

Abstract
The parameters of the spontaneous deleterious mutation process remain poorly known, despite their importance. Here, we report the results of a mutation accumulation experiment performed on panmictic populations of Drosophila melanogaster without any genetic manipulations. Two experimental populations were kept for 30 generations under relaxed natural selection. Each generation, 100 pairs were formed randomly, and every fecund pair contributed a son and a daughter to the next generation. Comparison with two controls, one cryopreserved and the other kept as the experimental populations but with long generation time, showed that the number of surviving offspring per female declined by 0.2% and 2.0% per generation under benign and harsh, competitive conditions, respectively. Thus, the mutational pressure on fitness may be strong and depends critically on the conditions under which fitness is assayed.
The intensive spontaneous deleterious mutation process may be crucial for human genetics (1), conservation biology (2), maintenance of genetic variability at the molecular (3) and phenotypic (4) levels, and the evolution of reproduction (5). However, the relevant parameters are controversial, with evidence both for (6–10) and against (11–13) high genomic deleterious mutation rates in multicellular eukaryotes. The necessary data can be obtained by assaying the consequences of accumulation of mutations under relaxed selection. The results of mutation accumulation experiments (except in refs. 13 and 14, in which low mutation rates were reported) recently were questioned (12) because mutations accumulated and the fitness was measured in weak, genetically altered organisms, either deeply inbred or carrying balancer chromosomes, while adaptation in control populations, which would cause overestimation of the mutation rates, could not be ruled out.
Here, we report the results of an experiment that is free from these problems. Four fitness-related traits were measured after 10, 20, and 30 generations of relaxed selection in two experimental panmictic populations of Drosophila melanogaster and, simultaneously, in two controls. Without selection, every generation the mean fitness in an outbred population is decremented by the mutational pressure U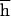 , and the average number of mutant alleles per genome is incremented by U, where U is the diploid genomic deleterious mutation rate and
, and the average number of mutant alleles per genome is incremented by U, where U is the diploid genomic deleterious mutation rate and 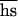 is the arithmetic mean of the product of dominance coefficient of a new heterozygous mutation h times coefficient of selection against a new homozygous mutation s. Our main objective was to estimate U
is the arithmetic mean of the product of dominance coefficient of a new heterozygous mutation h times coefficient of selection against a new homozygous mutation s. Our main objective was to estimate U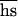 , and the rate of decline of the mean fitness is enough for this. In contrast, because the number of mutations in a genome cannot be assayed directly, measuring of U,
, and the rate of decline of the mean fitness is enough for this. In contrast, because the number of mutations in a genome cannot be assayed directly, measuring of U, 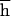 , and
, and 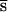 separately also must involve a much more difficult estimate of the increase of the variance in fitness, and the unknown variance of deleterious effects among new mutations must be somehow taken into account (6, 12). If U
separately also must involve a much more difficult estimate of the increase of the variance in fitness, and the unknown variance of deleterious effects among new mutations must be somehow taken into account (6, 12). If U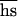 is indeed so low that relaxed selection causes no significant decline in the mean fitness (11–13), it makes little sense to measure the increase of its variance.
is indeed so low that relaxed selection causes no significant decline in the mean fitness (11–13), it makes little sense to measure the increase of its variance.
MATERIALS AND METHODS
Experimental Populations and Controls.
In July 1995, we sampled 160 mated females from a large population of D. melanogaster near Ithaca, NY. Their virgin offspring were placed in a cage. Samples of the eggs laid in this cage were cryopreserved (frozen control) using the procedure (15) that is apparently nonmutagenic (16). Two sets of 100 mated females were sampled from the cage. One experimental [Middle Class Neighborhood One (MCN-1)] and one control (cold control) population were started from the first set, and the other experimental population (MCN-2) was started from the second set. A sample from the same wild population obtained in August 1996 also was studied during the third measurement of fitness and was found to be very similar to the controls.
MCN-2, but not MCN-1 or cold control, carried sigma rhabdovirus, which causes hypersensitivity of flies to CO2. Sigma virus is transmitted vertically and occurs with low frequencies in many wild populations of D. melanogaster (17). Perhaps, it was present in one or few founders of MCN-2. During generations 15–25, the rate of sigma virus infection in MCN-2 grew from ≈5 to ≈70%.
Mutation Accumulation.
The experimental populations were kept under a 12/12 light cycle, 25°C, and 50% humidity in 95 × 25-mm vials with 10 ml of medium containing 7 g of agar, 50 g of brewer’s yeast, 60 g of sucrose, 20 g of corn flour, 2 ml of propionic acid, and 3 g of charcoal powder per 1 liter of water (optimal conditions). A tiny grain of live dry baker’s yeast was placed in a vial just before the flies were introduced. The flies were handled under CO2 anesthesia until generation 15 and under cold anesthesia after then. A generation lasted 11 days. On day 1, 100 pairs of the offspring from the previous generations were placed in separate vials. On day 2, the pairs were transferred individually into fresh vials (old vials were placed in 16°C and stored as back-ups). After 10–30 eggs were laid, the parents were discarded, and the eggs were counted. Virgin offspring from each vial were counted, and sexes were separated. On day 11, 1 female and 1 male were chosen randomly from each sibship (a few extra flies were randomly recruited to replace absent offspring from sterile vials), and 100 pairs were randomly formed. The cold control population was kept using the same breeding scheme at 16°C, where development took 25 days and virgin adults were stored for another 75 days. More details are available on request.
Reference Lines.
Two reference lines, marked with e (ebony) allele or with dke (dark eye) allele, were used to assay productivity under competitive conditions. These lines were created by backcrossing mutants, which occurred spontaneously, on flies from the same Ithaca wild population 6 times and were chosen from over 20 others because of high larval fitnesses. In the course of the experiment, the reference lines were kept at 16°C under high density with generation time ≈30 days.
Preparation of the Flies for Fitness Assays.
Extra offspring, not recruited for the next generation, were sampled from both experimental populations after 10, 20, and 30 generations. Simultaneously, the cold control that went through two, three, and four generations before the first, second, and third measurement of fitness, was sampled, and the frozen control flies appeared from the cryopreserved eggs activated 10 days earlier, with the recovery rates 18%, 11%, and 8% during the first, second, and third measurement, respectively. Each sample consisted of 100–200 virgin flies. To abolish possible difference of microflorae among the samples, they were rotated within the same set of vials so that every two samples used, in different days, the same food in the course of at least 1 day. After this, pairs were formed randomly within samples, and the flies laid eggs simultaneously. These eggs developed under low density (30–40 flies per vial) and optimal conditions into the intermediate generation, introduced to avoid maternal effects on fitness (18). At 5–7 days old, the flies were mated individually (either randomly within a sample or within sibships to produce inbred flies) and laid eggs from which “terminal” flies developed under low density and optimal conditions. Only cold anesthesia was used on terminal flies, and no anesthesia was used during 24 h before any fitness assay. Reference flies were produced and treated in the same way.
Productivity Under Benign Conditions.
The virgin females were kept (10–20 females per vial) during the first 3 days. On day 3, they were mated individually (outbred females were mated to unrelated males, and inbred females were mated to their double first cousins), and each pair was placed in a vial. At 1600 ± 1 h on day 5, each pair was transferred into the one experimental vial (95 × 28 mm), moved into the second experimental vial at 1600 ± 1 h on day 6, and discarded on day 7 at the same time. All of the flies were handled synchronously by different workers, and a pair spent 48 ± 0.5 h in two successive experimental vials. The pupae were counted 7–8 days later. Approximately 100 females of each type were used in each measurement.
Productivity Under Competitive Conditions.
Experiments of two types were performed. During the experiments of the first type, females were mated as above and kept for the first 4 days under optimal conditions and low density. On day 5, males were removed, and 6 or 12 of experimental or control females were placed in fresh vials together with the same numbers of mated reference females. On day 6, females from each vial were transferred to an experimental vial. With 12 or 24 females, standard vials were used with 10 ml of medium containing 10 g of agar, 20 g of brewer’s yeast, 20 g of sucrose, 10 g of corn flour, and 2 ml of propionic acid per 1 liter of water. The flies were discarded on day 7. Females spent 24 ± 0.5 h in an experimental vial. Live dry baker’s yeast (≈0.1 mg per female), consumed before the females were discharged, was placed in an experimental vial. These conditions were chosen to ensure tough competition (19). The offspring of both phenotypes were counted. During each measurement of fitness, 20 vials were used for each combination of the type of the experimental or control flies, reference line, and density.
After generation 30, we performed the experiment of the second type, to estimate variance components of the competitive ability in MCN-1 and in both controls (20, 21). Half-sib design was used, with random dam effect nested within random sire effect. Each sire was mated with five dams, which were 4–5 days old at the time of mating, in the successive days. After mating, each dam was paired with a mated e reference female, as above, and together they were placed into two successive experimental vials for 24 h in each and discarded afterward. Small (45 × 13 mm) vials were used with 0.75 ml of medium containing 10 g of agar, 17.5 g of brewer’s yeast, 17.5 g of sucrose, 7.5 g of corn flour, and 2 ml of propionic acid per 1 liter of water. Live dry baker’s yeast (≈0.2 mg) was placed in an experimental vial. There were 26, 61, and 65 sires in frozen control, cold control, and MCN-1, respectively.
Motility.
Males were kept with females under low density and optimal conditions for 3–4 days. After this, sets of 10 males were placed in separate vials. On days 6–8, escape response (22) was measured in these males. Each set was placed in a small compartment at the top of the experimental tube (500 × 40 ×40 mm), and 30 s later the tube was turned upside down, and the sliding wall separating the compartment from the rest of the tube was removed. The moments at which each male climbed 100 mm and 200 mm were recorded. Approximately 30 groups of males of each type were assayed during each measurement of fitness.
Longevity.
On day 2, 150 females and 150 males were placed in Plexiglas boxes (150 × 150 × 150 mm). Each box had one vial of food, replaced daily, and was kept under optimal conditions. Dead flies were removed and counted daily. Most measurements of longevity involved three boxes for each kind of flies.
Decline of Mean Fitness.
For each trait, the relative differences between its average value in an experimental population and the mean of the corresponding averages in the two controls were calculated, and their linear regressions on generation number, forced through zero, are reported below, together with their SEs. Regressions significantly different from zero (P < 0.01) are boldfaced.
RESULTS
Residual Selection.
Total opportunity for selection, the sum of mortality and infecundity rates, was ≈20% in both experimental populations (Fig. 1). Heritabilities of mortality, calculated through midparent regression (20), were 0.25 ± 0.07 in MCN-1 and 0.20 ± 0.07 in MCN-2 so that the genetic load was below 5%. The effective size of the experimental populations was 400 because each of 200 individuals contributed two offspring to the next generation (23). Thus, the inbreeding coefficient grew by 1/800 per generation, reaching ≈4% by the end of the experiment.
Figure 1.

Egg-to-imago mortalities in the experimental populations (MCN-1, solid lines; MCN-2, broken lines) and their linear regressions on generation number t (0.1506 + 0.0012t in MCN-1 and 0.1829 + 0.0021t in MCN-2). The data for generation 15 are absent because back-up vials were used. The infecundity rate was ≈1% in MCN-1 and ≈2% in MCN-2.
Fecundity.
Data on the number of surviving offspring per female (average of two days of measurement) are presented in Table 1. We call this trait “fecundity” because offspring mortality was low. Average per day fecundity of control flies exceeded 100, reflecting both high genetic quality of the flies and good experimental conditions. In generation 30, we also studied the fecundity of inbred females, obtained by brother–sister matings of the flies from MCN-1 or the frozen control, and mated to their double first cousins. Distributions of their fecundities, together with that on the fecundities of the corresponding outbred flies and of females obtained from the frozen control (female) × MCN-1 (male) cross, are presented in Fig. 2.
Table 1.
Fecundity
| Generation | Frozen control | Cold control | MCN-1 | MCN-2 |
|---|---|---|---|---|
| 10 | 105.04 ± 1.68 (4) | 103.90 ± 1.51 (6) | 96.68 ± 2.28 (7) | 92.99 ± 1.88 (4) |
| 20 | 97.53 ± 1.42 (2) | 88.62 ± 1.99 (1) | 91.06 ± 1.96 (4) | 90.47 ± 1.52 (3) |
| 30 | 105.30 ± 1.83 (7) | 108.15 ± 1.94 (4) | 99.69 ± 1.93 (6) | 93.68 ± 2.49 (3) |
| Regression coefficient | 0.00049 | −0.00049 | −0.00225 | −0.00376 |
| ±0.00044 | ±0.00050 | ±0.00056 | ±0.00054 |
Fecundities deviating from the mean by more than 3 SD (calculated for all nonsterile females) were discounted. The total numbers of discounted females are given in parentheses.
Figure 2.

Distributions of fecundities of the outbred and inbred frozen control females, outbred and inbred MCN-1 females, and females originated in the frozen control × MCN-1 cross in the third measurement of fitness-related traits.
Competitive Ability.
Logarithms of the ratios of the numbers of wild-type over reference offspring in the experiments of the first type are presented in Table 2. We call this trait “competitive ability” because it mostly reflects larval survival, which was only ≈10% due to intense competition. In each measurement, distribution of the competitive ability among the vials was close to Gaussian, and practically no vials had offspring of only one type. Fig. 3 shows the decline of mean fitness when the data with both reference lines are pooled. Overall, if the effects of density and of reference line are accounted for, competitive abilities in both controls were very similar (the regressions were 0.0014 ± 0.0026 for frozen control and −0.0014 ± 0.0026 for cold control), whereas the change of competitive ability was −0.021 ± 0.0032 and −0.024 ± 0.0027 per generation in MCN-1 and MCN-2, respectively.
Table 2.
Competitive ability
| Generation | Frozen control | Cold control | MCN1 | MCN2 |
|---|---|---|---|---|
| Dark eye reference line, Density 6:6 | ||||
| 10 | −0.332 ± 0.205 | −0.276 ± 0.115 | −0.480 ± 0.109 | −0.689 ± 0.112 |
| 20 | 0.618 ± 0.157 | 0.151 ± 0.266 | 0.581 ± 0.213 | −0.209 ± 0.114 |
| 30 | −0.926 ± 0.293 | −0.611 ± 0.175 | −1.468 ± 0.285 | −1.474 ± 0.163 |
| Regression coefficient | 0.0002 | −0.0002 | −0.0128 | −0.0264 |
| ±0.0060 | ±0.0052 | ±0.0059 | ±0.0035 | |
| Dark eye reference line, Density 12:12 | ||||
| 10 | −0.083 ± 0.152 | −0.165 ± 0.159 | −0.854 ± 0.189 | −0.561 ± 0.232 |
| 20 | 0.654 ± 0.153 | 0.414 ± 0.312 | 0.107 ± 0.155 | −0.337 ± 0.114 |
| 30 | −1.005 ± 0.177 | −1.258 ± 0.189 | −1.736 ± 0.283 | −2.058 ± 0.190 |
| Regression coefficient | 0.0044 | −0.0048 | −0.0248 | −0.0359 |
| ±0.0042 | ±0.0055 | ±0.0059 | ±0.0051 | |
| Ebony reference line, Density 6:6 | ||||
| 10 | 0.269 ± 0.161 | 0.248 ± 0.167 | 0.117 ± 0.142 | 0.264 ± 0.236 |
| 20 | 0.808 ± 0.189 | −0.041 ± 0.182 | 0.627 ± 0.154 | 0.146 ± 0.276 |
| 30 | −0.268 ± 0.153 | −0.192 ± 0.121 | −1.106 ± 0.326 | −1.307 ± 0.216 |
| Regression coefficient | 0.0049 | −0.0050 | −0.0165 | −0.0272 |
| ±0.0046 | ±0.0043 | ±0.0063 | ±0.0065 | |
| Ebony reference line, Density 12:12 | ||||
| 10 | 1.127 ± 0.161 | 0.962 ± 0.138 | 0.399 ± 0.194 | 0.903 ± 0.197 |
| 20 | 0.523 ± 0.249 | 0.203 ± 0.216 | 0.263 ± 0.264 | |
| 30 | −1.673 ± 0.255 | −1.030 ± 0.266 | −2.108 ± 0.249 | −1.513 ± 0.282 |
| Regression coefficient | −0.0037 | 0.0048 | −0.0293 | −0.0057 |
| ±0.0061 | ±0.0057 | ±0.0073 | ±0.0066 | |
The missing entry is due to an experimental error.
Figure 3.

Means, SEs, and linear regressions on the generation number of the differences between the competitive abilities of MCN-1 (solid lines) or MCN-2 (broken lines) and the average competitive ability of the two controls.
Table 3 presents the data from the experiment of the second type. Because many (44%) vials contained offspring of only one phenotype, the competitive ability of experimental or control flies was characterized by the ratio of the number of wild-type offspring to the total number of offspring. SEs were found through bootstrapping across males (21). If all of the offspring of a sire were pooled together, the average logarithms of the ratio of their number to that of the reference offspring in the same vials were 1.21 ± 0.25 in frozen control, 0.85 ± 0.12 in cold control, and 0.59 ± 0.13 in MCN-1, which is similar to what was observed in the first experiment.
Table 3.
Mean and variance components of competitive ability in small vials
| Frozen control | Cold control | MCN-1 | |
|---|---|---|---|
| Mean | 0.713 ± 0.048 | 0.632 ± 0.032 | 0.563 ± 0.032 |
| Sire | 0.026 ± 0.010 | 0.025 ± 0.021 | 0.018 ± 0.013 |
| Dam | 0.040 ± 0.034 | 0.059 ± 0.018 | 0.058 ± 0.018 |
| Residual | 0.063 ± 0.028 | 0.057 ± 0.014 | 0.067 ± 0.015 |
Motility.
Data on the speed (cm/s) of males running 100 mm upward are presented in Table 4 with group averages treated as independent observations. There was no difference between experimental and control males on 200-mm distance (data not reported).
Table 4.
Motility
| Generation | Frozen control | Cold control | MCN-1 | MCN-2 |
|---|---|---|---|---|
| 10 | 2.80 ± 0.065 | 2.62 ± 0.089 | 2.59 ± 0.085 | 2.44 ± 0.080 |
| 20 | 2.58 ± 0.088 | 2.55 ± 0.116 | 2.51 ± 0.084 | 2.44 ± 0.111 |
| 30 | 2.70 ± 0.131 | 2.72 ± 0.097 | 2.61 ± 0.082 | 2.70 ± 0.106 |
| Regression coefficient | 0.00067 | −0.00007 | −0.00187 | −0.00291 |
| ±0.00078 | ±0.00075 | ±0.00069 | ±0.00075 |
Longevity.
Average longevities of control females and males were 20.5 days and 35.9 days, respectively, and no significant mutational pressure on longevity was detected in the experimental populations (data not reported).
DISCUSSION
In agreement with others (11, 13), we have found little or no mutational pressure on the fitness-related traits assayed under benign noncompetitive conditions. In contrast, the mutational pressure on the competitive ability of larvae was substantial, ≈2% per generation (Table 2). In Mukai et al. (6), the mean viability of flies homozygous with second chromosomes sheltered from selection declined, relative to the mean viability of heterozygous flies, by ≈0.5% per generation (Fig. 2). If mutations with minor effects have h ≈ 0.4 (6, 24), this implies ≈2% per generation decline of fitness if mutations accumulate in the whole diploid genome because ≈40% of D. melanogaster genes are on the second chromosome. Thus, the agreement between our data and that from Mukai et al. (6) is quite remarkable, despite many differences in the respective experimental designs.
Because only a fraction of mutations affect larval performance under laboratory conditions (25), the mutational pressure on the total fitness in nature may be even larger than 2% per generation. The impact on fitness of homozygous deleterious alleles is magnified under harsh competitive conditions (19, 26–28), and the same is natural for heterozygous alleles because, in the struggle for existence, “any variation in the least degree injurious would be rigidly destroyed” (29). This may resolve the current controversy over the magnitude of the parameters of the spontaneous deleterious mutation process (30).
In contrast with other mutation accumulation experiments performed on D. melanogaster, we relaxed selection by purely demographical means and, therefore, studied vigorous genetically unmanipulated outbred flies. We used two different controls, both with rather limited opportunity for selection, which produced very similar results. One-time mortality of cryopreserved embryos was probably mostly random, and selection during a few generations in cold control was prevented demographically. Thus, the decline of competitive ability we observed can hardly be explained by adaptation of control flies. Rather weak residual selection in the experimental population, not assayed in any mutation accumulation experiments performed before, apparently can only restrict the accumulation of deleterious mutations and, thus, lead to underestimation of the mutation pressure.
Obviously, the decline of fitness we observed can be caused both by point mutations and by movements of transposable elements (31). Destruction of adaptive linkage disequilibria that might be present in the original natural population (32) could also contribute to this decline in the first few generations. However, such disequilibria are apparently quite rare. Because the performance of MCN-2 was only slightly below that of MCN-1, the impact of sigma virus on fitness seems to be small.
Marginally significant decline of motility (Table 4) is apparently the first observation of the mutational pressure on a behavioral trait. Some inbreeding depression of motility also was observed (average speeds were 2.52 ± 0.128 in inbred frozen control and 2.44 ± 0.130 in inbred MCN-1 males after 30 generations). No mutational pressure, as well as inbreeding depression (data not reported), was detected in longevity, indicating that it was not very sensitive to the genetic quality of the flies under the conditions used in our experiment.
We did not attempt to estimate the deviation from linearity in the decline of the competitive ability (epistasis) because the performance of reference flies, relative to both the controls and the experimental flies, improved with time (Table 2). In further experiments, reference populations should be kept, like controls, under conditions that preclude adaptation. The declines of fecundity and motility were too small to measure epistasis.
Although providing apparently robust estimates of U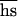 , our data can lead to only very tentative conclusions regarding the separate parameters. We detected (Table 3) no increase of the additive genetic variance in competitive ability, which is four times the sire variance component (20). This suggests that the decline of the mean is due to many mutations with small effects so that U is high. However, we can only exclude (with 95% confidence) the growth of additive variance that exceeds 0.04. Because the mean competitive ability declined by 0.1 in the same experiment, the maximal growth of the additive variance consistent with our data provides (6) the minimal estimate of U of only ≈0.01.
, our data can lead to only very tentative conclusions regarding the separate parameters. We detected (Table 3) no increase of the additive genetic variance in competitive ability, which is four times the sire variance component (20). This suggests that the decline of the mean is due to many mutations with small effects so that U is high. However, we can only exclude (with 95% confidence) the growth of additive variance that exceeds 0.04. Because the mean competitive ability declined by 0.1 in the same experiment, the maximal growth of the additive variance consistent with our data provides (6) the minimal estimate of U of only ≈0.01.
The increase of inbreeding depression in fecundity in the course of accumulation of mutations (Fig. 2) suggests that at least some new deleterious mutations are at least partially recessive (33). Although the mutational pressure on outbred fitness PO = U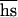 was 0.0019 ± 0.0013, this pressure on inbred fitness PI = U
was 0.0019 ± 0.0013, this pressure on inbred fitness PI = U 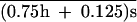 (with probability 0.125 flies produced by brother–sister mating are homozygous with either allele; no epistasis is assumed) was 0.0047 ± 0.0024. From this, we can estimate U
(with probability 0.125 flies produced by brother–sister mating are homozygous with either allele; no epistasis is assumed) was 0.0047 ± 0.0024. From this, we can estimate U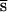 = 8(PI − 0.75PO) = 0.026 ± 0.023. If s and h were uncorrelated, comparison of PI and PO would also yield
= 8(PI − 0.75PO) = 0.026 ± 0.023. If s and h were uncorrelated, comparison of PI and PO would also yield 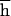 = 0.07 ± 0.069. However, because of negative covariance of h and s (24), this is an underestimate because
= 0.07 ± 0.069. However, because of negative covariance of h and s (24), this is an underestimate because 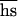 =
= 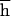
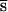 + Cov(h,s).
+ Cov(h,s).
High mutational pressures on fitness imply, in particular, that organisms from captive populations will soon become incapable of living in the wild. These pressures can be resisted if selection is allowed to operate in captivity (34), which is inconsistent with equal size of all families, the strategy recommended to minimize inbreeding (35). Also, the consequences of relaxed selection in civilized human populations may become significant after not so many generations (1).
Acknowledgments
We thank S. Caldwell and P. Steponkus for cryopreserving the flies, G. Gilchrist and R. Huey for suggestions on measuring fitness, C. McCulloch for advice on data analysis, C. Bergam, C. Hutter, F. Kondrashov, and N. Spiridonov for help with the experiments, and B. Charlesworth, D. Houle, P. Keightley, S. Nuzhdin, J. Peck, and an anonymous referee for useful suggestions. This project was supported by National Science Foundation Grant DEB-9531949.
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
Abbreviation: MCN, middle class neighborhood.
References
- 1.Crow J F. Proc Natl Acad Sci USA. 1997;94:8380–8386. doi: 10.1073/pnas.94.16.8380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lande R. Evolution. 1994;48:1460–1469. doi: 10.1111/j.1558-5646.1994.tb02188.x. [DOI] [PubMed] [Google Scholar]
- 3.Charlesworth B. Genet Res. 1996;68:131–149. doi: 10.1017/s0016672300034029. [DOI] [PubMed] [Google Scholar]
- 4.Houle D, Morikawa B, Lynch M. Genetics. 1996;143:1467–1483. doi: 10.1093/genetics/143.3.1467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hurst L D, Peck J R. Trends Ecol Evol. 1996;11:46–52. doi: 10.1016/0169-5347(96)81041-x. [DOI] [PubMed] [Google Scholar]
- 6.Mukai T, Chigusa S T, Mettler L E, Crow J F. Genetics. 1972;72:335–355. doi: 10.1093/genetics/72.2.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Neel J V, Satoh C, Goriki K, Fujita M, Takahashi N, Asakawa J, Hazama R. Proc Natl Acad Sci USA. 1986;83:389–393. doi: 10.1073/pnas.83.2.389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Johnston M O, Schoen D J. Science. 1995;267:226–229. doi: 10.1126/science.267.5195.226. [DOI] [PubMed] [Google Scholar]
- 9.Charlesworth D, Lyons E E, Litchfield L B. Proc R Soc Lond B. 1994;258:209–214. [Google Scholar]
- 10.Houle D, Hoffmaster D K, Assimacopoulos S, Charlesworth B. Nature (London) 1992;359:58–60. doi: 10.1038/359058a0. [DOI] [PubMed] [Google Scholar]
- 11.Fernandez J, Lopez-Fanjul C. Genetics. 1996;143:829–837. doi: 10.1093/genetics/143.2.829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Keightley P D. Genetics. 1996;144:1993–1999. doi: 10.1093/genetics/144.4.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Keightley P D, Caballero A. Proc Natl Acad Sci USA. 1997;94:3823–3827. doi: 10.1073/pnas.94.8.3823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kibota T T, Lynch M. Nature (London) 1996;381:694–696. doi: 10.1038/381694a0. [DOI] [PubMed] [Google Scholar]
- 15.Steponkus P L, Caldwell S. Cryo Lett. 1993;14:377–382. [Google Scholar]
- 16.Houle D, Kondrashov A S, Yampolsky L Yu, Caldwell S, Steponkus P L. Genet Res. 1997;69:209–213. doi: 10.1017/s0016672397002760. [DOI] [PubMed] [Google Scholar]
- 17.Wyers F, Petitjean A M, Dru P, Gay P, Contamine D. J Virol. 1995;69:4463–4470. doi: 10.1128/jvi.69.7.4463-4470.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mousseau T A, Dingle H. Ann Rev Entomol. 1991;36:511–534. [Google Scholar]
- 19.Kondrashov A S, Houle D. Proc R Soc Lond B. 1994;258:221–227. doi: 10.1098/rspb.1994.0166. [DOI] [PubMed] [Google Scholar]
- 20.Falconer D S. Introduction to Quantitative Genetics. 3rd ed. Harlow: Longman; 1989. [Google Scholar]
- 21.Searle S R, Casela G, McCulloch C E. Variance Components. New York: Wiley; 1992. [Google Scholar]
- 22.Engel J E, Wu C-F. J Neurosci. 1996;16:3486–3499. doi: 10.1523/JNEUROSCI.16-10-03486.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Crow J F, Kimura M. An Introduction to Population Genetics Theory. New York: Harper & Row; 1970. [Google Scholar]
- 24.Crow J F, Simmons M J. In: The Genetics and Biology of Drosophila. Ashburner M, Carson H L, Thompson N J Jr, editors. 3C. New York: Academic; 1983. pp. 1–35. [Google Scholar]
- 25.Hedrick P W, Murrai E. In: The Genetics and Biology of Drosophila. Ashburner M, Carson H L, Thompson N J Jr, editors. 3C. New York: Academic; 1983. pp. 61–104. [Google Scholar]
- 26.Scott T M, Koehn R K. J Exp Mar Biol Ecol. 1990;135:109–116. [Google Scholar]
- 27.Jimenez J A, Hughes K A, Alaks G, Graham L, Lacy R C. Science. 1994;266:271–273. doi: 10.1126/science.7939661. [DOI] [PubMed] [Google Scholar]
- 28.Ritland K. Genome. 1996;39:1–8. doi: 10.1139/g96-001. [DOI] [PubMed] [Google Scholar]
- 29.Darwin C. On the Origin of Species. London: Murray; 1859. [Google Scholar]
- 30.Peck J R, Eyre-Walker A. Nature (London) 1997;387:135–136. doi: 10.1038/387135a0. [DOI] [PubMed] [Google Scholar]
- 31.Nuzhdin S V, Pasyukova E G, Mackay T F C. Proc R Soc Lond B. 1996;263:823–831. doi: 10.1098/rspb.1996.0122. [DOI] [PubMed] [Google Scholar]
- 32.Zapata C, Alvarez G. Evolution. 1992;46:1900–1917. doi: 10.1111/j.1558-5646.1992.tb01177.x. [DOI] [PubMed] [Google Scholar]
- 33.Deng H-W, Lynch M. Genetics. 1996;144:349–360. doi: 10.1093/genetics/144.1.349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hedrick P W. Heredity. 1994;73:363–372. doi: 10.1038/hdy.1994.183. [DOI] [PubMed] [Google Scholar]
- 35.Borlase S C, Loebel D A, Frankham R, Nurthen R K, Briscoe D A, Daggard G E. Conserv Biol. 1993;7:122–131. [Google Scholar]