

Abstract
Intraportal serotonin infusion enhances net hepatic glucose uptake (NHGU) during glucose infusion but blunts nonhepatic glucose uptake and can cause gastrointestinal discomfort and diarrhea at high doses. Whether the serotonin precursor 5-hydroxytryptophan (5-HTP) could enhance NHGU without gastrointestinal side effects during glucose infusion was examined in conscious 42-h-fasted dogs, using arteriovenous difference and tracer ([3-3H]glucose) techniques. Experiments consisted of equilibration (−120 to −30 min), basal (−30 to 0 min), and experimental (EXP; 0–270 min) periods. During EXP, somatostatin, fourfold basal intraportal insulin, basal intraportal glucagon, and peripheral glucose (to double the hepatic glucose load) were infused. In one group of dogs (HTP, n = 6), saline was infused intraportally from 0 to 90 min (P1), and 5-HTP was infused intraportally at 10, 20, and 40 μg·kg−1·min−1 from 90 to 150 (P2), 150 to 210 (P3), and 210 to 270 (P4) min, respectively. In the other group (SAL, n = 7), saline was infused intraportally from 0 to 270 min. NHGU in SAL was 14.8 ± 1.9, 18.5 ± 2.3, 16.3 ± 1.4, and 19.7 ± 1.6 μmol·kg−1·min−1 in P1–P4, whereas NHGU in 5-HTP averaged 16.4 ± 2.6, 18.5 ± 1.4, 20.8 ± 2.0, and 27.6 ± 2.6 μmol·kg−1·min−1 (P < 0.05 vs. SAL). Nonhepatic glucose uptake (μmol·kg−1·min−1) in SAL was 30.2 ± 4.3, 36.8 ± 5.8, 44.3 ± 5.8, and 54.6 ± 11.8 during P1–P4, respectively, whereas in HTP the corresponding values were 26.3 ± 6.8, 44.9 ± 10.1, 47.5 ± 11.7, and 51.4 ± 13.2 (not significant between groups). Intraportal 5-HTP enhances NHGU without significantly altering nonhepatic glucose uptake or causing gastrointestinal side effects, raising the possibility that a related agent might have a role in reducing postprandial hyperglycemia.
Keywords: glycemia, liver, portal vein, leptin, adiponectin
Poor postprandial glycemic control is associated with elevated rates of all-cause mortality in individuals with type 2 diabetes (45), and thus control of postprandial hyperglycemia is an attractive pharmaceutical target. Intraportal infusion of serotonin (5-hydroxytryptamine or 5-HT) enhances net hepatic glucose uptake (NHGU) in the hyperinsulinemic, hyperglycemic conscious dog (28), indicating that 5-HT might be effective in reducing postprandial hyperglycemia. However, elevated circulating 5-HT is associated with intestinal cramping and diarrhea (3). Thus, for potential therapeutic use, agents that raise tissue levels of 5-HT without elevating blood 5-HT excessively are desirable.
Intraperitoneal administration of 5-hydroxytryptophan (5-HTP), the immediate precursor of 5-HT, has a hypoglycemic effect in mice that is apparently related to intrahepatic accumulation of 5-HT (8). We have demonstrated that intraportal infusion of the selective serotonin reuptake inhibitor (SSRI) fluvoxamine, which prolongs the action of endogenous 5-HT, stimulates NHGU (27). One multicenter study comparing fluvoxamine and 5-HTP in treatment of depression determined that 5-HTP was more effective (37), suggesting that serotonin deficiency (which can be corrected by administration of 5-HTP but not an SSRI) may exist in some individuals. Similarly, experimental models of diabetes are reported to have reduced levels of 5-HT (26). Thus we hypothesized that 5-HTP would enhance NHGU, thereby directing increased amounts of glucose in the liver as opposed to the peripheral circulation, without elevating circulating 5-HT sufficiently to cause adverse symptoms. We therefore examined the ability of 5-HTP given intraportally to conscious dogs in which the pancreatic hormones and hepatic glucose load were fixed at postprandial levels.
RESEARCH DESIGN AND METHODS
Animals and surgical procedures
Studies were carried out on 13 conscious 42-h-fasted mongrel dogs of either sex with a mean weight of 23.1 ± 0.6 kg. Diet was as previously described (36), the animals were housed in a facility approved by the United States Department of Agriculture, and the protocol was approved by the Vanderbilt University Medical Center Institutional Animal Care and Use Committee. The 42-h fast was used to reduce hepatic glycogen concentrations to a stable minimum, which reduces variability in the animals’ response to hyperglycemia and hyperinsulinemia. Dogs tolerate a fast of this duration well, with only a slight decline in blood glucose (~0.2–0.3 mM change) and little change in glucagon (~10%) between 18 and 42 h of fasting (30).
Approximately 16 days before study, each dog underwent a laparotomy for placement of ultrasonic flow probes (Transonic Systems, Ithaca, NY) around the portal vein and the hepatic artery, as well as for insertion of silicone rubber catheters for sampling in a hepatic vein, the portal vein, and a femoral artery and for infusion in a splenic and a jejunal vein as described elsewhere (36, 42). Criteria for study were as previously described (36).
On the morning of the study, catheters and flow probe leads were exteriorized from their subcutaneous pockets (36). The splenic and jejunal catheters were used for intraportal infusion of insulin (Eli Lilly, Indianapolis, IN), glucagon (GlucaGen; Bedford Laboratories, Bedford, OH), and 5-HTP (Sigma-Aldrich, St. Louis, MO). Angiocaths (Deseret Medical, Sandy, UT) were inserted in three peripheral veins.
Experimental design
Each experiment consisted of a 90-min equilibration period (−120 to −30 min), a 30-min basal period (−30 to 0 min), and a 270-min experimental period (0–270 min) divided into four subperiods (P1, 0–90 min; P2, 90–150 min; P3, 150–210 min, and P4, 210–270 min). At −120 min, a primed (38 μCi), continuous (0.35 μCi/min) infusion of [3-3H]glucose (NEN, Cambridge, MA) and a continuous infusion of indocyanine green dye (Sigma; 5 μg·kg−1·min−1) were begun in all dogs, with the exception of one that did not receive [3-3H]glucose. At 0 min, a constant peripheral infusion of somatostatin (0.8 μg·kg−1·min−1; Bachem, Torrance, CA) was begun to suppress endogenous insulin and glucagon secretion. Insulin was infused intraportally at 1.2 mU·kg−1·min−1 (4-fold basal), and glucagon (0.55 ng·kg−1·min−1) was replaced intraportally in basal amounts. In addition, a primed, continuous variable-rate infusion of 50% dextrose was begun through a peripheral vein to maintain the hepatic glucose load twofold basal. During P1, all dogs received intraportal normal saline infusion. At the end of P1, the dogs were divided into two groups. In the saline (SAL) group (n = 7), the intraportal saline infusion continued for the remainder of the study. In the HTP group (n = 6), 5-HTP was infused in the portal vein at 10, 20, and 40 μg·kg−1·min−1 during P2, P3, and P4, respectively. The infusion rates were the same as the infusion rates for 5-HT in our previous study (28). The portal route of infusion was chosen to target the 5-HTP at the liver as much as possible. In addition, we did not administer an inhibitor of peripheral aromatic L-amino acid decarboxylase (AADC). An AADC inhibitor would have reduced the conversion of 5-HTP to 5-HT in peripheral tissues, making more 5-HTP available to cross the blood-brain barrier (10). The liver has relatively high levels of AADC (21, 44) and thus should synthesize substantial amounts of 5-HT from the substrate.
Femoral artery, portal vein, and hepatic vein blood samples were taken every 15–30 min throughout the study, as previously described (36). Arterial blood samples were also taken every 5 min throughout the experimental period to monitor the glucose level (36). After completion of each experiment, the animal was killed with an overdose of pentobarbital sodium.
Processing and analysis of samples
Hematocrit, blood glucose, lactate, glycerol, plasma glucose, nonesterified fatty acids (NEFA), insulin, glucagon, cortisol, catecholamines, and [3H]glucose were measured as described previously (36, 42). 5-HT concentrations were determined on whole blood by an HPLC-amperometric assay (38) with a coefficient of variation (CV) of 4%, as previously reported (28). Plasma leptin concentrations were analyzed by a sandwich ELISA assay using an anti-canine leptin antibody as previously described (20) with inter- and intra-assay CV <4%. Plasma adiponectin was measured using a commercial mouse/rat ELISA kit (Otsuka Pharmaceuticals, Tokushima, Japan) optimized for analysis of canine samples, with intra-assay CV of 6.7–8.5% and interassay CV of 3.4–7.5% (35).
Calculations and data analysis
Hepatic blood flow was measured using ultrasonic flow probes and by use of indocyanine green extraction. The two methods yielded similar results, but the data reported here were calculated with the ultrasonic-determined flows because their measurement did not require an assumption regarding the relative contribution of arterial and portal flow to total hepatic blood flow.
The rate of glucose delivery to the liver (or hepatic glucose load), net hepatic substrate balance, net hepatic fractional substrate extraction, net hepatic carbon retention, hepatic sinusoidal insulin and glucagon concentrations, and nonhepatic glucose uptake (non-HGU) were calculated as described previously (33). During the 1st h of glucose infusion, the non-HGU was corrected for the glucose required to fill the pool, using a pool fraction of 0.65 (6) and assuming that the volume of distribution for glucose equaled the volume of the extracellular fluid, or ~22% of the dog’s weight (43). For all glucose balance calculations, glucose concentrations were converted from plasma to blood values by using correction factors (ratio of the blood to the plasma concentration), as previously established in our laboratory (18). Hepatic glucose uptake (in dpm·kg−1·min−1) was calculated as was NHGU, except that tritiated glucose measurements were used. The results were divided by the inflowing glucose specific activity (dpm/μmol glucose). Glucose rates of appearance (Ra) and disappearance (Rd) were calculated with a two compartment model using dog parameters previously described (7, 25).
Statistical analysis
All data are presented as means ± SE. Time course data were analyzed with repeated-measures ANOVA, with Tukey’s test for post hoc comparisons (SigmaStat; Jandel Scientific). Statistical significance was accepted at P < 0.05. Results for P1–P4 are the means of the three samples taken during the last 30 min of each period, when steady-state conditions prevailed.
RESULTS
5-HT, hormone, and catecholamine concentrations
Arterial blood 5-HT concentrations remained basal throughout the experiments in SAL and during P1–P3 in HTP, but the concentration increased significantly during P4 (Table 1). The arterial 5-HT concentration in HTP during P4 was similar to those evident in four dogs during the first 2 h after an oral glucose tolerance test consisting of 1.5 mg glucose/kg (M. C. Moore and A. D. Cherrington, unpublished observations). Net hepatic 5-HT balance remained basal in SAL throughout the experiments. In the HTP group, net hepatic 5-HT balance did not change from basal during P1–P3, but net hepatic 5-HT uptake increased significantly during P4.
Table 1.
Serotonin and hormone data
| Parameter and Group | Basal Period | Experimental Period
|
|||
|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | ||
| Arterial blood 5-HT, mg/l | |||||
| SAL | 0.9±0.2 | 0.8±0.2 | 0.8±0.2 | 0.8±0.2 | 0.7±0.2 |
| HTP | 1.0±0.2 | 1.0±0.2 | 1.1±0.2 | 1.3±0.3 | 2.0±0.6* |
| Net hepatic 5-HT balance, μg·kg−1·min−1 | |||||
| SAL | 2.2±0.5 | 1.7±1.0 | 0.4±1.5 | −0.1±2.2 | 1.2±1.4 |
| HTP | −2.8±2.9 | −3.3±1.9 | −4.0±1.8 | −2.8±3.1 | −7.4±3.4* |
| Arterial plasma insulin, pmol/l | |||||
| SAL | 46±8 | 108±18 | 110±11 | 129±13 | 135±17 |
| HTP | 27±3 | 120±9 | 123±11 | 143±11 | 155±6 |
| Hepatic sinusoidal insulin, pmol/l | |||||
| SAL | 128±21 | 417±67 | 425±45 | 399±47 | 414±37 |
| HTP | 181±15 | 411±34 | 359±39 | 383±44 | 386±40 |
| Arterial plasma glucagon, ng/l | |||||
| SAL | 46±7 | 41±5 | 38±5 | 38±4 | 35±8 |
| HTP | 33±4 | 39±4 | 28±3 | 32±3 | 34±2 |
| Hepatic sinusoidal glucagon, ng/l | |||||
| SAL | 54±8 | 58±8 | 56±5 | 52±5 | 48±4 |
| HTP | 45±7 | 44±4 | 42±4 | 47±4 | 42±5 |
| Arterial plasma cortisol, nmol/l | |||||
| SAL | 74±24 | 64±18 | 55±14 | 43±10 | 62±10 |
| HTP | 82±21 | 66±12 | 51±12 | 72±33 | 116±43 |
| Arterial plasma norepinephrine, pg/ml | |||||
| SAL | 185±39 | 154±33 | 152±52 | 177±39 | 173±44 |
| HTP | 114±38 | 139±59 | 115±57 | 140±65 | 142±49 |
| Arterial plasma epinephrine, pg/ml | |||||
| SAL | 149±55 | 78±33 | 55±14 | 74±33 | 75±39 |
| HTP | 102±43 | 93±39 | 79±43 | 112±55 | 61±38 |
Values are means ± SE. 5-HT, 5-hydroxytryptamine. SAL dogs (n = 7) received intraportal saline infusion during periods 1–4 (P1–P4); HTP dogs (n = 6) received saline during P1 and intraportal 5-hydroxytryptophan (5-HTP) at 10, 20, and 40 μg·kg−1·min−1, respectively, during P2–P4. Negative values indicate net hepatic uptake.
P < 0.05 vs. SAL.
The plasma insulin levels increased approximately three- to fourfold and remained stable during P1–P4 in both groups (Table 1). Arterial and hepatic sinusoidal plasma glucagon concentrations were basal and indistinguishable in both groups throughout the experiments (Table 1).
Arterial plasma cortisol concentrations remained basal in SAL throughout P1–P4. In HTP, the mean cortisol concentrations remained statistically unchanged from basal during P1–P4, but one animal did demonstrate a substantial increase from basal (84–271 nmol/l) during P4. This dog also exhibited a rise in arterial 5-HT concentrations to nearly threefold basal compared with the 40% increase observed in the other five dogs in the HTP group. Arterial plasma concentrations of epinephrine and norepinephrine did not change significantly during P1–P4 in either group (Table 1).
Plasma leptin and adiponectin concentrations were available on four animals in each group. The concentrations of both hormones were similar during the basal period in the two groups, and they did not change significantly during the study in either group (Table 2).
Table 2.
Plasma leptin and adiponectin concentrations
| Leptin, ng/ml
|
Adiponectin, μg/ml
|
|||||
|---|---|---|---|---|---|---|
| Blood Vessel and Group | Basal period | Experimental period
|
Basal period | Experimental period
|
||
| P3 | P4 | P3 | P4 | |||
| Artery | ||||||
| SAL | 13.1±0.9 | 13.6±1.3 | 13.9±1.0 | 15.2±2.2 | 12.8±1.9 | 13.5±2.7 |
| HTP | 12.1±1.3 | 13.5±1.8 | 13.9±1.7 | 13.3±2.5 | 12.6±1.6 | 12.6±3.4 |
| Portal vein | ||||||
| SAL | 12.8±0.6 | 14.3±1.3 | 14.5±1.2 | 19.2±8.0 | 16.2±4.0 | 16.0±2.8 |
| HTP | 11.9±1.5 | 13.6±1.6 | 13.7±2.0 | 16.9±3.4 | 14.2±2.9 | 12.2±1.3 |
| Hepatic vein | ||||||
| SAL | 12.8±0.5 | 14.2±1.2 | 14.3±1.2 | 18.4±2.9 | 20.7±5.4 | 20.1±5.8 |
| HTP | 11.8±1.3 | 13.0±1.5 | 13.4±1.6 | 17.3±2.6 | 14.6±2.5 | 12.9±1.6 |
Values are means ± SE. Samples were available from only 4 dogs/group and were collected only during the basal period, P3, and P4. SAL dogs received intraportal saline infusion during P1–P4; HTP dogs received saline during P1 and intraportal 5-HTP at 20 and 40 μg·kg−1·min−1, respectively, during P3 and P4. There were no significant differences between groups.
Hepatic blood flow, blood glucose concentrations, and hepatic glucose load
Portal vein blood flow decreased modestly in both groups during P1 as a response to somatostatin infusion (Table 3) and did not change significantly thereafter. There was a concomitant increase in hepatic artery flow. As a consequence, total hepatic blood flow was relatively stable throughout the experiments, with no differences between groups at any time.
Table 3.
Hepatic blood flows
| Group and Blood Vessel | Basal Period | Experimental Period
|
|||
|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | ||
| Hepatic artery | |||||
| SAL | 6.1±0.6 | 7.7±0.6 | 8.6±0.8 | 9.2±1.1 | 9.7±1.4 |
| HTP | 5.7±0.9 | 6.0±1.2 | 7.3±2.1 | 7.8±2.0 | 7.9±1.9 |
| Portal vein | |||||
| SAL | 24.2±2.1 | 20.3±2.1 | 21.0±2.1 | 20.6±2.2 | 20.4±2.1 |
| HTP | 26.6±2.4 | 21.2±1.2 | 22.9±1.6 | 21.9±1.5 | 21.2±1.3 |
Values are means ± SE. (Units are ml·kg−1·min−1). SAL dogs (n = 7) received intraportal saline infusion during P1–P4; HTP dogs (n = 6) received saline during P1 and intraportal 5-HTP at 10, 20, and 40 μg·kg−1·min−1, respectively, during P2–P4. There were no significant differences between groups.
Arterial blood glucose levels in SAL increased from a basal value of 4.6 ± 0.1 to 9.3 ± 0.1 mmol/l during all experimental periods (Fig. 1). In HTP, the arterial glucose concentration increased from 4.3 ± 0.2 to 9.0 ± 0.1 mmol/l. There were no significant differences between groups. In addition, the hepatic glucose loads were not significantly different between groups at any time (Fig. 1), increasing from 145 ± 9 (basal) to 271 ± 20 μmol·kg−1·min−1 during P1–P4 in SAL and from 137 ± 16 to 257 ± 18 μmol·kg−1·min−1 in HTP.
Fig. 1.
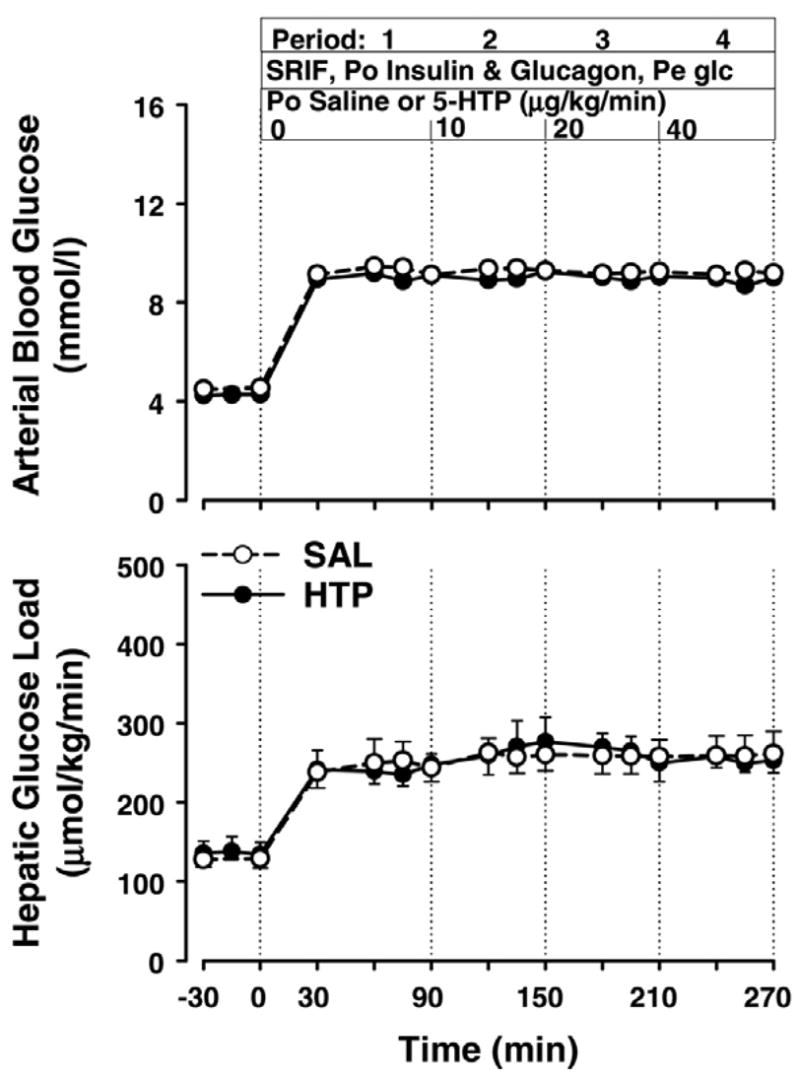
Arterial blood glucose and hepatic glucose load in dogs receiving somatostatin (SRIF), intraportal (Po) infusions of insulin (4-fold basal) and glucagon (basal), peripheral (Pe) glucose (Glc) infusion to double the hepatic glucose load, and intraportal infusion of 5-hydroxytryptophan [5-HTP (HTP) group; n = 6 dogs] at the rates shown or saline (SAL; n = 7). There were no significant differences between groups.
Net hepatic glucose balance, net hepatic fractional glucose extraction, and endogenous glucose Ra
The groups exhibited a similar rate of net hepatic glucose output during the basal period. Coincident with the start of the experimental period, they switched from net production to net uptake, with the rates being no different between groups during P1 and P2 (Fig. 2). In both groups, NHGU during P2 averaged 18.5 μmol·kg−1·min−1. Subsequently, the rate of NHGU remained relatively stable in SAL (mean rate 19.7 ± 1.6 μmol·kg−1·min−1 in P4), whereas in HTP it increased to 27.6 ± 2.6 μmol·kg−1·min−1 (P < 0.05 vs. SAL). Similarly, the net hepatic fractional extraction (FE) of glucose did not differ significantly between groups during P1 and P2, averaging 0.065 ± 0.007 and 0.074 ± 0.013 during P2 in SAL and HTP, respectively. Subsequently, FE changed little in SAL (0.060 ± 0.007 and 0.073 ± 0.006 during P3 and P4, respectively), but it increased in HTP (0.084 ± 0.014 and 0.110 ± 0.014 during P3 and P4, respectively; P < 0.05 vs. SAL during P4). The rate of unidirectional (tracer-determined) hepatic glucose uptake did not differ significantly between groups during any period, although there was a tendency (P = 0.07) for it to be greater in HTP vs. SAL during P4 (Table 4). However, compared with SAL, HTP demonstrated significant enhancement of hepatic glucose uptake between P1 and each of the last two periods (Δ between P1 and P3: 2.2 ± 2.1 vs. 11.3 ± 2.7 μmol·kg−1·min−1 in SAL vs. HTP, respectively, P < 0.05; Δ between P1 and P4: 5.3 ± 1.4 vs. 17.6 ± 3.0 μmol·kg−1·min−1, P < 0.005; Table 4).
Fig. 2.
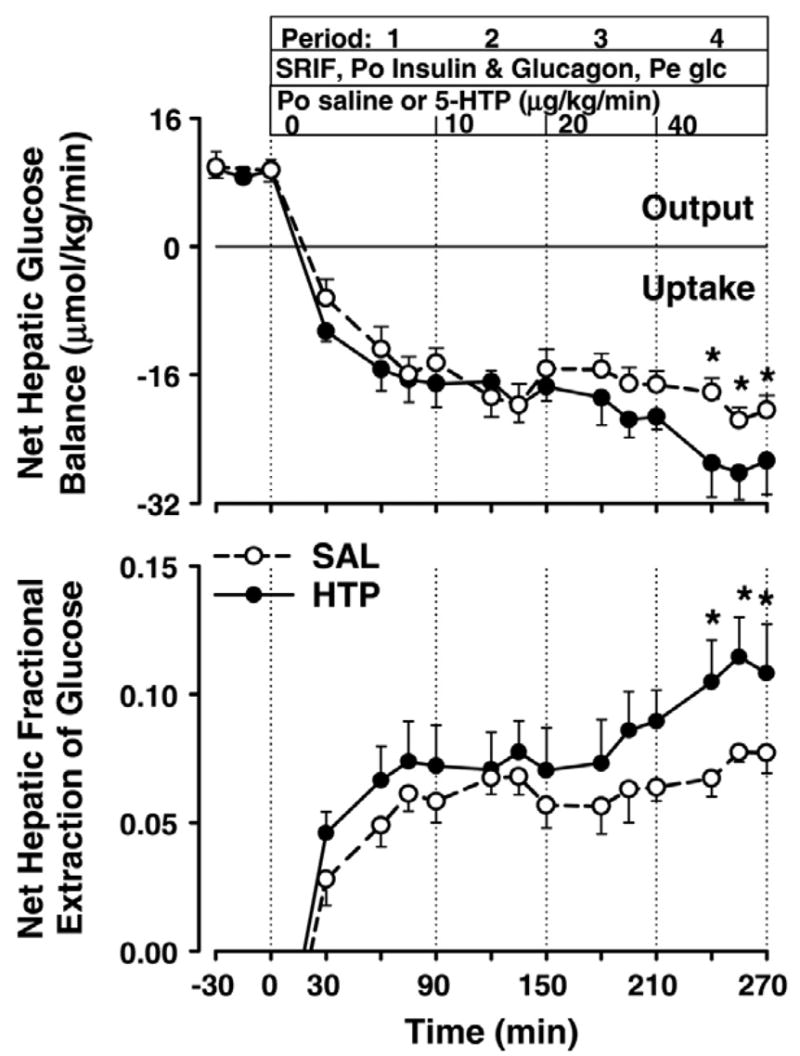
Net hepatic glucose uptake and fractional extraction of glucose. See legend to Fig. 1 for description of study conditions. *P < 0.05 between groups.
Table 4.
Tracer-determined unidirectional HGU and glucose Rd
| Group and parameter | Basal Period | Experimental Period
|
|||
|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | ||
| HGU | |||||
| SAL | 0.7±0.6 | 14.3±3.1 | 17.0±4.0 | 16.5±3.9 | 19.6±3.0 |
| HTP | 0.1±0.9 | 10.2±3.3 | 13.2±1.5 | 21.5±2.2 | 27.9±5.3 |
| Glucose Rd | |||||
| SAL | 13.6±0.7 | 51.3±7.7 | 59.8±8.0 | 66.7±8.7 | 80.8±12.5 |
| HTP | 12.9±0.6 | 53.4±12.4 | 65.3±14.5 | 71.2±13.4 | 84.2±11.8 |
Values are means ± SE; n = 7 for SAL group, and n = 5 for HTP group. Units are μmol·kg−1·min−1. HGU, hepatic glucose uptake; Rd, rate of disappearance. SAL dogs received intraportal saline infusion during P1–P4; HTP dogs received saline during P1 and intraportal 5-HTP at 10, 20, and 40 μg·kg−1·min−1, respectively, during P2–P4. P = 0.07 between groups for HGU during P4.
Endogenous glucose Ra decreased similarly in response to hyperglycemia and hyperinsulinemia in both groups (from 14.8 ± 1.0 to 3.4 ± 1.3 μmol·kg−1·min−1 in SAL and from 15.6 ± 3.5 to 3.6 ± 2.4 μmol·kg−1·min−1 in HTP), remained suppressed throughout the experimental periods and did not differ significantly between groups at any time (data not shown).
Total and nonhepatic glucose disposal
The glucose infusion rate in both groups increased steadily in a time-dependent manner during P1–P4, with no differences between groups (45.0 ± 5.3, 55.2 ± 7.4, 60.6 ± 6.9, and 73.7 ± 11.6 μmol·kg−1·min−1 in SAL and 42.7 ± 7.9, 63.4 ± 10.4, 68.3 ± 10.4, and 79.0 ± 11.4 μmol·kg−1·min−1 in HTP; Fig. 3). Similarly, both glucose Rd (Table 4) and non-HGU (Fig. 3) increased with time and did not differ between groups during any period. Non-HGU averaged 30.2 ± 4.3, 36.8 ± 5.8, 44.3 ± 5.8, and 54.6 ± 11.8 μmol·kg−1·min−1 during P1–P4 in SAL and 26.3 ± 6.8, 44.9 ± 10.8, 47.4 ± 11.8, and 51.4 ± 13.2 μmol·kg−1·min−1 in HTP.
Fig. 3.
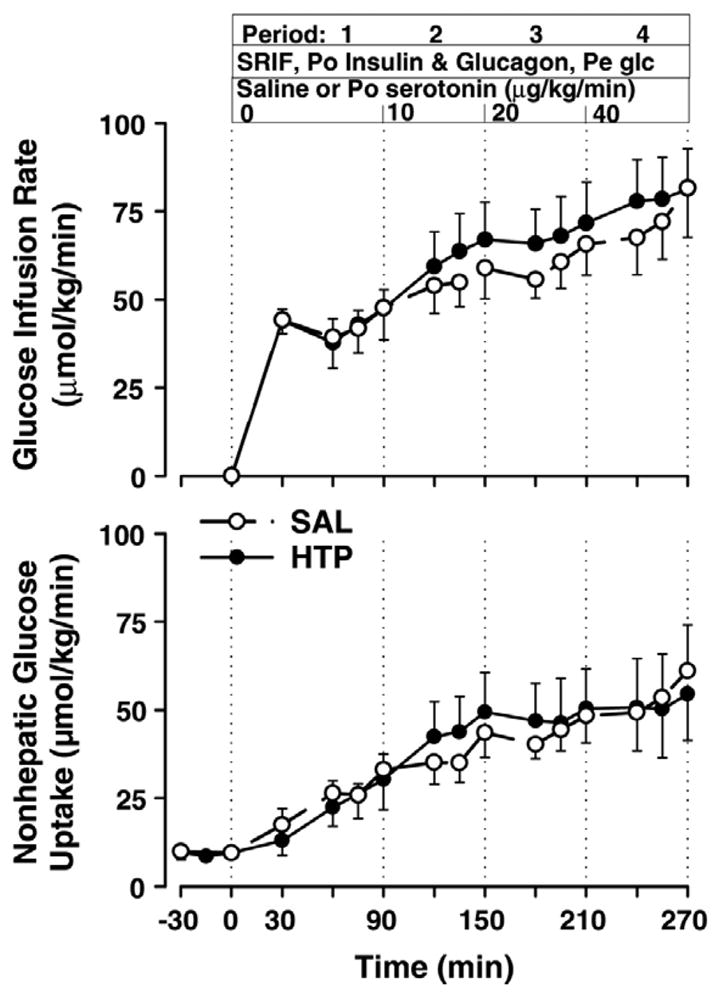
Glucose infusion rate and nonhepatic glucose uptake. See legend to Fig. 1 for description of study conditions. There were no significant differences between groups.
Lactate metabolism and net hepatic carbon retention
Basal arterial blood lactate concentrations were significantly higher in SAL than HTP, but both groups demonstrated a significant increase in lactate levels in response to hyperglycemia and hyperinsulinemia (Table 5). The lactate concentrations continued to be lower in HTP than SAL throughout the study, but this did not reach statistical significance during P1 and P4. Both groups demonstrated a switch from net hepatic lactate uptake to output at the onset of the experimental period, and the rates in the two groups did not differ significantly at any time.
Table 5.
Arterial lactate, glycerol, and NEFA concentrations and net hepatic balances
| Group | Basal Period | Experimental Period
|
|||
|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | ||
| Arterial blood lactate, μmol/l | |||||
| SAL | 536±74 | 1,060±49 | 975±72 | 981±75 | 1,028±91 |
| HTP | 298±52* | 802±148 | 665±92* | 685±90* | 816±135 |
| Net hepatic lactate balance, μmol·kg−1·min−1 | |||||
| SAL | −9.6±2.2 | 6.1±3.1 | 2.5±2.3 | 1.4±2.5 | 2.8±2.3 |
| HTP | −6.1±0.8 | 6.6±1.8 | 4.6±0.9 | 2.0±0.9 | 2.2±0.5 |
| Arterial blood glycerol, μmol/l | |||||
| SAL | 86±16 | 29±3 | 28±5 | 28±6 | 30±7 |
| HTP | 64±7 | 25±4 | 19±4 | 22±5 | 26±6 |
| Net hepatic glycerol uptake, μmol·kg−1·min−1 | |||||
| SAL | 1.7±0.4 | 0.4±0.0 | 0.4±0.1 | 0.4±0.1 | 0.4±0.1 |
| HTP | 1.5±0.2 | 0.4±0.1 | 0.2±0.1 | 0.3±0.2 | 0.5±0.2 |
| Arterial plasma NEFA, μmol/l | |||||
| SAL | 789±103 | 132±23 | 96±20 | 86±27 | 83±28 |
| HTP | 938±66 | 122±29 | 71±12 | 60±16 | 68±19 |
| Net hepatic NEFA uptake, μmol·kg−1·min−1 | |||||
| SAL | 2.4±0.6 | 0.2±0.1 | 0.1±0.1 | 0.2±0.1 | 0.2±0.1 |
| HTP | 3.3±0.2 | 0.6±0.3 | 0.0±0.0 | 0.3±0.2 | 0.0±0.1 |
Values are means ± SE. NEFA, nonesterified fatty acid. SAL dogs (n = 7) received intraportal saline infusion during P1–P4; HTP dogs (n = 6) received saline during P1 and intraportal 5-HTP at 10, 20, and 40 μg·kg−1·min−1, respectively, during P2–P4. Negative values for balance indicate net hepatic lactate uptake; rates of net hepatic uptake are shown as positive values for substrates for which there was no net release by the liver.
P < 0.05 vs. SAL.
The rates of net hepatic carbon retention in SAL and HTP were similar during P1 and P2 (11.6 ± 1.9 and 17.0 ± 2.7 μmol·kg−1·min−1 in SAL and 13.1 ± 2.6 and 16.7 ± 0.9 μmol·kg−1·min−1 in HTP; Fig. 4). However, during P3 and P4, the rates in HTP (20.1 ± 2.4 and 26.2 ± 2.3 μmol·kg−1·min−1) were greater (P < 0.05) than those in SAL (15.4 ± 2.0 and 17.5 ± 1.8 μmol·kg−1·min−1).
Fig. 4.
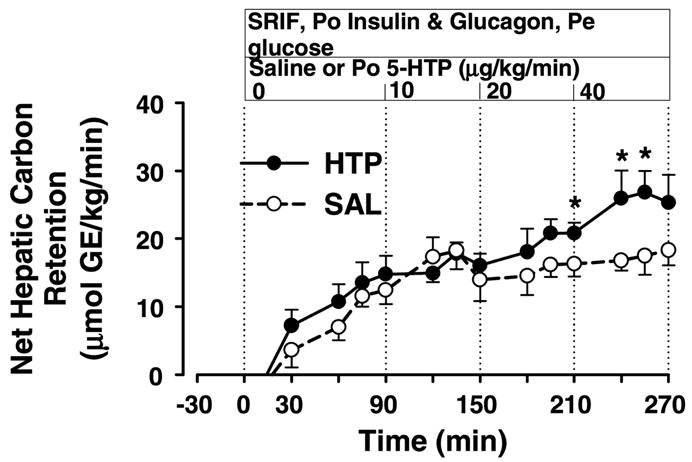
Net hepatic carbon retention. See legend to Fig. 1 for description of study conditions. GE, glucose equivalents. *P < 0.05 between groups.
Glycerol and NEFA metabolism
Arterial glycerol concentrations were reduced ~55–65% by hyperglycemia and hyper-insulinemia in both groups, and net hepatic glycerol uptake fell in parallel with the glycerol levels, remaining suppressed throughout the experimental period (Table 5). Arterial NEFA concentrations and net hepatic NEFA uptake changed in a manner similar to glycerol, decreasing ~80–90% during P1 in both groups and then remaining low throughout the remainder of the experimental period.
DISCUSSION
A twofold increase in the hepatic glucose load in the presence of fourfold basal insulin and basal glucagon concentrations resulted in NHGU of ~15.5 μmol·kg−1·min−1 in the two groups. Between P1 and P4, the rate of NHGU increased ~33% in SAL. During infusion of 5-HTP at 10 μg·kg−1·min−1, there was no enhancement in NHGU compared with SAL. However, during 5-HTP infusion at 20 and 40 μg·kg−1·min−1, NHGU increased 29 and 40%, respectively, over the corresponding rates in SAL (P < 0.05 for both increments). By the end of the experimental period, NHGU in HTP had increased 68% over the rate during P1 (P < 0.05 vs. the increase in SAL). Tracer measurements confirmed that there was a significant enhancement of hepatic glucose uptake over the rate during P1 associated with 5-HTP delivery, and there was a strong tendency for hepatic glucose uptake to differ between groups during P4. Similar to NHGU, net hepatic carbon retention was virtually identical between the groups during P1 and P2. During P3 and P4, net hepatic carbon retention was 31 and 50% greater, respectively, in HTP than in SAL (P < 0.05 for both). In this initial study, we merely wanted to determine whether 5-HTP, given at any rate that would bring about 5-HT concentrations in the physiological range, would induce NHGU. Therefore, we administered 5-HTP in three increasing steps during P2–P4. As a result, it is not possible to separate the effects of time from those of dosage. Nevertheless, it is possible to conclude definitively that 5-HTP impacted NHGU, hepatic glucose uptake, and hepatic carbon retention.
The effects of 5-HTP on the liver could have resulted either from a direct effect of 5-HT or from a secondary signal initiated elsewhere. In regard to a potential direct effect of 5-HT on hepatocytes, the liver contains AADC enzyme for conversion of 5-HTP to 5-HT, and this is enzyme is reported to have high efficiency at physiological substrate concentrations (13). Moreover, the 5-HT2B receptor is known to be expressed in greatest abundance in the liver and kidney in humans (4), and isoforms of the 5-HT3 receptor are also expressed in the human liver (23, 31). The actions of 5-HT within the liver have not been well studied. However, in the kidney and intestinal tract (nonneural tissues where 5-HT is synthesized and its actions have been more fully explored), 5-HT acts via autocrine/paracrine mechanisms (13, 24). Thus the potential exists for a direct hepatic action of 5-HT or an intrahepatic reflex elicited by 5-HT.
In regard to the possibility of indirect effects of 5-HT on the liver, 5-HT is commonly involved in afferent vagal signaling (e.g., see Ref. 16). The paracrine effects of 5-HT in the intestine involve stimulation of vagal afferents, with a reflex stimulation of pancreatic secretion (24). Niijima (32) determined that intraportal injection of 5-HT resulted in a decrease in the afferent firing rate in the hepatic branch of the vagus nerve and a stimulation of efferent firing in the pancreatic branch of the vagus, similar to the effect of intraportal glucose injection, suggesting a mechanism by which 5-HT could elicit a neural signal enhancing NHGU. Conversion of 5-HTP to 5-HT occurs in serum and many tissues, including the intestines, kidney, adrenals, and certain blood vessel walls (34, 39, 40, 50). Although we have not compared NHGU during peripheral venous vs. intraportal infusion of 5-HT, 5-HT elicited a greater change in afferent firing in the common hepatic branch of the vagus (which innervates the liver but also portions of the gastrointestinal tract), or a shorter period of latency, when injected in the jugular rather than the hepatic portal vein of rats, suggesting that 5-HT was acting primarily on nerve fibers outside the hepatoportal region (17). Additionally, rather than acting via a peripheral afferent neural signal, 5-HTP could have reached the central nervous system. 5-HTP crosses the blood-brain barrier, and the brain is rich in AADC (39). An increase in the brain 5-HT level could have resulted in a centrally mediated neural signal to the liver.
Another possible mechanism for the action of 5-HTP is via stimulation of the release of other hormones such as leptin (46, 47), which impacts upon insulin signaling and sensitivity in the liver (19, 22), or adiponectin (48), a hormone with insulin-sensitizing actions (49). Therefore, we measured these hormones in a subset of animals from each group. We were unable to identify any consistent changes related to 5-HTP administration, although we cannot rule out the possibility that 5-HTP increases the binding of leptin to its soluble receptor, thereby modulating its actions (5).
Because 5-HTP infusion was associated with a similar rate of non-HGU between groups but an enhancement of liver glucose uptake during P3 and P4, there was a tendency for whole body glucose disposal to be increased in the HTP group during those two periods. This tendency was evidenced in both the glucose infusion rate (~6–8 μmol·kg−1·min−1 greater in HTP than SAL during P3 and P4) and the glucose Rd (~4 μmol·kg−1·min−1 greater in HTP than SAL during P3 and P4), but neither parameter was statistically different between the groups (P = 0.4 for both). We have previously observed under a variety of conditions (e.g., Refs. 1, 9, 27, and 29) that, when NHGU is enhanced, there is a reciprocal reduction in nonhepatic (primarily skeletal muscle) glucose uptake such that there is little impact on whole body glucose disposal. Although the lack of reciprocity in the current study may simply have resulted from difficulty in observing a small change against the background of a high rate of non-HGU, it deserves further examination in the future. Impaired splanchnic (primarily liver) glucose uptake in response to glucose feeding is one of the defects associated with type 2 diabetes (2). We studied 5-HTP with the goal of determining whether it would be effective in increasing the proportion of a carbohydrate load extracted by the liver, thereby reducing the load to be disposed of in the peripheral tissues. Thus we did not expect 5-HTP to alter total body uptake. Nevertheless, the current data suggest that an optimal dosage or form of 5-HTP, or a related agent, might increase total glucose disposal.
This reciprocity of tissue glucose uptake (liver vs. muscle) was evident during 5-HT infusion at 20 and 40 μg·kg−1·min−1 (28), suggesting that 5-HT itself does not block the effect. However, differences in the circulating 5-HT concentrations are a possible explanation for differences in non-HGU with 5-HT and 5-HTP infusion. Circulating 5-HT rose substantially more during 5-HT infusion (28) than during 5-HTP infusion (~4-fold basal at the highest 5-HT infusion rate in contrast to the maximum 2-fold basal rise with 5-HTP). A direct effect of 5-HT to reduce peripheral glucose uptake is unlikely because 5-HT has either no effect (41) or a stimulatory effect (14) on glucose uptake in isolated skeletal muscle. The reduction of non-HGU with the higher concentrations of 5-HT infusion could have resulted from a change in muscle perfusion (41). In addition, at the higher rates of 5-HT infusion, we observed significant increases in cortisol and catecholamines (28), and this likely brought about a relative peripheral insulin resistance. The association of elevated 5-HT with increased levels of catecholamines and cortisol has been described in the clinical literature (15). No such elevation of stress hormones was observed in the HTP group, probably because of the more modest rise in circulating 5-HT. In the one dog with a threefold basal increase in 5-HT during 5-HTP infusion, there was a significant increase in cortisol and catecholamines during P4. However, that dog’s NHGU and non-HGU during P4 did not differ from the others in the group (NHGU 18.8 vs. a group mean of 27.6 ± 2.6 μmol·kg−1·min−1; non-HGU 60.8 vs. the group mean of 51.4 ± 13.2 μmol·kg−1·min−1). This agrees with previous data demonstrating that chronic elevation of cortisol brings about peripheral insulin resistance and markedly enhances NHGU (12), but acute elevations of cortisol have no such effect (11).
In conclusion, intraportal infusion of 5-HTP enhanced NHGU without blunting nonhepatic glucose disposal under hyperglycemic hyperinsulinemic conditions. We did not examine the effects of peripheral 5-HTP infusion, and thus it is not possible to speculate whether the effects observed were specific to the intraportal route of delivery. There were no significant increases in cortisol or catecholamines during 5-HTP delivery, and the dogs evidenced no diarrhea or signs of distress. Thus 5-HTP or related agents may provide a tool for reducing the proportion of the glucose load that must be disposed of in the peripheral tissues postprandially in individuals with type 2 diabetes or impaired glucose tolerance.
Acknowledgments
We thank Jon Hastings and the members of the Vanderbilt Diabetes Research and Training Center Hormone Core Laboratory (Wanda Snead, Angelina Penaloza, and Eric Allen) for technical support.
GRANTS
This work was supported by a Research Award from the American Diabetes Association to M. C. Moore and by National Institutes of Health (NIH) Grants R01 DK-58134 and SP-60-AM-20593. The serotonin analyses were performed by the Center for Molecular Neuroscience Neurochemistry Core, supported in part by NIH Grant P30 HD-15022.
References
- 1.Adkins BA, Myers SR, Hendrick GK, Stevenson RW, Williams PE, Cherrington AD. Importance of the route of intravenous glucose delivery to hepatic glucose balance in the conscious dog. J Clin Invest. 1987;79:557–565. doi: 10.1172/JCI112847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Basu A, Basu R, Shah P, Vella A, Johnson CM, Jensen M, Nair KS, Schwenk WF, Rizza RA. Type 2 diabetes impairs splanchnic uptake of glucose but does not alter intestinal glucose absorption during enteral glucose feeding: additional evidence for a defect in hepatic glucokinase activity. Diabetes. 2001;50:1351–1362. doi: 10.2337/diabetes.50.6.1351. [DOI] [PubMed] [Google Scholar]
- 3.Bearcroft CP, Perrett D, Farthing MJ. Postprandial plasma 5-hydroxytryptamine in diarrhoea predominant irritable bowel syndrome: a pilot study. Gut. 1998;42:42–46. doi: 10.1136/gut.42.1.42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bonhaus DW, Bach C, DeSouza A, Salazar FHR, Matsuoka BD, Zuppan P, Chan HW, Eglen RM. The pharmacology and distribution of human 5-hydroxytryptamine2B (5-HT2B) receptor gene products: comparison with 5-HT2A and 5-HT2C receptors. Br J Pharmacol. 1995;115:662–628. doi: 10.1111/j.1476-5381.1995.tb14977.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Brabant G, Nave H, Horn R, Anderwald C, Müller G, Roden M. In vivo and in vitro evidence for a hepatic modulation of the leptin signal in rats. Eur J Clin Invest. 2004;34:831–837. doi: 10.1111/j.1365-2362.2004.01427.x. [DOI] [PubMed] [Google Scholar]
- 6.Cowan JS, Hetenyi G., Jr Glucoregulatory responses in normal and diabetic dogs recorded by a new tracer method. Metabolism. 1971;20:360–372. doi: 10.1016/0026-0495(71)90098-9. [DOI] [PubMed] [Google Scholar]
- 7.Dobbins RL, Davis SN, Neal DW, Cobelli C, Jaspan J, Cherrington AD. Compartmental modeling of glucagon kinetics in the conscious dog. Metabolism. 1995;44:452–459. doi: 10.1016/0026-0495(95)90051-9. [DOI] [PubMed] [Google Scholar]
- 8.Endo Y. Suppression and potentiation of 5-hydroxytryptophan-induced hypoglycaemia by alpha-monofluoromethyldopa: correlation with the accumulation of 5-hydroxytryptamine in the liver. Br J Pharmacol. 1987;90:161–165. doi: 10.1111/j.1476-5381.1987.tb16836.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Galassetti P, Shiota M, Zinker BA, Wasserman DH, Cherrington AD. A negative arterial-portal vein glucose gradient decreases skeletal muscle glucose uptake in the conscious dog. Am J Physiol Endocrinol Metab. 1998;275:E101–E111. doi: 10.1152/ajpendo.1998.275.1.E101. [DOI] [PubMed] [Google Scholar]
- 10.Gijsman HJ, van Gerven JM, de Kam ML, Schoemaker RC, Pieters MS, Weemaes M, de Rijk R, van der Post J, Cohen AF. Placebo-controlled comparison of three dose-regimens of 5-hydroxytryptophan challenge test in healthy volunteers. J Clin Psychopharmacol. 2002;22:183–189. doi: 10.1097/00004714-200204000-00012. [DOI] [PubMed] [Google Scholar]
- 11.Goldstein RE, Reed GW, Wasserman DH, Williams PE, Lacy DB, Buckspan R, Abumrad NN, Cherrington AD. The effects of acute elevations in plasma cortisol levels on alanine metabolism in the conscious dog. Metabolism. 1992;41:1295–1303. doi: 10.1016/0026-0495(92)90099-v. [DOI] [PubMed] [Google Scholar]
- 12.Goldstein RE, Wasserman DH, McGuinness OP, Lacy DB, Cherrington AD, Abumrad NN. Effects of chronic elevation in plasma cortisol on hepatic carbohydrate metabolism. Am J Physiol Endocrinol Metab. 1993;264:E119–E127. doi: 10.1152/ajpendo.1993.264.1.E119. [DOI] [PubMed] [Google Scholar]
- 13.Hafdi Z, Couette S, Comoy E, Prie D, Amiel C, Friedlander G. Locally formed 5-hydroxytryptamine stimulates phosphate transport in cultured opossum kidney cells and in rat kidney. Biochem J. 1996;320:615–621. doi: 10.1042/bj3200615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hajduch E, Rencurel F, Balendran A, Batty IH, Downes CP, Hundal HS. Serotonin (5-hydroxytryptamine), a novel regulator of glucose transport in rat skeletal muscle. J Biol Chem. 1999;274:13563–13568. doi: 10.1074/jbc.274.19.13563. [DOI] [PubMed] [Google Scholar]
- 15.Heitkemper M, Jarrett M, Cain K, Shaver J, Bond E, Woods NF, Walker E, Bond EF, Chang L. Increased urine catecholamines and cortisol in women with irritable bowel syndrome. Am J Gastroenterol. 1996;91:906–913. [PubMed] [Google Scholar]
- 16.Hillsley K, Grundy D. Serotonin and cholecystokinin activate different populations of rat mesenteric vagal afferents. Neurosci Lett. 1998;255:63–66. doi: 10.1016/s0304-3940(98)00690-9. [DOI] [PubMed] [Google Scholar]
- 17.Horn CC, Friedman MI. Separation of hepatic and gastrointestinal signals from the common “hepatic” branch of the vagus. Am J Physiol Regul Integr Comp Physiol. 2004;287:R120–R126. doi: 10.1152/ajpregu.00673.2003. [DOI] [PubMed] [Google Scholar]
- 18.Hsieh PS, Moore MC, Neal DW, Emshwiller M, Cherrington AD. Rapid reversal of the effects of the portal signal under hyperinsulinemic conditions in the conscious dog. Am J Physiol Endocrinol Metab. 1999;276:E930–E937. doi: 10.1152/ajpendo.1999.276.5.E930. [DOI] [PubMed] [Google Scholar]
- 19.Huang W, Dedousis N, Bhatt BA, O’Doherty RM. Impaired activation of phosphatidylinositol 3-kinase by leptin is a novel mechanism of hepatic leptin resistance in diet-induced obesity. J Biol Chem. 2004;279:21695–21700. doi: 10.1074/jbc.M401546200. [DOI] [PubMed] [Google Scholar]
- 20.Ishioka K, Soliman MM, Sagawa M, Nakadomo F, Shibata H, Honjoh T, Hashimoto A, Kitamura H, Kimura K, Saito M. Experimental and clinical studies on plasma leptin in obese dogs. J Vet Med Sci. 2002;64:349–353. doi: 10.1292/jvms.64.349. [DOI] [PubMed] [Google Scholar]
- 21.Kubovcakova L, Krizanova O, Kvetnansky R. Identification of the aromatic L-amino acid decarboxylase gene expression in various mice tissues and its modulation by immobilization stress in stellate ganglia. Neuroscience. 2004;126:375–380. doi: 10.1016/j.neuroscience.2004.04.005. [DOI] [PubMed] [Google Scholar]
- 22.Lam NT, Lewis JT, Cheung AT, Luk CT, Tse J, Wang J, Bryer-Ash M, Kolls JK, Kieffer TJ. Leptin increases hepatic insulin sensitivity and protein tyrosine phosphatase 1B expression. Mol Endocrinol. 2004;18:1333–1345. doi: 10.1210/me.2002-0193. [DOI] [PubMed] [Google Scholar]
- 23.Li T, Leng XS, Weng SG, Peng JR, Wei YH, Mou DC, Wang WX, Zhu JY. Expression of 5-hydroxytamine receptors in hepatic stellate cell and action of 5-hydroxytamine on biological characteristics of hepatic stellate cell. Zhonghua Wai Ke Za Zhi. 2003;41:175–179. [PubMed] [Google Scholar]
- 24.Li Y, Wu XY, Zhu JX, Owyang C. Intestinal serotonin acts as paracrine substance to mediate pancreatic secretion stimulated by luminal factors. Am J Physiol Gastrointest Liver Physiol. 2001;281:G916–G923. doi: 10.1152/ajpgi.2001.281.4.G916. [DOI] [PubMed] [Google Scholar]
- 25.Mari A. Estimation of the rate of appearance in the non-steady state with a two-compartment model. Am J Physiol Endocrinol Metab. 1992;263:E400–E415. doi: 10.1152/ajpendo.1992.263.2.E400. [DOI] [PubMed] [Google Scholar]
- 26.Martin FJ, Miguez JM, Aldegunde M, Atienza G. Effect of streptozotocin-induced diabetes mellitus on serotonin measures of peripheral tissues in rats. Life Sci. 1995;56:51–59. doi: 10.1016/0024-3205(94)00407-j. [DOI] [PubMed] [Google Scholar]
- 27.Moore MC, DiCostanzo CA, Dardevet D, Lautz M, Farmer B, Neal DW, Cherrington AD. Portal infusion of a selective serotonin reuptake inhibitor enhances hepatic glucose disposal in conscious dogs. Am J Physiol Endocrinol Metab. 2004;287:E1057–E1063. doi: 10.1152/ajpendo.00313.2004. [DOI] [PubMed] [Google Scholar]
- 28.Moore MC, Geho WB, Lautz M, Farmer B, Neal DW, Cherrington AD. Portal serotonin infusion and glucose disposal in conscious dogs. Diabetes. 2004;53:14–20. doi: 10.2337/diabetes.53.1.14. [DOI] [PubMed] [Google Scholar]
- 29.Moore MC, Hsieh PS, Neal DW, Cherrington AD. Nonhepatic response to portal glucose delivery in conscious dogs. Am J Physiol Endocrinol Metab. 2000;279:E1271–E1277. doi: 10.1152/ajpendo.2000.279.6.E1271. [DOI] [PubMed] [Google Scholar]
- 30.Moore MC, Pagliassotti MJ, Wasserman DH, Goldstein R, Asher J, Neal DW, Cherrington AD. Hepatic denervation alters the transition from the fed to the food-deprived state in conscious dogs. J Nutr. 1993;123:1739–1746. doi: 10.1093/jn/123.10.1739. [DOI] [PubMed] [Google Scholar]
- 31.Niesler B, Frank B, Kapeller J, Rappold GA. Cloning, physical mapping and expression analysis of the human 5-HT3 serotonin receptor-like genes HTR3C, HTR3D and HTR3E. Gene. 2003;310:101–111. doi: 10.1016/s0378-1119(03)00503-1. [DOI] [PubMed] [Google Scholar]
- 32.Niijima A. Visceral afferents and metabolic function. Diabetologia Suppl. 1981;20:325–330. [PubMed] [Google Scholar]
- 33.Nishizawa M, Moore MC, Shiota M, Gustavson SM, Snead WL, Neal DW, Cherrington AD. Effect of intraportal glucagon-like peptide-1 on glucose metabolism in conscious dogs. Am J Physiol Endocrinol Metab. 2003;284:E1027–E1036. doi: 10.1152/ajpendo.00503.2002. [DOI] [PubMed] [Google Scholar]
- 34.Nomura T, Inoue K, Creveling CR, Komatsu F, Ohta N, Chino T, Karasawa N, Nagatsu I. Immunocytochemical localization of aromatic L-amino acid decarboxylase and catechol-O-methyltransferase in blood vessel wall of the human dental pulp. Brain Res. 1996;735:314–316. doi: 10.1016/0006-8993(96)00790-1. [DOI] [PubMed] [Google Scholar]
- 35.Omachi A, Ishioka K, Shibata H, Honjoh T, Kimura K, Saito M. Canine adiponectin: measurement of its serum/plasma concentration by ELISA and effects of obesity. Vet Biochem. 2004;41:31–37. [Google Scholar]
- 36.Pagliassotti MJ, Holste LC, Moore MC, Neal DW, Cherrington AD. Comparison of the time courses of insulin and the portal signal on hepatic glucose and glycogen metabolism in the dog. J Clin Invest. 1996;97:81–91. doi: 10.1172/JCI118410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Poldinger W, Calanchini B, Schwarz W. A functional-dimensional approach to depression: serotonin deficiency as a target syndrome in a comparison of 5-hydroxytryptophan and fluvoxamine. Psychopathology. 1991;24:53–81. doi: 10.1159/000284698. [DOI] [PubMed] [Google Scholar]
- 38.Pussard E, Guigueno N, Adam O, Giudicelli JF. Validation of HPLC-amperometric detection to measure serotonin in plasma, platelets, whole blood, and urine. Clin Chem. 1996;42:1086–1091. [PubMed] [Google Scholar]
- 39.Rahman MK, Nagatsu T, Kato T. Aromatic L-amino acid decarboxylase activity in central and peripheral tissues and serum of rats with L-DOPA and L-5-hydroxytryptophan as substrates. Biochem Pharmacol. 1981;30:645–649. doi: 10.1016/0006-2952(81)90139-8. [DOI] [PubMed] [Google Scholar]
- 40.Rahman MK, Nagatsu T, Kato T. Determination of aromatic L-amino acid decarboxylase in serum of various animals by high-performance liquid chromatography with electrochemical detection. Life Sci. 1981;28:485–492. doi: 10.1016/0024-3205(81)90141-7. [DOI] [PubMed] [Google Scholar]
- 41.Rattigan S, Dora KA, Colquhoun EQ, Clark MG. Serotonin-mediated acute insulin resistance in the perfused rat hindlimb but not in incubated muscle: a role for the vascular system. Life Sci. 1993;53:1545–1555. doi: 10.1016/0024-3205(93)90563-i. [DOI] [PubMed] [Google Scholar]
- 42.Satake S, Moore MC, Igawa K, Converse M, Farmer B, Neal DW, Cherrington AD. Direct and indirect effects of insulin on glucose uptake and storage by the liver. Diabetes. 2002;51:1663–1671. doi: 10.2337/diabetes.51.6.1663. [DOI] [PubMed] [Google Scholar]
- 43.Steele R, Wall JS, DeBodo RC, Altszuler N. Measurement of size and turnover rate of body glucose pool by the isotope dilution method. Am J Physiol. 1956;187:15–24. doi: 10.1152/ajplegacy.1956.187.1.15. [DOI] [PubMed] [Google Scholar]
- 44.Sumi-Ichinose C, Hasegawa S, Ichinose H, Sawada H, Kobayashi K, Sakai M, Fujii T, Nomura H, Nomura T, Nagatsu I, Hagino Y, Fujita K, Nagatsu T. Analysis of the alternative promoters that regulate tissue-specific expression of human aromatic L-amino acid decarboxylase. J Neurochem. 1995;64:514–524. doi: 10.1046/j.1471-4159.1995.64020514.x. [DOI] [PubMed] [Google Scholar]
- 45.United Kingdom Prospective Diabetes Study Group. Intensive blood-glucose control with sulphonylureas or insulin compared with conventional treatment and risk of complications in patients with type 2 diabetes (UKPDS 33) Lancet. 1998;352:837–853. [PubMed] [Google Scholar]
- 46.Yamada J, Sugimoto Y, Ujikawa M. Effects of insulin and adrenalectomy on elevation of serum leptin levels induced by 5-hydroxytryptophan in mice. Biol Pharm Bull. 2003;26:1491–1493. doi: 10.1248/bpb.26.1491. [DOI] [PubMed] [Google Scholar]
- 47.Yamada J, Sugimoto Y, Ujikawa M, Goko H, Yagura T. Hyperleptinemia elicited by the 5-HT precursor, 5-hydroxytryptophan in mice: involvement of insulin. Life Sci. 2003;73:2335–2344. doi: 10.1016/s0024-3205(03)00641-6. [DOI] [PubMed] [Google Scholar]
- 48.Yamakawa J, Takahashi T, Itoh T, Kusaka K, Kawaura K, Wang XQ, Kanda T. A novel serotonin blocker, sarpogrelate, increases circulating adiponectin levels in diabetic patients with arteriosclerosis obliterans. Diabetes Care. 2003;26:2477–2478. doi: 10.2337/diacare.26.8.2477. [DOI] [PubMed] [Google Scholar]
- 49.Yamauchi T, Kamon J, Minokoshi Y, Ito Y, Waki H, Uchida S, Yamashita S, Noda M, Kita S, Ueki K, Eto K, Akanuma Y, Froguel P, Foufelle F, Ferre P, Carling D, Kimura S, Nagai R, Kahn BB, Kadowaki T. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat Med. 2002;8:1288–1295. doi: 10.1038/nm788. [DOI] [PubMed] [Google Scholar]
- 50.Zhu MY, Juorio AV. Aromatic L-amino acid decarboxylase: biological characterization and functional role. Gen Pharmacol. 1995;26:681–696. doi: 10.1016/0306-3623(94)00223-a. [DOI] [PubMed] [Google Scholar]