

Abstract
The pure hereditary spastic paraplegias (HSPs) are a group of conditions in which there is progressive length-dependent degeneration of the distal ends of the corticospinal tract axons, resulting in spastic paralysis of the legs. Pure HSPs are most frequently inherited in an autosomal dominant pattern and are commonly caused by mutations in either the SPG4 gene spastin or in the SPG3A gene atlastin. To identify binding partners for spastin, we carried out a yeast two-hybrid screen on a brain cDNA library, using spastin as bait. Remarkably, nearly all of the positive interacting prey clones coded for atlastin. We have verified the physiological relevance of this interaction using co-immunoprecipitation, GST-pull down and intracellular co-localisation experiments. We show that the spastin domain required for binding to atlastin lies within the N-terminal 80 residues of the protein, a region that is only present in the predominantly cytoplasmic full-length spastin isoform. These data suggest that spastin and atlastin function in the same biochemical pathway and that it is the cytoplasmic function of spastin which is important for the pathogenesis of HSP. They also provide further evidence for a physiological and pathological role for spastin in membrane dynamics.
INTRODUCTION
The hereditary spastic paraplegias (HSPs) are a diverse group of genetically determined neurodegenerative conditions that all share the principal clinical feature of progressive spastic paralysis of the lower limbs. They are conventionally classified as pure or, when spastic paraplegia is accompanied by other prominent clinical manifestations, complex [1-3]. Pure HSP is generally regarded as the most common type in Northern Europe and North America [4]. In the pure HSPs, the most important cells involved are the corticospinal tract motor neurons (often termed “upper motor neurons”) and the clinical picture of progressive spastic paraplegia is reflected in pathological findings of progressive, length dependent, distal “dying-back” degeneration of the corticospinal tract axons [1]. Identification of the genes involved in HSPs and elucidation of the function of the corresponding proteins is beginning to provide important insights into molecular pathways that are important for axonal maintenance and motor neuronal function.
Eight autosomal dominant pure HSP loci have been mapped by genetic linkage studies [reviewed in 1, 2 and 5]. Rapid progress has been made in identifying the disease genes at these loci, with 5, atlastin (SPG3A), spastin (SPG4), NIPA1 (SPG6), KIF5A (SPG12) and HSP60 (SPG13) having been isolated to date [6-10]. Mutations in spastin and atlastin are the most frequent and are estimated to account for 40% and 10% of autosomal dominant pure HSP families respectively [5, 6, 11-13].
The spastin transcript is widely expressed within the nervous system and in non-neuronal tissues, but is most strongly expressed in subclasses of neurons [7, 14, 15]. Immunolocalisation studies in a variety of tissues and cell types have generally suggested that the protein has both a cytoplasmic and nuclear distribution [14, 15, 16, 17]. Recently it has become clear that this is because of differential localisation of two main isoforms of the protein, an N-terminal truncated form being predominantly nuclear, whereas a less abundant full-length form is actively exported from the nucleus to the cytoplasm [18]. This cytoplasmic distribution was described as punctate and perinuclear in one study [16], while in another, spastin was found in cytoplasmic cellular regions containing dynamic microtubules, which in a neuronal cell line included the distal axon and axonal branch points [17]. In human nervous tissues, spastin was located in the cytoplasm and synaptic terminals of several neuronal cell types, including motor neurons, although in other neuronal types it was detected in the nucleus [15].
With regard to the cytoplasmic function of spastin, two main, perhaps related, cellular roles have been proposed. Spastin is an AAA ATPase closely related in sequence to p60 katanin, a microtubule severing protein, and a variety of data suggest that spastin is likely to have a role in severing microtubules [19, 20]. In cultured cells expressing epitope-tagged spastin, expression of the protein was associated with the appearance of broken microtubule bundles within the cytoplasm. In these experiments, the protein localised to discrete punctate cytoplasmic structures not corresponding to known organelles, but reminiscent of the punctate staining seen in some antibody studies of the endogenous protein [19]. Further, expression of mutated ATPase-defective spastin resulted in striking cytoplasmic filaments that contained bundled microtubules [19, 21]. These results are compatible with the localization of endogenous spastin to cytoplasmic regions rich in dynamic microtubules. Data from Drosophila models also support a relationship between Drosophila spastin (D-spastin) and microtubule organisation in neurons and other tissues [22, 23, 24].
Aside from its role in microtubule regulation, it has also been suggested that spastin may be involved in intracellular membrane traffic events. This suggestion was first made when it was recognised that spastin has an N-terminal microtubule interacting and trafficking (MIT) domain [25, 26]. Most, if not all, other proteins that contain this domain have roles in membrane traffic [26]. Evidence for involvement of spastin in membrane traffic events was recently strengthened by our identification of the endosomal protein CHMP1B as a binding partner for spastin [27]. CHMP1B localises to endosomes and is associated with ESCRT-III (endosomal sorting complex required for transport-III), a protein complex which functions in the sorting of mono-ubiquinated proteins to the multivesicular body, a pre-lysosomal endocytic compartment [reviewed in 27]. This study also identified a Golgi membrane protein, gp25L2, as a possible binding partner for spastin, although this interaction has not yet been characterised fully [27]. Since many membrane traffic events are linked to microtubule-based transport, it is not inconceivable that the membrane traffic and microtubule regulating roles of spastin may be inter-related.
The atlastin gene consists of 14 exons which code for a protein that is 558 amino acids long [6]. The gene is expressed in many body tissues, with highest expression in brain and spinal cord [6]. With rare exceptions, mutations in atlastin cause childhood onset HSP. Most atlastin mutations published to date have been missense mutations, with one frameshift mutation described [28-34].
The atlastin protein is a novel GTPase that is similar to members of the dynamin family of large GTPases [6]. It has two putative transmembrane domains with membrane topology resulting in the N- and C-termini being exposed to the cytoplasm [35]. The GTPase site is active and is formed from residues at 3 motifs, the P-loop, DxxG and RD motifs. The protein may oligomerise to form homotetramers [35].
The cellular distribution of atlastin has been studied in rat brain sections [35]. Here, atlastin positive cells were most abundant in lamina V of the cerebral cortex, with both the neuronal soma and dendritic tree containing atlastin. At the subcellular level in cultured rat subcortical neurons, atlastin was predominantly located in the cis-Golgi apparatus, with punctate staining also seen in axons and dendrites. Some limited co-localisation with endoplasmic reticulum (ER) markers was also seen [35]. The precise functional role of atlastin at the Golgi membrane or in other membrane organelle sites remains to be clarified.
In this study, we set out to gain additional insights into the function of spastin, by identifying new binding partners for the protein. Our previous spastin yeast two-hybrid screen was directed against an erythroleukaemia cDNA library [27]. In order to identify binding partners which might be important for spastin’s function in the central nervous system, we repeated the yeast two-hybrid screen using a fetal brain cDNA library. Remarkably, nearly all of the positive interacting clones contained atlastin cDNA, strongly suggesting that these two HSP proteins interact with each other. We have confirmed this potential interaction by co-immunoprecipitation and pull-down experiments, along with co-localisation studies using epitope-tagged wild-type and mutant proteins in neuronal and non-neuronal cell lines. Together, these data provide the first experimental evidence that two proteins involved in HSP physically interact. They suggest that spastin and atlastin participate in the same biochemical pathway and that the molecular pathology of spastin and atlastin HSP is related. They also provide further evidence that spastin is involved in membrane traffic events.
RESULTS
Yeast two-hybrid, GST pull-down and co-immunoprecipitation experiments identify atlastin as a binding partner of spastin
We used full-length spastin protein as a ‘bait’ to screen a high-complexity human yeast two-hybrid fetal brain cDNA library (Clontech), using methods described previously [27]. This screen generated 15 positive diploid clones that grew on selective media lacking adenine and were positive for β-galactosidase activity. When these primary interactions were retested in fresh yeast, 8 were found to be both reproducible and bait specific, giving strong β-galactosidase activity and strong growth on selective medium lacking tryptophan, leucine and adenine (-W/L/A). Remarkably, all but two of the positive ‘prey’ clones contained atlastin coding sequence.
In order to verify the relevance of the putative interaction between spastin and atlastin, we first carried out glutathione-S-transferase (GST) pull-down experiments. GST-spastin successfully pulled down atlastin from a lysate of cells transiently transfected with a mammalian expression vector containing c-Myc-tagged atlastinΔTM (myc-atlastinΔTM; figure 1), indicating that the two proteins are capable of a biochemical interaction (figure 2A). We next characterised the spastin protein domains necessary for interaction with atlastin, by using yeast-two hybrid and GST pull-down experiments. We tested a prey atlastin vector against yeast two-hybrid bait vectors containing inserts coding for several truncated forms of spastin, spastinΔAAA, spastinΔN1 and spastinΔN2 (figure 1). We found that yeast two-hybrid interaction was maintained with the spastinΔAAA bait vector, but lost when the spastinΔN1 and spastinΔN2 baits were used (figure 2B). Pull-down assays using GST-atlastinΔTM and cell lysates from cells transfected with the mutant forms of spastin were also consistent with these results; GST-atlastinΔTM was able to pull-down myc-spastinΔAAA and myc-spastinK388R (a form of spastin containing a disease-associated missense mutation), but not myc-spastinΔN1 or spastinΔN2-GFP, from lysates of cells transfected with the relevant construct (figure 2C). These data indicate that the region of spastin responsible for the interaction with atlastin lies at the N-terminal of the protein, from residues 1 to 80. Recent data have shown that spastin has two translation initiation sites, the first at codon 1 and the second at codon 87, resulting in the production of two main spastin isoforms, a rare, predominantly cytoplasmic full-length form (approximately 68kD size) and a much more abundant shorter, predominantly nuclear isoform (approximately 60kD) [18]. The situation is further complicated by the presence of alternative splicing events which result in the presence of minor exon 4-deleted versions of the long and short isoforms (at approximately 64kD and 55kD respectively). Our domain interaction data therefore suggest that only the long (i.e. 68 and 64kD) isoform version of spastin are capable of interacting with atlastin.
Figure 1. Constructs used in this study.

Schematic diagram showing the domain structure of wild-type (WT) spastin and atlastin, along with the structure of other inserts used in constructs. Amino acid numbering is given above each construct, while the position of the K388R point mutation examined in the study is shown in grey font above the WT spastin illustration. HR=hydrophobic region, MIT=microtubule interacting and trafficking domain, AAA=AAA ATPase cassette, GTPase=GTPase cassette, TM=transmembrane domain.
Figure 2. Spastin and atlastin interact biochemically.

A) GST-spastin was used in a pull-down experiment using cell lysate from HeLa cells transfected with myc-atlastinΔTM. A band the correct size for myc-atlastinΔTM was seen in the pull-down lane (lane 4), but not in lanes from experiments lacking GST protein (lane 1), experiments using empty-GST instead of GST-spastin (lane 2), or experiments in which no cell lysate was added (lane 3). B) Investigation of spastin domains responsible for the interaction with atlastin was carried out using yeast two-hybrid. Control column 1 shows the growth of diploid colonies on media lacking tryptophan and leucine (-W/L) to select for the presence of both bait and prey vectors. Column 2 shows the growth of diploid colonies on media lacking tryptophan, leucine and adenine (-W/A/L), which requires the activation of the Ade reporter gene, while control column 3 shows the growth of diploid colonies on media lacking tryptophan, leucine and histidine (-W/H/L), which requires the activation of the His reporter gene. In each case spastin or a spastin mutant was used as the ‘bait’ protein, while atlastin was used as ‘prey’. Full-length spastin and spastinΔAAA interact with atlastin, but spastinΔN1 and spastinΔN2 do not. No interaction was present with a non-specific prey (LSM2). C) GST-pull down experiments using GST-atlastinΔTM versus cell lysates from cells transfected with the stated constructs confirmed the yeast two-hybrid results. The GST-fusion protein was able to pull-down myc-spastinΔAAA and myc-spastinK388R, but not myc-spastinΔN1 or spastinΔN2-GFP. D) Co-immunoprecipitation of transiently transfected myc-spastin and atlastin-GFP in HEK293T cells. 24 hours post transfection, tagged proteins were immunoprecipitated (IP) using an anti-GFP polyclonal antibody and immunoblotted (IB) with an anti-myc monoclonal antibody. A band of the expected size for transfected full-length myc-spastin was co-immunoprecipitated with atlastin-GFP. No band was present when the anti-GFP antibody was omitted from the immunoprecipitation reaction. E) Attempted co-immunoprecipitation of transiently transfected myc-spastin and vector only EGFP as a negative control for D). Myc-spastin was not co-immunoprecipitated with GFP. For parts D) and E), input lanes show α-myc and α-GFP immunoblots of input fractions, confirming expression of the relevant proteins in the cell lysates used for the immunoprecipitations. F) Endogenous spastin and atlastin are present in NSC34 cells. Spastin was identified using the S51 spastin antibody. This antibody reveals 3 bands in this blot, corresponding to different isoforms of spastin- full-length (68kD, arrow to right of lane marks position), short form (60kD) and short form variant lacking exon 4 (55kD). The full-length isoform lacking exon 4 (64kD) is of such low abundance that it is not seen on this blot. Using the 5409 anti-atlastin antibody, a single band of approximately 55kD was seen. G) Under denaturing conditions, atlastin is strongly immunoprecipitated from NSC34 cells by the 5409 atlastin antibody. The left lane shows the input cell lysate, while the right lane shows the immunoprecipitate. Both were blotted with the atlastin antibody. H) Endogenous atlastin and myc-spastin are co-immunoprecipitated from NSC34 cells. Anti-atlastin 5409 antibody was used to immunoprecipitate under denaturing conditions and immunoblotting was performed using anti-myc. A band of the size expected for myc-spastin is present in the input lysate and in the immunoprecipitate. This band was absent when a spurious control antibody (anti-GFP) was used to carry out the immunoprecipitation. The asterisks mark the position of immunoglobulin bands- these were also present when the primary anti-myc antibody was omitted from the immunoblotting procedure (not shown). The position of relevant marker bands is shown to the left of each blot.
We next attempted to verify the physiological relevance of the spastin-atlastin interaction by co-immunoprecipitation experiments. We co-immunoprecipited epitope-tagged myc-spastin and atlastin-GFP, by transiently expressing these proteins in HEK293T cells and immunoprecipitating with anti-GFP antibody. Subsequent immunobloting of the precipitate with anti-myc revealed a band of the size expected for myc-spastin (c. 68kD; figure 2D). This band was not present when the GFP antibody was omitted from the immunoprecipitation reaction, or when the immunoprecipitation was carried out using cells co-transfected with myc-spastin and GFP-empty vector (figure 2E).
Finally, we attempted to carry out co-immunoprecipitation experiments using antibodies to endogenous spastin and atlastin. We used two published antibodies in these experiments. The first was S51, a rabbit polyclonal anti-spastin antibody (a kind gift of Elena Rugarli) [17]. This antibody can detect the main spastin isoforms in NSC34 cell lysates (figure 2F) and is capable of immunoprecipitating spastin. The second antibody was a rabbit polyclonal anti-atlastin antibody (5409- a kind gift of Craig Blackstone) [35]. This antibody detects a single strong band at approximately the size predicted for atlastin (c. 55kD) on Western blotting of NSC34 cell lysate (figure 2F). On immunoprecipitation of NSC34 cells under non-denaturing conditions using either the spastin S51 antibody or the atlastin antibody, we were unable to demonstrate any co-immunoprecipitation of the endogenous putative binding partner. Since we were concerned that immunoprecipitation of atlastin by the atlastin antibody was weak under non-denaturing conditions, we investigated the capacity of this antibody to immunoprecipitate atlastin from NSC34 cells under denaturing conditions and found strong immunoprecipitation (figure 2G). We therefore carried out co-immunoprecipitation experiments under denaturing conditions, treating the cells with the chemical crosslinker DSP before lysis, in order to preserve protein-protein interactions. However, we were still unable to demonstrate any co-immunoprecipitation of endogenous spastin (data not shown).
We hypothesized that the reason we could not demonstrate any interaction between the endogenous proteins was because of the very low abundance of full-length 68kD spastin (figure 2F), the form capable of interacting with atlastin. In view of this, we explored whether provision of additional full-length exogenous spastin might allow us to demonstrate co-immunoprecipitation with endogenous atlastin and we found that this was indeed the case. When we used the atlastin antibody under denaturing conditions to immunoprecipitate atlastin from NSC34 cells expressing myc-spastin, we found that myc-spastin was co-immunoprecipitated (figure 2H). No myc-spastin was co-immunoprecipitated when a spurious immunoprecipitating antibody was used (figure 2H).
In summary, we were able to identify an interaction between spastin and atlastin by yeast two-hybrid, GST-pull down and co-immunoprecipitation experiments. These results provide very strong evidence that spastin and atlastin are capable of binding to each other, and that this interaction has physiological relevance.
Epitope tagged atlastin and spastin co-localise in HeLa and NSC34 cells
Having established the presence of an interaction between spastin and atlastin, we next went on to determine whether there was any co-localisation between the proteins in mammalian cells, and if so where the site of the interaction was, by examining the intracellular distribution of epitope-tagged proteins.
Before examining whether spastin and atlastin showed intracellular co-localisation, we established the intracellular distribution in mammalian cells of epitope-tagged versions of atlastin, since this has not previously been described. To minimise the potential for tag-induced changes in distribution, we made constructs expressing either N-terminal c-Myc- or C-terminal GFP-tagged versions of full-length atlastin and examined the expression pattern of each construct by immunofluorescence using a confocal microscope. In each case, epitope-tagged atlastin appeared in a reticulate pattern throughout the cytoplasm of transiently transfected HeLa cells. This reticulate staining pattern strongly co-localised with calreticulin, a marker of the endoplasmic reticulum (ER) (figure 3A-F), and with antibodies to two other ER proteins, PDI (protein disulphide isomerase), a lumenal protein, and sec61α, an ER membrane protein (data not shown). In view of the interaction between spastin and atlastin, and the relationship between spastin and microtubules, we also examined whether atlastin co-localised with tubulin. We found partial co-localisation between epitope-tagged atlastin and the anti-tubulin antibody YL1/2 (figure 3G-I), although there was no gross effect of atlastin expression on microtubule architecture. This co-localisation between atlastin and microtubules is perhaps not surprising, since the ER is known to have a close association with the microtubule framework. We also tested for co-localisation between epitope-tagged atlastin and a battery of other antibody markers, which included EEA1 (for early endosomes), M6PR (for late endosomes and trans-Golgi network), Lamp1 (for lysosomes), GM130 (for the Golgi apparatus) and TGN46 (for the trans-Golgi network). We found some limited co-localisation between atlastin-GFP and M6PR (figure 3J-L) and GM130 (figure 3M-O), although we cannot be sure whether this might simply reflect the fact that the organelles these markers detect are located close to the microtubule organising centre, where the expression of ER markers is also very intense.
Figure 3. Epitope-tagged wild-type and mutant atlastin co-localises with ER and microtubule markers.

The images show Hela cells labelled with the antibody marker shown in panels A, D, G, J, M. Panels B, E, H, K, N show the expression pattern in the same cells, transiently transfected with the construct shown in each panel. The colour of text in each of these panels represents the image colour in the corresponding merged panels, C, F, I, L, O. Both N-terminal myc-tagged (panels A-C) and C-terminal GFP-tagged (panels D-F) atlastin co-localise tightly with calreticulin. Wild-type atlastin also partially co-localises with the microtubule marker YL1/2 (panels G-I). This co-localisation was strongest in the perinuclear region. There is also partial co-localisation with the late endosomal mannose 6 phosphate receptor (M6PR, panels J-L) and the Golgi marker GM130 (panels M-O), particularly in a focal perinuclear region where atlastin showed strong expression. Cells were fixed 24 hours after transfection. Size bars indicate 10μm.
In addition to experiments in HeLa cells, we examined the expression pattern of epitope-tagged atlastin in a mouse motor neuronal cell type, NSC34. This hybrid cell type is derived from a fusion between mouse spinal cord motor neurons and a neuroblastoma cell line and has many of the anatomical, pharmacological and electrical characteristics of neuronal cells [36]. For c-Myc and -GFP tagged atlastin, the expression pattern in NSC34 cells was very similar to that of the corresponding protein in HeLa cells. Atlastin expression was present both in the cell body and neurite extensions (data not shown).
Having established the intracellular distribution of epitope-tagged atlastin, we examined co-localisation of epitope-tagged spastin and atlastin in HeLa cells (figure 4A-G) and NSC34 cells (figure 4H-J). We found partial but definite co-localisation between the spastin and atlastin proteins and this did not depend on the nature or location of the epitope tag used. The co-localisation in HeLa cells was typically in cytoplasmic puncta/small tubules, which also co-localised with the ER marker calreticulin (figure 4A-G). In NSC34 cells, co-localisation occurred in the soma and in neurites (figure 4 H-J).
Figure 4. Epitope-tagged atlastin and spastin co-localise in HeLa and NSC34 cells.

Hela cells (panels A-G, K-W) or NSC34 cells (panels H-J) were co-transfected with myc-spastin and atlastin-GFP (panels A-J), myc-spastin alone (panels K-M), myc-spastinK388R and atlastin-GFP (panels N-Q), myc-spastinK388R alone (panels R-T), or myc-spastin and GFP-VPS4-EQ (panels U-W). The cells shown in panels D-G and K-T were also labelled with the ER marker anti-calreticulin (shown in panels F, L, P, S). Panels C, G, J, M, Q, T and W show the merged images of the cells shown in panels A and B, D-F, H and I, K and L, N-P, R and S, U and V respectively, with the colour of the text in each panel indicating the image colour in the merged picture. In each of the cells shown in panels A-J there is definite co-localisation between the spastin and atlastin proteins. Both the co-localised proteins (panels D-G) and myc-spastin expressed alone (panels K-M) also co-localise with calreticulin. In NSC34 cells (panels H-J), co-localisation was seen in both the soma and neurite extensions. In cells expressing mutant spastin and wild-type atlastin (panels N-Q), atlastin and calreticulin are redistributed to co-localise with the mutant spastin in filaments. Exogenous atlastin expression is not necessary for calreticulin to be redistributed in this way (panels R-T). A proportion of myc-spastin expression was also on endosomes, since it partially co-localised with GFP-VPS4-EQ (panels U-W, arrowheads). Cells were fixed 24 hours after transfection. Size bars indicate 10μm.
Since epitope-tagged spastin co-localised with atlastin on the ER, we next determined whether some epitope-tagged spastin is present on the ER in the absence of exogenous atlastin, or whether expression of atlastin caused a redistribution of spastin to the ER. We found significant co-localisation in puncta and tubules of a subset of myc-spastin with calreticulin (figure 4K-M), but this was not as strong as the ER localisation when myc-spastin was co-expressed with atlastin-GFP (figure 4D-G), suggesting that co-expression with atlastin caused recruitment of spastin to the ER, perhaps because additional binding sites were provided there by atlastin expression. When GFP-spastin and spastin-GFP were expressed alone, co-localisation with calreticulin was also present, but subjectively more limited than with myc-spastin.
We next examined whether atlastin co-localised with ATPase defective forms of spastin. We examined the effects of atlastin expression on a disease-causing spastin mutation, K338R. The spastin K388R mutation abrogates ATPase function and is associated with the development of an abnormal, filamentous phenotype when expressed in cultured cells. The abnormal filaments partially co-localise with markers of microtubules [19, 27]. We found that the spastin mutant retained a filamentous appearance when co-expressed with epitope-tagged atlastin and that the distribution of atlastin was dramatically changed so that the protein showed strong co-localisation with the abnormal filaments (figure 4N-Q). In NSC34 cells, the abnormal filaments extended into neurite processes, and strong co-localisation was seen here, as well as in the soma (data not shown). The redistribution of atlastin was accompanied by a redistribution of calreticulin, which also co-localised with the mutant spastin filaments (figure 4P). A similar redistribution of the ER was seen when myc-spastinK388R was expressed in HeLa cells in the absence of exogenous atlastin (figure 4R-T).
We have previously suggested that some spastin expression may be on endosomes, based on data that show an interaction between spastin and the endosomal protein CHMP1B. We have also described colocalisation between myc-spastin and a GFP-tagged endosomal marker RhoB [27]. However, our previous co-localisation experiments used a GFP-tagged RhoB construct (Clontech) that was inappropriate, since it contained a c-Myc epitope. We therefore reinvestigated the endosomal localisation of transiently expressed spastin by examining HeLa cells co-transfected with myc-spastin and N-terminal GFP-tagged mammalian VPS4-E235Q (a kind gift of Paul Whitely) [37]. The ATPase-defective GFP-VPS4-E235Q protein accumulates on swollen endosomes which retain components of the ESCRT-III complex on their limiting membranes [see 37]. We found significant partial co-localisation between myc-spastin and GFP-VPS4-E235Q, strongly suggesting that a subpopulation of spastin is present on endosomal membranes (figure 4U-W). It therefore seems likely that spastin can be present on at least two membrane populations, endosomes and the ER.
In order to gain more information on the composition of the filaments associated with ATPase-defective spastin, we investigated them at the ultrastructural level. We transiently co-transfected HeLa cells with myc-spastinK388R and atlastin-GFP and examined the cells under the electron microscope. We found bundles of microtubules in the cytoplasm of co-transfected cells (figure 5A). Individual microtubules within these bundles had an abnormal appearance, having electron-dense striations. We also saw cytoplasmic structures with an abnormal honeycomb-like appearance in these cells (figure 5B) and these structures were sometimes continuous with the ER (figure 5B) or nuclear envelope (figure 5C). In some sections the honeycomb structures were contiguous with the abnormal microtubule bundles (figure 5D).
Figure 5. The ultrastructure of the filaments associated with ATPase-defective spastin.

At the electron microscope level, cells co-transfected with myc-spastinK388R and atlastin-GFP had abnormal bundles of microtubules in the cytoplasm (panel A). The microtubules had an unusual electron dense coating. Dense honeycomb structures were also seen and in places these were continuous with the ER (arrowhead, panel B), or with the nuclear membrane (arrows, panel C). In some cases they were contiguous with the abnormal microtubule bundles (arrow, panel D). All panels show HeLa cells that were fixed 24 hours after transfection. Size bar represents 200nm, all pictures are at the same magnification.
DISCUSSION
In this study we used a yeast two-hybrid approach to identify potential interacting partners for spastin, a protein mutated in autosomal dominant pure hereditary spastic paraplegia. Remarkably, this approach identified atlastin, another protein mutated in autosomal dominant pure hereditary spastic paraplegia, as a potential spastin binding partner. We verified the relevance of this interaction in mammalian cells by a variety of approaches, which together provide compelling evidence that spastin and atlastin are true binding partners. This interaction is the first demonstration of a direct functional relationship between two proteins involved in HSP. Its presence strongly suggests that spastin and atlastin participate in the same biochemical pathway and that the molecular pathology of spastin- and atlastin- HSP is linked.
One issue that our study was not able to conclusively address was the subcellular site of the spastin-atlastin interaction. Our data with epitope-tagged proteins suggests that they interact on the ER, since we found that epitope-tagged atlastin strongly co-localised with ER markers (and to a lesser extent with the Golgi apparatus), that epitope-tagged full-length spastin showed partial, but definite, co-localisation with the ER marker calreticulin, and that when co-expressed, epitope-tagged full-length spastin and atlastin co-localised with each other and with calreticulin. It is possible that a subset of endogenous spastin might be located on the ER or the contiguous microtubule network - previous immunolocalisation studies have not specifically examined whether a proportion of the cytoplasmic expression of spastin might be on or very close to the ER. Although a previous immunolocalisation study found some endogenous atlastin on the ER, the main location of the endogenous protein was on the Golgi apparatus, a site which was less prominent with epitope-tagged atlastin [35]. Explanations for this discrepancy could include an immunofluorescence cross-reaction of the atlastin antibody with another protein, the presence of ER and Golgi forms of atlastin with different sensitivity/available epitopes for the antibody, or effects of the epitope tag on localisation of the protein. However, the localisation that we observed was independent of the site or nature of the epitope-tag used. Transient expression of atlastin may have had an effect on its distribution, with over-expression saturating ER to Golgi transport pathways or affecting the balance between anterograde and retrograde ER/Golgi pathways, resulting in a relative increase in ER staining compared to the endogenous pattern. Finally, the continuous distribution of atlastin that we saw throughout the ER was not typical of the aggregated clumps characteristic of over-expressed proteins that are misfolded in the ER. Further studies with additional spastin and atlastin antibodies will be required to conclusively resolve the subcellular location of the spastin-atlastin interaction.
Whether the localisation of native atlastin is predominantly at the ER, at the Golgi, or at both, the fact that it interacts with spastin makes it very likely that spastin is active at more than one membrane site. We have previously identified the endosomal protein CHMP1B as a binding partner for spastin and we show here that full-length spastin partially co-localises with GFP-VPS4-E235Q, an endosomal marker [27]. A picture is therefore emerging of spastin as a microtubule regulating protein that can be recruited by different adaptor proteins to several membrane sites, where it may play a role in local regulation of microtubules, perhaps coupled to movement of membrane bound organelles (especially in axons) and/or the re-modelling of organelle architecture, or even have an entirely separate role unrelated to its microtubule severing function. The factors that influence recruitment of spastin to one site versus another are not yet known, but their elucidation will be important in further understanding the exact physiological role of the protein.
Spastin may also have other functions unrelated to membrane interactions. Several studies using antibodies to endogenous spastin have suggested that, in some cell types or at some stages of the cell cycle, spastin may have a nuclear localisation [14, 15, 17]. This nuclear spastin predominantly consists of the short N-terminal truncated isoform and its function is currently unknown [18]. The scenario of an AAA family member having multiple sites of action is not unprecedented, with several documented examples of individual AAA proteins exerting a specific function, often involving regulation of substrate protein conformation, at multiple intracellular sites of action [38, 39].
We examined the spastin domains that mediate the interaction with atlastin and the region responsible resided within the first 80 residues of the protein (figure 6). Thus, only the predominantly cytoplasmic, full-length spastin isoform contains the region responsible for binding to atlastin, suggesting that it is the function of this isoform which is of direct importance for the pathogenesis of HSP. Interestingly, the full-length isoform is enriched in brain and spinal cord, the anatomical sites of HSP pathology [18]. The atlastin binding region overlaps the domain (residues 50-87) that is responsible for binding to microtubules and the centrosomal protein NA14, but is distinct from the region that interacts with CHMP1B (residues 81-194) [17, 19, 27]. Thus it appears that the different spastin locations and functions discussed above are reflected in the presence of binding sites for different adaptor proteins in its N-terminal region.
Figure 6. Cartoon diagram of interaction between spastin and atlastin.
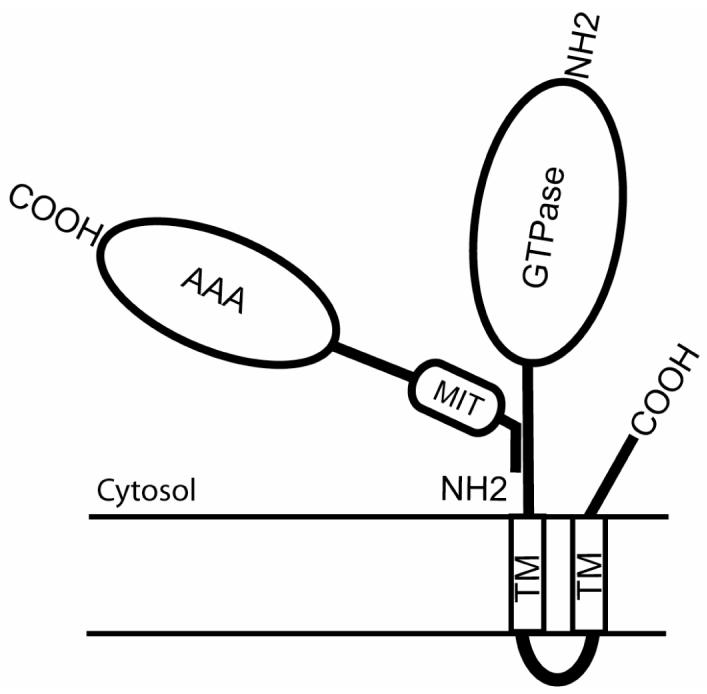
The region of atlastin responsible for binding to spastin lies N-terminal to the first transmembrane domain (TM). For spastin, the region responsible for binding to atlastin resides within the first 80 residues of the protein. This overlaps the region of the protein (residues 50-87) that is responsible for binding to microtubules, but is distinct from the MIT and AAA ATPase domains.
Our study also presents more information on the composition of the intracellular filaments seen on expression of ATPase-defective spastin mutants. Although it is unlikely that these filaments are relevant to those spastin mutations which cause haplo-insufficiency and it is possible that they represent an over-expression artefact, their composition may be relevant to HSP caused by spastin ATPase missense mutations. At the light microscope level, epitope-tagged atlastin and the ER marker calreticulin were both redistributed to the abnormal microtubule bundles. On electron microscopy in cells transfected with myc-spastinK388R and atlastin-GFP, we observed bundled microtubules and abnormal honeycomb structures, which were sometimes seen in association with microtubule bundles. In some cases, it was apparent that the honeycomb structures were continuous with normal ER and with the nuclear envelope, suggesting that they may represent abnormal ER. It is not clear whether the reorganisation of the ER that we have seen is a consequence of direct binding between mutant spastin and atlastin, or whether it is an indirect effect via reorganisation of the microtubule network associated with the ER, although it seems clear that the filaments are closely associated with membrane components.
In summary, we have identified two HSP proteins, spastin and atlastin, as binding partners. This suggests that the pathogenesis of HSP caused by mutation in these two proteins is fundamentally related and provides the first experimental evidence that HSP may be caused by abnormalities in proteins that are members of the same biochemical pathway. A more detailed understanding of the function of this pathway will not only give key insights into the causes of neurodegeneration in HSP and perhaps other neurological conditions, but should also elucidate molecular mechanisms that are important for long tract neuronal maintenance.
MATERIALS AND METHODS
Construction of Yeast two-hybrid, GST- and Immunofluorescence Vectors
Spastin yeast two-hybrid and mammalian expression constructs were constructed as described previously [27, 40]. Atlastin constructs were amplified by proof-reading Pfx polymerase (Invitrogen) PCR from IMAGE clone 3869877, using Gateway™ (Invitrogen) recombination cloning system-compatible primers designed from the reference cDNA sequence for each gene. For truncated cDNA constructs, Pfx PCR was carried out using appropriate Gateway™ (Invitrogen) recombination cloning system-compatible primers designed from within the reference cDNA. Gateway™ recombination cloning system pENTR201 and/or pENTR207 entry vectors containing the amplified cDNAs were constructed according to the manufacturer’s instructions. Sequence of entry vector constructs was verified by sequencing on an ABI377 or 3700 sequencer, using BigDyeDT chemistry (Applied Biosystems).
The atlastin transcripts were then subcloned into Gateway™-compatible vectors pcDNA3-C-Myc and/or, pE-GFP-N, which contained 5′ -myc, or 3′ -GFP epitopes, respectively. The orientation of the atlastin sequences within the constructs was verified by direct sequencing on an ABI377 or 3700 sequencer.
For production of the GST-fusion cDNA constructs, full length spastin cDNA and atlastinΔTM cDNA were amplified by PCR using the Pfu Turbo® polymerase enzyme (Stratagene). The primers used in the reaction were designed to produce a PCR product with 5′ and 3′ ends compatible with the GST gene fusion vector, pGEX-4T-3 (Amersham Biosciences). After restriction with Not-I and Sal-I (New England Biolabs), the PCR product was ligated into the expression vector using T4 DNA ligase (New England Biolabs). The sequence of the constructs was verified as above.
Production of GST-fusion proteins
Transformation of the GST-fusion construct into BL21(DE3)pLysS cells was performed according to the manufacture’s instructions (Invitrogen). Production of large-scale bacterial sonicates and subsequent GST-fusion protein purification was performed according to the Amersham Biosciences’ GST gene fusion system instructions.
Yeast two-hybrid library screening, reporter assays and identification of interacting proteins
Yeast two-hybrid screening with the spastin bait clone was carried out as previously described, except the screen was carried out against a fetal brain cDNA Matchmaker library (Clontech) [27]. To identify interacting proteins, prey inserts were directly amplified from yeast using vector-specific primers, as previously described, and 3-5μl of each PCR product was sequenced as described above [27].
Prey sequences were searched against locally held versions of the Homo sapiens Unigene and the EMBLminus databases using an automated BLAST [41] algorithm. Custom-built Perl modules and scripts were used to prefilter and format the raw BLAST output, and determine whether the 5′ sequence read overlapped with the protein coding region of a gene. Only matches to known protein coding regions and to ESTs that have no defined open reading frame were included as positives. Matches to 3′ UTRs and genomic DNA with no associated gene prediction were excluded. Singleton, unspliced ESTs that did not correspond to any gene predictions were also excluded.
Reconfirmation and specificity testing for yeast two-hybrid assays
To retest each interaction in fresh yeast, prey PCR product was cloned using Gateway™ recombination cloning system into the Gateway™ compatible pGAD-G vector, as described above. Competent MATα PJ69-4A yeast cells were transformed and mated as previously described. They were mated with either the relevant bait, two different irrelevant baits or an empty bait vector, and then incubated for >5 hours on YPAD plates. Diploids were selected on SD -Ura/Leu prior to testing for reporter activation by replicating onto SD -Ura/Leu/Ade.
Antibodies
Monoclonal anti-myc primary antibody (mouse clone 4A6) was obtained from Upstate (New York). Monoclonal anti-microtubule antibody (rat clone YL1/2) and anti-GFP rabbit polyclonal antibodies were obtained from abcam (Cambridge). Rabbit polyclonal anti-calreticulin antibody was obtained from Calbiochem (San Diego). Sheep polyclonal anti-TGN46 was obtained from Serotec (Oxford), rabbit polyclonal anti-M6PR and mouse monoclonal anti-Lamp1 antibodies were available in house. Mouse monoclonal anti-EEA1 and mouse monoclonal anti-GM130 were obtained from BD Transduction Laboratories (Oxford). Peroxidase conjugated secondary antibodies for western blotting were obtained from Sigma. Alexafluor-labelled secondary antibodies for immunofluorescence were obtained from Molecular Probes (Oregon). The S51 rabbit anti-spastin polyclonal antibody was a kind gift of Elena Rugarli, the rabbit anti-atlastin 5409 antibody was a kind gift of Craig Blackstone.
Cell Culture, Transfection and Immunofluorescence
HeLa cells were seeded onto sterile glass coverslips in 6 well plates (approximately 0.5×105 cells/well). After 24 hours the cells were transfected with approximately 400ng of vector DNA, using the Effectene® transfection reagent (Qiagen), according to the manufacturer’s instructions. Following transfection, cells were incubated for 24 or 48 hours in RPMI-1640 medium (Sigma) containing 10% fetal bovine serum (FBS) (Sigma), 100U/ml penicillin and 100μg/ml streptomycin (Sigma) and 2mM l-glutamine (Sigma). NSC34 cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) (Life Technologies), supplemented in the same way as the RPMI-1640 medium used for HeLa cell. NSC34 cells were seeded at 1×105 into 6 well plates containing glass coverslips pre-coated with Poly-D-Lysine (Sigma). They were then transiently transfected with Effectene® transfection reagent (Qiagen), according to the manufacturer’s instructions, for 24-48 hours before fixation.
Following transfection, cells were washed in PBS (Sigma) and fixed with 4% formaldehyde in PBS, at room temperature for 15 minutes. Fixed cells were washed in PBS before being permeabilized in PBS containing 0.1% Triton X-100 (Sigma) or 0.05% saponin (Sigma- used with M6PR and Lamp1 antibodies) at room temperature for 5-10 minutes. Permeabilized cells were then washed 3 times in PBS, incubated for 15 minutes in a blocking solution (PBS, 10% FBS, ±0.05% saponin) and then transferred to blocking solution containing the appropriate epitope-specific antibody at an appropriate dilution. After 60 minutes incubation, coverslips were washed 3 times in blocking solution and were then incubated for 60 minutes in blocking buffer containing the appropriate secondary antibodies, at a concentration of 1/300. Cover slips were then washed 3 times in PBS and once in distilled H2O, after which they were mounted in Vectashield™ (Vector Laboratories Inc.) medium on a glass slide. Stained samples were analysed on a Zeiss 510 Meta confocal microscope. Images were recorded using LSM Image Analyzer software and data were subsequently processed using Adobe Photoshop and Illustrator programmes.
Electron Microscopy
Fixation and processing of sections for microscopy was carried out according to previously described methods [42].
Co-immunoprecipitation Experiments
For the attempted co-immunoprecipitation of endogenous spastin and atlastin under non-denaturing conditions, untransfected NSC34 cells were plated into three 15cm plates at 1×107. Twenty four hours later confluent cells were washed in PBS and lysed in 1ml ice-cold NP40 lysis buffer (see [27] for recipe). A 100μl aliquot was then removed for use as a concentrated input fraction. The volume of NSC34 lysate was made up to 10ml and this was split into two fractions. Fraction 1 represented total cell lysate while fraction 2 was centrifuged at 13000g for 5 min and supernatant removed for use as a soluble fraction. Both fractions were then divided further into 2.5ml samples for positive/control co-immunoprecipitations. Anti-atlastin or anti-spastin polyclonal antiserum was then added to + fractions at 1/100, while anti-GFP (ab290 abcam) antibody was added to control fractions. Fractions were then incubated on a rocking table at 4°C for 4 hours. Following antibody incubations 200μl of a 50% slurry of protein-A sepharose beads (Sigma) in NP40 lysis buffer was added to all fractions and incubated overnight on a rocking table at 4°C. Following protein-antibody complex incubations protein-A sepharose beads were isolated by brief centrifugation and then washed 5x for 20 minutes in 10ml NP40 lysis buffer. The final wash supernatant was then removed and beads were resuspended in 2x SDS-PAGE sample buffer. SDS-PAGE was then performed and immunoblotting carried out with appropriate antisera.
For transiently transfected HEK293T cells, cells were plated into two 15cm plates at 1×107 and 24 hours later cells were co-transfected with atlastin-GFP and myc-spastin or pEGFP and myc-spastin. 24 hours post transfection cells were lysed in 1ml ice-cold lysis buffer and a 100μl aliquot removed for use as a concentrated input fraction. Cells were then processed as described for NSC34 cells. An anti-GFP polyclonal antibody (ab290, abcam) was added to + antibody fractions at 1/1000 and +/- antibody fractions were then incubated on a rocking table at 4°C for 4 hours. Further processing was carried out as above.
A modified protocol was used for the co-immunoprecipitations carried out under denaturing conditions. These experiments were performed in NSC34 cells using denaturing conditions in the presence of a chemical cross linker Dithiobis[succinnimidyl propionate] (DSP) (Pierce). NSC34 cells were harvested in PBS containing 1mM DSP and incubated for 30min at room temperature. 1M Tris HCl pH7.5 was added to a final concentration of 10mM to quench DSP activity and cells were incubated for a further 15min at room temperature. Cells were re-suspended in 100μl per 1×107 cells denaturing lysis buffer containing 1% SDS, 50mM Tris HCl pH 7.4, 5mM EDTA, DNAse I (Sigma) and protease inhibitors (Roche). Denatured lysate was then diluted into non-denaturing lysis buffer containing 1% Triton X-100, 50mM Tris HCl pH 7.4, 300mM NaCl, 0.5mM EDTA and protease inhibitors to a final concentration of 0.1% SDS. Total cell lysate was cleared by centrifugation at 13000rpm for 10min and the soluble fraction retained. Protein-A sepharose beads were pre-coupled to 20μg of the anti-atlastin (5409) polyclonal antibody, and to a spurious control antibody (anti-GFP, abcam 6556), in the presence of DSP. DSP activity was quenched with 10mM Tris-HCl, as above. Antibody-coupled protein-A sepharose beads were then added to soluble cell lysates. Antigen-antibody complexes were allowed to form at 4°C for two hours. Beads were then isolated and washed several times in non-denaturing lysis buffer. DSP cross-linking was cleaved by addition of SDS-PAGE sample buffer containing 5% β-mercaptoethanol at 100°C for 5min and proteins were resolved by SDS-PAGE.
GST-fusion Protein Pull-down Experiments
Glutathione sepharose beads (Amersham Biosciences) were washed in a pull down buffer that consisted of 1% NP40, PBS solution with protease inhibitors (Complete Mini protease inhibitor cocktail tablets, Roche Diagnostic) and made up to a 50% slurry in this buffer. HeLa cells were transiently transfected with expression vector constructs of epitope-tagged spastin or atlastin, using the Effectene® Transfection kit (Qiagen), as described above. Cells were harvested 24 to 48 hours after transfection into the wash buffer and freeze/thawed to -20°C.
The pull down reaction consisted of 50ul of 50% glutathione sepharose beads, 500ug cell cytosol and 100ug GST-fusion protein. The total volume was made up to 0.5 mL in a 1.5 mL tube with pull down buffer (1% NP40, PBS, 5mM MgCl2 and 5mM ATP). The GST negative control used an equi-molar amount of GST protein. Tubes were incubated at 4°C on a rotating wheel overnight. The beads were the spun at max speed for 15 seconds in a bench top centrifuge then washed with 1 mL of wash buffer. This cycle was repeated four times. Prior to the fourth wash, the pull down mix was transferred to clean 1.5 mL tubes to eliminate any possible binding of protein to the interior of the plastic tube. Protein bound to the beads was eluted after four wash/spin cycles with 55 uL of SDS sample buffer (188mM TRIS, 6% SDS, 30% glycerol, 0.03% bromophenol blue and 10% beta-mercaptoethanol, pH 6.8). The samples were heated to 95°C for 5 minutes and then spun at maximum speed in a bench top centrifuge for 15 seconds. The supernatants were loaded on a 10% SDS polyacrylamide gel and immunoblotted with anti-myc or anti-GFP primary antibody as appropriate.
Acknowledgements
The pGBDU-C series of vectors and the PJ69-4A MATa and MATα yeast strains were kindly provided by Dr Philip James (Department of Biomolecular Chemistry, University of Wisconsin, Madison, WI 53706-1532, USA.). The S51 spastin antibody was a kind gift of Dr Elena Rugarli (Telethon Institute, Naples, Italy), the atlastin antibody was a kind gift of Craig Blackstone (NIH, Maryland, USA), the GFP-VPS4EQ construct was a kind gift of Paul Whitely (Univeristy of Bath, UK) and NSC34 cells were a kind gift of Dr Neil Cashman (University of Toronto, Canada). This work has been supported by grants from the Wellcome Trust and Medical Research Council. ER is a Wellcome Trust Advanced Clinical Fellow. TLE is supported by a Raymond and Beverly Sackler Fellowship and an award from the Cambridge Commonwealth Trust.
REFERENCES
- 1.Harding AE. The hereditary ataxias and related disorders. Edinburgh: Churchill Livingstone; 1984. [Google Scholar]
- 2.Fink JK. Hereditary spastic paraplegia. In: Rimoin DL, Pyeritz RE, Connor JM, Korf BR, editors. Emory and Rimoin’s Principles and Practice of Medical Genetics. 4th edition London, UK: Harcoart; 2002. pp. 3124–3145. [Google Scholar]
- 3.Reid E. Science in motion: common molecular pathological themes emerge in the hereditary spastic paraplegias. J. Med. Genet. 2003;40:35–39. doi: 10.1136/jmg.40.2.81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Polo JM, Calleja J, Combarros O, Berciano J. Hereditary ataxias and paraplegias in Cantabria, Spain. An epidemiological and clinical study. Brain. 1991;114:855–866. doi: 10.1093/brain/114.2.855. [DOI] [PubMed] [Google Scholar]
- 5.Fink JK. The hereditary spastic paraplegias: nine genes and counting. Arch Neurol. 2003;60:1045–1049. doi: 10.1001/archneur.60.8.1045. [DOI] [PubMed] [Google Scholar]
- 6.Zhao X, Alvarado D, Rainier S, Lemons R, Hedera P, Weber CH, Tukel T, Apak M, Heiman-Patterson T, Ming L, et al. Mutations in a newly identified GTPase cause autosomal dominant hereditary spastic paraparesis. Nature Genet. 2001;29:326–331. doi: 10.1038/ng758. [DOI] [PubMed] [Google Scholar]
- 7.Hazan J, Fonknechten N, Mavel D, Paternotte C, Samson D, Artiguenave F, Davoine CS, Cruaud C, Dürr A, Wincker P, et al. Spastin, a new AAA protein, is altered in the most frequent form of autosomal dominant spastic paraplegia. Nature Genet. 1999;23:296–303. doi: 10.1038/15472. [DOI] [PubMed] [Google Scholar]
- 8.Rainier S, Chai JH, Tokarz D, Nicholls RD, Fink JK. NIPA1 gene mutations cause autosomal dominant hereditary spastic paraplegia (SPG6) Am J. Hum. Genet. 2003;73:967–971. doi: 10.1086/378817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Reid E, Kloos M, Ashley-Koch A, Hughes L, Bevan S, Svenson IK, Lennon Graham F, Gaskell PC, Dearlove A, Pericak-Vance MA, et al. A Kinesin Heavy Chain (KIF5A) Mutation in Hereditary Spastic Paraplegia (SPG10) Am. J. Hum. Genet. 2002;71:1189–1194. doi: 10.1086/344210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hansen JJ, Durr A, CournuRebeix I, Georgopoulos C, Ang D, Nielsen MN, Davoine CS, Brice A, Fontaine B, Gregersen N, Bross P. Hereditary spastic paraplegia SPG13 is associated with a mutation in the gene encoding the mitochondrial chaperonin Hsp60. Am. J. Hum. Genet. 2002;70:1328–1332. doi: 10.1086/339935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fonknechten N, Mavel D, Byrne P, Davoine CS, Cruaud C, Boentsch D, Samson D, Coutinho P, Hutchinson M, McMonagle P, et al. Spectrum of SPG4 mutations in autosomal dominant spastic paraplegia. Hum. Mol. Genet. 2000;9:637–644. doi: 10.1093/hmg/9.4.637. [DOI] [PubMed] [Google Scholar]
- 12.Meijer IA, Hand CK, Cossette P, Figlewicz DA, Rouleau GA. Spectrum of SPG4 mutations in a large collection of North American families with hereditary spastic paraplegia. Arch. Neurol. 2002;59:281–286. doi: 10.1001/archneur.59.2.281. [DOI] [PubMed] [Google Scholar]
- 13.Sauter S, Miterski B, Klimpe S, Bonsch D, Schols L, Visbeck A, Papke T, Hopf HC, Engel W, Deufel T, et al. Mutation analysis of the spastin gene (SPG4) in patients in Germany with autosomal dominant hereditary spastic paraplegia. Hum. Mutat. 2002;20:127–132. doi: 10.1002/humu.10105. [DOI] [PubMed] [Google Scholar]
- 14.Charvin D, Cifuentes-Diaz C, Fonknechten N, Joshi V, Hazan J, Melki J, Betuing S. Mutations of SPG4 are responsible for a loss of function of spastin, an abundant neuronal protein localized in the nucleus. Hum. Mol. Genet. 2003;12:71–78. doi: 10.1093/hmg/ddg004. [DOI] [PubMed] [Google Scholar]
- 15.Wharton SB, McDermott CJ, Grierson AJ, Wood JD, Gelsthorpe C, Ince PG, Shaw PJ. The cellular and molecular pathology of the motor system in hereditary spastic paraparesis due to mutation of the spastin gene. J. Neuropathol. Exp. Neurol. 2003;62:1166–1177. doi: 10.1093/jnen/62.11.1166. [DOI] [PubMed] [Google Scholar]
- 16.Beetz C, Brodhun M, Moutzouris K, Kiehntopf M, Berndt A, Lehnert D, Deufel T, Bastmeyer M, Schickel J. Identification of nuclear localisation sequences in spastin (SPG4) using a novel Tetra-GFP reporter system. Biochem. Biophys. Res. Commun. 2004;318:1079–1084. doi: 10.1016/j.bbrc.2004.03.195. [DOI] [PubMed] [Google Scholar]
- 17.Errico A, Claudiani P, D’Addio M, Rugarli EI. Spastin interacts with the centrosomal protein NA14, and is enriched in the spindle pole, the midbody, and the distal axon. Hum. Mol. Genet. 2004;13:2121–2132. doi: 10.1093/hmg/ddh223. [DOI] [PubMed] [Google Scholar]
- 18.Claudiani P, Riano E, Errico A, Andolfi G, Rugarli EI. Spastin subcellular localization is regulated through usage of different translation start sites and active export from the nucleus. Exp. Cell Res. 2005;309:358–369. doi: 10.1016/j.yexcr.2005.06.009. [DOI] [PubMed] [Google Scholar]
- 19.Errico A, Ballabio A, Rugarli EI. Spastin, the protein mutated in autosomal dominant hereditary spastic paraplegia, is involved in microtubule dynamics. Hum. Mol. Genet. 2002;11:153–163. doi: 10.1093/hmg/11.2.153. [DOI] [PubMed] [Google Scholar]
- 20.Evans KJ, Gomes ER, Reisenweber SM, Gundersen GG, Lauring BP. Linking axonal degeneration to microtubule remodeling by Spastin-mediated microtubule severing. J Cell Biol. 2005;168:599–606. doi: 10.1083/jcb.200409058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.McDermott CJ, Grierson AJ, Wood JD, Bingley M, Wharton SB, Bushby KM, Shaw PJ. Hereditary spastic paraparesis: disrupted intracellular transport associated with spastin mutation. Ann. Neurol. 2003;54:748–759. doi: 10.1002/ana.10757. [DOI] [PubMed] [Google Scholar]
- 22.Trotta N, Orso G, Rossetto MG, Daga A, Broadie K. The hereditary spastic paraplegia gene, spastin, regulates microtubule stability to modulate synaptic structure and function. Curr. Biol. 2004;14:1135–1147. doi: 10.1016/j.cub.2004.06.058. [DOI] [PubMed] [Google Scholar]
- 23.Sherwood NT, Sun Q, Xue M, Zhang B, Zinn K. Drosophila spastin regulates synaptic microtubule networks and is required for normal motor function. PLoS Biol. 2004;2:e429. doi: 10.1371/journal.pbio.0020429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Roll-Mecak A, Vale RD. The Drosophila homologue of the hereditary spastic paraplegia protein, spastin, severs and disassembles microtubules. Curr Biol. 2005;15:650–655. doi: 10.1016/j.cub.2005.02.029. [DOI] [PubMed] [Google Scholar]
- 25.Patel H, Cross H, Proukakis C, Hershberger R, Bork P, Ciccarelli FD, Patton MA, McKusick VA, Crosby AH. SPG20 is mutated in Troyer syndrome, an hereditary spastic paraplegia. Nature Genet. 2002;31:347–348. doi: 10.1038/ng937. [DOI] [PubMed] [Google Scholar]
- 26.Ciccarelli FD, Proukakis C, Patel H, Cross H, Azam S, Patton MA, Bork P, Crosby AH. The identification of a conserved domain in both spartin and spastin, mutated in hereditary spastic paraplegia. Genomics. 2003;81:437–441. doi: 10.1016/s0888-7543(03)00011-9. [DOI] [PubMed] [Google Scholar]
- 27.Reid E, Connell J, Edwards TL, Duley S, Brown SE, Sanderson CM. The hereditary spastic paraplegia protein spastin interacts with the ESCRT-III complex-associated endosomal protein CHMP1B. Hum. Mol. Genet. 2005;14:19–38. doi: 10.1093/hmg/ddi003. [DOI] [PubMed] [Google Scholar]
- 28.Muglia M, Magariello A, Nicoletti G, Patitucci A, Gabriele AL, Conforti FL, Mazzei R, Caracciolo M, Ardito B, Lastilla M, et al. Further evidence that SPG3A gene mutations cause autosomal dominant hereditary spastic paraplegia. Ann. Neurol. 2002;51:794–795. doi: 10.1002/ana.10185. [DOI] [PubMed] [Google Scholar]
- 29.Tessa A, Casali C, Damiano M, Bruno C, Fortini D, Patrono C, Cricchi F, Valoppi M, Nappi G, Amabile GA, et al. SPG3A: An additional family carrying a new atlastin mutation. Neurology. 2002;59:2002–2005. doi: 10.1212/01.wnl.0000036902.21438.98. [DOI] [PubMed] [Google Scholar]
- 30.Dalpozzo F, Rossetto MG, Boaretto F, Sartori E, Mostacciuolo ML, Daga A, Bassi MT, Martinuzzi A. Infancy onset hereditary spastic paraplegia associated with a novel atlastin mutation. Neurology. 2003;61:580–581. doi: 10.1212/01.wnl.0000078189.73611.df. [DOI] [PubMed] [Google Scholar]
- 31.Wilkinson PA, Hart PE, Patel H, Warner TT, Crosby AH. SPG3A mutation screening in English families with early onset autosomal dominant hereditary spastic paraplegia. J Neurol Sci. 2003;216:43–45. doi: 10.1016/s0022-510x(03)00210-7. [DOI] [PubMed] [Google Scholar]
- 32.Sauter SM, Engel W, Neumann LM, Kunze J, Neesen J. Novel mutations in the Atlastin gene (SPG3A) in families with autosomal dominant hereditary spastic paraplegia and evidence for late onset forms of HSP linked to the SPG3A locus. Hum Mutat. 2004;23:98. doi: 10.1002/humu.9205. [DOI] [PubMed] [Google Scholar]
- 33.D’Amico A, Tessa A, Sabino A, Bertini E, Santorelli FM, Servidei S. Incomplete penetrance in an SPG3A-linked family with a new mutation in the atlastin gene. Neurology. 2004;62:2138–2139. doi: 10.1212/01.wnl.0000127698.88895.85. [DOI] [PubMed] [Google Scholar]
- 34.Hedera P, Fenichel GM, Blair M, Haines JL. Novel Mutation in the SPG3A Gene in an African American Family With an Early Onset of Hereditary Spastic Paraplegia. Arch. Neurol. 2004;61:1600–1603. doi: 10.1001/archneur.61.10.1600. [DOI] [PubMed] [Google Scholar]
- 35.Zhu PP, Patterson A, Lavoie B, Stadler J, Shoeb M, Patel R, Blackstone C. Cellular Localization, Oligomerization, and Membrane Association of the Hereditary Spastic Paraplegia 3A (SPG3A) Protein Atlastin. J. Biol. Chem. 2003;278:49063–49071. doi: 10.1074/jbc.M306702200. [DOI] [PubMed] [Google Scholar]
- 36.Cashman NR, Durham HD, Blusztajn JK, Oda K, Tabira T, Shaw IT, Dahrouge S, Antel JP. Neuroblastoma x spinal cord (NSC) hybrid cell lines resemble developing motor neurons. Dev. Dyn. 2002;194:209–221. doi: 10.1002/aja.1001940306. [DOI] [PubMed] [Google Scholar]
- 37.Whitley P, Reaves BJ, Hashimoto M, Riley AM, Potter BV, Holman GD. Identification of mammalian Vps24p as an effector of phosphatidylinositol 3,5-bisphosphate-dependent endosome compartmentalization. J. Biol. Chem. 2003;278:38786–38795. doi: 10.1074/jbc.M306864200. [DOI] [PubMed] [Google Scholar]
- 38.Dreveny I, Pye VE, Beuron F, Briggs LC, Isaacson RL, Matthews SJ, McKeown C, Yuan X, Zhang X, Freemont PS. p97 and close encounters of every kind: a brief review. Biochem. Soc. Trans. 2004;32:715–720. doi: 10.1042/BST0320715. [DOI] [PubMed] [Google Scholar]
- 39.Whiteheart SW, Matveeva EA. Multiple binding proteins suggest diverse functions for the N-ethylmaleimide sensitive factor. J. Struct. Biol. 2004;146:32–43. doi: 10.1016/j.jsb.2003.09.015. [DOI] [PubMed] [Google Scholar]
- 40.Horton RM, Hunt HD, Ho SN, Pullen JK, Pease LR. Engineering hybrid genes without the use of restriction enzymes: gene splicing by overlap extension. Gene. 1989;77:61–68. doi: 10.1016/0378-1119(89)90359-4. [DOI] [PubMed] [Google Scholar]
- 41.Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J. Mol. Biol. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- 42.Bright NA, Reaves BJ, Mullock BM, Luzio JP. Dense core lysosomes can fuse with late endosomes and are re-formed from the resultant hybrid organelles. J. Cell Sci. 1997;110:2027–2040. doi: 10.1242/jcs.110.17.2027. [DOI] [PubMed] [Google Scholar]