

Abstract
Adult Xenopus laevis frogs made transgenic by restriction enzyme-mediated integration were bred to test the feasibility of establishing lines of frogs that express transgenes. All of the 19 animals raised to sexual maturity generated progeny that expressed the transgene(s). The patterns and levels of expression of green fluorescent protein transgenes driven by a viral promoter, rat promoter, and four X. laevis promoters were all unaffected by passage through the germ line. These results demonstrate the ease of establishing transgenic lines in X. laevis.
Xenopus laevis is a model system for many types of biological questions; however, studies in this organism have been hampered by the lack of genetic tools. Recently, an efficient method for generating transgenic X. laevis has been described (1). The method involves restriction enzyme-mediated integration of DNA into demembranated sperm nuclei and transplantation of the nuclei into unfertilized eggs. This technology has been employed to study regulatory regions of genes (2, 3) and assess the effects of overexpression of wild-type and dominant-negative forms of gene products (1, 4). To date, these studies have been conducted only in the first-generation (F0) animals produced by nuclear transplantation. One drawback is that embryonic phenotypes have been difficult to study in F0 transgenic animals because of the high prevalence of abnormalities caused by the restriction enzyme-mediated integration procedure. Also, different integration events can result in highly variable levels of expression in different F0 animals. Such variability of expression can often complicate the interpretation of results. For many types of experiments, it would be ideal to have large numbers of normal developing animals with equal levels of transgene expression.
This study was designed to address the feasibility of establishing transgenic lines of X. laevis. We show here that every transgenic animal that was raised to sexual maturity produced progeny that faithfully expressed the transgene(s). These results demonstrate that lines of X. laevis expressing various transgenes can be readily generated.
Materials and Methods
Preparation, Rearing, and Breeding of Transgenic Animals.
F0 transgenic animals were prepared by using the restriction enzyme-mediated integration method (5) with minor modifications (4). F0 animals expressing the transgenes were identified by their green fluorescent protein (GFP) fluorescence at 2–7 days of development. Tadpoles were raised under standardized conditions (4). In the heat-shock experiments, 8-day-old tadpoles were transferred to 0.1× MMR (10 mM NaCl/0.2 mM KCl/0.1 mM MgCl2/0.2 mM CaCl2/0.5 mM Hepes, pH 7.5) solution that had been prewarmed to 33°C and then held at this temperature for 1 h before being returned to a solution at room temperature. Females to be bred were primed with 50 units of pregnant mare serum gonadotropin (Sigma) the day before mating and 300–350 units of human chorionic gonadotropin (Sigma) just before mating. The males received 100 units of human chorionic gonadotropin just before mating. Matings were conducted by placing a transgenic frog with a wild-type frog of the opposite sex into a bucket and harvesting the eggs the next morning.
Plasmids.
All constructs were made in pCS2+ vectors (6). The construct carrying the simian cytomegalovirus (CMV) promoter/enhancer (pCS2+GFP*) has been described (4). The construct with the X. laevis γ-crystallin promoter driving GFP (Cry1GFP3) was a gift of Robert Grainger (University of Virginia, Charlottesville) and is here designated xγCrys:GFP. The construct with the X. laevis neural β-tubulin promoter driving the fusion construct of the tau protein and GFP (NβTτGFP) was the gift of Enrique Amaya (Wellcome/CRC Institute, Cambridge, U.K.). Various other constructs were made by amplifying cDNAs or genomic DNAs by PCR using Pfu DNA polymerase (Stratagene). The GFP-luciferase fusion construct (pCS2+GFP-luc) was made by amplifying CMV:GFP from pCS2+GFP* with primers 5′-ATGGACGGGCCCTCTTCGCTATTACGCCAGTCG-3′ and 5′-CTATTTGTAGAGCTCATCCATGCCATGTGTAATCC-3′ and amplifying luciferase from the pGL2-basic plasmid (Promega) with primers 5′-GGATGAGCTCTACAAAATGGAAGACGCCAAAAACATAAAGAAAGG-3′ and 5′-ATGGACGCGGCCGCATGATAAGATACATTGATGAGTTTGG-3′. Both fragments were digested with SacI and ligated. The ligation product was then amplified by using the GFP internal primer 5′-AAGGTGATGCAACATACGG-3′ and the same reverse luciferase primer. This fragment was cloned into the NcoI and NotI sites of pCS2+GFP*. pCS2+GFPtag, a GFP vector designed for C-terminal GFP fusion constructs, was made by amplifying GFP carrying a point mutation (Ser-65 → Cys) by using primers 5′-GATCCCATCGATCCACCATGAGTAAAGG-3′ and 5′-GAGGAATTCTATTTGTATAGTTCATCCATGCC-3′ and cloning the fragment into the EcoRI and XbaI sites of pCS2+. A construct carrying the herpes simplex virus thymidine kinase (HSVtk) promoter (pCS2+[HSVtk]GFPtag) was used in this study as a shuttle cloning vector and was made by amplifying the HSVtk promoter from the pRLtkluc vector (Promega) by using primers 5′-ACGCCAGTCGACAGATCTAAATGAGTCTTCGGACCTC-3′ and 5′-GACTGTTGTCAGAAGAATCAAGC-3′ and cloning the fragment into the SalI and HindIII sites of pCS2+GFPtag. The construct carrying the rat tubulin α-1 promoter (pCS2+[rTα1]GFPtag) was made by digesting the plasmid pBluescript SK T alpha1GHPh (7), which was kindly provided by Steven Goldman (Cornell University Medical Center, New York), with HindIII and XbaI. The ends were blunted with the Klenow fragment of DNA polymerase and the fragment containing the promoter was cloned into blunted BglII and MluI sites of pCS2+[HSVtk]GFPtag. The construct carrying the X. laevis hsp70 promoter (pCS2+[Hsp70]GFP-luc) was made by amplifying 578 bp of upstream sequences from the hsp70 gene (8) from genomic DNA by using the primers 5′-ACGCCAGTCGACCCGTTTAGCAGGAAATAGCCTTGG-3′ and 5′-GACTGTAAGCTTGCGCTCCTTACAGTTTGCTTTTCG-3′ and cloning the fragment into the SalI and HindIII sites of pCS2+GFP-luc. The construct carrying the X. laevis larval keratin promoter (pCS2+[Ker]GFP) was made by amplifying 922 bp of upstream sequences from the larval keratin gene (9) from genomic DNA by using the primers 5′-ACGTGTCGACGCAACATACAAAGAGTAACGG-3′ and 5′-CCCAAGCTTGCCTGCAGCTTTGACAGCC-3′ and cloning the fragment into the SalI and HindIII sites of pCS2+GFP(S65C).
PCR Typing, GFP Detection, and Luciferase Measurements.
PCR typing of transgenic animals was performed as described (4) by using the GFP-specific primers 5′-AAGGTGATGCAACATACGG-3′ and 5′-ACAGGGCCATCGCCAATTG-3′, the growth hormone (GH) cDNA-specific primers 5′-CGTGCTCGAGTTAAATGGTGCAGTTGCTTTC-3′ and 5′-GCTAGAATTCACAGCCACCATGGCTACAGGGTTCTGCTC-3′, and the gene B cDNA-specific (10) primers 5′-CCAGAGATCTGCATGGCTCA-3′ and 5′-TTCCCAGCAACAGCCCGAAGTGG-3′. GFP fluorescence in whole tadpoles was monitored with a MZ12 fluorescence dissecting microscope and a LEI-750 charge-coupled device video camera system (Leica). GFP expression in cryosections was detected by means of a rabbit anti-GFP polyclonal antibody (CLONTECH) and a Cy3-conjugated anti-rabbit secondary antibody (Jackson ImmunoResearch), and was photographed with a RTE-CCD-1317-K11 digital camera (Princeton Instruments) and iplab spectrum (Signal Analytics Corporation, Vienna, VA) software. For luciferase measurements, animals were anesthetized with 0.01% 3-aminobenzoic acid ethyl ester, placed in tubes, and frozen in liquid nitrogen. Luciferase activities were assayed as described (11) by using a Monolight 2010 luminometer (Analytical Luminescence Laboratory, San Diego).
Results
Nineteen F0 founder animals carrying GFP transgenes driven by various promoters were selected as embryos because of their strong fluorescence and raised to sexual maturity. Their transgenic status was confirmed by PCR of tadpole tail tissue. Among the animals raised were ones carrying GFP transgenes driven by the simian CMV immediate early promoter (CMV), the rat tubulin α-1 promoter (rTα1), and various X. laevis promoters: neural β-tubulin (xNβT), γ-crystallin (xγCrys), larval keratin (xKer), and hsp70 (xHsp70) promoters. The transgenic frogs were crossed with wild-type frogs, and the progeny of F0 (F1) were screened by fluorescence during the first week of development. The GFP-positive and -negative progeny were counted to estimate the number of integration events in the founders (Table 1). Expression of transgenes ranged from 50% to over 98% of F1 animals; thus, the number of integration events in different founders likely ranged from one to greater than four.
Table 1.
Transgenic lines
| Line | F0
|
F1
|
|||
|---|---|---|---|---|---|
| Promoter | Reporter | Sex | GFP+/total | % GFP+ | |
| 3 | CMV | GFP | F | 201/269 | 75 |
| 7 | CMV | GFP | F | 213/273 | 78 |
| 8 | CMV | GFP | F | 32/48 | 67 |
| 10 | CMV | GFP | F | 182/204 | 89 |
| 12 | CMV | GFP | F | 256/260 | 98 |
| 14 | CMV | GFP | M | 186/339 | 55 |
| 15* | CMV | GFP | M | 311/323 | 96 |
| 16 | CMV | GFP | M | 95/169 | 56 |
| 17 | CMV | GFP | M | 313/351 | 89 |
| 18 | CMV | GFP | M | 221/287 | 77 |
| 1 | CMV | GFP | M | 252/504 | 50 |
| CMV | xGH | ||||
| 9 | rTα1 | GFP | F | 91/92 | 99 |
| 13 | rTα1 | GFP | F | 278/398 | 70 |
| 2 | xNβT | TauGFP | M | 202/272 | 74 |
| 6 | xNβT | TauGFP | F | 210/432 | 49 |
| 11 | xNβT | TauGFP | M | 210/279 | 75 |
| 19 | xKer | GFP | M | 126/251 | 50 |
| 4* | xγCrys | GFP | F | 187/252 | 74 |
| CMV | GeneB | ||||
| 5 | xγCrys | GFP | F | 281/540 | 52 |
| xHsp70 | GFP-luc | ||||
*Deceased.
In animals prepared by the current transgenesis procedure, more than one DNA molecule usually integrates at each chromosomal site (1). Such a pattern of integration agrees with the high incidence of coexpression seen in F0 animals when different constructs are used together (4). The pattern of transgene inheritance that we observed also suggests that multiple DNA molecules usually integrate together at each chromosomal site. Three of the F0 animals (Table 1; lines 1, 4, and 5) were prepared by using two different cDNA constructs. In two cases, the two transgenes cosegregated. In line 1, which has both GFP and GH (xGH) driven by CMV promoters, all animals selected by their fluorescence tested positive by PCR for both transgenes (10/10), and all animals selected by their lack of fluorescence tested negative by PCR for both transgenes (10/10). In addition, every animal that was fluorescent also developed the characteristic phenotype of GH overexpression (see below). In line 5, which has GFP driven by the xγCrys promoter and a fusion construct of GFP and luciferase (GFP-luc) driven by the xHsp70 promoter, all animals selected because of their fluorescent eyes also had luciferase activity (20/20), and all animals selected because of their lack of eye fluorescence also lacked luciferase activity (20/20). Thus, in both lines 1 and 5, the two transgenes must have integrated very near to each other and most likely integrated together into the same chromosomal locus. In contrast, in line 4, which has GFP driven by the xγCrys promoter and a thyroid hormone-inducible gene of unknown function (gene B) driven by the CMV promoter, the pattern of inheritance was consistent with the founder having both a site where the two different transgenes integrated and a second site where only the gene B transgene (or transgenes) integrated. All 10 of the animals that were selected because of their fluorescent eyes tested positive by PCR with GFP and gene B-specific primers. However, the 10 animals that were selected because of their lack of eye fluorescence all tested negative by PCR with GFP primers, but two tested positive with gene B primers.
To determine the frequency of integration events that did not express the transgenes, animals were separated into fluorescent and nonfluorescent groups, and individuals were screened by PCR with GFP-specific primers. All fluorescent animals (88 F1 animals derived from 15 different founders) tested positive by PCR, and all nonfluorescent animals (144 F1 animals derived from 15 different founders) tested negative by PCR; thus, there was no evidence for an integration event that did not express the transgene.
The pattern of fluorescence for the individual founder animals during tadpole stages was not recorded. However, the pattern and levels of fluorescence observed in CMV:GFP transgenic tadpoles in the F1 generation (Fig. 1 B–L) were comparable to those observed in typical F0 animals (Fig. 1 M–O). All siblings derived from F0 parents in which the pattern of inheritance suggested a single integration event (Table 1, lines 1, 14, 16, and 19) had similar levels of fluorescence, whereas siblings from lines with multiple integration events (lines 3, 7, 10, 12, 15, 17, and 18) had variable levels of fluorescence. Indeed, 30 of 64 animals from line 17 that were screened initially as lacking GFP fluorescence were found on rescreening to have GFP expression that was so low that it was detectable only in the olfactory epithelia (Fig. 1K), which is the tissue where the CMV promoter drives the strongest expression. Similar low levels of expression were also observed in some F0 tadpoles (Fig. 1O).
Figure 1.

CMV:GFP transgenic tadpoles. (A) Control nontransgenic tadpole. (B–L) F1 CMV:GFP tadpoles from 10 different lines. Numbers in the right bottom corner represent the line number in Table 1. (K) Weak-expressing tadpole from line 17. (J) A strong-expressing tadpole from line 17. (M–O) Three F0 CMV:GFP tadpoles. All panels represent dorsal views of 8-day-old tadpoles taken at the same 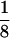 -s exposure, except for K, which was taken at ¼-s exposure. (Scale bar represents 500 μm.)
-s exposure, except for K, which was taken at ¼-s exposure. (Scale bar represents 500 μm.)
The tissue specificity of expression of the various promoters was maintained in the F1 progeny. The X. laevis neural β-tubulin (Fig. 2A) and γ-crystallin (Fig. 2B) promoters drove expression in the brain and lens, respectively, starting during embryogenesis and lasting through metamorphosis. The X. laevis larval keratin promoter drove expression in the skin (Fig. 2C) and epithelial linings of the buccal cavity (data not shown) during larval life. A rat promoter for tubulin α-one showed expression in the nervous system (Fig. 2D), and later also in the growing limbs (Fig. 2E), an expression pattern that is identical to that seen in F0 animals (data not shown). All four tissue-specific promoters drove expression in the F1 generation at levels comparable to those typically seen in the F0 generation (data not shown).
Figure 2.

F1 tadpoles transgenic for tissue-specific promoters. (A) xNβT:TauGFP line 2, dorsal view of an 8-day-old tadpole. (B) xγCryst:GFP line 5, dorsal view of an 8-day-old tadpole. The arrowhead points to the fluorescent lens. The lens of the other eye was obscured because of the orientation of the tadpole. (C) xKer:GFP line 19, immunohistochemical detection of GFP (red), with 4′,6-diamidino-2-phenylindole counterstain (blue), in transverse section through the head of a stage 42 (12) tadpole. The arrowhead points to the skin and the arrow points to the brain. (D) rTα1:GFP line 13, dorsal view of an 8-day-old animal. (E) rTα1:GFP line 13, lateral view of a stage 54 tadpole. The arrowhead points to the forelimb and the arrow points to the hindlimb. [Scale bars represent 500 μm (A, B, and D), 20 μm (C), and 1 mm (E).]
A study of the variability of expression between different integration events and in F1 relative to F0 animals was performed by using line 5, which carries xHsp70:GFP-luc and xγCrys:GFP transgenes integrated at a single locus. Different F1 transgenic tadpoles, which were identified by their fluorescent lenses, had very similar levels of expression of the xHsp70:GFP-luc transgene at various times after heat shock (Fig. 3); however, F0 tadpoles showed a much greater variability of expression. The founder of line 5 was chosen because it had fluorescent lenses and therefore was presumed to carry the xHsp70:GFP-luc transgene. It was not selected because of the expression level of the xHSP70:GFP-luc transgene. The progeny of this animal had luciferase levels that were low but within the range that is observed in F0 animals.
Figure 3.

Luciferase activity of xHSP70:GFP-luc transgenes in 18 F0 animals 8 h after heat shock (○) and 5 F1 transgenic animals (line 5) assayed at each time point after heat shock (●).
For line 1, which carries GFP and X. laevis GH (xGH) both under the control of the CMV promoter, F2 animals were generated and compared with the F1 animals. The founder (F0) for this line was an 8-month-old moribund male that showed severe gigantism. Its testes were used to fertilize wild-type eggs. Half of the resultant progeny carried both transgenes, whereas the other half carried neither (see above), which is suggestive of a single integration event of both transgenes together. Transgenic F1 animals displayed hyperplastic brachial arches as young tadpoles (Fig. 4A), increased size and weight as tadpoles and frogs, and skeletal deformities as adults; the typical phenotype seen in F0 animals carrying CMV:xGH transgenes (H.H. and D.D.B., unpublished results). Siblings that did not carry the transgenes did not develop this phenotype (Fig. 4B). Two different transgenic F1 males were killed and their testes were used to fertilize wild-type eggs. Half of the F2 progeny from each cross (300 of 612 and 95 of 200) expressed the GFP transgene (Fig. 4C) at levels comparable to those seen in F1 animals (Fig. 4A). All of the F2 animals expressing GFP also developed the brachial arch hyperplasia associated with CMV:xGH transgene expression (Fig. 4C), whereas those that did not express GFP had normal brachial arches (Fig. 4D).
Figure 4.

Comparison of 2-week-old F1 and F2 tadpoles from line 1. Two-week-old tadpoles that expressed GFP in the F1 generation (A) and F2 generation (C) were larger and showed hyperplastic brachial arches, whereas animals that did not express GFP in the F1 generation (B) and F2 generation (D) were smaller and had normal brachial arches. Each panel shows the same animal twice: in a reflected light view on top, and a GFP fluorescence view on the bottom. Brackets highlight the enlarged brachial arches. (Scale bar represents 1 mm.)
Discussion
The development of transgenic methodology for X. laevis (1) greatly expands the usefulness of this model organism. Many studies can be performed in the original F0 animals derived by nuclear transplantation. F0 transgenics in X. laevis are ideal to study gene regulatory regions (2, 3) because large numbers of transgenic animals can be easily and inexpensively generated, and comparing individuals with different integration events in some cases can strengthen rather than weaken the analysis. Analyses of gene function are also possible by using F0 transgenics in X. laevis (1, 4); however, because of differing levels of transgene expression in different animals, gene function studies require the use of strong promoters that give reproducible patterns and levels of expression and the analysis of many individuals to ascertain phenotypes.
Even strong promoters such as CMV show differences in expression depending on the integration event (Fig. 1). In transgenic lines where the pattern of inheritance suggests a single integration event, all transgenic F1 animals have similar levels of expression. For example, in line 5, which has the xHsp70 promoter driving GFP-luc and a pattern of inheritance that suggests a single integration event (Table 1), different F1 transgenic animals have very similar levels of expression at various times after heat shock (Fig. 3). In contrast, the variability of expression for this construct is much greater in F0 tadpoles that carry different integration events. As expected, in transgenic lines in which the pattern of inheritance suggested multiple integration events, different levels of expression were apparent among the F1 animals.
Having multiple animals with similar levels of expression will be useful in assessing phenotypes associated with the overexpression of gene products. We show here that both F1 and F2 animals that carry GH transgenes acquire the enhanced growth and hyperplastic brachial arch phenotypes that result from overexpression of this hormone. Although the effect of GH is variable in F0 animals, with some animals grossly affected and others unaffected, the F1 and F2 animals that carry the same integration event have a consistently severe phenotype (H.H. and D.D.B., unpublished results).
The patterns of inheritance (Table 1) suggest that in both lines 1 and 5 a single integration event occurred in which two different cDNAs integrated into the same chromosomal site. In these lines, both pairs of transgenes were expressed after passage through the germ line. Every animal in line 1 that expressed GFP also developed the characteristic GH overexpression phenotype. Every animal in line 5 that had fluorescent lenses could be induced to express the Hsp70:GFPluc transgene. This finding demonstrates that multiple transgenes integrated into the same chromosomal site are coexpressed. Such coexpression of a marker transgene and a test transgene, especially in lines that carry a single integration event, will be particularly useful for many applications.
All of the transgenic founders produced F1 progeny that expressed the transgenes, and in the one case tested, this expression was unaltered by a second passage through the germ line. We have found no evidence for integrations that were silenced by passage through the germ line. Even though the F0 animals that were raised were preselected because of their strong expression of the transgenes, many lines had multiple integration events that would not have been the basis for selection in the F0 animals. In no case did we find an F1 animal that lacked transgene expression but had the transgene DNA. We did find examples of low expression in F1 animals; however, such low expression also occurs in F0 animals.
This study shows that transgenic lines of X. laevis can be made reliably. All of the transgenic F0s raised to maturity were successfully mated. The youngest frogs naturally mated in this study were 11-month-old females. Males can be crossed before this age either by using macerated testes to fertilize eggs in vitro, as we did here for an 8-month-old frog, or by injecting sperm nuclei preparations into unfertilized eggs, as has been done for males as young as 5 months old (N.M.-A., unpublished work). All of the adults produced progeny that expressed the transgenes faithfully. The ability to make stable lines of X. laevis expressing different transgenes expands the use of the current transgenesis methodology and the usefulness of X. laevis as a model organism.
Acknowledgments
We thank Enrique Amaya, Marnie Halpern, and members of the Brown laboratory for critical comments on the manuscript. This work was supported by a Jane Coffin Childs Fund for Medical Research postdoctoral fellowship to N.M.-A. and grants from the National Institutes of Health and the G. Harold and Leila Y. Mathers Charitable Trust to D.D.B. The research performed by H.H. and D.L.B. was in partial fulfillment of the requirements for the Ph.D. degree at John Hopkins University.
Abbreviations
- GFP
green fluorescent protein
- CMV
cytomegalovirus
- GH
growth hormone
- F0
first generation
- F1
progeny of F0
- F2
progeny of F1
Footnotes
See commentary on page 14189.
References
- 1.Kroll K L, Amaya E. Development (Cambridge, UK) 1996;122:3173–3183. doi: 10.1242/dev.122.10.3173. [DOI] [PubMed] [Google Scholar]
- 2.Knox B E, Schlueter C, Sanger B M, Green C B, Besharse J C. FEBS Lett. 1998;423:117–121. doi: 10.1016/s0014-5793(98)00018-0. [DOI] [PubMed] [Google Scholar]
- 3.Casey E S, Tada M, Fairclough L, Wylie C C, Heasman J, Smith J C. Development (Cambridge, UK) 1999;126:4193–4200. doi: 10.1242/dev.126.19.4193. [DOI] [PubMed] [Google Scholar]
- 4.Huang H, Marsh-Armstrong N, Brown D D. Proc Natl Acad Sci USA. 1999;96:962–967. doi: 10.1073/pnas.96.3.962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Amaya E, Kroll K L. In: Methods in Molecular Biology. Sharp P, Mason I, editors. Totowa, NJ: Humana; 1999. pp. 393–414. [DOI] [PubMed] [Google Scholar]
- 6.Turner D L, Weintraub H. Genes Dev. 1994;8:1434–1447. doi: 10.1101/gad.8.12.1434. [DOI] [PubMed] [Google Scholar]
- 7.Gloster A, Wu W, Speelman A, Weiss S, Causing C, Pozniak C, Reynolds B, Chang E, Toma J G, Miller F D. J Neurosci. 1994;14:7319–7330. doi: 10.1523/JNEUROSCI.14-12-07319.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bienz M. EMBO J. 1984;3:2477–2483. doi: 10.1002/j.1460-2075.1984.tb02159.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Miyatani S, Winkles J, Sargent T, Dawid I D. J Cell Biol. 1986;103:1957–1965. doi: 10.1083/jcb.103.5.1957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wang Z, Brown D D. J Biol Chem. 1993;268:16270–16278. [PubMed] [Google Scholar]
- 11.Brasier A R, Tate J E, Habener J F. BioTechniques. 1989;7:1116–1122. [PubMed] [Google Scholar]
- 12.Nieuwkoop P D, Faber J. Normal Table of Xenopus laevis (Daudin) Amsterdam: Elsevier Biomedical; 1956. pp. 1–252. [Google Scholar]