

Abstract
Structural variation (copy number variation [CNV] including deletion and duplication, translocation, inversion) of chromosomes has been identified in some individuals with autism spectrum disorder (ASD), but the full etiologic role is unknown. We performed genome-wide assessment for structural abnormalities in 427 unrelated ASD cases via single-nucleotide polymorphism microarrays and karyotyping. With microarrays, we discovered 277 unbalanced CNVs in 44% of ASD families not present in 500 controls (and re-examined in another 1152 controls). Karyotyping detected additional balanced changes. Although most variants were inherited, we found a total of 27 cases with de novo alterations, and in three (11%) of these individuals, two or more new variants were observed. De novo CNVs were found in ∼7% and ∼2% of idiopathic families having one child, or two or more ASD siblings, respectively. We also detected 13 loci with recurrent/overlapping CNV in unrelated cases, and at these sites, deletions and duplications affecting the same gene(s) in different individuals and sometimes in asymptomatic carriers were also found. Notwithstanding complexities, our results further implicate the SHANK3-NLGN4-NRXN1 postsynaptic density genes and also identify novel loci at DPP6-DPP10-PCDH9 (synapse complex), ANKRD11, DPYD, PTCHD1, 15q24, among others, for a role in ASD susceptibility. Our most compelling result discovered CNV at 16p11.2 (p = 0.002) (with characteristics of a genomic disorder) at ∼1% frequency. Some of the ASD regions were also common to mental retardation loci. Structural variants were found in sufficiently high frequency influencing ASD to suggest that cytogenetic and microarray analyses be considered in routine clinical workup.
Introduction
Autism (MIM 209850) is a neurodevelopmental disorder that manifests in the first three years of life. The group of pervasive developmental disorders (PDDs), also termed autism spectrum disorders (ASDs), includes autism as well as PDD-not otherwise specified (PDD-NOS) and Asperger's disorder. The three core characteristics of the ASDs are impairments of reciprocal social interactions, problems in communication, and a restricted range of behaviors and interests. Asperger's disorder differs from autism in that individuals have an absence of clinically significant cognitive and language delay. ASDs are observed in all populations with an incidence of 6 cases per 1000, with about four times more males diagnosed than females.1 The estimated prevalence of autism in siblings is 5%–10%.2
ASDs are etiologically heterogeneous. They are associated with a recognized cause in about 10% of cases, most commonly with fragile X (MIM 300624) and Rett syndrome (MIM 312750), tuberous sclerosis (MIM 191100), and other medical genetic conditions. Heritability estimates for ASDs, as determined from twin and family studies, are ∼90%,3 and linkage scans have mapped candidate risk loci.4
Based on a recent systematic review, cytogenetically detectable chromosome abnormalities are found in 7.4% (129/1749) of ASD cases with a range from 0% to 54%.5,6 The highest occurrence of events is observed in syndromic forms of ASD.6 Balanced translocations and inversions accounted for 17% (22/129) of rearrangements. Whereas the most frequent anomaly observed is maternally derived duplication of chromosome 15q11–q13 in 1%–3% of cases,7 little is known about the proportion of inherited compared to spontaneous karyotypic changes at other sites. With chromosome abnormalities as the initial step to identify ASD candidate loci, mutations have most convincingly been reported in SHANK3 on chromosome 22q13,8,9 two neuroligin (NLGN3 and NLGN4) genes on the X chromosome,10 and the neurexin 1 gene on chromosome 2p16.4,11,12 Recent data have also revealed that submicroscopic copy number variants (CNVs) can have a role in ASD,4,13 and de novo CNVs seem to be a more common risk factor in sporadic compared with familial forms of ASD.13,14
Our objective was to determine the potential contribution of all forms of genomic structural variation in ASD. We therefore used both high-resolution microarray technologies and karyotyping, allowing detection of both unbalanced and balanced as well as submicroscopic and cytogenetically visible structural variants. We also built a new Autism Chromosome Rearrangement Database (ACRD) allowing integration of ours and all other molecular information with the wealth of karyotypic data gathered over the years.15 We have identified several new candidate ASD-susceptibility loci, some of which encode proteins that are known to function within the synapse. Although the association of these variants with ASD is often compelling, there are many genotypic and phenotypic complexities. Our data, however, do indicate that high-resolution microarray analysis can be a powerful tool for clinical categorization and diagnosis in ASD.
Material and Methods
DNA Samples and Population Structure
The study included 427 ASD families (Table S1 available online). All cases met Autism Diagnostic Interview-Revised (ADI-R) and Autism Diagnostic Observation Schedule (ADOS) criteria on a clinical best estimate.16 Of these, 32 carried a cytogenetic chromosome rearrangement, 18 were detected through this study by karyotyping 313 of 413 idiopathic samples, and 14 were already known to carry karyotypic anomalies (Tables S1 and S2). Affected and unaffected siblings were also assessed, and 56% (237/427) had one child (simplex) and 44% (189/427) had more than one child (multiplex) with ASD. This ratio of multiplex to simplex is artificially elevated, reflecting our earlier interest in analyzing families in linkage studies.4 Most cases were screened for fragile X mutations (75%), and if detected, they were not included. Experiments were performed on blood DNA (80%) or otherwise lymphoblasts. Population ancestry was estimated via STRUCTURE17 (Table S1).
Affymetrix GeneChip Human Mapping 500K Array and Karyotyping Experiments
For each sample, approximately 500,000 SNPs were genotyped with the combined two-chip Affymetrix NspI and StyI GeneChip Human Mapping Commercial or Early Access Arrays according to standard protocols.9,18 Data have been submitted to the Gene Expression Omnibus database (accession GSE9222). Karyotypes were generated by standard clinical diagnostic protocols.
Characterization of Copy Number Variation
NspI/StyI array scans were analyzed for CNV content with a combination of DNA Chip Analyzer (dChip),4,19 Copy Number Analysis for GeneChip (CNAG),20 and Genotyping Microarray based CNV Analysis (GEMCA)21 (Table 1).
Table 1.
Summary of CNV in ASD and Controls
| Popgen Controls |
Autism Probands |
|||||
|---|---|---|---|---|---|---|
| All CNVs |
All CNVs |
Autism Specifica |
||||
| Full | Stringentb | Full | Stringentb | Full | Stringentb | |
| # Samples | 500 | 500 | 427 | 427 | 427 | 427 |
| # CNVs | 3695 | 1558 | 3396 | 1315 | 889 | 277 |
| CNV/Genomec | 7.4 | 3.1 | 8.0 | 3.1 | 2.1 | 0.65 |
| Mean/Median Size (kb) | 315/151 | 470/224 | 390/162 | 603/219 | 518/121 | 1082/194 |
| % Gain/Loss | 59/41% | 70/30% | 58/42% | 62/38% | 61/39% | 57/43% |
| Overlapping CNV/Loci (%)d | 3005/333 (81%) | 1226/142 (78%) | 2734/277 (80%) | 983/94 (74%) | 398/122 (44%) | 31/13 (11%) |
| >1 Mb CNV(%) | 343 (9%) | 250 (16%) | 339 (10%) | 212 (16%) | 63 (7%) | 32 (12%) |
Not seen in the initial 500 controls. There were 50 probands run in a previous study with the Affymetrix 10K array.4 The 10K array detected five different CNVs in five individuals. Of these putative CNVs, three were confirmed by the current 500K experiments, one was a false positive, and one had no probe coverage on the 500K platform so its status is unknown.
Stringent data set as called by >1 algorithms or arrays. Analysis with dChip was performed in batches of ∼100 probands. For CNAG version 2.0, we set the reference pool to include all samples and performed an automatic batch pair-wise analysis with sex-matched controls. For GEMCA, we used two designated DNA samples (NA10851 and NA15510) as references for pair-wise comparison to all proband experiments. We further filtered these results by including only those CNVs that were common to both pair-wise experiments. In all instances, CNVs were merged if they were detected in the same individual by more than one algorithm with the outside probe boundaries.
CNV/genome breakdown by algorithm: dChip Merged (3.0/genome), CNAG Merged (5.6/genome), GEMCA (5.5/genome). Validation experiments with q-PCR and FISH are described in the text. Another form of validation comes from examining the trios where we can demonstrate inheritance in 48 (maternal is 25, paternal is 23) of the autism-specific stringent data set. Also from the trios, 148 confirmed regions (inheritance assignment) in the stringent data set that overlap with controls (maternal is 65, paternal is 83).
Represents the total number of overlapping and/or recurrent CNVs, the number of overlapping/CNV loci, and the percentage of overlapping CNVs, out of the total data set.
Controls and the ACRD
Initial control samples consisted of (1) CNVs observed in 500 Europeans from the German PopGen project22 and (2) entries in the Database of Genomic Variants (containing 8006 CNVs at 3933 loci).23 A CNV was considered ASD specific if it was >10 kb, contained at least three probes, and at least 20% of its total length was unique when compared to the controls. To further test whether CNVs found in probands were specific to ASD, we also compared to CNV found in a cohort of 1152 nondisease controls of European origin from the Ontario population.24 In addition, we established the ACRD that had 834 putative CNVs or breakpoints mapped to the genome.
CNV Validation Experiments and Balanced Rearrangement Breakpoint Mapping
PCR validation of CNV calls was performed with Quantitative Multiplex PCR of short fluorescent fragments (QMPSF) or SYBR-Green I-based real-time quantitative PCR (qPCR) with controls at the ACCN1, CFTR, or FOXP2 loci. At least two independent qPCR assays were required for confirmation of a CNV. Balanced rearrangements were mapped primarily with FISH.25 The microdel program26 was used to score CNV losses.
Statistical Analysis
To assess the significance of the frequency of recurrent CNVs in ASD probands and controls, a 2-sided Fischer's exact test was used.
Results
Structural Variation Characteristics in ASD Cases
We initially tested 427 ASD probands for CNV content including 413 idiopathic cases (18 of which were found by karyotyping to have rearrangements) and 14 others that were enrolled based on prior knowledge of a cytogenetic abnormality (Table S1). We used the Affymetrix 500k SNP array because it provided the highest resolution screen available for both SNP genotype and CNV data. By using the SNPs, we could categorize the ancestry of the samples (to guide selection of controls) and found 90.3%, 4.5%, 4.5%, and 0.7% to have European, European/mixed, Asian, or African backgrounds, respectively.
To maximize CNV discovery, we used three calling algorithms (Figure S1) and merged sample-specific results between them to identify a “full” data set of 3396 independent CNVs (∼8 CNVs per genome, mean size 390 kb) (Table 1). To minimize potential false positives, we generated a second data set whereby a CNV needed to be detected by two or more algorithms and/or on both the NspI or StyI microarrays (see Figure S2 for representative data).27 This “stringent” data set contained 1315 CNVs (∼3 CNVs per genome, mean size 603 kb), of which we were able to validate 10/10 (5 deletions and 5 duplications) randomly selected CNVs via q-PCR. In total, we validated 97% (89/92) of CNVs tested in the stringent data set.
We then examined 500 European control samples for their CNV content and found similar numbers of CNVs (3695 in the full and 1558 in the stringent data set) to those in the ASD cases (Table 1). This suggested that germline chromosome instability was not a significant contributing mechanism. We also compared the ASD CNVs against the 500 European controls and the Database of Genomic Variants (a repository of structural variation in “nondisease” populations)23 to establish ASD-specific CNV data sets. Most of our subsequent analysis then focused on the 277 CNVs in our stringent autism-specific category, which mapped across all the 22 autosomes and X (but not Y) chromosome (Figure 1; Table S3). Additional ASD-relevant CNV data are also found in the other categories (Table 1; Table S2; and in the ACRD).
Figure 1.
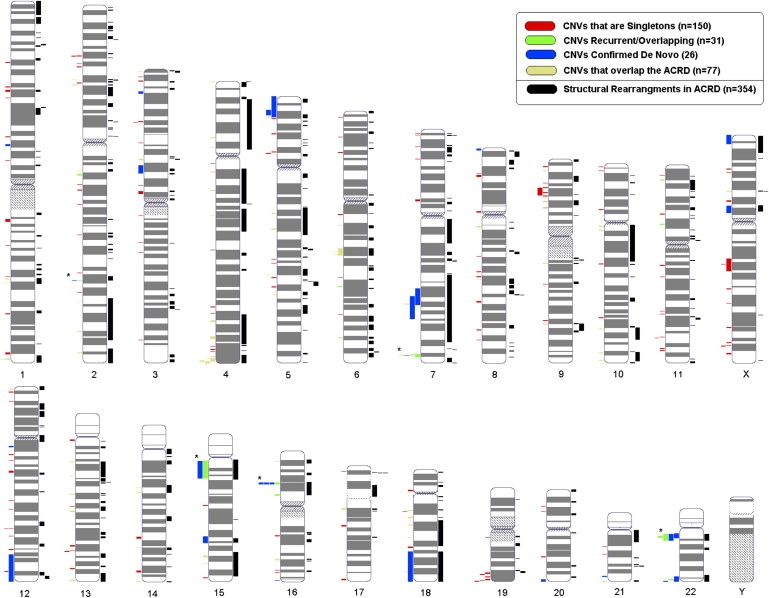
Genome-wide Distribution of CNVs
CNVs from the Autism Chromosome Rearrangement Database (ACRD) are plotted to the right of each chromosome (black). CNV data from the autism-specific stringent data set from the current study are shown to the left of the chromosome and is categorized as de novo (blue), overlapping/recurrent (green), CNVs overlapping with structural variation from the ACRD (yellow), and singleton CNVs (red). Note that five CNVs belong to both de novo and the recurrent categories and these are denoted by an asterisk (see Tables 2 and 3). All CNV data sets are described in Table 1 and the characteristics of the 277 CNVs depicted here are described in Table S3. The raw data from all are present in the Gene Expression Omnibus database.
Wide-ranging prevalence frequencies of cytogenetically detectable chromosomal abnormalities in ASD and the inability of microarray scans to find balanced abnormalities prompted us to also perform karyotyping. Karyotyping (and FISH) also provided the ability to characterize the chromosomal context (e.g., ring chromosomes) of some of the CNV regions, something not possible with microarrays alone. We therefore examined consecutive idiopathic cases where blood was available and found that 5.8% (18/313) cases had balanced (11) or unbalanced (7) karyotypes. We note that all unbalanced karyotypic changes described in this study were also found by microarray analysis and are included in the CNV statistics, and further that microarray analysis confirmed all balanced karyotypic changes (i.e., there were no additional gains or losses at the breakpoints below cytogenetic resolution). The genomic characteristics of all CNVs are shown in the ACRD (see Figure S3 as one example). We note that from our data, a CNV loss and gain will typically equate to a standard deletion or duplication, but they could also be more complex.18 In some cases, a duplication of only part of a gene could lead to its disruption, and there are also positional effects on gene expression to consider.28
De Novo, Overlapping/Recurrent, and Inherited Structural Variants
We initially prioritized structural variants found in ASD cases to possibly be etiologic if they were not in our initial control set and (1) de novo in origin (27 cases) (Table 2), (2) overlapping (27 cases at 13 loci) in two or more unrelated samples (Table 3), (3) recurrent (same breakpoints) in two or more unrelated samples (four cases at two loci) (Table 3), or (4) inherited (the remainder). We found CNVs at previously reported ASD loci: NLGN4 and 22q11.2, 15q11–q13, SHANK3, and NRXN1 in categories 1, 2, 3, and 4, respectively. ASD structural variants found in controls (e.g., NRXN1) could also be involved.
Table 2.
Twenty-Seven ASD Cases with De Novo Rearrangements
| FamID | Sex | Typea | Chromosomeb | Size (bp)c | CNV | Genesd | Phenotype Comments | |
|---|---|---|---|---|---|---|---|---|
| 1 | SK0181-004 | M | CHR (SPX) | 3p14.1-p13 (a) | 5,346,900 | loss | 13 genes | IQ/LOFe 78, language mod delay,f speech 2+,g RB2+,h dysmorph 3+,i congenital unilateral sensorineural hearing loss |
| t(6;14)(q13;q21) (k) | N/A | none | 11 genes | |||||
| 2 | SK0152-003 | M | CHR (MPX) | 3p25.1-p24.3 (a) | 1,409,600 | loss | 12 genes | LOF 31, RL/ELj mod/sev delay, speech 3+, RB2+, dysmorph 0, hypotonia affecting FM & GM development |
| 5p15.31-p15.2 (a) | 3,429,389 | loss | 8 genes | |||||
| 12q12 (a) | 422,842 | loss | 4 genes | |||||
| t(5;7)(p15p13) (k) | N/A | none | CDH18 | |||||
| 3 | SK0215-006 | M | CHR (SPX) | 1p21.3 (a) | 1,092,500 | loss | DPYD whole | LOF 31, language sev delay, RB3+, dysmorph N/A |
| 4 | SK0205-004 | F | CHR (SPX) | 5p15.33-p15.2 (k) | 13,800,984 | loss | 46 genes | IQ/LOF & language mild delay, RB1+, dysmorph 3+, Cri du Chat syndrome |
| 5 | SK0083-003 | M | CHR (SPX) | 7q31.1-q31.31 (k) | 11,023,507 | loss | 25 genes | IQ/LOF 56, language sev, speech 3+, RB1+, dysmorph 3+, febrile seizures |
| 6 | SK0131-003 | F | CHR (SPX) | 7q31.1-q32.2 (k) | 15,486,722 | loss | >50 genes | IQ/LOF 74, RL/EL mod/sev delay, speech 2+, RB2+, dysmorph 3+, microcephaly |
| 7 | SK0243-003 | M | CHR (SPX) | 15q23-q24.2 (k) | 4,289,500 | loss | >50 genes | LOF severely impaired, language sev delay, RB2+, dysmorph 3+, severe scoliosis, hiatal hernia |
| 8 | SK0073-003 | F | CHR (SPX) | 15q11.2-q13.3 (k) | 11,922,600 | gain | >50 genes | IQ/LOF 49, RL/EL mod delay, RB3+, dysmorph N/A, premature (34 wks), hypotonia |
| 9 | SK0245-005 | M | CHR (SPX) | 15q11.2-q13.3 (k) | 11,871,747 | gain | >50 genes | IQ/LOF 47, RL/EL sev delay, RB3+, dysmorph N/A |
| 10 | SK0218-003 | F | CHR (MPX) | 18q21.32-q23 (k) | 20,358,999 | loss | >50 genes | LOF severely impaired, RL/EL nonverbal, RB3+, dysmorph 3+, seizures, microcephaly, strabismus, hypotonia, cleft palate, club feet |
| 11 | NA0039-000 | F | CHR (SPX) | 22q13.31-q13.33 (k) | 3,231,700 | loss | 41 genes | LOF < 40, language sev delay, RB0, dysmorph 2+, hypotonia, mega cisterna magna, submucous cleft palate, single umbilical artery; father has balanced reciprocal translocation (14;22), proband inherited der(22), sister with ADHD also has unbalanced karyotype with der(14) |
| 12 | NA0097-000 | F | CHR (SPX) | Xp22.33- p22.31 (a) | 5,825,311 | loss | 21 genes+NLGN4 | IQ/LOF 117, language average, speech 2+, RB1+, dysmorph 0, carries maternal balanced reciprocal 11;12 translocation |
| 13 | SK0283-003 | F | CHR (SPX) | 47,XX, ring chr1 (k) | N/A | gain | >50 genes | IQ/LOF 30, RL/EL sev delay, RB2+, dysmorph 3+, microcephaly, blood dyscrasia, failure to thrive |
| 14 | SK0195-003 | M | CHR (SPX) | t(5;8;17)(q31.1;q24.1;q21.3) (k) | N/A | none | 5 genes | IQ/LOF unknown, language nonverbal, RB2+, dysmorph 3+, bifid uvula |
| 15 | SK0306-004 | F | SPX | 2q32.1 (a) | 97,130 | loss | None | IQ/LOF 68, RL/EL mild delay, speech 2+,i RB2+, dysmorph 3+, severe hypotonia |
| 16 | NA0002-000 | M | SPX | 7q36.2 (a) | 66,462 | loss | DPP6 exonic | IQ/LOF unknown, RL/EL sev delay, RB3+, dysmorph 0 |
| 17 | SK0262-003 | M | SPX | 8p23.3 (a) | 791,089 | gain | DLGAP2 exonic | IQ/LOF 68, RL/EL mod delay, RB4+, dysmorph N/A |
| 18 | MM0278-003 | M | SPX | 12q24.21-q24.33 (a) | 18,218,000 | gain | >50 genes | IQ/LOF 24, language sev delay, speech 1+, RB3+, dysmorph 1+, seizures; scoliosis, lower limb anomaly |
| 19 | NA0067-000 | M | SPX | 16q24.3 (a) | 265,667 | loss | ANKRD11 exonic | IQ/LOF unknown, language mod delay, RB3+, dysmorph 2+, motor delay, alloimmune ITP |
| 20 | MM0088-003 | F | MPX | 16p11.2 (a) | 675,829 | loss | 28 genes | IQ/LOF 82, RL/EL mod delay, RB3+, dysmorph 0 |
| 21 | SK0102-004 | M | SPX | 16p11.2 (a) | 432,600 | gain | 24 genes | IQ/LOF 39, RL/EL sev delay, speech 2+, RB3+, dysmorph 2+, epilepsy, scoliosis, diaphragmatic hernia |
| 22 | SK0019-004 | M | SPX | 16p11.2 (a) | 675,829 | loss | 28 genes | IQ/LOF 93, RL/EL average, speech 2+, RB3+, dysmorph 1, hyperphagia and severe obesity |
| 23 | SK0244-003 | M | SPX | 21q22.3 (a) | 353,936 | gain | 4 genes | IQ/LOF 80, RL/EL mild delay, RB1+, dysmorph N/A |
| 24 | MM0109-003 | F | SPX | 20q13.33 (a) | 1,427,661 | gain | 44 genes | IQ/LOF 27, language nonverbal, RB3+, dysmorph 1+ |
| 22q13.33 (a) | 276,702 | loss | 13 genes+SHANK3 | |||||
| 25 | SK0119-003k | M | MPX | 22q11.21 (a) | 2,771,300 | loss | >50 genes | IQ/LOF 77, RL/EL mod/sev delay, dysmorph 3+, velocardiofacial syndrome |
| 26 | SK0297-003 | M | SPX-MZ | 22q11.21 (a) | 4,281,262 | gain | >50 genes | IQ/LOF 75, RL/EL average, speech 1+, RB3+, dysmorph 2+, seizures, MZ twin (discordant for ASD) |
| 27 | SK0306-004 | F | SPX | Xp11.23-p11.22 (a) | 4,643,367 | gain | >50 genes | IQ/LOF 68, RL/EL mild delay, speech 2+, RB2+, dysmorph 3+, severe hypotonia |
Probands with abnormal karyotypes (CHR) (1–14) are separated from probands belonging to simplex (SPX) and multiplex (MPX) families with normal karyotypes (15–27). Families are grouped and sorted based on simplex (SPX), multiplex (MPX), and chromosomal abnormalities (CHR). Simplex families with affected monozygotic twins are denoted as SPX-MZ. Some cases here are also recurrent and appear in Table 3 and some of the family pedigrees are shown in Figure 2 and Figure S2. For multiplex families, the de novo events were not detected in affected siblings.
De novo event detected by either karyotype (k) or microarray (a).
All de novo CNV detected by the array were validated with qPCR, whereas translocations and larger deletions/duplications detected by karyotyping were confirmed by FISH. In all cases where an unbalanced change occurs, CNV size is based on array results. The breakpoints have not been accurately defined, and CNVs may be smaller or larger than posted.
If the CNV intersects only a single gene (suggesting that it may disrupt the gene), the term “exonic” is used, and if the CNV encompasses the entire gene, the term “whole” is used. The term “intronic” is used for CNV that overlaps noncoding parts of a single gene.
IQ/LOF, level of functioning denotes average of Vineland Social, Communication, and daily living scores and nonverbal IQ, when available.
Language was rated as average, nonverbal, mild, mod (moderately delayed), or sev (severely delayed).
Speech refers to the severity of impaired speech intelligibility, most likely resulting from oral motor apraxia (1+, mild; 2+, moderate; 3+, severe unintelligibility; or 0, intelligible speech).
RB: repetitive behavior score was derived from ADI and ADOS ratings (1+, mild; 2+, moderate; 3+, severe repetitive behaviors; or 0, no repetitive behaviors).
Dysmorphology scores were based on anthropometric measurement abnormalities and qualitative features documented either by a clinical geneticist or a developmental pediatrician. Anomalies were reviewed by a single clinical geneticist and assigned score: 0, not dysmorphic; 1+, mild; 2+, moderate; 3+, severe degree of dysmorphism. Children with known genetic syndromes received a score of 3+. N/A, not assessed for dysmorphic features.
RL, receptive language; EL, expressive.
SK0119-003 originally entered the study with an ASD diagnosis but upon re-examination after CNV detection was assessed to be below cutoffs for ASD.
Table 3.
Recurrent and Overlapping Loci in ASD
| Chromosome | FamID | Sex | Typea | Size (bp)b | CNV | Origin | Genesc | Phenotype Comments | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2q14.1 | SK0147-003 | F | SPX | 478,370 | loss | paternal | DPP10 exonic | IQ/LOFd unknown, RL/ELe sev delay, RB2+,f dysmorph 3+,g neurofibromatosis type 1, abnormal EEG |
| SK0288-003 | F | SPX-MZ | 105,120 | gain | paternal | DPP10 intronic | IQ/LOF 73, RL/EL average/mild delay, RB 2+, dysmorph N/A | ||
| 2 | 2q32.1 | SK0306-004 | F | SPX | 97,130 | loss | de novo | none | IQ/LOF 68, RL/EL mild delay, speech 2+,h RB2+, dysmorph 3+, severe hypotonia |
| NA0030-000 | M | SPX | 112,323 | loss | maternal | none | IQ/LOF 69, RL/EL mod delay, RB2+, dysmorph 1+, seizures, medicated for OCD, motor incoordination | ||
| 3 | 6q22.31 | MM0220-003 | M | MPX | 318,000 | gain | paternal | PLN whole | IQ/LOF 82, language average, dysmorph 2+, high myopia, fine motor difficulties |
| NA0025-000 | M | SPX | 293,989 | gain | paternal | PLN whole | IQ/LOF unknown, language: regression/mild delay, RB2+, dysmorph 0 | ||
| 4 | 7q36.2 | SK0190-003 | M | SPX | 1,780,000 | gain | maternal | DPP6 whole | IQ/LOF 55, RL/EL sev delay, speech 2+, RB1+, dysmorph N/A, parents 1st cousins |
| SK0115-003 | M | SPX | 274,000 | gain | unknown | DPP6 exonic | IQ/LOF 86, RL/EL average, RB2+, ysmorph N/A, | ||
| SK0058-003 | M | MPX | 16,788 | gain | maternal | DPP6 intronic | IQ/LOF 111, RL/EL average, RB1+, ysmorph N/A | ||
| NA0002-000 | M | SPX | 66,462 | loss | de novo | DPP6 exonic | IQ/LOF unknown, RL/EL severe delay, RB3+, dysmorph 0 | ||
| 5 | 8q11.23 | SK0143-003 | M | SPX | 285,200 | gain | unknown | UNQ9433 whole, RB1CC1 exonic | IQ/LOF 65, RL/EL sev delay, speech 2+, RB2+, dysmorph 2+, seizures, hypoplastic left heart syndrome, left hemidiaphragm paralysis |
| MM0236-004 | M | MPX | 271,679 | gain | unknown | RB1CC1 exonic | IQ/LOF 84, language average, RB1+, dysmorph 0, central auditory processing difficulty | ||
| 6 | 9p24.1 | SK0270-003 | M | SPX | 38,900 | loss | unknown | none | IQ/LOF 67, RL/EL mild/mod delay, speech 1+, RB3+, dysmorph N/A |
| MM0103-003 | M | MPX | 34,950 | loss | paternal | none | IQ/LOF 100, language mild delay, speech 0, RB2+, dysmorph 0, twin preg (other twin lost in 1st trimester), premature (34 wks with respiratory distress syndrome), club feet | ||
| 7 | 11p12 | MM0272-003 | M | MPX | 262,938 | loss | maternal | none | IQ/LOF 74, language mild delay, speech 3+, RB2+, dysmorph N/A, seizures, unilateral congenital ptosis |
| SK0167-003 | F | MPX | 192,846 | loss | unknown | none | IQ/LOF 62, RL/EL average/mild delay, speech apraxia 2+, dysmorph N/A | ||
| 8 | 13q21.32 | SK0023-003 | M | SPX | 189,438 | gain | unknown | PCDH9 intronic | IQ/LOF 82, RL/EL mod/sev delay, speech 2+, RB2+, dysmorph N/A; seizures |
| MM0299-003 | F | MPX | 172,401 | gain | paternal | PCDH9 intronic | IQ/LOF 43, language nonverbal, RB2+, dysmorph 0, hypotonia, gags/chokes, fine motor delay | ||
| 9 | 15q11.2-q13.3 | SK0073-003 | F | CHR | 11,922,600 | gain | de novo | >50 genes | IQ/LOF 49, RL/EL mod delay, RB3+, dysmorph N/A, premature (34 wks) |
| SK0245-005 | M | CHR | 11,871,747 | gain | de novo | >50 genes | IQ/LOF 47, RL/EL sev delay, RB3+, dysmorph N/A | ||
| 10 | 16p12.2 | MM0109-003 | F | SPX | 1,246,288 | gain | maternal | 8 genes | IQ/LOF 27, language nonverbal, RB3+, dysmorph 1+ |
| MM0289-003 | F | MPX | 802,555 | loss | maternal | 5 genes | IQ/LOF 45, language delay, RB2+, dysmorph 0 | ||
| 11 | 16p11.2 | NA0133-000 | F | SPX | 525,319 | gain | maternal | 29 genes | IQ/LOF & language moderate delay, speech 2+, RB1+, dysmorph 0, early motor delay |
| SK0102-004 | M | SPX | 432,600 | gaini | de novo | 24 genes | IQ/LOF 39, RL/EL sev delay, speech 2+, RB3+, dysmorph 2+, epilepsy, congenital diaphragmatic hernia | ||
| MM0088-003 | F | MPX | 675,829 | loss | de novo | 32 genes | IQ/LOF 82, RL/EL mod delay, RB3+, dysmorph 1+ | ||
| SK0019-004 | M | SPX | 675,829 | loss | de novo | 32 genes | IQ/LOF 93, RL/EL average, speech 2+, RB3+, dysmorph 1+, hyperphagia and severe obesity | ||
| 12 | 22q11.2 | SK0119-003j | M | MPX | 2,771,300 | loss | de novo | >50 genes | IQ/LOF 77, RL/EL mod/sev delay, dysmorph 3+, velocardiofacial syndrome |
| SK0091-004 | F | MPX | 4,281,262 | gain | paternal | >50 genes | IQ/LOF 92, RL/EL average/mod delay, RB3+, dysmorph 0, placental insufficiency | ||
| SK0297-003 | M | SPX-MZ | 4,281,262 | gain | de novo | >50 genes | IQ/LOF 74, language average; RB3+, dysmorph 2+, seizures, MZ twin (discordant for ASD) | ||
| SK0323-003 | M | MPX | 743,100 | gain | unknown | 7 genes | IQ/LOQ 43, language sev delay, RB4+, dysmorph N/A | ||
| 13 | 22q13.31 | SK0123-004 | M | MPX | 601,528 | gain | maternal | none | IQ 93, language mod delay, RB4+, dysmorph 0, nonidentical triplet, Hirschsprung disease |
| MM0102-003 | M | MPX | 80,380 | loss | maternal | none | IQ/LOF 60, language mild delay, dysmorph 0, weakness & hypotonia of arms |
Probands are ordered by chromosome location. Families are grouped based on simplex (SPX), multiplex (MPX), and chromosomal abnormalities (CHR). Simplex families with affected monozygotic twins are denoted as SPX-MZ. The de novo cases also appear in Table 2 and some of the family pedigrees are shown in Figure 2 and Figure S2.
All recurrent and overlapping CNVs were detected by the array validated with qPCR. The breakpoints have not been accurately defined, and CNVs may be smaller or larger than noted.
If the CNV intersects only a single gene (suggesting that it may disrupt the gene), the term “exonic” is used, and if the CNV encompasses the entire gene, the term “whole” is used. The term “intronic” is used for CNV that overlaps noncoding parts of a single gene.
IQ/LOF (level of functioning) denotes average of Vineland Social, Communication and daily living scores and nonverbal IQ, when available.
Language (RL, receptive language; EL, expressive language) was rated as average, nonverbal, or mild, mod (moderately), or sev (severely) delayed.
RB (repetitive behavior) score was derived from ADI and ADOS ratings (1+, mild; 2+, moderate; 3+, severe repetitive behaviors; or 0, no repetitive behaviors).
Dysmorphology scores were based on anthropometric measurement abnormalities and qualitative features documented either by a clinical geneticist or a developmental pediatrician. Anomalies were reviewed by a single clinical geneticist and assigned score 0, not dysmorphic; 1+, mild; 2+, moderate; 3+, severe degree of dysmorphism. Children with known genetic syndromes received a score of 3+. N/A, not assessed for dysmorphic features.
Speech refers to the severity of impaired speech intelligibility, most likely resulting from oral motor apraxia (1+, mild; 2+, moderate; 3+, severe unintelligibility).
CNV is called by only one algorithm.
SK0119-003 originally entered the study with an ASD diagnosis but upon re-examination after CNV detection was assessed to be below cutoffs for ASD.
By testing random families, a de novo CNV rate of 7.1% (4/56) and 2.0% (1/49) was observed in our idiopathic simplex and multiplex families, respectively, similar to rates in another project.13 This is likely a minimum value because many smaller CNVs will be beyond detection with the current technology. The literature13,29 and our previous work18 have shown a <1% spontaneous CNV mutation rate in nondisease samples. Regarding balanced alterations, there was parental information for 13 of 18 cases discovered to carry cytogenetic anomalies (Table S1), and 7 (6 simplex, 1 multiplex) of these were de novo in origin. Because only 1/7 (from a simplex family) was balanced and directly interrupting a gene, we estimate that this class of rearrangements had much less of a contribution than CNVs to the total rate of de novo and inherited structural variation in our ASD cohort.
Our collective data identified 27 de novo cases (Table 2), and in three of these, two or more events were identified. Notably in family SK0152 (Figure 2A), there were four de novo events. In MM0109 (Figure 2B), there were two de novo CNVs, one leading to haploinsufficiency of SHANK3.
Figure 2.
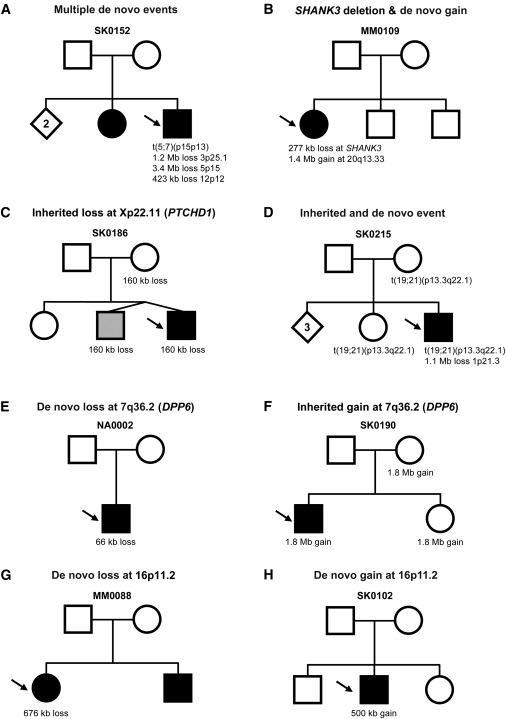
Examples of Complexities of Structural Variants Observed in ASD Families
Males are denoted by squares and females by circles. The size of each de novo or inherited event is shown below each family member. Arrows identify the proband, open shapes are unaffected, and filled have ASD diagnosis (gray denotes developmental delay but not a definitive ASD diagnosis). Diamonds indicate number of older unaffected siblings of unspecified gender. For ASD cases, probands may have multiple de novo events (A), including rearrangements overlapping genes known to be associated with ASD such as SHANK3 (B). Probands may also inherit chromosome X deletions (at PTCHD1) from female carriers (C) or have an inherited translocation in addition to an unrelated de novo deletion (D). Overlapping events in unrelated probands may be either de novo (E) or inherited (F) at the DPP6 locus. Recurrent de novo events in unrelated probands may also be either losses (G) or gains (H) at chromosome 16p11.2. Additional ASD families with CNVs at DPP6 and 16p11.2 are shown in Figures S4 and S5, respectively.
The 13 loci where overlapping ASD-specific CNVs were found are more likely to have a role in ASD susceptibility because they arise in two or more unrelated families. In six loci, gains and losses were observed at the same locus (Table 3), suggesting that general gene dysregulation may be involved. In those cases with shared CNVs not overlapping with a known medical genetic locus (see below), phenotypic re-examination yielded no obvious genotype-phenotype correlation beyond a trend for larger sporadic abnormalities to generally have a more complex clinical presentation. We also note that in the “full” data set, there are 397 overlapping CNVs at 122 loci, among which other bona fide candidate ASD loci may exist.
By using q-PCR or by assessing SNP patterns, we confirmed 196 inherited CNVs (90 maternal and 106 paternal). No subgrouping of these demonstrated obvious parent-of-origin effects (the two chromosome 15q11–q13 duplications detected were both de novo in origin). We did detect a 160 kb deletion in a male inherited from a carrier mother, leading to a null PTCHD1 in the proband and his dizygotic twin brother (Figure 2C). We also found instances where apparently balanced inherited translocations were accompanied by de novo deletions elsewhere in the genome in the offspring (e.g., DPYD; Figure 2D).
Candidate ASD-Susceptibility Genes and Loci Identified
To further validate our findings, we compared our de novo and recurrent/overlapping CNVs from ASD cases to an additional 1152 matched controls. This sample consists of samples from Ontario, Canada, of European origin.24 We analyzed these in an identical manner to our ASD data and were unable to find CNV matching identically to the 27 de novo cases nor to the 13 recurrent/overlapping ASD cases, further emphasizing the potential importance of these regions. There was, however, one new CNV overlapping the DPP6 locus found in this control set (see below).
New ASD candidates identified were those with a structural change (either de novo, found in two or more unrelated ASD cases, or for the X chromosome an allele being transmitted maternally from an unaffected carrier) specific to that gene, including ANKRD11, DLGAP2, DPP6, DPP10, DPYD, PCDH9, and PTCHD1 (Tables 2 and 3). As previously mentioned, NLGN4, SHANK3, and NRXN1 were also identified. The PCDH9 and NRXN1 genes are also found as CNVs in controls in the Database of Genomic Variants. This suggests that there could also be other important CNVs that are not included in the 277 ASD-specific variants on which we based most of our analyses.
Additional positional candidate genes identified were those found interrupted by balanced cytogenetic breakpoints including NEGR1, PIP5K1B, GABRG1, KLHL3, STK3, ST7, and SATB2 (Table S2). Moreover, 77 CNVs in our stringent data set overlapped with the ACRD, providing a second line of evidence for involvement (Figure 1). For example, a 4.6 Mb de novo duplication at Xp11.23–11.22 was detected in a female SK0306-004 (Table 2) and a male in the database.30 There is a wealth of other data that needs further analysis at the population level, such as the finding of an ∼100 kb deletion at 2q37.3 (previously shown to be involved in ASD6,7 that occurs in twice the frequency in ASD cases compared to controls) (Table S2).
DPP6 and DPP10 emerge as being positional and functional candidates. DPP6 (∼1.5 Mb in size at 2q14.1) and DPP10 (∼1.3 Mb at 7q36.2) code for accessory transmembrane dipeptidyl peptidase-like subunits that affect the expression and gating of Kv4.2 channels (KCND2). Kv4.2 channels function in regulation of neurotransmitter release and neuronal excitability in the glutamatergic synapse31 at the same sites where SHANK3 and the NLGN gene products are found. Moreover, we have mapped autism balanced breakpoints near KCND2 at 7q31 (see the ACRD).25
For DPP10, there are inherited CNV gains and losses (Table 3; Figure S4). De novo and inherited CNVs were found at the multitranscript DPP6 gene (CNV in 4/427 ASD probands versus 1/1652 controls [initial 500 controls plus 1152 additional controls; Fischer exact test p value = 0.016]). A 66 kb de novo loss encompassing exons 2 and 3 is found in a male in family NA0002 (Figure 2E). In family SK0190, the male proband and an unaffected female sibling both carry a CNV gain inherited from an unaffected mother (Figure 2F) that encompassed the entire DPP6. A 270 kb gain is found in SK0115-003 that extends across the first exon (which may disrupt the functional gene), and SK0058-003 carries a maternally inherited 16 kb intronic CNV gain (Figure S4).14
Medical Genetics
We identify structural variants overlapping loci involved in medical genetic conditions including Waardenburg Type IIA (MIM 193510; 3p14.1), speech and language disorder (MIM 602081; 7q31), mental retardation (MR) (15q23–q24, 16p11.2), and velocardialfacial syndrome (VCFS) (MIM 192430; 22q11.2), among others (Tables 2 and 3; Table S2). Identification of the structural variant at these loci led to clinical reassessment and either identification or refinement of the diagnosis, for additional syndromic features. Other instances (e.g., SK0186-PTCHD1 deletion) (Figure 2C) prompted retesting of the entire family and eventually a diagnosis of mild ASD in a previously undiagnosed sibling. This family would therefore move from a simplex to multiplex designation, underscoring the need for cautious interpretation of our statistics in any medical genetic application.
The identification of a de novo deletion (2.7 Mb) at 22q11.2 in two brothers with ASD led to their re-examination and diagnosis for VCFS. The retesting also further defined the siblings to be at opposite ends of the ASD spectrum (Figure S5). Larger duplications (4.3 Mb) of this same region in two other ASD families (SK0289 and SK0091) do not cause VCFS (Table 3). However, in SK0091, the variant was inherited from a normal father and was not found in an affected male sibling. Other studies also reveal differential phenotypes, some having ASD associated with different duplications and deletions at this locus.32
We also discovered a recurrent ∼500 kb duplication at 16p11.2 in two ASD families (SK0102 and NA0133) (Figure 2; Figure S5). The same region was recently shown to be involved in mental retardation, aortic valve development, and seizure disorder.33 As with DPP6/DPP10 and 22q11.2, there are carriers of these structural variants without ASD. At the same loci, we also detected a recurrent 676 kb de novo deletion in two ASD families (MM0088 and SK0019). Family SK0019 is a simplex family with the deletion in the proband (Figure S5), whereas in family MM0088, the deletion is detected in only one of two ASD siblings (Figure 2G). This brings a combined discovery rate of 4/427 16p11.2 CNVs in ASD and 0/1652 controls (2-sided Fischer's exact test p value = 0.002). We note that the 16p11.2 deletions and duplications may be reciprocal events, and we are testing this by fine-mapping the breakpoints.
Discussion
Our genome-wide analysis of structural variation yields many new results indicating that chromosomal abnormalities have a substantial role in ASD. In light of these and other findings,4,13,34 we recommend that genomic analysis, in particular with high-resolution microarrays, be used in research studies of ASD and possibly in clinical assessment once we understand what the implications of these variants are. Such structural variation data could initially serve to focus clinical examination in a search for undetected syndromes. In cases where the loci detected show high penetrance such as haploinsufficiency of SHANK38,9 and 16p11.2, clinical diagnostic testing could be considered.
In our cohort, we observed that ∼7% of randomly selected idiopathic cases harbor de novo genomic CNV rearrangements that are detectable by the microarray platform we used (including ASD cases with balanced translocation or inversion changes that affect genes increases the number slightly). Of these, we found that ∼11% of cases actually carry two or more de novo events. Moreover, in other families (and in some cases in the general population), there were non-ASD carriers of the variants, as well as ASD siblings without the variant (Figure 2). These observations, coupled with the newly recognized abundance of structural variants in the genome35 having a comparatively high new mutation rate,29 cautions against assigning causation to all de novo rearrangements found. Certainly in some instances additional independent risk factors need to be considered, including those being genetic, epigenetic, environmental, or stochastic in origin.36 Our finding of X-linked CNVs being maternally transmitted to males and that most structural variants detected on X are inherited from parents may contribute to explain gender prevalence differences and the high degree of heritability in ASD.
With the genome-wide scanning approach, we have identified numerous new putative ASD loci (Tables 2 and 3; Figure 1). Three themes that emerge include the identification of (1) loci that often contain genes functioning in the postsynaptic density (PSD), (2) and/or chromosomal regions previously shown to be involved in mental retardation, and (3) new loci where general dysregulation of gene expression may be involved.
First, we found CNVs that further implicate the SHANK3, NLGN4, and NRXN1-PSD genes and also identify others including DPP6 and DPP10 (and PCDH9, RPS6KA2, RET from the larger data set). Neuroligins have been shown to bind to the SHANK3 scaffolding protein and are required for the maturation of glutamatergic synapses. Moreover, the neurexins are ligands of the neuroligins, thus supporting a potential pathogenic construct of a network of interrelated molecules, from the trans-synaptic interaction between the neurexins and neuroligins to the PSD complex that includes SHANK3. Given that DPP6 and DPP10 are gating channels regulating neurotransmitter release, a functional complex emerges, in which perturbations at a number of potential PSD molecules acting alone or in combination may lead to ASD.37,38
Second, we identified at least two ASD loci (15q24, 16p11.2) that overlap with known MR sites,33,39,40 suggesting that some of the genes causing MR also cause autism (we also found inherited CNVs at three other MR loci: IDS, IL1RAPL1, and TSPAN7). We did not detect structural variants at other chromosome loci associated with anomalies in syndromic forms of ASD such as 7q11.23 (MIM 609757) and 17p11.2 (MIM 610883), most likely because we limited our cases to more strictly defined ASD.
The 16p11.2 CNV region is particularly interesting because it is found at near to 1% frequency in our autism cohort and not controls. It also exhibits characteristics of a genomic disorder, including being flanked by a pair of segmental duplications with >99% identity, which likely mediate the deletion and duplication events through nonallelic homologous recombination (Figure S3). We found both duplication and deletions associated with ASD at this locus. Although no obvious genotype-phenotype correlation is so far obvious in the four families we have identified (Figure S5B), the deletion may be more penetrant because some unaffected individuals carry the duplication. Twenty-four genes map to the CNV interval, including five postsynaptic density (PSD) genes (DOC2A, ALDOA, MAPK3, CORO1A, and CDIPT) and two candidates for seizure (SEZ6L2 and QPRT). The fact that both duplications and deletions exert phenotype suggests that some genes in the interval are sensitive to dosage.
Third, we discovered six unrelated ASD cases that had either CNV gains or losses in unrelated individuals at the same locus (including 16p11.2) (Table 3). This suggests that fine tuning of gene expression of critical genes in these regions can be crucial for the development of speech and language and/or social communication, as seems to be the case for SHANK38,9 and genes in the Williams-Beuren syndrome locus (MIM 194050).41 In some cases, duplication of the Williams-Beuren syndrome region also exhibits an ASD-like phenotype (MIM 609757).
Our understanding of the full etiologic role of structural variation in ASD will require genomic and phenotypic analyses of more cases (and their families) and population controls.42 To achieve the desired numbers, integration of multiple data sets will likely be necessary; hence, our establishing the ACRD resource as a first step. From our current data, it is already apparent that for a proportion of individuals, it will be possible to describe their ASD based on the underlying structural characteristics of their genome. The detection rate of structural variation and ability to discern the associated outcome will only increase as new higher-resolution genome-scanning methods become available and accessible.
Supplemental Data
Supplemental data include five figures and three tables and can be found with this article online at http://www.ajhg.org/.
Supplemental Data
Web Resources
The URLs for data presented herein are as follows:
The Autism Chromosome Rearrangement Database, http://projects.tcag.ca/autism/
The Database of Genomic Variants, http://projects.tcag.ca/variation/
Gene Expression Omnibus, http://www.ncbi.nlm.nih.gov/geo/
Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/sites/entrez?db=OMIM
Accession Numbers
Microarray data have been deposited in the Gene Expression Omnibus database under accession GSE9222.
Acknowledgments
We thank Lili Senman, Carolyn Noakes, Michelle Lee, Beata Stachowiak, Kathy Whitten, Barbara Noble, and The Centre for Applied Genomics for technical assistance. The work is supported by Genome Canada/Ontario Genomics Institute, the Canadian Institutes of Health Research (CIHR), the McLaughlin Centre for Molecular Medicine, the Canadian Institute of Advanced Research, Autism Speaks, the McMaster Children's Hospital Foundation, the Hospital for Sick Children (SickKids) Foundation, and the German National Genome Research Network (NGFN). C.R.M. is supported by the SickKids Foundation and the National Alliance for Research on Schizophrenia and Depression (NARSAD). L.F. is supported by Canadian Institute for Health Research (CIHR) Fellowship. D.P. is supported by the Netherlands Organization for Scientific Research and the Royal Netherlands Academy of Arts and Sciences. L.Z. is supported by a Health Scholar award from the Alberta Heritage Foundation for Medical Research and is a CIHR New Investigator. S.W.S. is an Investigator the GlaxoSmithKline-CIHR Pathfinder Chair in Genetics and Genomics at SickKids and the University of Toronto.
References
- 1.Chakrabarti S., Fombonne E. Pervasive developmental disorders in preschool children: confirmation of high prevalence. Am. J. Psychiatry. 2005;162:1133–1141. doi: 10.1176/appi.ajp.162.6.1133. [DOI] [PubMed] [Google Scholar]
- 2.Jones M.B., Szatmari P. Stoppage rules and genetic studies of autism. J. Autism Dev. Disord. 1988;18:31–40. doi: 10.1007/BF02211816. [DOI] [PubMed] [Google Scholar]
- 3.Bailey A., Le Couteur A., Gottesman I., Bolton P., Simonoff E., Yuzda E., Rutter M. Autism as a strongly genetic disorder: evidence from a British twin study. Psychol. Med. 1995;25:63–77. doi: 10.1017/s0033291700028099. [DOI] [PubMed] [Google Scholar]
- 4.Szatmari P., Paterson A.D., Zwaigenbaum L., Roberts W., Brian J., Liu X.Q., Vincent J.B., Skaug J.L., Thompson A.P., Autism Genome Project Consortium (AGPC) Mapping autism risk loci using genetic linkage and chromosomal rearrangements. Nat. Genet. 2007;39:319–328. doi: 10.1038/ng1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Xu J., Zwaigenbaum L., Szatmari P., Scherer S.W. Molecular cytogenetics of autism. Curr. Genomics. 2004;5:347–364. [Google Scholar]
- 6.Vorstman J.A., Staal W.G., van Daalen E., van Engeland H., Hochstenbach P.F., Franke L. Identification of novel autism candidate regions through analysis of reported cytogenetic abnormalities associated with autism. Mol. Psychiatry. 2006;11:18–28. doi: 10.1038/sj.mp.4001781. [DOI] [PubMed] [Google Scholar]
- 7.Veenstra-Vanderweele J., Christian S.L., Cook E.H. Autism as a paradigmatic complex genetic disorder. Annu. Rev. Genomics Hum. Genet. 2004;5:379–405. doi: 10.1146/annurev.genom.5.061903.180050. [DOI] [PubMed] [Google Scholar]
- 8.Durand C.M., Betancur C., Boeckers T.M., Bockmann J., Chaste P., Fauchereau F., Nygren G., Rastam M., Gillberg I.C., Anckarsater H. Mutations in the gene encoding the synaptic scaffolding protein SHANK3 are associated with autism spectrum disorders. Nat. Genet. 2007;39:25–27. doi: 10.1038/ng1933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Moessner R., Marshall C.R., Sutcliffe J.S., Skaug J., Pinto D., Vincent J., Zwaigenbaum L., Fernandez B., Roberts W., Szatmari P., Scherer S.W. Contribution of SHANK3 mutations to autism spectrum disorder. Am. J. Hum. Genet. 2007;81:1289–1297. doi: 10.1086/522590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jamain S., Quach H., Betancur C., Rastam M., Colineaux C., Gillberg I.C., Soderstrom H., Giros B., Leboyer M., Gillberg C., Bourgeron T. Mutations of the X-linked genes encoding neuroligins NLGN3 and NLGN4 are associated with autism. Nat. Genet. 2003;34:27–29. doi: 10.1038/ng1136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chubykin A.A., Liu X., Comoletti D., Tsigelny I., Taylor P., Sudhof T.C. Dissection of synapse induction by neuroligins: effect of a neuroligin mutation associated with autism. J. Biol. Chem. 2005;280:22365–22374. doi: 10.1074/jbc.M410723200. [DOI] [PubMed] [Google Scholar]
- 12.Feng J., Schroer R., Yan J., Song W., Yang C., Bockholt A., Cook E.H., Skinner C., Schwartz C.E., Sommer S.S. High frequency of neurexin 1beta signal peptide structural variants in patients with autism. Neurosci. Lett. 2006;409:10–13. doi: 10.1016/j.neulet.2006.08.017. [DOI] [PubMed] [Google Scholar]
- 13.Sebat J., Lakshmi B., Malhotra D., Troge J., Lese-Martin C., Walsh T., Yamrom B., Yoon S., Krasnitz A., Kendall J. Strong association of de novo copy number mutations with autism. Science. 2007;316:445–449. doi: 10.1126/science.1138659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhao X., Leotta A., Kustanovich V., Lajonchere C., Geschwind D.H., Law K., Law P., Qiu S., Lord C., Sebat J. A unified genetic theory for sporadic and inherited autism. Proc. Natl. Acad. Sci. USA. 2007;104:12831–12836. doi: 10.1073/pnas.0705803104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gillberg C. Chromosomal disorders and autism. J. Autism Dev. Disord. 1998;28:415–425. doi: 10.1023/a:1026004505764. [DOI] [PubMed] [Google Scholar]
- 16.Risi S., Lord C., Gotham K., Corsello C., Chrysler C., Szatmari P., Cook E.H., Leventhal B.L., Pickles A. Combining information from multiple sources in the diagnosis of autism spectrum disorders. J. Am. Acad. Child Adolesc. Psychiatry. 2006;45:1094–1103. doi: 10.1097/01.chi.0000227880.42780.0e. [DOI] [PubMed] [Google Scholar]
- 17.Falush D., Stephens M., Pritchard J.K. Inference of population structure using multilocus genotype data: linked loci and correlated allele frequencies. Genetics. 2003;164:1567–1587. doi: 10.1093/genetics/164.4.1567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Redon R., Ishikawa S., Fitch K.R., Feuk L., Perry G.H., Andrews T.D., Fiegler H., Shapero M.H., Carson A.R., Chen W. Global variation in copy number in the human genome. Nature. 2006;444:444–454. doi: 10.1038/nature05329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhao X., Li C., Paez J.G., Chin K., Janne P.A., Chen T.H., Girard L., Minna J., Christiani D., Leo C. An integrated view of copy number and allelic alterations in the cancer genome using single nucleotide polymorphism arrays. Cancer Res. 2004;64:3060–3071. doi: 10.1158/0008-5472.can-03-3308. [DOI] [PubMed] [Google Scholar]
- 20.Nannya Y., Sanada M., Nakazaki K., Hosoya N., Wang L., Hangaishi A., Kurokawa M., Chiba S., Bailey D.K., Kennedy G.C., Ogawa S. A robust algorithm for copy number detection using high-density oligonucleotide single nucleotide polymorphism genotyping arrays. Cancer Res. 2005;65:6071–6079. doi: 10.1158/0008-5472.CAN-05-0465. [DOI] [PubMed] [Google Scholar]
- 21.Komura D., Shen F., Ishikawa S., Fitch K.R., Chen W., Zhang J., Liu G., Ihara S., Nakamura H., Hurles M.E. Genome-wide detection of human copy number variations using high-density DNA oligonucleotide arrays. Genome Res. 2006;16:1575–1584. doi: 10.1101/gr.5629106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Krawczak M., Nikolaus S., von Eberstein H., Croucher P.J., El Mokhtari N.E., Schreiber S. PopGen: population-based recruitment of patients and controls for the analysis of complex genotype-phenotype relationships. Community Genet. 2006;9:55–61. doi: 10.1159/000090694. [DOI] [PubMed] [Google Scholar]
- 23.Iafrate A.J., Feuk L., Rivera M.N., Listewnik M.L., Donahoe P.K., Qi Y., Scherer S.W., Lee C. Detection of large-scale variation in the human genome. Nat. Genet. 2004;36:949–951. doi: 10.1038/ng1416. [DOI] [PubMed] [Google Scholar]
- 24.Zogopoulos G., Ha K.C., Naqib F., Moore S., Kim H., Montpetit A., Robidoux F., Laflamme P., Cotterchio M., Greenwood C. Germ-line DNA copy number variation frequencies in a large North American population. Hum. Genet. 2007;122:345–353. doi: 10.1007/s00439-007-0404-5. [DOI] [PubMed] [Google Scholar]
- 25.Scherer S.W., Cheung J., MacDonald J.R., Osborne L.R., Nakabayashi K., Herbrick J.A., Carson A.R., Parker-Katiraee L., Skaug J., Khaja R. Human chromosome 7: DNA sequence and biology. Science. 2003;300:767–772. doi: 10.1126/science.1083423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kohler J.R., Cutler D.J. Simultaneous discovery and testing of deletions for disease association in SNP genotyping studies. Am. J. Hum. Genet. 2007;81:684–699. doi: 10.1086/520823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pinto D., Marshall C., Feuk L., Scherer S.W. Copy-number variation in control population cohorts. Hum. Mol. Genet. 2007;16:168–173. doi: 10.1093/hmg/ddm241. [DOI] [PubMed] [Google Scholar]
- 28.Stranger B.E., Forrest M.S., Dunning M., Ingle C.E., Beazley C., Thorne N., Redon R., Bird C.P., de Grassi A., Lee C. Relative impact of nucleotide and copy number variation on gene expression phenotypes. Science. 2007;315:848–853. doi: 10.1126/science.1136678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lupski J.R. Genomic rearrangements and sporadic disease. Nat. Genet. 2007;39:S43–S47. doi: 10.1038/ng2084. [DOI] [PubMed] [Google Scholar]
- 30.Bonnet C., Gregoire M.J., Brochet K., Raffo E., Leheup B., Jonveaux P. Pure de-novo 5 Mb duplication at Xp11.22-p11.23 in a male: phenotypic and molecular characterization. J. Hum. Genet. 2006;51:815–821. doi: 10.1007/s10038-006-0023-3. [DOI] [PubMed] [Google Scholar]
- 31.Wong W., Newell E.W., Jugloff D.G., Jones O.T., Schlichter L.C. Cell surface targeting and clustering interactions between heterologously expressed PSD-95 and the Shal voltage-gated potassium channel, Kv4.2. J. Biol. Chem. 2002;277:20423–20430. doi: 10.1074/jbc.M109412200. [DOI] [PubMed] [Google Scholar]
- 32.Ensenauer R.E., Adeyinka A., Flynn H.C., Michels V.V., Lindor N.M., Dawson D.B., Thorland E.C., Lorentz C.P., Goldstein J.L., McDonald M.T. Microduplication 22q11.2, an emerging syndrome: clinical, cytogenetic, and molecular analysis of thirteen patients. Am. J. Hum. Genet. 2003;73:1027–1040. doi: 10.1086/378818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ghebranious N., Giampietro P.F., Wesbrook F.P., Rezkalla S.H. A novel microdeletion at 16p11.2 harbors candidate genes for aortic valve development, seizure disorder, and mild mental retardation. Am. J. Med. Genet. A. 2007;143:1462–1471. doi: 10.1002/ajmg.a.31837. [DOI] [PubMed] [Google Scholar]
- 34.Jacquemont M.L., Sanlaville D., Redon R., Raoul O., Cormier-Daire V., Lyonnet S., Amiel J., Le Merrer M., Heron D., de Blois M.C. Array-based comparative genomic hybridisation identifies high frequency of cryptic chromosomal rearrangements in patients with syndromic autism spectrum disorders. J. Med. Genet. 2006;43:843–849. doi: 10.1136/jmg.2006.043166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Feuk L., Carson A.R., Scherer S.W. Structural variation in the human genome. Nat. Rev. Genet. 2006;7:85–97. doi: 10.1038/nrg1767. [DOI] [PubMed] [Google Scholar]
- 36.Skuse D.H. Rethinking the nature of genetic vulnerability to autistic spectrum disorders. Trends Genet. 2007;23:387–395. doi: 10.1016/j.tig.2007.06.003. [DOI] [PubMed] [Google Scholar]
- 37.Zoghbi H.Y. Postnatal neurodevelopmental disorders: meeting at the synapse? Science. 2003;302:826–830. doi: 10.1126/science.1089071. [DOI] [PubMed] [Google Scholar]
- 38.Mitchell K.J. The genetics of brain wiring: from molecule to mind. PLoS Biol. 2007;5:e113. doi: 10.1371/journal.pbio.0050113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ballif B.C., Hornor S.A., Jenkins E., Madan-Khetarpal S., Surti U., Jackson K.E., Asamoah A., Brock P.L., Gowans G.C., Conway R.L. Discovery of a previously unrecognized microdeletion syndrome of 16p11.2-p12.2. Nat. Genet. 2007;39:1071–1073. doi: 10.1038/ng2107. [DOI] [PubMed] [Google Scholar]
- 40.Sharp A.J., Selzer R.R., Veltman J.A., Gimelli S., Gimelli G., Striano P., Coppola A., Regan R., Price S.M., Knoers N.V. Characterization of a recurrent 15q24 microdeletion syndrome. Hum. Mol. Genet. 2007;16:567–572. doi: 10.1093/hmg/ddm016. [DOI] [PubMed] [Google Scholar]
- 41.Somerville M.J., Mervis C.B., Young E.J., Seo E.J., del Campo M., Bamforth S., Peregrine E., Loo W., Lilley M., Perez-Jurado L.A. Severe expressive-language delay related to duplication of the Williams-Beuren locus. N. Engl. J. Med. 2005;353:1694–1701. doi: 10.1056/NEJMoa051962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yu C.E., Dawson G., Munson J., D'Souza I., Osterling J., Estes A., Leutenegger A.L., Flodman P., Smith M., Raskind W.H. Presence of large deletions in kindreds with autism. Am. J. Hum. Genet. 2002;71:100–115. doi: 10.1086/341291. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.