

Abstract
Lipochitooligosaccharides (LCOs) are a novel class of plant growth regulators that activate in tobacco protoplasts the expression of AXI1, a gene implicated in auxin signaling. Transient assays with a chimeric PAXI-GUS expression plasmid revealed that the N-octadecenoylated monosaccharide GlcN has all structural requirements for a biological active glycolipid, whereas the inactive N-acylated GalN epimer inhibits LCO action. Specific inhibition of LCO and auxin action shows that both signals are transduced within the tobacco cell via separate pathways that converge at or before AXI1 transcription. Cytokinin is suggested to be a common effector of LCO and auxin signaling. We also show that activation of AXI1 correlates with growth factor-induced cell division.
Lipochitooligosaccharides (LCOs) secreted by rhizobia play a key role in the induction of nodules on the roots of legumes by activating signaling events that, at very low concentration, initiate cell division at distinct sites in the root (1, 2). Possibly these signals affect regulation of the plant cell cycle (3), and evidence is accumulating that LCOs play a general role as plant growth regulators (4–6). Previously we have developed methods to synthesize LCOs and demonstrated that these signaling molecules, which trigger nodulation in legumes, also promote cell division in tobacco protoplasts grown in the absence of auxin and cytokinin (7). Notably, glycolipid signals carrying fatty acids with double bonds in the trans configuration were efficient at femtomolar concentration to alleviate the requirement for micromolar concentrations of auxin and cytokinin to sustain growth of cultured tobacco protoplasts. LCOs increased the levels of AXI1 transcripts in protoplasts grown in the absence of auxin. The AXI1 gene is auxin- and cytokinin-responsive and its deregulated expression uncouples protoplasts from the normal effects of auxin on cell division (8).
In this study, we have investigated the minimum structural requirements for LCOs to induce cell division of tobacco protoplasts. We have used the AXI1 promoter in transient expression assays to show that multiple signaling pathways for AXI1 expression exist in tobacco cells.
MATERIALS AND METHODS
Synthesis and Purification of Glycolipids.
Chitooligosaccharides (Seikagaku Tokyo) were deacetylated at the nonreducing end using recombinant NodB protein as described (9). The primary amines were purified on a minicolumn of AG50 W-X4 (hydrogen form, Bio-Rad; ref. 10). Aminosugars (GalN and GlcN, Sigma) and the deacetylated chitooligosaccharides GlcN β-1,4-GlcNAc up to GlcN (β-1,4-GlcNAc)5 were N-acylated with trans-9-octadecenoic acid (Sigma) by a described procedure (11). Elaidic acid (51 μmol) was dissolved in acetonitrile (1 ml) and treated with triethylamine (56 μmol, ICN) and 2-chloro-1-methylpyridinium iodide (51 μmol, Sigma). The reaction was stirred under argon for 15 min at 30°C. To attach the fatty acid to the free amino group of the carbohydrate backbone, a solution of carbohydrate (17 μmol) in dimethyl sulfoxide (2 ml) was added to the reaction mixture. The mixture was stirred for 2 hr at 30°C under argon. Acylated products were obtained by HPLC purification on a preparative C18 column using an acetonitrile gradient as eluant. Depending on the length of the carbohydrate backbone, the yield varies between 15% and 30%.
Construction of the Expression Plasmid PAXI-GUS.
The expression plasmid PAXI-GUS (GUS, β-glucuronidase) contains the 2.5-kb promoter fragment of the Arabidopsis thaliana AXI1 gene (At AXI1a) fused to the GUS A reporter gene (12). The gene At AXI1a was isolated from an Arabidopsis λ Gem11 genomic DNA library using AXI1 cDNA from tobacco (8) as a probe. At AXI1a was subcloned into pBluescript (Stratagene), and the coding region was removed by digestion with NotI, which cleaves within the initiation codon and vector. The DNA fragment carrying the GUS gene together with the poly(A) addition sequence was released from pRT103GUS (13) by cutting with NcoI. To insert the reporter gene into the expression vector, all ends were filled with Klenow polymerase. Transient expression of PAXI-GUS induced by auxin mimics the auxin-responsive expression of the endogenous tobacco AXI1 as judged by Northern blot analysis (data not shown).
Protoplast Bioassay.
Protoplasts were isolated from tobacco (SR1) leaf tissue. Cell division assays were performed as described (7). Protoplasts were transfected with the expression plasmid PAXI-GUS using PEG 4000 treatment as described (14). Glycolipids dissolved in dimethyl sulfoxide were diluted in Jensen medium containing 1% dimethyl sulfoxide, and were added, at various concentrations, in a final volume of 50 μl to 3 × 105 transfected protoplasts (5 ml). Protoplasts were incubated for 48 hr, and transiently expressed GUS was determined as described (12).
RESULTS AND DISCUSSION
Effect of Carbohydrate Moiety on LCO Activity.
LCOs of rhizobial origin usually consist of a tetra- or pentameric backbone of β-1,4-linked GlcNAc residues (15). To study the effect of the carbohydrate chain length on the ability of LCOs to stimulate cell division of tobacco protoplasts, we N-acylated glucosamine and terminally deacetylated chitooligosaccharides (chitobiose to chitohexaose) with trans-9-octadecenoic acid. The structural identity of these synthetic glycolipids was confirmed by C18 reverse-phase HPLC, capillary GLC, and TLC (7). Exogenously supplied GlcN-containing glycolipids stimulated auxin-independent growth of cultured tobacco mesophyll protoplasts at nanomolar to femtomolar concentrations, depending on the chain-length of the carbohydrate moiety (Fig. 1A). The N-acylated monosaccharide GlcN stimulates maximal rates of cell division at 10−9 M and shows half-maximal activity at 10−12 M, indicating that additional β-1,4-linked GlcNAc residues in the carbohydrate moiety are not necessarily required for biological activity. Since the free fatty acid alone is not active in the cell division assay (7), the presence of a GlcN-containing carbohydrate residue in the signaling molecule is essential for mitogenic activity. In contrast, a GalN moiety that differs from GlcN only in the orientation of the hydroxyl group at C-4 renders the corresponding glycolipid biologically inactive (Fig. 1A). This suggests that a putative binding protein can distinguish between this single point of difference.
Figure 1.
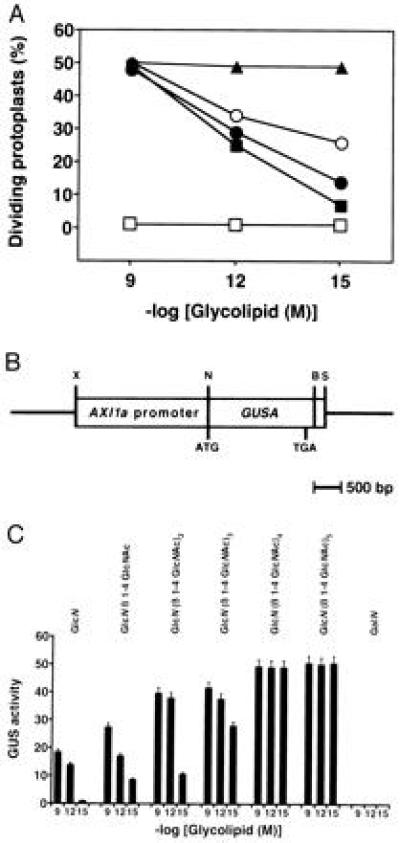
Effect of carbohydrate chain-length on the ability of synthetic glycolipids to stimulate division of tobacco protoplasts and to activate the AXI1 promoter from Arabidopsis in transient expression assays. The response of cultured protoplasts to glycolipids at concentrations of 10−9, 10−12, and 10−15 M was determined in the absence of auxin and in the presence of 1 μM kinetin. (A) Cell division assays. Carbohydrate moieties were N-acylated with trans-9-octadecenoic acid: GlcN (β-1,4-GlcNAc)3–5 (▴); GlcN (β-1,4-GlcNAc)2 (○); GlcN β-1,4-GlcNAc (•); GlcN (▪); GalN (□). Protoplast division assays were performed as described (7). Data shown are representative of three independent experiments performed in duplicate. (B) Structure of the PAXI-GUS chimeric gene used for the transfection of tobacco protoplasts. The AXI1a promoter was fused to the GUS A reporter gene. B, BamHI; N, NcoI; S, SacI; X, XhoI. (C) Transient GUS expression assays. Protoplasts were transfected with the expression plasmid containing the chimeric PAXI-GUS gene. GUS activity was measured in the cells after 48 hr as described (12). Data are expressed as nanomoles of 4-methyl-umbelliferone released per microgram of protein per hour and represent averages of three independent experiments. Carbohydrate moieties, N-acylated with trans-9-octadecenoic acid are indicated.
Recent studies have shown that LCOs activate the expression of AXI1 in tobacco protoplasts, conferring on these cells the ability to grow in the absence of auxin (7). AXI1 is believed to be an auxin-responsive transcriptional activator involved in transmission of mitogen signals to the nucleus (16). Since deregulated AXI1 expression uncouples tobacco cells from the normal effects of auxin on cell proliferation (8, 17), it is likely that this putative transcription factor plays an important role in auxin signaling. To monitor the effects of different growth factors on AXI1 expression, the reporter gene GUS A was fused to the AXI1a promoter from A. thaliana to form the chimeric PAXI-GUS expression plasmid (Fig. 1B). Tobacco protoplasts transfected with PAXI-GUS were incubated with different glycolipids at 10−9 M, 10−12 M, and 10−15 M concentration in the absence of auxin but in the presence of cytokinin. After 48 hr, the level of transiently expressed GUS was determined and found to increase with increasing chain-length of the carbohydrate backbone. However, while the N-acylated GlcN was still able to induce significant GUS activity, the elaidoylated GalN epimer was inactive (Fig. 1C). Together, the overall pattern of PAXI-GUS expression stimulated by different glycolipids correlates with the induction of cell division by the same signals (Fig. 1A).
LCO and Auxin Transduce Their Signals via Separate Pathways.
To examine the possibility that the N-acylated GalN can compete with GlcN-containing glycolipids in activating the AXI1a promoter, we tested this compound for its inhibitory effect in transient GUS expression assays. We therefore preincubated tobacco protoplasts, cultured in the presence of kinetin, with an excess of this potential inhibitory analog prior to adding N-acyl GlcN, N-acyl GlcN (β-1,4-GlcNAc)4, or auxin. N-Acyl GalN was efficient in inhibiting the glycolipid-activated expression of PAXI-GUS, whereas in the same assay the action of the auxin α-naphthalene acetic acid (1-NAA) was not affected (Fig. 2A). This observation suggests that in tobacco cells these GlcN-containing glycolipids use a signaling mechanism to induce AXI1 expression that is different from the auxin signaling pathway. This interpretation is further supported by the finding that in transient assays the anti-auxin β-naphthalene acetic acid (2-NAA; refs. 18 and 19) inhibits the action of 1-NAA, but not the ability of LCOs to induce the AXI1a promoter (Fig. 2B).
Figure 2.
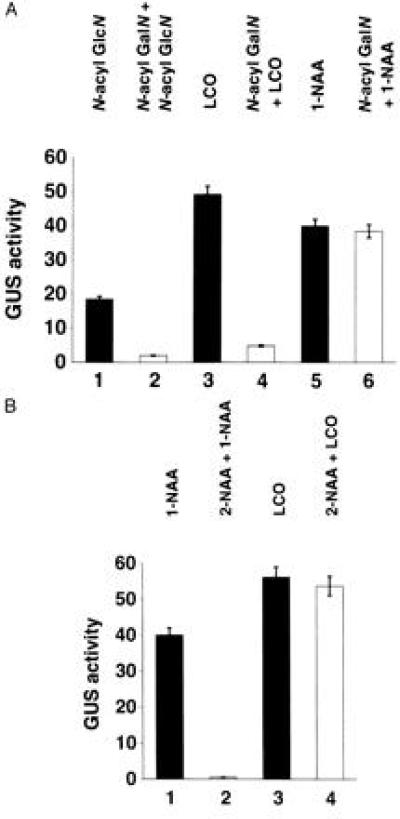
Different signaling pathways leading to the activation of AXI1 transcription can be selectively blocked by specific inhibitors. Tobacco protoplasts were transfected with the expression plasmid containing PAXI-GUS. AXI promoter activity was monitored by measuring the level of transiently expressed GUS (12). GUS activity is expressed as nanomoles of 4-methyl-umbelliferone released per microgram of protein per hour. (A) Effect of a GalN-containing glycolipid on LCO and auxin action. Cells grown in the presence of 1 μM kinetin were either untreated (solid bars) or treated with the inhibitor N-acyl GalN (open bars). Growth regulators in addition to kinetin are indicated on top. For the N-acyl GalN inhibition experiments, protoplasts were treated with this glycolipid for 20 min before addition of the different growth factors. Lanes: 1, N-acyl GlcN at 10−9 M; 2, N-acyl GalN at 10−8 M followed by N-acyl GlcN at 10−9 M; 3, N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M; 4, N-acyl GalN at 10−12 M followed by N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M; 5, 1-NAA at 5 × 10−6 M; 6, N-acyl GalN at 10−5 M followed by 1-NAA at 5 × 10−6 M. Data represent means of three independent experiments performed in duplicate. (B) Effect of 2-NAA on auxin and LCO action. Cells grown in the presence of 1 μM kinetin were treated either with (open bars) or without (solid bars; ref. 18) the anti-auxin 2-NAA. Growth regulators in addition to kinetin are indicated on top. Protoplasts were treated with 2-NAA for 20 min before addition of the different growth factors. Lanes: 1, 1-NAA at 5 × 10−6 M; 2, 2-NAA at 5 × 10−5 M followed by 1-NAA at 5 × 10−6 M; 3, N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M; 4, 2-NAA at 5 × 10−5 M followed by N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M. Data shown represent averages of three independent experiments.
Cytokinin Modulates the Level of AXI1 Induction.
In the absence of cytokinin, the phytohormone auxin is not sufficient to induce the AXI1a promoter (data not shown), and LCOs only weakly stimulate AXI expression in transient protoplast assays. Thus, cytokinin is additionally required in combination with auxin or LCOs to maximally stimulate PAXI-GUS expression (Fig. 3). To test this further and to obtain more information on cytokinin action, we used the cytokinin inhibitor 4-cyclobutyl-2-methylpyrrolo-[2,3-d]-pyrimidine (CBMPP) in transient assays (20). Exposure of transfected tobacco protoplasts to CBMPP specifically blocked cytokinin and reduced transiently expressed GUS to LCO- and auxin-induced levels. Furthermore, we found that treatment of transfected cells with the anti-cytokinin failed to block the activation of PAXI-GUS synergistically induced by auxin and LCOs (Fig. 3). It appears, therefore, that cytokinin may function as an effector necessary for LCOs and auxin to reach their full gene-activating ability. However, it is not clear how cross-talk with the effects of LCOs and auxin is mediated at the molecular level.
Figure 3.
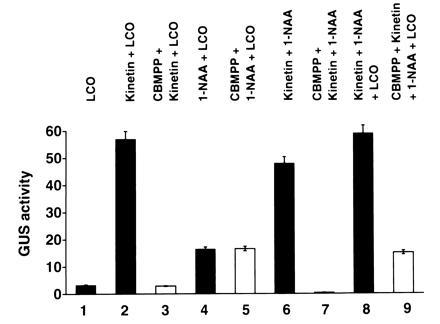
Effect of a cytokinin antagonist on LCO and phytohormone action. Transfected protoplasts were either untreated (solid bars) or treated with CBMPP (open bars; ref. 20). Growth regulators in culture medium are indicated on top. For the inhibition experiments, cells were pretreated for 20 min with the anti-cytokinin before the addition of the different growth regulators. Lanes: 1, N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M; 2, N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M and kinetin at 10−6 M; 3, anti-cytokinin CBMPP at 10−5 M followed by the addition of N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M and kinetin at 10−6 M; 4, N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M and 1-NAA at 5 × 10−6 M; 5, CBMPP at 10−5 M followed by N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M and 1-NAA at 5 × 10−6 M; 6, 1-NAA at 5 × 10−6 M and kinetin at 10−6 M; 7, CBMPP at 10−5 M followed by 1-NAA at 5 × 10−6 M and kinetin at 10−6 M; 8, N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M, 1-NAA at 5 × 10−6 M, and kinetin at 10−6 M; 9, CBMPP at 10−5 M followed by N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M, 1-NAA at 5 × 10−6 M, and kinetin at 10−6 M. Values are expressed as nanomoles of 4-methyl-umbelliferone released per microgram of protein per hour and represent averages of three independent experiments performed in duplicate.
Activation of AXI1 Correlates with Growth Factor-Induced Cell Division.
The finding that different mitogenic plant growth factors use distinct signaling pathways to activate AXI1 expression suggests that these pathways may also be involved in the control of cell division. Using the protoplast division assay, we could show that the inhibitors N-acyl GalN, 2-NAA, and CBMPP, which were used to inhibit the induction of AXI1, significantly affect LCO, auxin, and cytokinin also in their cell division-promoting activity (Fig. 4). This indicates that the multiple signaling pathways leading to the induction of AXI1 expression correspond to mitogenic signaling pathways. Although the exact role of AXI1 in stimulating mitogen-induced cell division remains to be defined, our results suggest that AXI1 participates in mitogenic signaling.
Figure 4.
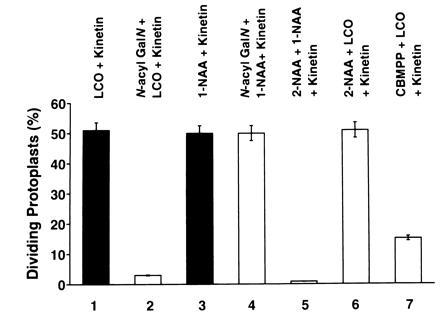
Effect of specific inhibitors on the ability of LCOs and phytohormones to induce cell division. Tobacco protoplasts were either untreated (solid bars) or treated with specific inhibitors (open bars) for 20 min before the addition of the different growth regulators. Growth regulators in culture medium are indicated on top. Protoplast bioassays were performed as described (7). Lanes: 1, N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M and kinetin at 10−6 M; 2, N-acyl GalN at 10−12 M followed by the addition of N-acyl GlcN(β-1,4-GlcNAc)4 at 10−15 M and kinetin at 10−6 M; 3, 1-NAA at 5 × 10−6 M followed by kinetin at 10−6 M; 4, N-acyl GalN at 10−5 M followed by 1-NAA at 5 × 10−6 M and kinetin at 10−6 M; 5, 2-NAA at 5 × 10−5 M followed by 1-NAA at 5 × 10−6 M and kinetin at 10−6 M; 6, 2-NAA at 5 × 10−5 M followed by N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M and kinetin at 10−6 M; 7, CBMPP at 10−5 M followed by N-acyl GlcN (β-1,4-GlcNAc)4 at 10−15 M and kinetin at 10−6 M. Data shown are means of two independent experiments performed in duplicate.
Multiple Signaling Pathways Regulate Cell Growth.
The emerging picture from our study is that tobacco cells have independent growth-promoting pathways for LCOs, auxin, and cytokinin leading to the activation of AXI1 expression. These pathways can be separately blocked by specific inhibitors. The inputs of LCO and auxin signals converge at a common target upstream, or at the level of AXI1 induction. Both signals, however, require the input from the cytokinin effector pathway for full mitogenic activity (Fig. 5). Although a number of proteins potentially involved in auxin signaling have been recently identified (21), little is known about how the extracellular LCO stimulus is converted into intracellular signals controlling events regulating cell growth. Since it is energetically favorable for LCOs to become embedded in lipid bilayers of cellular membranes, this might explain why these signals are active in one billion-fold lower amounts than auxin and cytokinin. Identification of the mechanism by which the LCO signal is perceived and subsequently propagated within the cell should provide insight how this novel class of plant growth regulators controls cell proliferation.
Figure 5.
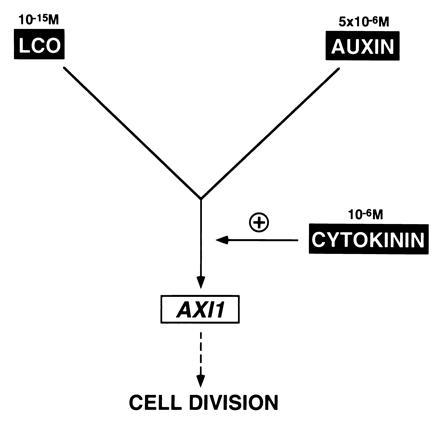
Schematic interpretation of the data. Induction of AXI1 by LCOs involves a signaling route independent from the auxin pathway. LCOs and auxin require the presence of cytokinin to stimulate maximal AXI transcription that correlates with cell division. The exogenously applied concentrations of growth factors are indicated.
Acknowledgments
We thank K. Palme for the cytokinin inhibitor; J. Dangl for providing a genomic DNA library from Arabidopsis; and A. Müller and A. Vente for helpful discussions. This work was supported by grants from Deutsche Forschungsgemeinschaft (H.L.) and the European Union (J. Schmidt).
Footnotes
Abbreviations: LCO, lipochitooligosaccharide; NAA, naphthalene acetic acid; GUS, β-glucuronidase; CBMPP, 4-cyclobutyl-2-methylpyrrolo-[2,3-d] pyrimidine.
References
- 1.Lerouge P, Roche P, Faucher C, Maillet F, Truchet G, Promé J C, Dénarié J. Nature (London) 1990;344:781–784. doi: 10.1038/344781a0. [DOI] [PubMed] [Google Scholar]
- 2.Truchet G, Roche P, Lerouge P, Vasse J, Camut S, de Billy F, Promé J C, Dénarié J. Nature (London) 1991;351:670–673. [Google Scholar]
- 3.Yang W-C, de Blank C, Meskiene I, Hirt H, Bakker J, van Kammen A, Franssen H, Bisseling T. Plant Cell. 1994;6:1415–1426. doi: 10.1105/tpc.6.10.1415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Schmidt J, Wingender R, John M, Wieneke U, Schell J. Proc Natl Acad Sci USA. 1988;85:8578–8582. doi: 10.1073/pnas.85.22.8578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.De Jong A J, Heidstra R, Spaink H P, Hartog M V, Meijer E A, Hendriks T, Lo Schiavo F, Terzi M, Bisseling T, Van Kammen A, De Vries S C. Plant Cell. 1993;5:615–620. doi: 10.1105/tpc.5.6.615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Schmidt J, Röhrig H, John M, Wieneke U, Stacey G, Koncz C, Schell J. Plant J. 1993;4:651–658. [Google Scholar]
- 7.Röhrig H, Schmidt J, Walden R, Czaja I, Miklasevics E, Wieneke U, Schell J, John M. Science. 1995;269:841–843. doi: 10.1126/science.269.5225.841. [DOI] [PubMed] [Google Scholar]
- 8.Hayashi H, Czaja I, Lubenow H, Schell J, Walden R. Science. 1992;258:1350–1353. doi: 10.1126/science.1455228. [DOI] [PubMed] [Google Scholar]
- 9.John M, Röhrig H, Schmidt J, Wieneke U, Schell J. Proc Natl Acad Sci USA. 1993;90:625–629. doi: 10.1073/pnas.90.2.625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Röhrig H, Schmidt J, Wieneke U, Kondorosi E, Barlier I, Schell J, John M. Proc Natl Acad Sci USA. 1994;91:3122–3126. doi: 10.1073/pnas.91.8.3122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Nicolaou K C, Bockovich N J, Carcanague D R, Hummel C W, Even L F. J Am Chem Soc. 1992;114:8701–8702. [Google Scholar]
- 12.Jefferson R A, Kavanaugh T A, Bevan M. EMBO J. 1987;6:3901–3907. doi: 10.1002/j.1460-2075.1987.tb02730.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Töpfer R, Pröls M, Schell J, Steinbiss H-H. Plant Cell Rep. 1988;7:225–228. doi: 10.1007/BF00272529. [DOI] [PubMed] [Google Scholar]
- 14.Walden R, Czaja I, Schmülling T, Schell J. Plant Cell Rep. 1993;12:551–554. doi: 10.1007/BF00233058. [DOI] [PubMed] [Google Scholar]
- 15.Dénarié J, Cullimore J. Cell. 1993;74:951–954. doi: 10.1016/0092-8674(93)90717-5. [DOI] [PubMed] [Google Scholar]
- 16.Walden R, Röhrig H, Schmidt J, John M, Schell J. In: Recent Advances in Plant Molecular Biology. Bansal K C, editor. India: Oxford and IBH Publishing Company; 1996. in press. [Google Scholar]
- 17.Walden R, Hayashi H, Lubenow H, Czaja I, Schell J. EMBO J. 1994;13:4729–4736. doi: 10.1002/j.1460-2075.1994.tb06798.x. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 18.Macdonald H, Jones A M, King P J. J Biol Chem. 1991;266:7393–7399. [PubMed] [Google Scholar]
- 19.Watahiki M K, Mori H, Yamamoto K T. Physiol Plant. 1995;94:566–574. [Google Scholar]
- 20.Iwamura H, Masuda N, Koshimizu K, Matsubara S. Phytochemistry. 1979;18:217–222. [Google Scholar]
- 21.Millner P A. Curr Opin Cell Biol. 1995;7:224–231. doi: 10.1016/0955-0674(95)80032-8. [DOI] [PubMed] [Google Scholar]