

Diverse mechanisms for transcription antitermination are widespread (24, 47). Recently, a new type, in which the ribosome and the TnaC leader peptide control Rho-dependent transcription termination upstream of the tnaAB operon, was characterized for Escherichia coli (Fig. 1) (1, 34, 49). Remarkably, the nascent leader peptide directs formation of a stereospecific tryptophan-binding site near the peptidyl transferase center. Bound tryptophan inhibits peptidyl transferase activity, preventing release factor cleavage of TnaC-peptidyl-tRNAPro (TnaC-tRNAPro). The stalled ribosome blocks the RNA entry site for Rho factor, allowing transcription readthrough. In this issue, Luis Rogelio Cruz-Vera and Charles Yanofsky show that conserved TnaC residues Asp-16 and Pro-24 are required for tryptophan inhibition of ribosome release (9). This paper exhibits the in vitro methods developed in the Yanofsky laboratory for studying nascent peptide control of ribosome function.
FIG. 1.

tnaAB operon control region, drawn to scale. The transcription initiation site (11) is indicated as +1. The 319-nt transcribed leader region is denoted as tnaL (44). Coding regions for the leader peptide (tnaC) and for the amino-terminal portion of tryptophanase (tnaA) are shown as open boxes (truncated for tnaA). DNA sites for binding cyclic AMP (cAMP) receptor protein (CRP) and RNA polymerase (RNAP) are depicted as filled boxes (11). The boxA-rut element (16, 44) in RNA serves as an entry point for Rho factor, and sites for Rho-dependent transcription termination (43) within the leader region are indicated as vertical arrows. The nascent TnaC leader peptide causes conformational shifts within the ribosome to form a tryptophan-binding site. Bound tryptophan inhibits peptidyl transferase activity, preventing release of 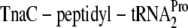 and thereby stalling the ribosome at the tnaC termination codon. Rho factor, blocked by the ribosome from the boxA-rut element, is unable to load onto the RNA to catalyze transcription termination. In the absence of tryptophan,
and thereby stalling the ribosome at the tnaC termination codon. Rho factor, blocked by the ribosome from the boxA-rut element, is unable to load onto the RNA to catalyze transcription termination. In the absence of tryptophan, 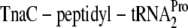 is cleaved and released, allowing Rho to terminate transcription prior to tnaAB transcript synthesis.
is cleaved and released, allowing Rho to terminate transcription prior to tnaAB transcript synthesis.
Those attending Stanford's graduate course “Gene Action” learned a four-step approach to studying regulation. First, understand the physiology by measuring gene expression during culture in different media and in response to metabolic shifts. Second, understand the genetics by characterizing mutants in which regulatory responses are altered. Third, understand the biochemistry by reconstituting regulation in vitro. Fourth, with accumulated information, materials, and assays in hand, return to in vivo studies to try to validate hypotheses. I think of these as “Yanofsky's Postulates.” The study of tnaAB regulation exemplifies this approach.
Physiology.
Tryptophanase (tryptophan-indole lyase), which catalyzes reversible hydrolysis of tryptophan to indole plus ammonium pyruvate (40), allows E. coli to use tryptophan as the sole carbon and nitrogen source, excreting indole. Indeed, tryptophan is one of the few amino acids efficiently catabolized from complex media (4, 38).
The tnaAB operon encodes tryptophanase (10) and a tryptophan permease essential for catabolism (39, 51). Expression is induced by extracellular tryptophan (51) and is subject to strong catabolite repression (2). This indicates that catabolism is the primary physiological function, although additional roles have been suggested (3, 41; reviewed in reference 49). tnaAB is the only operon whose expression is induced by tryptophan (29). Expression is not subject to general nitrogen regulation (15).
The 319-nucleotide (nt) transcribed leader region (tnaL) encodes the 24-residue TnaC regulatory peptide and contains sites for Rho-dependent transcription termination (Fig. 1) (43, 44). Catabolite repression acts exclusively at the cyclic AMP-activated promoter (11, 18, 26), whereas tryptophan induction acts exclusively by transcription antitermination within the leader region (18, 44). Expression is induced efficiently by 1-methyl-tryptophan, which is not charged onto tRNATrp (51). Thus, tryptophanyl-tRNATrp is not the inducer.
Rho-dependent transcription termination.
Rho factor catalyzes transcription termination, both in vitro and in vivo, at discrete sites within the 220-nt region between tnaC and tnaA (Fig. 1) (19, 43). A boxA-like element overlaps the carboxyl-terminus-coding region of the tnaC gene (44), and a rut (Rho utilization) element lies immediately downstream (16). boxA-like sequences are auxiliary elements for some Rho-dependent terminators (5).
Constitutive tnaAB expression results from substitutions and deletions altering the boxA-rut element, lesions in the rho structural gene, and treatment with the Rho inhibitor bicyclomycin (14, 16, 32, 44, 50). Similarly, boxA-rut alterations and bicyclomycin reduce Rho-dependent transcription termination in vitro (19, 43). Thus, tryptophan induction results from antitermination control within the leader region.
TnaC leader peptide.
Alterations that block or prematurely terminate tnaC translation result in low, uninducible tnaAB expression (14, 16, 45), whereas alterations that extend tnaC translation beyond the native UGA termination codon result in high constitutive expression (16, 44). Mutant tnaC alleles act in cis, demonstrating that the TnaC peptide is not a diffusible regulator (14, 44).
Findings to this point suggested a concrete hypothesis: a critical region of nascent TnaC “may act on its translating ribosome and, by affecting elongation, stalling, or release of the ribosome, modulate access of Rho factor to the nascent transcript” (14). This hypothesis was enabled by the work of Paul Lovett and colleagues, who characterized leader peptide control of antibiotic resistance enzyme synthesis (33). Since the boxA-rut element overlaps the tnaC carboxyl-terminus-coding region (Fig. 1), a ribosome stalled at the tnaC UGA termination codon will block Rho entry and thereby prevent transcription termination.
Attention was focused on the single Trp residue at position 12, initially to determine if tryptophanyl-tRNATrp is involved in regulation (14, 16) and subsequently to characterize its essential structural role in ribosome pausing, as summarized below. Further analysis of tnaC missense alterations identified the TnaC carboxyl-terminal half as important for regulation (14, 16), and TnaC sequences from other species share similarity in this region (14, 27). Indeed, parallel experiments with the Proteus vulgaris tna regulatory region have reinforced several observations made with E. coli (27, 28).
Leader peptide release in vivo.
Polypeptide release is catalyzed by termination codon-specific release factors RF1 (UAG and UAA) and RF2 (UGA and UAA), the activities of which are enhanced by RF3 (36). Loss of RF3 increases uninduced levels of tnaAB expression, whereas RF3 overexpression inhibits expression (52). When the UGA codon is converted to UAG, a mutant allele of the prfA gene encoding RF1 increases the uninduced expression level (31). Furthermore, tryptophan inhibits LacZ synthesis from a construct in which the tnaC UGA termination codon abuts the lacZ gene (30). Thus, tryptophan regulates tnaAB expression by inhibiting ribosome release from the tnaC termination codon.
Regulation reconstituted in vitro.
Physiology and genetics produced a solid hypothesis. For its test, Feng Gong and Charles Yanofsky developed an assay based on the cell-free coupled transcription-translation (S-30) system of Geoffrey Zubay (53). Added tryptophan or 1-methyl-tryptophan induces reporter synthesis from a wild-type template but not from tnaC mutant templates in which the codon for residue Trp-12 is replaced by one for Arg (W12R) or in which the initiation codon is destroyed. The Rho inhibitor bicyclomycin increases the uninduced level of expression, and antiterminated transcript accumulates in the presence of tryptophan or bicyclomycin (18). Thus, the cell-free system mimics the essential regulatory features as deduced from in vivo analyses.
For a stalled ribosome to inhibit transcription termination, leader peptide translation must coordinate with transcription elongation (47, 49). Experiments with the S-30 system show that this coupling results from a transcription pause just past the boxA-like element (21).
Leader peptide release in vitro.
In the absence of the inducer, the free TnaC leader peptide accumulates in cell-free reactions. In the presence of the inducer, TnaC-tRNAPro is detected (18) and remains bound to the ribosome (17). TnaC-tRNAPro is hydrolyzed upon addition of RF2, but only when the inducer is removed (17). Mutant TnaC(W12R)-tRNAPro is not detected even in the presence of the inducer (19). In S-30 extracts depleted of RF2, both wild-type and mutant TnaC-tRNAPro are detected in the absence of the inducer, and antiterminated transcript accumulates (19). Thus, the inducer inhibits RF2-mediated cleavage of TnaC-tRNAPro. The ribosomal inducer-binding site is stereospecific, because TnaC transfer to puromycin, an analog of A site-bound tRNA, is blocked only by l-tryptophan and closely related compounds (17).
TnaC residues Trp-12 and Pro-24 are essential for tryptophan-induced accumulation of TnaC-tRNAPro, as is their spacing (20). However, deletions, insertions, or missense alterations in the amino-terminal half have little effect on tryptophan induction (20), in accordance with earlier mutational analyses (14, 16).
Remarkably, insertion of a Trp codon between those for Pro-24 and termination causes accumulation of TnaC-tRNAPro even in the absence of the inducer, whereas insertion of a Phe codon blocks accumulation even in the presence of the inducer (20). Thus, tryptophanyl-tRNATrp delivers the inducer to the binding site, whereas phenylalanyl-tRNAPhe delivers a competitive inhibitor. This indicates that the leader peptide-directed inducer-binding site is near the A site in the ribosome, close to the peptidyl transferase center (20). This region binds several antibiotics, many of which, like tryptophan, contain aromatic groups (e.g., puromycin and chloramphenicol).
Leader peptide-ribosome interactions.
The exit tunnel accommodates at least 30 residues of nascent polypeptide in alpha-helical conformation (34, 42, 46). The SecM polypeptide undergoes a regulatory translation arrest at residue Pro-166, in which the essential residue Trp-155 lies near a constriction in the exit tunnel formed by protrusions of proteins L4 and L22 and domain II of 16S rRNA (1, 34, 37).
To determine where TnaC interacts, TnaC-tRNAPro-ribosome complexes were purified from S-30 reactions (8). Isolated complexes contain RF2, and tryptophan removal releases cleaved TnaC. In complexes depleted of RF2, TnaC residue Lys-11 cross-links to residue A750 in domain II of 23S rRNA, showing that residue Trp-12 is near the exit tunnel constriction. Wild-type TnaC but not TnaC(W12R) protects residue A788 from methylation, demonstrating that TnaC induces a conformational change displacing 23S rRNA domain II (8).
Tryptophan-binding site.
Next, RF2-depleted complexes were used to study interactions of the inducer tryptophan (6). Free tryptophan inhibits both methylation of residue A2572, which is required for peptidyl transferase activity, and TnaC transfer from tRNAPro to puromycin, which requires peptidyl transferase activity. Sparsomycin, a peptidyl transferase inhibitor, enhances methylation of residue A2059; this methylation is also inhibited by tryptophan. Tryptophan has none of these effects in complexes formed with the TnaC(W12R) mutant (6) or with mutant ribosomes with exit tunnel substitutions in 23S rRNA or L22 (7). This reinforces the conclusion that the nascent peptide-induced tryptophan-binding site is at or near the peptidyl transferase center (6).
Leader peptide residues Asp-16 and Pro-24.
This brings us to the featured work (9). Among available TnaC sequences, the critical residues Trp-12 and Asp-16 (14) are invariant, and Pro-24 is strongly conserved (9). Strikingly, arginine attenuator peptides from fungi share invariant residues Asp-12, Asp-16, and Ala-24 (12, 25). Thus, residues corresponding to these positions appear important in pause-directed formation of a metabolite binding site.
Results from the in vitro assays described above reveal that residues Asp-16 and Pro-24 are required both for tryptophan-induced TnaC-tRNAPro accumulation in S-30 extracts (shown previously for Pro-24 [20]) and for tryptophan-inhibited peptidyl transfer to puromycin in isolated, RF2-depleted ribosome complexes (9). Thus, like invariant residue Trp-12, these positions are critical for the TnaC leader peptide to induce formation of the ribosomal tryptophan-binding site.
Unlike ribosome complexes with wild-type or W12R mutant TnaC (6), those with D16A or P24A mutant TnaC do not exhibit sparsomycin-enhanced methylation of 23S rRNA residue A2059 (9). Sparsomycin does bind, because it inhibits peptidyl transfer to puromycin in D16A and P24A complexes. Thus, the locations of the D16A and P24A peptides in the exit tunnel appear to differ from those of the wild-type and W12R forms, indicating that these residues contribute to proper TnaC-exit tunnel interaction.
Finally, P24A mutant TnaC-tRNAAla exhibits increased transfer to puromycin, indicating that the carboxyl-terminal Pro residue may reduce peptidyl transferase activity, reduce the rate of transfer to RF2 (9), and increase ribosome stalling (35). In this scenario, tryptophan binding to the site induced by the TnaC leader peptide acts by further inhibiting an already compromised peptidyl transferase activity.
Broader implications.
The study of tryptophan metabolism has done much to advance our knowledge in biochemistry, genetics, and microbiology (48). Here, it has illuminated our understanding of ribosome structure, function, and regulation. The single-minded pursuit of mechanism for tnaAB control has generated an experimental and conceptual model for nascent peptide control in other contexts (1, 12, 33, 34, 37).
Back to the whole cell.
Overexpressed TnaC inhibits tnaAB induction (13) and inhibits growth (23), because accumulated 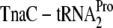 depletes this tRNA species (23). Changing codon Pro-24 from CCU to CCG results in depletion of the isoaccepting species
depletes this tRNA species (23). Changing codon Pro-24 from CCU to CCG results in depletion of the isoaccepting species 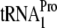 , whereas overproduction of the appropriate tRNA relieves inhibition in a codon-specific manner (23). Overexpressed RF3 and ribosome recycling factor inhibit chromosomal tnaAB induction (22, 52) and induce dropoff of overexpressed TnaC-tRNAPro (22). Further analysis employed ribosomal mutants screened for altered tnaAB expression or identified by others. These changes, in L22, L4, and 23S rRNA, all lie near the exit tunnel constriction and all influence tnaAB induction (7, 8). Thus, key elements of the hypothesis have been validated in vivo, thereby fulfilling the “fourth postulate.”
, whereas overproduction of the appropriate tRNA relieves inhibition in a codon-specific manner (23). Overexpressed RF3 and ribosome recycling factor inhibit chromosomal tnaAB induction (22, 52) and induce dropoff of overexpressed TnaC-tRNAPro (22). Further analysis employed ribosomal mutants screened for altered tnaAB expression or identified by others. These changes, in L22, L4, and 23S rRNA, all lie near the exit tunnel constriction and all influence tnaAB induction (7, 8). Thus, key elements of the hypothesis have been validated in vivo, thereby fulfilling the “fourth postulate.”
Acknowledgments
I am indebted to Charles Yanofsky for maintaining an exceptionally stimulating training environment and for continually supporting my independent research career.
The views expressed in this Commentary do not necessarily reflect the views of the journal or of ASM.
Footnotes
Published ahead of print on 16 May 2008.
REFERENCES
- 1.Beringer, M. 2008. Modulating the activity of the peptidyl transferase center of the ribosome. RNA 14795-801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Botsford, J. L., and R. D. DeMoss. 1971. Catabolite repression of tryptophanase in Escherichia coli. J. Bacteriol. 105303-312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Botsford, J. L., and R. D. DeMoss. 1972. Escherichia coli tryptophanase in the enteric environment. J. Bacteriol. 10974-80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chang, D. E., D. J. Smalley, D. L. Tucker, M. P. Leatham, W. E. Norris, S. J. Stevenson, A. B. Anderson, J. E. Grissom, D. C. Laux, P. S. Cohen, and T. Conway. 2004. Carbon nutrition of Escherichia coli in the mouse intestine. Proc. Natl. Acad. Sci. USA 1017427-7432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ciampi, M. S. 2006. Rho-dependent terminators and transcription termination. Microbiology 1522515-2528. [DOI] [PubMed] [Google Scholar]
- 6.Cruz-Vera, L. R., M. Gong, and C. Yanofsky. 2006. Changes produced by bound tryptophan in the ribosome peptidyl transferase center in response to TnaC, a nascent leader peptide. Proc. Natl. Acad. Sci. USA 1033598-3603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cruz-Vera, L. R., A. New, C. Squires, and C. Yanofsky. 2007. Ribosomal features essential for tna operon induction: tryptophan binding at the peptidyl transferase center. J. Bacteriol. 1893140-3146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cruz-Vera, L. R., S. Rajagopal, C. Squires, and C. Yanofsky. 2005. Features of ribosome-peptidyl-tRNA interactions essential for tryptophan induction of tna operon expression. Mol. Cell 19333-343. [DOI] [PubMed] [Google Scholar]
- 9.Cruz-Vera, L. R., and C. Yanofsky. 2008. Conserved residues Asp16 and Pro24 of TnaC-tRNAPro participate in tryptophan induction of tna operon expression. J. Bacteriol. 1904791-4797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Deeley, M. C., and C. Yanofsky. 1981. Nucleotide sequence of the structural gene for tryptophanase of Escherichia coli K-12. J. Bacteriol. 147787-796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Deeley, M. C., and C. Yanofsky. 1982. Transcription initiation at the tryptophanase promoter of Escherichia coli K-12. J. Bacteriol. 151942-951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fang, P., C. C. Spevak, C. Wu, and M. S. Sachs. 2004. A nascent polypeptide domain that can regulate translation elongation. Proc. Natl. Acad. Sci. USA 1014059-4064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gish, K., and C. Yanofsky. 1993. Inhibition of expression of the tryptophanase operon in Escherichia coli by extrachromosomal copies of the tna leader region. J. Bacteriol. 1753380-3387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gish, K., and C. Yanofsky. 1995. Evidence suggesting cis action by the TnaC leader peptide in regulating transcription attenuation in the tryptophanase operon of Escherichia coli. J. Bacteriol. 1777245-7254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Goldberg, R. B., and R. Hanau. 1980. Regulation of Klebsiella pneumoniae hut operons by oxygen. J. Bacteriol. 141745-750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gollnick, P., and C. Yanofsky. 1990. tRNATrp translation of leader peptide codon 12 and other factors that regulate expression of the tryptophanase operon. J. Bacteriol. 1723100-3107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gong, F., K. Ito, Y. Nakamura, and C. Yanofsky. 2001. The mechanism of tryptophan induction of tryptophanase operon expression: tryptophan inhibits release factor-mediated cleavage of TnaC-peptidyl-tRNAPro. Proc. Natl. Acad. Sci. USA 988997-9001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gong, F., and C. Yanofsky. 2001. Reproducing tna operon regulation in vitro in an S-30 system. Tryptophan induction inhibits cleavage of TnaC peptidyl-tRNA. J. Biol. Chem. 2761974-1983. [DOI] [PubMed] [Google Scholar]
- 19.Gong, F., and C. Yanofsky. 2002. Analysis of tryptophanase operon expression in vitro: accumulation of TnaC-peptidyl-tRNA in a release factor 2-depleted S-30 extract prevents Rho factor action, simulating induction. J. Biol. Chem. 27717095-17100. [DOI] [PubMed] [Google Scholar]
- 20.Gong, F., and C. Yanofsky. 2002. Instruction of translating ribosome by nascent peptide. Science 2971864-1867. [DOI] [PubMed] [Google Scholar]
- 21.Gong, F., and C. Yanofsky. 2003. A transcriptional pause synchronizes translation with transcription in the tryptophanase operon leader region. J. Bacteriol. 1856472-6476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gong, M., L. R. Cruz-Vera, and C. Yanofsky. 2007. Ribosome recycling factor and release factor 3 action promotes TnaC-peptidyl-tRNA dropoff and relieves ribosome stalling during tryptophan induction of tna operon expression in Escherichia coli. J. Bacteriol. 1893147-3155. [DOI] [PMC free article] [PubMed] [Google Scholar]
-
23.Gong, M., F. Gong, and C. Yanofsky. 2006. Overexpression of tnaC of Escherichia coli inhibits growth by depleting
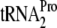 availability. J. Bacteriol. 1881892-1898. [DOI] [PMC free article] [PubMed] [Google Scholar]
availability. J. Bacteriol. 1881892-1898. [DOI] [PMC free article] [PubMed] [Google Scholar] - 24.Henkin, T. M., and C. Yanofsky. 2002. Regulation by transcription attenuation in bacteria: how RNA provides instructions for transcription termination/antitermination decisions. Bioessays 24700-707. [DOI] [PubMed] [Google Scholar]
- 25.Hood, H. M., C. C. Spevak, and M. S. Sachs. 2007. Evolutionary changes in the fungal carbamoyl-phosphate synthetase small subunit gene and its associated upstream open reading frame. Fungal Genet. Biol. 4493-104. [DOI] [PubMed] [Google Scholar]
- 26.Isaacs, H., Jr., D. Chao, C. Yanofsky, and M. H. Saier, Jr. 1994. Mechanism of catabolite repression of tryptophanase synthesis in Escherichia coli. Microbiology 1402125-2134. [DOI] [PubMed] [Google Scholar]
- 27.Kamath, A. V., and C. Yanofsky. 1992. Characterization of the tryptophanase operon of Proteus vulgaris. Cloning, nucleotide sequence, amino acid homology, and in vitro synthesis of the leader peptide and regulatory analysis. J. Biol. Chem. 26719978-19985. [PubMed] [Google Scholar]
- 28.Kamath, A. V., and C. Yanofsky. 1997. Roles of the tnaC-tnaA spacer region and Rho factor in regulating expression of the tryptophanase operon of Proteus vulgaris. J. Bacteriol. 1791780-1786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Khodursky, A. B., B. J. Peter, N. R. Cozzarelli, D. Botstein, P. O. Brown, and C. Yanofsky. 2000. DNA microarray analysis of gene expression in response to physiological and genetic changes that affect tryptophan metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 9712170-12175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Konan, K. V., and C. Yanofsky. 1997. Regulation of the Escherichia coli tna operon: nascent leader peptide control at the tnaC stop codon. J. Bacteriol. 1791774-1779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Konan, K. V., and C. Yanofsky. 1999. Role of ribosome release in regulation of tna operon expression in Escherichia coli. J. Bacteriol. 1811530-1536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Konan, K. V., and C. Yanofsky. 2000. Rho-dependent transcription termination in the tna operon of Escherichia coli: roles of the boxA sequence and the rut site. J. Bacteriol. 1823981-3988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lovett, P. S., and E. J. Rogers. 1996. Ribosome regulation by the nascent peptide. Microbiol. Rev. 60366-385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mankin, A. S. 2006. Nascent peptide in the “birth canal” of the ribosome. Trends Biochem. Sci. 3111-13. [DOI] [PubMed] [Google Scholar]
- 35.Muto, H., and K. Ito. 2008. Peptidyl-prolyl-tRNA at the ribosomal P-site reacts poorly with puromycin. Biochem. Biophys. Res. Commun. 3661043-1047. [DOI] [PubMed] [Google Scholar]
- 36.Nakamura, Y., and K. Ito. 2003. Making sense of mimic in translation termination. Trends Biochem. Sci. 2899-105. [DOI] [PubMed] [Google Scholar]
- 37.Nakatogawa, H., A. Murakami, and K. Ito. 2004. Control of SecA and SecM translation by protein secretion. Curr. Opin. Microbiol. 7145-150. [DOI] [PubMed] [Google Scholar]
- 38.Prüss, B. M., J. M. Nelms, C. Park, and A. J. Wolfe. 1994. Mutations in NADH:ubiquinone oxidoreductase of Escherichia coli affect growth on mixed amino acids. J. Bacteriol. 1762143-2150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sarsero, J. P., P. J. Wookey, P. Gollnick, C. Yanofsky, and A. J. Pittard. 1991. A new family of integral membrane proteins involved in transport of aromatic amino acids in Escherichia coli. J. Bacteriol. 1733231-3234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Snell, E. E. 1975. Tryptophanase: structure, catalytic activities, and mechanism of action. Adv. Enzymol. Relat. Areas Mol. Biol. 42287-333. [DOI] [PubMed] [Google Scholar]
- 41.Stancik, L. M., D. M. Stancik, B. Schmidt, D. M. Barnhart, Y. N. Yoncheva, and J. L. Slonczewski. 2002. pH-dependent expression of periplasmic proteins and amino acid catabolism in Escherichia coli. J. Bacteriol. 1844246-4258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Steitz, T. A. 2008. A structural understanding of the dynamic ribosome machine. Nat. Rev. Mol. Cell Biol. 9242-253. [DOI] [PubMed] [Google Scholar]
- 43.Stewart, V., R. Landick, and C. Yanofsky. 1986. Rho-dependent transcription termination in the tryptophanase operon leader region of Escherichia coli K-12. J. Bacteriol. 166217-223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Stewart, V., and C. Yanofsky. 1985. Evidence for transcription antitermination control of tryptophanase operon expression in Escherichia coli K-12. J. Bacteriol. 164731-740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stewart, V., and C. Yanofsky. 1986. Role of leader peptide synthesis in tryptophanase operon expression in Escherichia coli K-12. J. Bacteriol. 167383-386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Voss, N. R., M. Gerstein, T. A. Steitz, and P. B. Moore. 2006. The geometry of the ribosomal polypeptide exit tunnel. J. Mol. Biol. 360893-906. [DOI] [PubMed] [Google Scholar]
- 47.Yanofsky, C. 2000. Transcription attenuation: once viewed as a novel regulatory strategy. J. Bacteriol. 1821-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yanofsky, C. 2001. Advancing our knowledge in biochemistry, genetics, and microbiology through studies on tryptophan metabolism. Annu. Rev. Biochem. 701-37. [DOI] [PubMed] [Google Scholar]
- 49.Yanofsky, C. 2007. RNA-based regulation of genes of tryptophan synthesis and degradation, in bacteria. RNA 131141-1154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Yanofsky, C., and V. Horn. 1995. Bicyclomycin sensitivity and resistance affect Rho factor-mediated transcription termination in the tna operon of Escherichia coli. J. Bacteriol. 1774451-4456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yanofsky, C., V. Horn, and P. Gollnick. 1991. Physiological studies of tryptophan transport and tryptophanase operon induction in Escherichia coli. J. Bacteriol. 1736009-6017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yanofsky, C., V. Horn, and Y. Nakamura. 1996. Loss of overproduction of polypeptide release factor 3 influences expression of the tryptophanase operon of Escherichia coli. J. Bacteriol. 1783755-3762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zubay, G. 1973. In vitro synthesis of protein in microbial systems. Annu. Rev. Genet. 7267-287. [DOI] [PubMed] [Google Scholar]