in the TR− rats.
Fig. 5.
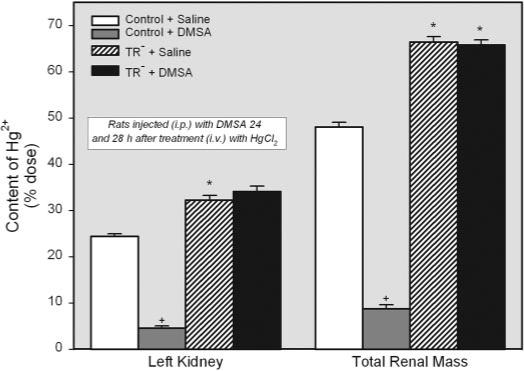
Content of Hg2+ (percentage of administered dose) in the left kidney and total renal mass of control and TR− rats injected i.v. with 0.5 μmol/kg HgCl2. Rats were injected i.p. with 100 mg/kg DMSA or 2 ml/kg saline 24 and 28 h after injection of HgCl2. Kidneys were harvested for determination of Hg2+ content after 48-h exposure to HgCl2. *, significantly different (p < 0.05) from the corresponding mean for control rats treated in the same manner. +, significantly different (p < 0.05) from the corresponding mean for the same strain of rats treated with saline.
The effect of DMSA on the concentration of Hg2+ in the various zones of the kidney was also examined (Fig. 6). Significant differences in the concentration of Hg2+ were detected among the four groups of rats mainly in the cortex and outer stripe of the outer medulla. The concentration of Hg2+ in the renal cortex was significantly greater in both groups of TR− rats than in the corresponding groups of control animals. In contrast, the concentration of Hg2+ in the outer stripe of the outer medulla was significantly less in both groups of TR− rats than in the corresponding groups of control animals. One of the most striking findings in the renal zones is that treatment with DMSA effectively reduced the concentration of Hg2+ in each of the zones of control rats yet had no effect on the concentration of Hg2+ in the TR− rats.
Fig. 6.
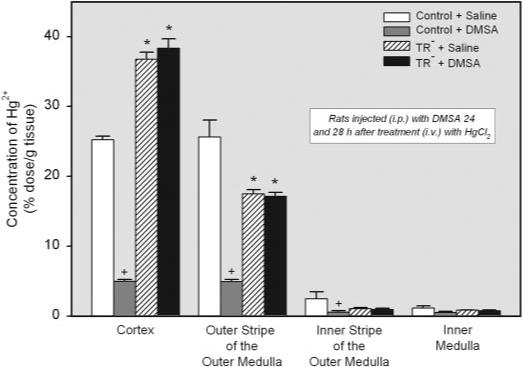
Concentration of Hg2+ (percent administered dose per gram of tissue) in the various zones of kidneys removed from the same animals. Rats were injected i.p. with 100 mg/kg DMSA or 2 ml/kg saline 24 and 28 h after injection of HgCl2. Kidneys were harvested for determination of Hg2+ content after 48-h exposure to HgCl2. *, significantly different (p < 0.05) from the corresponding mean for control rats treated in the same manner. +, significantly different (p < 0.05) from the corresponding mean for the same strain of rats treated with saline.
Effect of DMSA on the Burden of Hg2+ in Liver and Blood after Exposure to HgCl2
The hepatic burden of Hg2+ in the TR− rats treated with DMSA (approximately 8% of dose) was over 2-fold greater than that of control animals (approximately 3% of dose) (Fig. 7). Interestingly, treatment of animals with DMSA had little effect on the content of Hg2+ in the liver of either strain of rat.
Fig. 7.
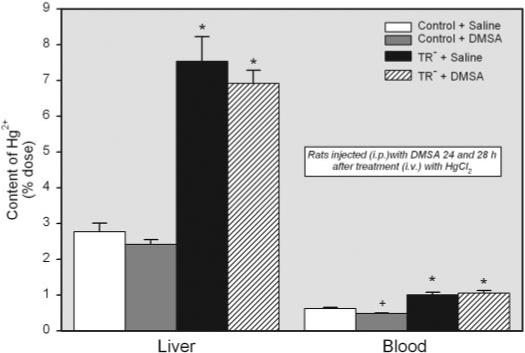
Content of Hg2+ in the liver and blood (percentage of administered dose) of control and TR− rats injected i.v. with 0.5 μmol/kg HgCl2 followed 24 and 28 h later by injection i.p. with 100 mg/kg DMSA. Livers and blood were harvested for determination of Hg2+ content 48 h after injection with HgCl2. Data represent mean ± S.E. of four rats. *, significantly different (p < 0.05) from the corresponding mean for control rats treated in the same manner. +, significantly different (p < 0.05) from the corresponding mean for the same strain of rats treated with saline.
When the content of Hg2+ in blood was measured, only approximately 1% of the dose was detected in blood of saline-injected TR− rats. A similar amount of Hg2+ was detected in the blood of corresponding saline-treated controls. Treatment with DMSA significantly reduced the amount of Hg2+ in blood of control animals but had no affect on that of TR− rats (Fig. 7).
Effect of DMSA on the Urinary and Fecal Excretion of Hg2+ Exposure to HgCl2
Urinary excretion of Hg2+ was over 12-fold greater in the control rats treated with DMSA than in the control rats treated with saline (Fig. 8A). Interestingly, there was little difference in the urinary excretion of Hg2+ between the two groups of TR− rats. Moreover, the urinary excretion of Hg2+ in the two groups of TR− rats was significantly less than that in the corresponding group of control rats.
Fig. 8.
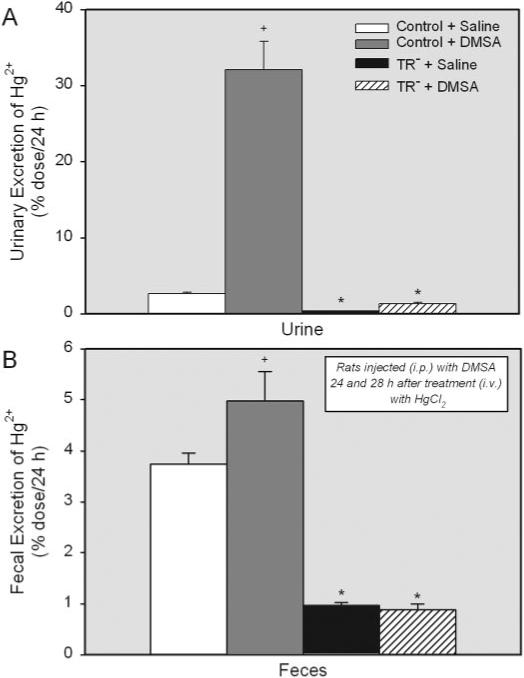
Content of Hg2+ excreted in urine (A) and feces (B) (percentage of administered dose) from control and TR− rats injected i.v. with 0.5 μmol/kg HgCl2 followed 24 and 28 h later by injection i.p. with 100 mg/kg DMSA. Data represent the amount of Hg2+ in urine and feces collected during the 24 h following treatment with DMSA. Data represent mean ± S.E. of four rats. *, significantly different (p < 0.05) from the corresponding mean for control rats treated in the same manner. +, significantly different (p < 0.05) from the corresponding mean for the same strain of rats treated with saline.
Fecal excretion of Hg2+ in the saline-injected control rats (approximately 3.5% of the dose) was greater than that in the corresponding TR− rats (approximately 1% of the dose) (Fig. 8B). DMSA increased significantly the fecal excretion of Hg2+ in control animals to approximately 5% of the dose. In TR− rats, DMSA did not alter significantly the amount of Hg2+ excreted in the feces.
Transport of Hg2+ in Inside-out Membrane Vesicles Expressing MRP2
ATP-dependent transport of DMPS-S-Hg-S-DMPS was significantly greater in inside-out membrane vesicles expressing MRP2 than in corresponding control vesicles (Fig. 9). In addition, the ATP-dependent transport of DMSA-S-Hg-S-DMSA was also significantly greater in membrane vesicles expressing MRP2 than in control vesicles (Fig. 9).
Fig. 9.
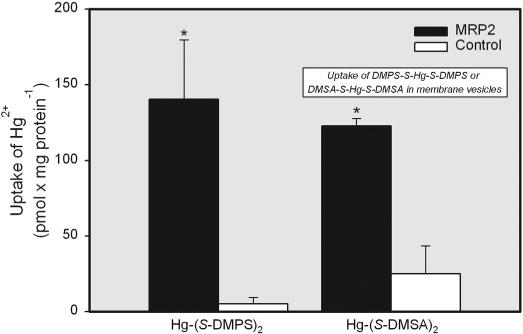
Transport of Hg2+ into inside-out membrane vesicles expressing MRP2. The transport of 5 μM Hg2+, conjugated to either 12.5 μM DMPS or DMSA, was measured at 37°C for 15 s in control and MRP2-expressing membrane vesicles. Data represent mean ± S.E. of two experiments, performed in triplicate. *, significantly different (p < 0.05) from the corresponding mean for control vesicles treated with the same mercuric conjugate.
Discussion
The purpose of the current study was to test the hypothesis that Mrp2 is involved in the export of DMPS and/or DMSA conjugates of Hg2+ from the intracellular compartment of proximal tubular cells into the tubular lumen. More specifically, we hypothesize that following administration of DMPS or DMSA, these chelators are taken up at the basolateral membrane of proximal tubular epithelial cells and form complexes with intracellular Hg2+, which are then exported into the tubular lumen via Mrp2. TR− rats were chosen as a model in which to test this hypothesis because of their lack of a functional Mrp2 (Jansen et al., 1985).
The current data clearly show differences in the disposition of Hg2+ between control and TR− rats that were not treated with DMPS or DMSA. In TR− rats injected only with saline, the levels of Hg2+ in the total renal mass, liver, and blood were greater than those in saline-treated control rats. Within the kidneys, the greatest amounts of Hg2+ were found in the cortex, although significant amounts of Hg2+ were also present in the outer stripe of the outer medulla. This pattern of renal disposition of Hg2+ is consistent with the zonal localization of the three segments of the proximal tubule in the kidneys. Moreover, this distribution parallels that of Mrp2 in the kidney (Peng et al., 1999; Schaub et al., 1997).
In addition, the amount of Hg2+ excreted in the urine and feces was significantly less in TR− rats than in controls. These differences in the handling of Hg2+ between saline-injected TR− rats and corresponding control rats suggest that Mrp2 and/or additional transporters altered in the TR− rats are involved in the proximal tubular elimination of Hg2+.
Numerous studies in humans have demonstrated that DMPS and DMSA reduce the body burden of Hg2+ rapidly and effectively (Aposhian, 1983). The results of the current study are consistent with these previous findings. DMPS and DMSA each reduced significantly the renal content of Hg2+ in control rats. Interestingly, in TR− rats, only DMPS was capable of reducing the renal burden of Hg2+. The percentage by which DMPS reduced the amount of Hg2+ in the kidneys of TR− rats (approximately a 38% reduction) was far less than that detected in control rats also treated with DMPS (approximately a 87% reduction). Since DMPS appears to be capable of reducing the renal burden of Hg2+ to a limited degree in TR− rats, we suggest that Mrp2 and at least one additional transporter (such as Mrp4, which is also localized in the luminal plasma membrane of proximal tubular epithelial cells; Van Aubel et al., 2002) are involved in the secretion of DMPS S-conjugates of Hg2+ into the lumen of the proximal tubule. Our conclusion about the involvement of Mrp2 in the luminal secretion of DMPS S-conjugates of Hg2+ is based primarily on the large body of data obtained from TR− rats that have implicated this carrier protein in the luminal secretion of other toxicants and metabolites.
Unlike DMPS, DMSA reduced effectively the renal burden of Hg2+ only in control animals (by approximately 82%). It had no effect on the renal disposition of Hg2+ in the TR− rats. These data indicate strongly that Mrp2 is the primary mechanism involved in the proximal tubular elimination of Hg2+ following treatment with DMSA. However, since a number of other transport proteins have been shown to be altered in TR− rats, we cannot conclude definitively that Mrp2 is the sole mechanism for the secretion of DMSA S-conjugates of Hg2+ into the lumen of the proximal tubule. Additional studies are clearly necessary to elucidate fully all the potential mechanisms responsible for the elimination of Hg2+ following chelation therapy.
It is clear that following administration of DMPS or DMSA to humans or animals exposed to Hg2+, the urinary excretion of mercuric ions is enhanced. Following treatment with either chelator, Hg2+ and the respective chelator have been detected in urine (Maiorino et al., 1991; Ruprecht, 1997). Moreover, data from isolated perfused S2 segments of the rabbit proximal tubule indicate that DMPS can extract mercuric ions that have accumulated in proximal tubular epithelial cells (Zalups et al., 1998). Although it is not clear from the present study as to what extent the amount of mercuric ions excreted in the urine were bound to DMPS or DMSA molecules, there are data from studies in humans showing that the rate of urinary excretion of Hg2+ is similar to that of DMPS (Maiorino et al., 1991). All of these data are consistent with our hypothesis that treatment with either DMPS or DMSA leads to mercuric ions being extracted from proximal tubular epithelial cells as conjugates of DMPS or DMSA, respectively.
DMPS and DMSA have been shown to be present in blood primarily as disulfides, bound to another DMPS or DMSA molecule, respectively, or to another sulfhydryl-containing molecule such as cysteine (Aposhian and Aposhian, 1990; Maiorino et al., 1991). Interestingly, studies in Sperber chickens have shown that DMPS-disulfides are taken up by and reduced within renal tubular cells. Once reduced, these chelators appear to form S-conjugates of Hg2+ by removing mercuric ions from intracellular ligands by a mechanism involving thiol competition (Stewart and Diamond, 1987, 1988; Klotzbach and Diamond, 1988; Diamond et al., 1988). These findings form an additional line of support for the hypothesis that, in humans, DMPS or DMSA taken up from the blood can form complexes with intracellular Hg2+ in renal proximal tubular epithelial cells, which are probable substrates of MRP2. Indeed, the current in vitro data from membrane vesicles demonstrate that DMPS S- and DMSA S-complexes of Hg2+ are transportable substrates of MRP2.
The actual structures of the DMPS S- or DMSA S-complexes of Hg2+ formed in proximal tubular cells are unclear presently. However, several studies have shown that stable DMPS S- and DMSA S-conjugates of Hg2+ are formed by a single mercuric ion bonding to the vicinal thiol groups on a single molecule of DMPS and DMSA, respectively (Rivera et al., 1989; Aposhian and Aposhian, 1990; Maiorino et al., 1991; Aposhian et al., 1995; Ruprecht, 1997). In contrast, George et al. (2004) argue that at least two molecules of DMPS or DMSA and two atoms of Hg2+ are required to form respective DMPS S- or DMSA S-conjugates of Hg2+. Regardless of the exact species of the complexes formed intracellularly, the findings from the current study and those of other studies show clearly that DMPS and DMSA are each capable of effectively reducing the renal and body burden of Hg2+ (Planas-Bohne, 1981; Aposhian, 1983; Aposhian et al., 1992; Zalups et al., 1992; Ruprecht, 1997).
In summary, the results of the present study apparently represent the first line of substantive evidence defining a role for Mrp2 in the renal cellular elimination and secretion of Hg2+ in rats treated with or without DMPS or DMSA. It should be reiterated that although Mrp2 is a likely participant in the secretion of DMPS S- or DMSA S-conjugates of Hg2+, one cannot rule out the participation of other mechanisms in this process. Only additional studies can resolve this possibility. Overall, however, the findings from the present study provide significant insights into the mechanisms by which DMPS or DMSA reduce the renal and body burden of Hg2+.
Acknowledgments
We thank Delon Barfuss (Georgia State University, Atlanta, GA) for providing the 203Hg2+.
This study was supported, in part, by the National Institutes of Health (National Institute of Environmental Health Sciences) Grants ES05157, ES05980, and ES11288 (to R.K.Z.) and ES015511 (to C.C.B.).
ABBREVIATIONS
- DMPS
2,3-dimercaptopropane-1-sulfonic acid
- DMSA
meso-2,3-dimercaptosuccinic acid
- Mrp
multidrug resistance protein.
References
- Aposhian HV. DMSA and DMPS: water soluble antidotes for heavy metal poisoning. Ann Rev Pharmacol Toxicol. 1983;23:193–215. doi: 10.1146/annurev.pa.23.040183.001205. [DOI] [PubMed] [Google Scholar]
- Aposhian HV, Aposhian MM. Meso-2,3-dimercaptosuccinic acid: chemical, pharmacological, and toxicological properties of an orally effective metal chelating agent. Annu Rev Pharmacol Toxicol. 1990;30:279–306. doi: 10.1146/annurev.pa.30.040190.001431. [DOI] [PubMed] [Google Scholar]
- Aposhian HV, Maiorino RM, Rivera M, Bruce DC, Dart RC, Hurlbut KM, Levine DJ, Zheng W, Fernando Q, Carter D, et al. Human studies with the chelating agents DMPS and DMSA. Clin Toxicol. 1992;30:505–528. doi: 10.3109/15563659209017938. [DOI] [PubMed] [Google Scholar]
- Aposhian HV, Maiorino RM, Gonzalez-Ramirez D, Zuniga-Charles M, Xu Z, Hurlbut KM, Junco-Munoz P, Dart RC, Aposhian MM. Mobilization of heavy metals by newer, therapeutically useful chelating agents. Toxicology. 1995;97:23–38. doi: 10.1016/0300-483x(95)02965-b. [DOI] [PubMed] [Google Scholar]
- Bahn A, Knabe M, Hagos Y, Rodiger M, Godehardt S, Graber-Neufeld DS, Evans KK, Burckhardt G, Wright SH. Interaction of the metal chelator 2,3-dimercapto-1-propoanesulfonate with the rabbit multispecific organic anion transporter 1 (rbOAT1). Mol Pharmacol. 2002;62:1128–1136. doi: 10.1124/mol.62.5.1128. [DOI] [PubMed] [Google Scholar]
- Belanger M, Westin A, Barfuss DW. Some health physics aspects of working with 203Hg in university research. Health Phys. 2001;80(Suppl 1):S28–S30. [PubMed] [Google Scholar]
- Bridges CC, Bauch C, Verrey F, Zalups RK. Mercuric conjugates of cysteine are transported by the amino acid transporter system b0,+: implications of molecular mimicry. J Am Soc Nephrol. 2004;15:663–673. doi: 10.1097/01.ASN.0000113553.62380.F5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bridges CC, Zalups RK. Molecular and ionic mimicry and the transport of toxic metals. Toxicol Appl Pharmacol. 2005;204:274–308. doi: 10.1016/j.taap.2004.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burckhardt BC, Drinkuth B, Menzel C, Konig A, Steffgen J, Wright SH, Burckhardt G. The renal Na+-dependent dicarboxylate transporter, NaDC-3 translocates dimethyl- and disulfhydryl-compounds and contributes to renal heavy metal detoxification. J Am Soc Nephrol. 2002;13:2628–2638. doi: 10.1097/01.asn.0000033463.58641.f9. [DOI] [PubMed] [Google Scholar]
- Chancy CD, Kekuda R, Huang W, Prasad P, Kuhnel JM, Sirotnak FM, Roon P, Ganapathy V, Smith SB. Expression and differential polarization of the reduced-folate transporter-1 and the folate receptor α in mammalian retinal pigment epithelium. J Biol Chem. 2000;275:20676–20684. doi: 10.1074/jbc.M002328200. [DOI] [PubMed] [Google Scholar]
- Cui Y, König J, Buchholz U, Spring H, Leier I, Keppler D. Drug resistance and ATP-dependent transport mediated by the apical multidrug resistance protein, MRP2, permanently expressed in human and canine cells. Mol Pharmacol. 1999;55:929–937. [PubMed] [Google Scholar]
- de Vries MH, Redegeld FAM, Koster AS, Noordhoek J, de Haan JG, Oude Elferink RPJ, Jansen PLM. Hepatic, intestinal and renal transport of 1-naphthol-β-d-glucuronide in mutant rats with hereditary-conjugated hyperbilirubinemia. Arch Pharmacol. 1989;340:588–592. doi: 10.1007/BF00260615. [DOI] [PubMed] [Google Scholar]
- Diamond GL, Klotzbach JM, Stewart JR. Complexing activity of 2,3-dimercapto-1-propanesulfonate and its disulfide auto-oxidation product in rat kidney. J Pharmacol Exp Ther. 1988;246:270–274. [PubMed] [Google Scholar]
- George GN, Prince RC, Gailer J, Buttigieg GA, Denton MB, Harris HH, Pickering IJ. Mercury binding to the chelation therapy agents DMSA and DMPS and the rational design of custom chelators for mercury. Chem Res Toxciol. 2004;17:999–1006. doi: 10.1021/tx049904e. [DOI] [PubMed] [Google Scholar]
- Institute of Laboratory Animal Resources . Guide for the Care and Use of Laboratory Animals. 7th ed. Institute of Laboratory Animal Resources, Commission on Life Sciences, National Research Council; Washington DC: 1996. [Google Scholar]
- Islinger F, Gekle M, Wright SH. Interaction of 2,3-dimercatpo-1-propane sulfonate with the human organic anion transporter hOAT1. J Pharmacol Exp Ther. 2001;299:741–747. [PubMed] [Google Scholar]
- Jansen PLM, Peters WH, Lamers WH. hereditary chronic conjugated hyperbilirubinemia in mutant rats caused by defective hepatic anion transport. Hepatology. 1985;5:573–579. doi: 10.1002/hep.1840050408. [DOI] [PubMed] [Google Scholar]
- Klotzbach JM, Diamond GL. Complexing activity and excretion of 2,3-dimercaptopropane sulfonate and other thiols by precolumn derivatization with bromabimane and column liquid chromatography. Am J Physiol. 1988;254:F871–F878. doi: 10.1152/ajprenal.1988.254.6.F871. [DOI] [PubMed] [Google Scholar]
- Leslie EM, Deeley RG, Cole SPC. Multidrug resistance proteins: role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol Appl Pharmacol. 2005;204:216–237. doi: 10.1016/j.taap.2004.10.012. [DOI] [PubMed] [Google Scholar]
- Maiorino RM, Dart RC, Carter DE, Aposhian HV. Determination and metabolism of dithiol chelating agents: XII. Metabolism and pharmokinetics of sodium 2,3-dimercaptopropane-1-sulfonate in humans. J Pharmacol Exp Ther. 1991;259:808–814. [PubMed] [Google Scholar]
- Masereeuw R, Notenboom S, Smeets PHE, Wouterse AC, Russel FGM. Impaired renal secretion of substrates for the multidrug resistance protein 2 in mutant transport-deficient (TR−) rats. J Am Soc Nephrol. 2003;14:2741–2749. doi: 10.1097/01.asn.0000094083.82845.fa. [DOI] [PubMed] [Google Scholar]
- Mayer R, Kartenbeck J, Buchler M, Jedlitschky G, Leier I, Keppler D. Expression of the MRP gene-encoded conjugate export pump in liver and its selective absence from the canalicular membrane in transport-deficient mutant hepatocytes. J Cell Biol. 1995;131:137–150. doi: 10.1083/jcb.131.1.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paulusma CC, Bosma PJ, Zaman GT, Bakker CT, Otter M, Scheffer GL, Scheper RJ, Borst P, Oude Elferink RP. Congenital jaundice in rats with a mutation in a multidrug resistance-associated protein gene. Science. 1996;271:1126–1128. doi: 10.1126/science.271.5252.1126. [DOI] [PubMed] [Google Scholar]
- peng K-C, Cluzeaud F, Bens M, Duong Van Huyen J-P, Wioland MA, Lacave R, Vandewalle A. Tissue and cell distribution of the multidrug resistance-associated protein (MRP) in mouse intestine and kidney. J Histochem Cytochem. 1999;47:757–767. doi: 10.1177/002215549904700605. [DOI] [PubMed] [Google Scholar]
- Planas-Bohne F. The effect of 2,3-dimercaptopropoane-1-sulfonate and dimercaptosuccinic acid on the distribution and excretion of mercuric chloride in rats. Toxicology. 1981;19:275–278. doi: 10.1016/0300-483x(81)90138-4. [DOI] [PubMed] [Google Scholar]
- Rivera M, Zheng W, Aposhian V, Fernando Determination and metabolism of dithiol chelating agents: VIII. Metal complexes of meso-dimercaptosuccinic acid. Toxicol Appl Pharmacol. 1989;100:96–106. doi: 10.1016/0041-008x(89)90094-x. [DOI] [PubMed] [Google Scholar]
- Ruprecht J. Scientific Monograph for Dimaval (DMPS) 6th ed Heyltex Corporation; Houston, TX: 1997. [Google Scholar]
- Schaub TP, Kartenbeck J, Konig J, Vogel O, Witzgall R, Kriz W, Keppler D. Expression of the conjugate export pump encoded by the mrp2 gene in the apical membrane of kidney proximal tubules. J Am Soc Nephrol. 1997;8:1213–1221. doi: 10.1681/ASN.V881213. [DOI] [PubMed] [Google Scholar]
- Smeets PHE, Van Aubel RAMH, Wouterse AC, Van Den Heuvel JJMW, Russel FGM. Contribution of multidrug resistance protein 2 (MRP2/ABCC2) to the renal excretion of p-aminohippurate (PAH) and identification of MRP4 (ABCC4) as a novel PAH transporter. J Am Soc Nephrol. 2004;15:2828–2835. doi: 10.1097/01.ASN.0000143473.64430.AC. [DOI] [PubMed] [Google Scholar]
- Stewart JR, Diamond GL. Renal tubular secretion of the alkanesulfonates 2,3-dimercapto-1-propane sulfonate. Am J Physiol. 1987;252:F800–F810. doi: 10.1152/ajprenal.1987.252.5.F800. [DOI] [PubMed] [Google Scholar]
- Stewart JR, Diamond GL. In vivo renal tubular secretion and metabolism of the disulfide of 2,3-dimercaptopropane-1-sulfonate. Drug Metab Dispos. 1988;16:189–195. [PubMed] [Google Scholar]
- Sugawara N, Lai YR, Sugaware C, Arizono K. Decreased hepatobiliary secretion of inorganic mercury, its deposition and toxicity in the Eisai hyperbilirubinemic rat with no hepatic canalicular organic anion transporter. Toxicology. 1998;126:23–31. doi: 10.1016/s0300-483x(97)00170-4. [DOI] [PubMed] [Google Scholar]
- Tanaka-Kagawa T, Naganuma A, Imura N. Tubular secretion and reabsorption of mercury compounds in mouse kidney. J Pharmacol Exp Ther. 1993;264:776–782. [PubMed] [Google Scholar]
- Van Aubel RA, Smeets Ph, Peters JG, Bindels RJ, Russel FG. The MRP4/ABCC4 gene encodes a novel apical organic anion transporter in human kidney proximal tubules: putative efflux pump for urinary cAMP and cGMP. J Am Soc Nephrol. 2002;13:595–603. doi: 10.1681/ASN.V133595. [DOI] [PubMed] [Google Scholar]
- Zalups RK. Influence of 2,3-dimercaptopropane-1-sulfonate (DMPS) and meso-2,3-dimercaptosuccinic acid (DMSA) on the renal disposition of mercury in normal and uninephrectomized rats exposed to inorganic mercury. J Pharmacol Exp Ther. 1993;267:791–800. [PubMed] [Google Scholar]
- Zalups RK. Molecular interaction with mercury in the kidney. Pharmacol Rev. 2000;52:113–143. [PubMed] [Google Scholar]
- Zalups RK, Barfuss DW, Kostyniak PJ. Altered intrarenal accumulation of mercury in uninephrectomized rats treated with methylmercury chloride. Toxicol Appl Pharmacol. 1992;115:174–182. doi: 10.1016/0041-008x(92)90321-i. [DOI] [PubMed] [Google Scholar]
- Zalups RK, Parks LD, Cannon VT, Barfuss DW. Mechanisms of action of 2,3-dimercaptopropane-1-sulfonate and the transport, disposition, and toxicity of inorganic mercury in isolated perfused segments of rabbit proximal tubules. Mol Pharmacol. 1998;54:353–363. doi: 10.1124/mol.54.2.353. [DOI] [PubMed] [Google Scholar]