

Abstract
Spontaneous calcium activity of neural progenitors is largely dependent on a paracrine signaling mechanism involving release of ATP and activation of purinergic receptors. Although it is well documented that, in mature astrocytes, cytokines modulate the expression levels of certain purinergic receptors, nothing is known about their impact during early stages of development. Here we provide evidence that conditioned medium from activated microglia as well as interleukin-1β, but not tumor necrosis factor-α, decrease the frequency of calcium oscillations and reduce the rate of “in vitro” migration of astrocyte progenitors. Such alterations were due to changes in activity of two purinergic P2 receptors, and not to the amount of released ATP. These results indicate that Interleukin-1β plays important role during early stages of CNS development, modulating calcium signaling and cell migration.
Keywords: cytokine, microglia, neurospheres, purinergic receptors, IL-1β, TNFα
1. Introduction
Studies on repair mechanisms have indicated that the inflammatory microenvironment is an important component involved in the engagement of endogenous and/or exogenous neural stem/progenitor cells. These include observations that neural progenitors cells (NPC) increase their proliferation and migrate toward pathologic sites derived from diverse etiologies in in vivo and in vitro experiments (Picard-Riera et al., 2002; Ben-Hur et al., 2003b; Kokaia and Lindvall, 2003; Imitola et al., 2004; Kelly et al., 2004; Pluchino et al., 2005). These observations have led some authors (Imitola et al., 2004) to propose that the inflammatory cues guide the behavior of potentially reparative stem cells. However, brain injury can also lead to the establishment of a “non-permissive” molecular milieu that could favor apoptotic cell death and would prevent migration, proliferation and differentiation of NPC (Fok-Seang et al., 1998; Fawcett and Asher 1999; Monje et al., 2003). Consequently, the ultimate effect of the neuroinflammatory milieu on the development and fate of neural progenitors is complex and likely to depend on a balance between proliferation and apoptotic-inducing factors, and differentiation-inducing factors. The inflammatory response in the central nervous system (CNS) involves the activation of microglia, which releases inflammatory mediators, such as acute phase proteins, eicosanoids, complement, chemokines, free radicals, and pro-inflammatory cytokines (Allan and Rothwell, 2001; Monje et al., 2003).
Several lines of evidence indicated that the migration of neuronal and glial progenitor cells is dependent on intracellular calcium elevation induced by the release of neurotransmitters and modulators (Komuro and Rakic, 1998; Komuro and Kumada, 2005; Kumada and Komuro, 2004; Scemes et al., 2003; Agresti et al., 2005a,b; Striedinger et al., 2007). At least for glial progenitor cell migration, stimulation of purinergic P2Y1 receptor by extracellular ATP have been proposed as a key component mediating their rate of migration (Scemes et al., 2003; Agresti et al., 2005a,b; Striedinger et al., 2007).
Because purinergic receptor expression and activity can be modulated by pro-inflammatory cytokines (John et al., 1999; Narcisse et al., 2005; Scemes, 2008), we evaluated the effects of conditioned medium of activated microglia (CM), as well as those induced by interleukin-1β (IL-1β) and tumor necrosis factor-α (TNFα) on astrocyte progenitor cell migration and calcium signaling. Our results indicate that, similarly to the effects of conditioned medium of activated microglia, the pro-inflammatory cytokine IL-1β, but not TNFα reduced the rate of in vitro progenitor cell migration. Moreover, the reduced cell migration induced by IL-1β was paralleled by decreased frequency of spontaneous calcium oscillations due to reduced activity but not expression level of the P2Y1 receptor.
2. Materials and Methods
Neurospheres and neural progenitor cultures
Neurospheres were prepared as previously described (Striedinger et al., 2007). Briefly, neural progenitor cells were obtained by aspiration of forebrain tissues of 14 day-old (E14) wild-type C57Bl/6 mouse embryos (time-pregnant C57Bl/6 females obtained from Charles River) and mechanically dissociated into single cells in ice-cold Hanks balanced solution (HBSS Ca2+- and Mg2+-free). The Albert Einstein College of Medicine (AECOM) Animal Care and Use Committee approved all experimental procedures performed. Viable cells were transferred to tissue culture dishes containing Dulbecco Modified Eagle Medium Nutrient Mixture F12 (DMEM-F12, Gibco, Invitrogen, Carlsbad, CA) supplemented with 5% B27 (Gibco), 1% antibiotics and 20ng/ml human recombinant epidermal growth factor (EGF; Sigma, St. Louis, MO), and allowed to grow into floating neurospheres. Culture medium was changed twice a week, and neurospheres were mechanically dissociated into smaller neurospheres once a week. Neurosphere cultures were maintained for no longer than 2 months. For in vitro cell differentiation, floating neurospheres were plated on glass bottom microwells (MaTek Co, Ashland, MA) coated with poly-D-lysine- (10μg/ml; Sigma) and fibronectin- (10μg/ml; Sigma) and bathed in DMEM-F12 in the absence of EGF. After adhesion of neurospheres to the substrate, cells migrating out of the neurospheres differentiate into glia and neuronal progenitors.
Conditioned medium from activated microglia
Purified cultures of murine microglia were kindly provided by Dr. Kostantin Dobrenis (Department of Neuroscience, Albert Einstein College of Medicine), and prepared as previously described (Dobrenis et al., 2005) from neocortex of 2-day postnatal C57Bl/6 mice. Purified microglia cultures were maintained in Macrophage-SPM Medium (Gibco) supplemented with 10ng/ml recombinant mouse granulocyte-macrophage colony-stimulating factor (GM-CSF; R&D, Minneapolis, MN), 5% fetal bovine serum (FBS) and 1% Penicillin-Streptomycin. Conditioned media (CM) from GM-CSF activated microglia were obtained by replacing the Macrophage medium with the neurospheres DMEM-F12 medium, which was then collected 24 hrs after medium substitution, filtered and stored at −20°C until further use.
ELISA Assay
The level of interleukin-1β (IL-1β) present on the microglia conditioned media (CM) was measured using the quantitative sandwich enzyme immunoassay technique (Quantikine, R&D Systems) as described by the manufacture. Briefly, 50μl of samples (CM and microglia cell lysates) and 50μl of a buffer solution were added to a 96 well plate pre-coated with a polyclonal antibody specific for the mouse IL-1β and incubated for 2 hr at room temperature (RT). After several washes, 100μl of a polyclonal HRP-conjugated antibody against mouse IL-1β were added to each well. Following 2hr incubation and several washes, 100μl chromogen-hydrogen peroxide solution was added to each well and reaction incubated for 30 min before the addition of a hydrochloric acid (stop) solution. Optical densities were obtained using a microplate reader (Dynex Revelation 4.21) and the concentration (pg/ml) of IL-1β calculated based on a standard curve. All samples were run in triplicates.
Cytokine treatment
Neurospheres (100–200μm diameters) were treated in suspension for 24 hrs with CM, IL-1β (20ng/mL; Cedarlane) or tumor necrosis factor-α (TNFα: 10ng/mL; Chemicon) and thereafter plated on coated dishes, in the continuous presence of these cytokines, for migration and calcium assays (see below). Parallel experiments were performed on untreated neurospheres. In some experiments the neutralizing IL-1 receptor antibody (100ng/mL; Antigenix Inc.) was used.
Migration assay
To evaluate the distances of emigration of progenitor cells from the neurospheres, the outgrowth index (OI) was calculated as the ratio between the distance of the nucleus of the foremost cells to the center of the sphere and the radius of the sphere, as previously described (Scemes et al., 2003; Striedinger et al., 2007). In vitro cell migration was induced by withdrawal of EGF from the culture medium of untreated and treated neurospheres plated on poly-D-lysine/fibronectin coated glass bottom microwells. Twenty-four hrs CM-, IL-1β-, or TNFα-treated floating neurospheres were then seeded on coated dishes and the OI measured daily during three days in culture in the continuous presence of these inflammatory mediators. In some experiments, neurospheres were treated with the P2X7R agonist (100 μM BzATP) as described above for the cytokines.
Spontaneous calcium oscillations
Spontaneous intracellular Ca2+ transients were measured in adherent, Cd44S immuno-identified astrocyte progenitors, as previously described (Striedinger et al., 2007). Briefly, adherent cultures of progenitors were incubated in serum-free DMEM-F12 containing monoclonal anti-Cd44S (1:200) antibodies for 30 min at 37°C. After several washes with DMEM-F12, the cultures were incubated with Alexa fluor 594-tagged secondary antibodies, diluted 1:500 and concomitantly loaded with the Ca2+ indicator Fluo-3-AM (5μM; Molecular Probes-Invitrogen) at 37oC for 40 min. Cells were then washed with Dulbecco’s phosphate buffered saline (DPBS, pH 7.4; Cellgro, Herndon, VA) and imaged on an epifluorescence microscope (Eclipse TE2000-S; Nikon, Japan), equipped with FITC and Texas red filter sets. Images were acquired with a CCD camera (Orca-ER; Hamamatsu, Japan). Fluo-3 fluorescence intensity emitted at one excitation wavelength (488 nm) was continuously acquired at a rate of 1.0 Hz for 5 min period using combined systems of filters and shutter (Lambda DG-4 Diaphot, Sutter Instruments Co., Burlingame, CA) driven by a computer through Metafluor software (Universal Imaging Systems, Downingtown, PA). Fluo-3 fluorescence intensity obtained from regions of interest placed on Cd44S positive cells were normalized to initial values and expressed as relative changes in fluorescence intensity (F/F0).
Ca2+ mobilization induced by P2 receptor agonists
P2 receptor agonists with affinity for the metabotropic receptor P2Y1R, 2-MeSATP (100nM; 2-methylthioadenosine 5′-triphosphate; Sigma-Aldrich, St Louis, MO) and for the ionotropic receptor P2X7R, BzATP (100μM; 3′O-4-bezoylbenzoyl-ATP; Sigma) were employed to evaluate the effects of IL-1β on P2Y1 and P2X7R-mediated Ca2+ mobilization of 3 day-old astrocyte Cd44S positive progenitors. For that, cells were loaded with the ratiometric Ca2+ indicator, Fura-2-AM (2.5 μM; Molecular Probes-Invitrogen) for 40 min. Fura-2 ratio values were translated into intracellular Ca2+ concentrations according to an in vitro calibration curve of the form: [Ca2+]i = KD{(R-Rmin)/(Rmax-R)} (F380min/F380max), where [Ca2+]i is the calculated intracellular calcium concentration, KD is the dissociation constant of free-Ca2+ for Fura-2 (KD = 224 nM; Grynkiewicz et al., 1985), R is the ratio intensity, Rmin is the ratio of the intensity obtained at zero calcium, Rmax is the ratio of the intensity at saturated calcium, F380min is the fluorescence intensity measured with zero calcium at 380nm, and F380max is the fluorescence intensity measured with saturated calcium at 380nm. At least three independent cultures of neurospheres were used to measure the amplitudes of Ca2+ transients induced by the P2R agonists.
ATP assays
The amount of ATP released in the bathing solution by untreated and cytokine-treated 3 day-adherent progenitor cells was measured for 2 min in 200μl DPBS, as previously described (Striedinger et al., 2007). The amount of ATP was measured using the luciferin/luciferase assay (Molecular Probes- Invitrogen). Fifty microliters of a buffered solution containing luciferin (50μM) and luciferase (1.25μg/ml) were placed in triplicates in a 96-well plate luminometer (Veritas,Turner) for background luminescence subtraction. Reactions were started by adding 5μl of the experimental samples and luminescence values acquired during 1 sec integration time. The amount of ATP in the samples was calculated from standard curves and normalized for the total amount of protein, using the BCA assay (Pierce, Rockford, IL).
Western Blots
Untreated and IL-1β-(20ng/ml) treated 3-day adherent progenitor cells were lysed in lyses buffer (150mM NaCl, 10mM Tris-HCl, 1% Triton-X-100, Roche complete protease inhibitor cocktail). Samples of whole cell lysates were mixed with 2XLaemli buffer and subjected to electrophoresis using a 4–20% SDS-PAGE mini-gel (BioRad). After transfer of proteins to nitrocelulose membranes (Schleicher & Schuell, Keene, NH), membranes were incubated for 1hr at RT with blocking solution (1XPBS, 0.5% Tween-Twenty, 2% non-fat dry milk) prior to exposure to primary antibodies. Membranes were blotted for 2hrs at RT with anti-P2Y1 (1:200) and anti-P2X7 (1:500) antibodies (Alomone labs) prepared in blocking solutions. After three washes (1XPBS, 0.5% Tween-Twenty), membranes were incubated with goat-anti-rabbit-HRP conjugated secondary antibodies (1:2000; Santa Cruz Biotechnology, Santa Cruz, CA) for 1hr at RT. After several washes, detection of bands was performed on X-ray films (Kodak, Rochester, NY) following incubation with enhanced chemiluminescence (ECL) reagents (Amersham Pharmacia Biotechnology, Piscataway, NJ). Membranes were re-probed with anti-GAPDH (1:2000; Fitzgerald). Quantification was performed by densitometric analysis of bands using Image-J and P2R levels normalized to that of GAPDH.
Statistical analyses
All data are expressed as mean±SE. GraphPad Prism version 4 was used for statistical analysis, consisting of one-way ANOVA analysis of variance, followed by Newman-Keuls’ multiple comparison test. T-test comparison was also employed in some cases.
3. Results
Conditioned medium from activated microglia (CM) and IL1-β but not TNFα decrease the migration distances of neural progenitor cells
We have previously reported (Striedinger et al., 2007) that the great majority (> 80%) of progenitor cells derived from fibronectin adherent neurospheres are of the astrocytic lineage that express the extracellular glycoprotein Cd44S (a marker for astrocyte progenitors: Rao and Mayer-Proschel, 1997; Liu et al., 2002; Liu and Rao, 2004).
Using this neurosphere system, we evaluated the effects of microglia conditioned medium and of two cytokines on the migration of astrocyte progenitors.
When adherent neurosphere cultures were exposed for 3 days to CM, a significant decrease on the distance of migration of astrocyte progenitor cells was observed compared to untreated cultures (Fig. 1A). Under these conditions, the outgrowth index (OI) of CM-treated neurospheres was 7.66±0.94 (N=29 neurospheres), while the OI of control neurospheres was 11.04±0.26 (N=66 neurospheres). To further investigate which of the main inflammatory mediators contained in the conditioned media from activated microglia was mediating the reduced OI, we treated neurospheres with two pro-inflammatory cytokines, IL-1β and TNFα that have been previously reported to be released from activated microglia (Aloisi et al., 1997; Degousee et al., 2001; Meme et al., 2006).
Figure 1. Effect of pro-inflammatory cytokines on the “in vitro” migration of neural progenitor cells.
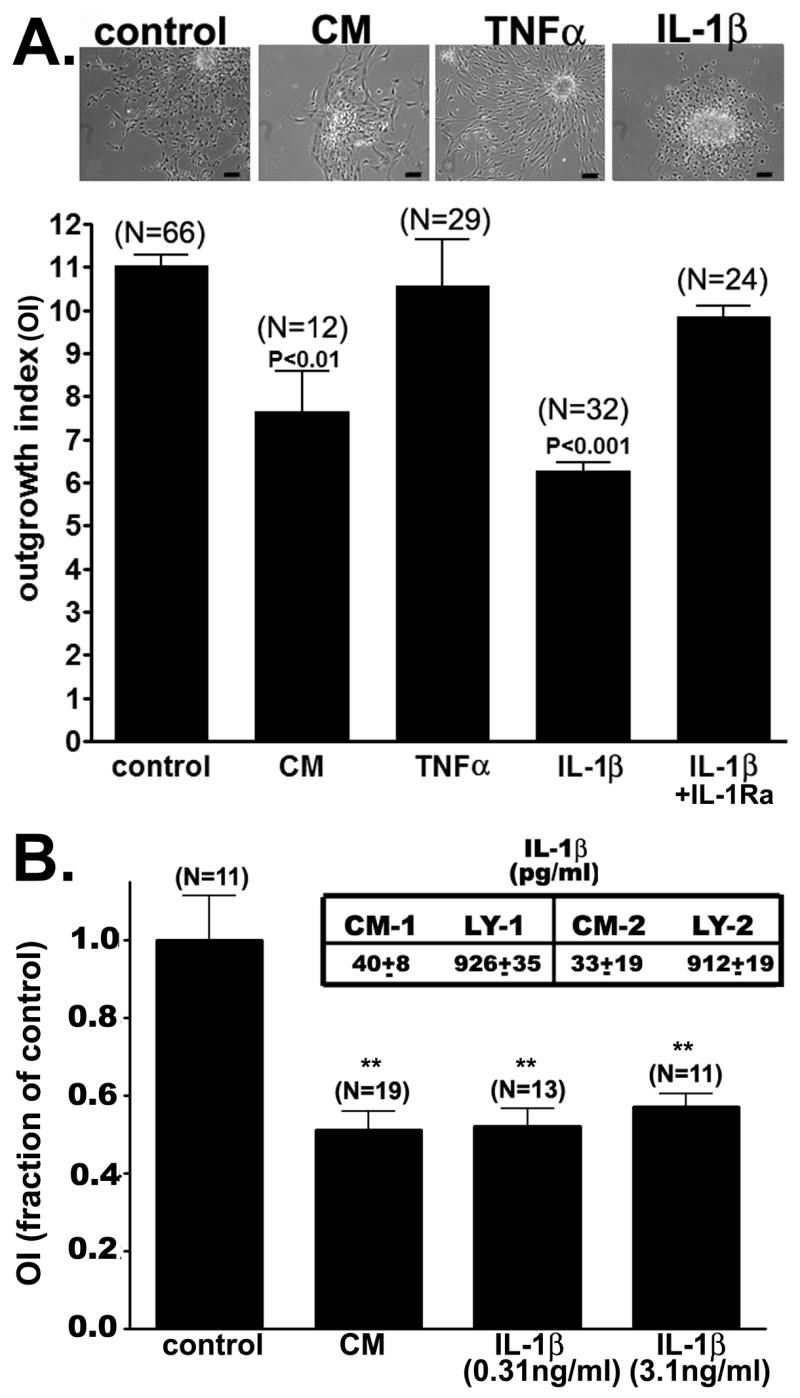
(A) Bar histograms showing the mean±SE values obtained for the outgrowth index from 3-days adherent neurospheres that were untreated (control) and treated with conditioned media from activated microglia (CM), IL-1β (20ng/ml), TNFα (10ng/ml) and with the neutralizing IL-1 receptor antibody (100ng/ml). Note that only CM and IL-1β caused a significant decrease in the outgrowth index compared to untreated cells (ANOVA followed by Newman-Keuls’ multiple comparison test; N represents the number of neurospheres used from at least three independent). (Top) Representative phase-contrast images obtained from 3-days adherent neurosphere cultures untreated (control) and treated with CM, IL-1β, and TNFα. Bars: 40 μm. (B) Bar histograms of the mean±SE values obtained for the relative changes in OI from 3-day adherent neurospheres untreated (control) and treated with CM containing 0.03ng/ml IL-1β and with IL-1β (0.31ng/ml and 3.1ng/ml). Inset shows the mean±SE values (in pg/ml) of IL-1β concentration measured in the conditioned media (CM) and in the lysate (LY) fractions obtained from two different cultures of microglia cells. (** P<0.001, ANOVA followed by Newman-Keuls’ multiple comparison test; N represents the number of neurospheres used).
After 3 days of adhesion to poly-D-lysine/fibronectin substrate, IL-1β-treated progenitors displayed a significant reduction of the outgrowth index (OI=6.27±0.20; N=32 neurospheres) compared to untreated cells (OI=11.04±0.26; N=66 neurospheres, p<0.001; Fig. 1A). This decreased OI was prevented by co-treating the neurospheres with the neutralizing IL-1 receptor antibody (OI=9.86±0.26; N=24 neurospheres). In contrast to the effects of IL-1β, after three days of adhesion, the outgrowth index of TNFα-treated progenitors (OI=10.58±1.06; N=29 neurospheres; Fig. 1A) was not significantly different to that of control, untreated progenitors.
Using an ELISA assay we quantified the amount of IL-1β present on the CM obtained from two cultures of microglia cells. As shown in the table displayed in Fig. 1B, about 30–40pg/ml IL-1β were detected in CM, and about 900pg/ml were found in the cytosol of microglia. To evaluate whether lower concentrations of IL-1β than those used in the experiments described above could also reduce the outgrowth index, neurospheres were treated for 3 days with 0.31 and 3.1 ng/ml IL-1β. Even at lowest concentration used (0.31ng/ml), IL-1β caused a similar reduction in the outgrowth index seen in cultures treated with CM (0.03ng/ml IL-1β) (Fig. 1B). These results suggest that the levels of IL-1β found in CM are sufficient to maximally reduce progenitor cell migration.
Given that 10ng/ml TNFα (a concentration similar - 23ng/ml - to that found in CM of mouse microglia: Meme et al., 2006) did not cause any effect on astrocyte progenitor cell migration, we did not quantify the levels of this cytokine in CM.
Spontaneous Ca2+ oscillations in astrocyte progenitors are attenuated by CM and IL-1β
We and others have previously shown that the frequency of spontaneous calcium oscillations of neural progenitor cells dictates their rate of migration (Komuro and Rakic, 1998; Kumada and Komuro, 2004; Komuro and Kumada, 2005; Scemes et al., 2003; Striedinger et al., 2007). To evaluate whether the effect of CM and IL-1β on the outgrowth index of astrocyte progenitors was related to reduced Ca2+ activity, we measured the number of astrocyte progenitors displaying spontaneous Ca2+ transients, as well as the frequencies of these Ca2+ oscillations. For that, we loaded the cells with the Ca2+ indicator Fluo-3-AM and recorded Ca2+ activity from Cd44S positive cells.
Our results indicate that the fraction of astrocyte progenitors (Cd44S+) displaying spontaneous Ca2+ oscillations recorded under control conditions (0.54± 0.04; N=4 experiments) was significantly reduced to 0.38±0.04 and to 0.38± 0.02 (P<0.05; ANOVA analysis of variance and Newman Keuls’ multiple comparison test) when treated with CM and with IL-1β, respectively. TNFα, in contrast, did not alter the fraction of astrocyte progenitors displaying spontaneous Ca2+ oscillations (0.55±0.09; p>0.05) compared to control untreated cells. The frequency of spontaneous Ca2+ oscillations was also greatly reduced after treating the cells with CM and IL-1β. As shown in Figure 2, the frequency of Ca2+ oscillations (10.81±0.34 mHz; N=571 cells) recorded from untreated cultures was significantly reduced in CM- (6.69±0.36 mHz; N=122 cells; p<0.001) and IL-1β-(7.09±0.35 mHz; N=203 cells; p<0.001) treated cells; TNFα did not significantly alter the frequency of spontaneous Ca2+ oscillations (11.17±0.63 mHz; N=161 cells; p >0.05) compared to that obtained from control cells.
Figure 2. Frequency of spontaneous Ca2+ oscillations in astrocyte progenitors exposed to cytokines.

(A) Example of intracellular Ca2+ oscillations recorded from Fluo-3-AM loaded progenitor cells that were untreated and treated with IL1-β. Each trace corresponds to a single cell. (B) Bar histograms showing the mean ± SE values of the frequency (mHz) of spontaneous intracellular Ca2+ oscillations recorded from 3 days-adherent astrocyte progenitors that were untreated (control) and treated with conditioned media from activated microglia (CM), IL-1β (20ng/ml), and TNFα (10ng/ml). Note the marked reduction in frequency of spontaneous Ca2+ activity in CM and IL-1β-treated progenitors compared to control (ANOVA followed by Newman-Keuls’ multiple comparison test).
These data indicate that the reduced frequency of spontaneous Ca2+ oscillations, together with the reduced number of Ca2+ oscillating astrocyte progenitors contribute to the reduced cell migration observed by treating the cells with CM and IL-1β.
IL-1β does not alter the amount of ATP released from progenitor cells but altered purinergic receptor agonist mediated calcium transients
Given that we have previously shown that spontaneous Ca2+ oscillations of astrocyte progenitors are dependent upon ATP release and subsequent activation of purinergic P2Y1 receptors (Scemes et al., 2003; Striedinger et al., 2007), we investigated whether IL-1β and TNFα affected the amount of ATP released from these cells. For that, we measured the amount of ATP present on the bathing solution of 3-days adherent progenitors using the luciferase assay. As shown in Figure 3, no significant changes in the amount of released ATP were observed compared to control untreated cells.
Figure 3. Effects of cytokines on the amount of ATP release.
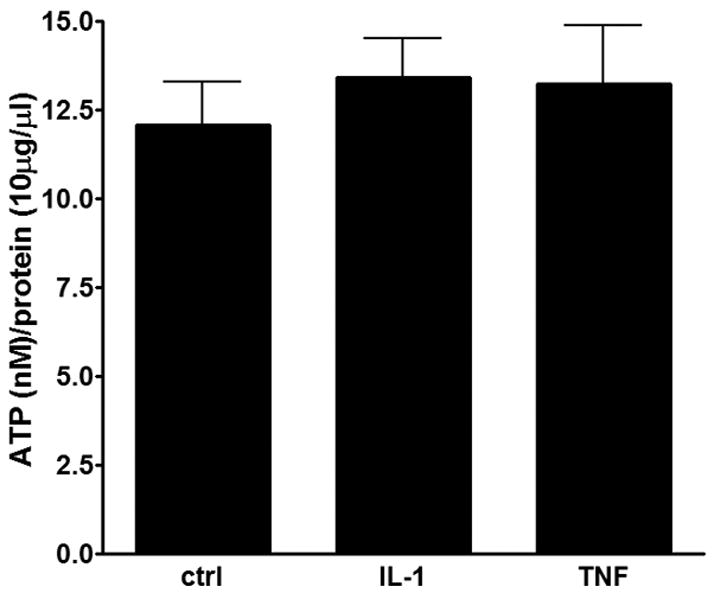
Bar histograms showing the mean±SE values obtained for the amount of ATP released from 3-days adherent progenitors that were untreated and treated with IL-1β and TNFα.
To evaluate whether changes in spontaneous calcium activity induced by IL-1β were related to changes in purinergic receptor activity, the amplitudes of Ca2+ transients induced by P2Y1 and P2X7 receptor agonists were measured on Cd44S positive progenitors derived from 3-days adherent neurospheres that were untreated or treated with this cytokine. As shown in Figure 4, intracellular Ca2+ levels mobilized by the P2Y1 receptor agonist 2-Me-S-ATP (100nM) were significantly reduced in IL-1β (394.71±35.87nM; N=175 cells; P<0.01) treated progenitors compared to controls (977±55.78nM; N=175 cells). In contrast to the action of P2Y1R agonist, the P2X7 R agonist BzATP (100μM) caused a significant sustained increase in intracellular Ca2+ levels in IL-1β-treated cells (612±45.06nM; N=192 cells; P<0.01; Fig. 4) compared to control cells.
Figure 4. Changes in purinergic P2 receptor activity induced by IL-1β.
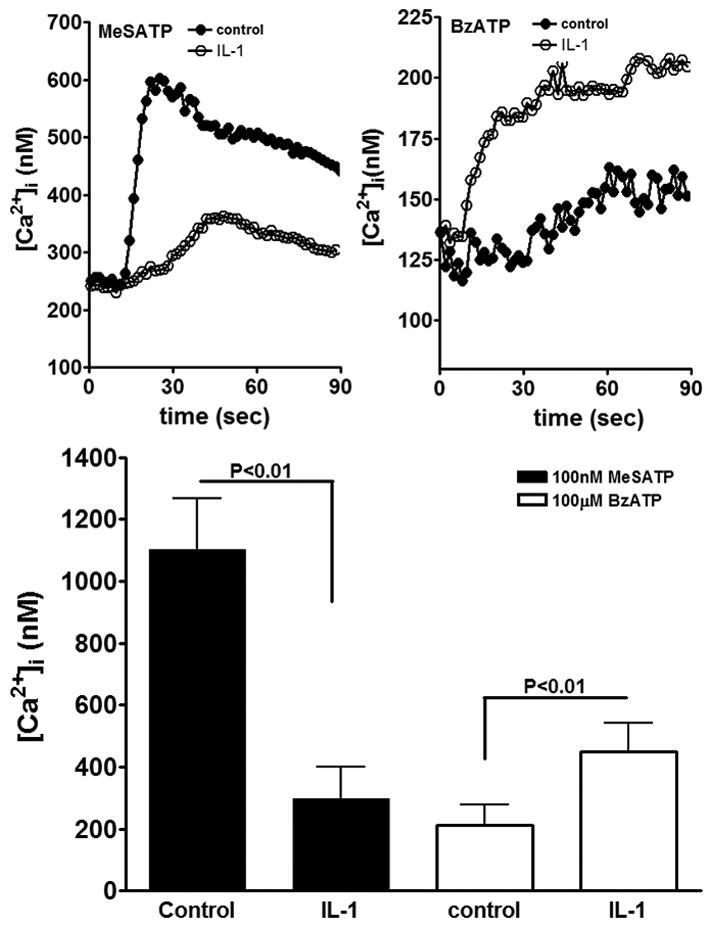
Time course of intracellular calcium changes induced by the two P2 receptor agonists showing the reduced response to the P2Y1R agonist (2-MeSATP) (A) and the increased response to the P2X7R agonist (B) in IL-1β-treated progenitors compared to controls (Student T test). (C) Bar histograms showing the mean±SE values obtained for the amplitudes of intracellular Ca2+ transients induced by 100nM MeSATP and 100μM BzATP in untreated (control) and IL-1β treated progenitors.
These data indicate that IL-1β attenuates P2Y1 receptor mediated intracellular calcium mobilization and increases the activity of P2X7 receptor.
In order to evaluate whether changes in activity of the two P2Rs were related to altered protein expression levels, western blots were performed in 3-days adherent progenitors untreated and treated with 20ng/ml IL-1β. Western blot analysis revealed that the expression levels of P2X7 and P2Y1 receptors were unchanged following IL-1β treatment, compared to control, untreated progenitors (Fig. 5).
Figure 5. Expression levels of P2 receptors.
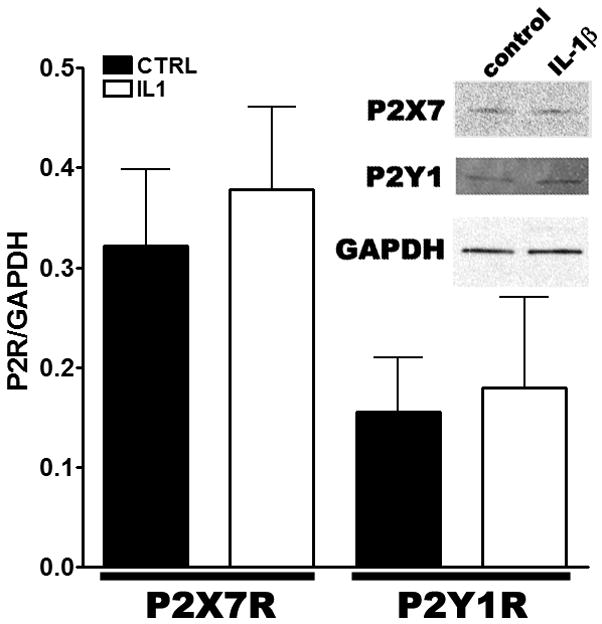
Bar histograms showing the expression levels of P2 (P2X7 and P2Y1) receptors normalized to that of GAPDH obtained from untreated (black bars) and IL-1β treated progenitors (white bars). IL-1β did not significantly changed P2 receptors expression levels (P>0.05; T-test; N=2–5). Inset. Representative Western blot showing the expression of the two P2 receptors in untreated (control) and IL-1β treated progenitors.
Contribution of P2R to neural progenitor cell migration
Regarding the contribution of these P2 receptors to astrocyte progenitor migration, it is likely that the P2Y1 and not the P2X7 receptors are involved in this process. This is because we have previously reported that a P2Y1R antagonist, MRS2179, but not the P2X7R antagonist KN62, reduced progenitor cell migration (Scemes et al., 2003). Further support that the P2X7R does not contribute to this process was obtained from experiments in which adherent neurospheres were treated for three days with BzATP (100μM). Thus, after 3 days of adhesion, BzATP-treated cells featured an outgrowth index (OI) of 12.35±0.82 (N=16 neurospheres from 3 independent experiments), which was not statistically different (P>0.05; unpaired T-test) from that recorded from untreated cells (OI=13.16±1.26; N=11 neurospheres from 3 independent experiments).
Thus, our previous (Scemes et al., 2003) and current results support the hypothesis that migration of astrocyte progenitors is dependent upon the proper function of P2Y1Rs and that the P2X7Rs do not play a major role in this process.
4. Discussion
A relevant aspect of the response of neural precursor cells to CNS inflammation is their migration towards lesion sites (Imitola et al., 2004), a feature that is also common of transplanted precursor cells (Ben-Hur et al., 2003b). Microglia and astrocytes are the main sources of pro-inflammatory cytokines in the CNS (Aloisi, 2001; Aloisi et al., 1992; Degousee et al., 2001; Gehrmann et al., 1995; Meme et al., 2007) and some of these inflammatory mediators, such as IL-1β and TNFα, were reported to affect in vitro neural progenitor cell proliferation (Wang et al., 2007; Widera et al., 2006) and migration (Ben-Hur et al., 2003a).
Besides the well established roles of cell adhesion and extracellular molecules on cell migration, other signaling mechanisms have been identified; most prominent is the intracellular Ca2+ fluctuation resultant from ion channel and membrane receptor activation (Komuro and Rakic, 1998; Kumada and Komuro, 2004). Spontaneous intracellular Ca2+ oscillations and waves are prominent events during early CNS development that modulate cell proliferation and migration (Komuro and Rakic, 1998; Owens and Kriegstein, 1998; Catsicas et al., 1998; Pearson et al., 2002; Scemes et al., 2003; Webb et al., 2003; Kumada and Komuro, 2004; Weissman et al., 2004; Agresti et al., 2005a,b). It has been suggested that during these early stages of development, glial cells, by releasing transmitters, would provide the necessary inputs leading to calcium transients and thus influencing neurogenesis (Komuro and Rakic, 1998; Weissman et al., 2004).
Using an in vitro model of neural cell development we have previously shown that migrating neural progenitor cells display spontaneous calcium oscillations that are blunted by bath application of apyrase, an ATP degrading enzyme, and by the specific P2Y1 receptor antagonist MRS-2179, which also reduced progenitor cell migration (Scemes et al., 2003). In this same study, we also showed that exogenous expression of P2Y1R in P2Y1R-deficient cells, rescued their frequency of Ca2+ oscillations and migration rate. Moreover, interference with ATP release machinery was also shown to attenuate the rate of in vitro astrocyte progenitor cell migration and calcium oscillations (Striedinger et al., 2007). These studies, therefore, strongly indicate that spontaneous Ca2+ oscillations mediated by the release of ATP acting on P2Y1 receptors influence the migration of astrocyte progenitor cells.
We here show that conditioned medium derived from activated microglia as well as IL-1β, but not TNFα, reduced the migration distances of astrocyte progenitor cell compared to controls, untreated cells. Evidence is also provided indicating that the reduced in vitro cell migration induced by IL-1β is accompany by a decrease in the frequency of spontaneous Ca2+ oscillations. It is likely that the changes in spontaneous Ca2+ oscillations observed in the present study are related to the altered P2Y1 receptor activity rather than changes in its expression levels or to the ATP release machinery. Evidence favoring such possibility is based on the observations that, in IL-1β treated progenitors, a P2Y1R agonist induced Ca2+ transients of smaller amplitudes than those recorded from untreated cells, and on measurements showing that neither the expression levels of P2Y1 receptors nor the amount of ATP released from progenitor cells were affected by this pro-inflammatory cytokine.
In contrast to the effect of IL-1β on P2Y1 receptor activity, we found that this cytokine enhanced Ca2+ mobilization induced by a P2X7R agonist. Because activation of P2X7 receptors, differently from P2Y1 receptors, leads to prolonged/sustained calcium elevation, it is likely that the reduced frequency of spontaneous Ca2+ oscillations recorded from IL-1 β-treated progenitors is a reflection of the combined changes in activity of these two P2 receptors. However, despite the increased activity of P2X7R, these receptors are unlikely to be involved in the migration of astrocyte progenitors. This is based on our current and previous studies showing that neither the P2X7R agonist BzATP (this study) nor its antagonist KN-62 (Scemes et al., 2003) had an impact on progenitor cell migration. Although the P2X7 receptors have been proposed to be a component of the inflammatory response in astrocytes (Narcisse et al., 2005), and a contributor to pathogenic Ca2+ entry (Yeung et al., 2006), and cell death (Zhang et al., 2005; Wang et al., 2004), further studies are needed to disclose the impact of these ionotropic receptors on neural progenitor cell (patho) physiology.
It is well established that activation of G-coupled surface membrane receptors is the initial step leading to cell polarization and migration triggered by a chemo-attractant. The downstream signal transduction pathway mediating cell motility and migration involves members of the Rho family of small GTPases including Rho, Rac and Cdc42, which are involved in cytoskeletal remodeling (Bokoch, 2005; Barber and Welch, 2006). Although nothing is known about the role of P2R on the triggering of small GTPAses during astrocyte progenitor cell migration, at least in platelet, it has been shown that stimulation of P2Y1R leads to rapid activation of Rac through a Gq and Ca2+-dependent pathway (Soulet et al., 2005). Given that Rac is primarily involved in lamelipodia protrusion at the leading edge (Bokoch, 2005; Barber and Welch, 2006), reduced activity of P2Y1R is expected to attenuate cell migration. Clearly, further studies are necessary to evaluate such hypothesis and to elucidate whether the deactivating effect of IL-1β on Rho GTPase seen in mature astrocytes (John et al., 2004) is also effective in reducing astrocyte progenitor cell migration, independently of P2Y1R. Nevertheless, because the effects of IL-1β on the activity of P2 receptors reported in the present study are very similar to what has been previously reported for mature astrocytes, i.e., decreased response to agonists of P2Y1 receptors (Scemes, 2008) and increased response of P2X7 receptors (Narcisse et al., 2005), it is likely that astrocyte progenitors acquire in early stages of CNS development, the ability to fully respond to IL-1β.
In summary, the present study indicates that IL-1β is a potent negative modulator of calcium signaling and cell migration in astrocyte progenitors, two important processes for proper CNS development and repair mechanisms during pathological conditions involving neuroinflammation.
Acknowledgments
We thank Dr. Kostantin Dobrenis, Department of Neuroscience at Albert Einstein College of Medicine, for providing the microglial cultures, Dr. Meng-Liang Zhao, at the Albert Einstein CFAR Immunology and Pathology Core (P50 AI051519), for the assistance with the ELISA assay, and Miss Melissa Aleksey for technical assistance with the neurosphere cultures. This work was supported by the National Multiple Sclerosis Society (PP1019) and NIH (NS052245) to ES.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Agresti C, Meomartini ME, Amadio S, Ambrosini E, Volonte C, Aloisi F, Visentin S. ATP regulates oligodendrocyte progenitor migration, proliferation, and differentiation: involvement of metabotropic P2 receptors. Brain Res Brain Res Rev. 2005a;48:157–165. doi: 10.1016/j.brainresrev.2004.12.005. [DOI] [PubMed] [Google Scholar]
- Agresti C, Meomartini ME, Amadio S, Ambrosini E, Serafini B, Franchini L, Volonte C, Aloisi F, Visentin S. Metabotropic P2 receptor activation regulates oligodendrocyte progenitor migration and development. Glia. 2005b;50:132–144. doi: 10.1002/glia.20160. [DOI] [PubMed] [Google Scholar]
- Allan SM, Rothwell NJ. Cytokines and acute neurodegeneration. Nat Rev Neurosci. 2001;2:734–744. doi: 10.1038/35094583. [DOI] [PubMed] [Google Scholar]
- Aloisi F, Penna G, Cerase J, Menéndez Iglesias B, Adorini L. IL-12 production by central nervous system microglia is inhibited by astrocytes. J Immunol. 1997;159:1604–1612. [PubMed] [Google Scholar]
- Aloisi F. Immune function of microglia. Glia. 2001;36:165–179. doi: 10.1002/glia.1106. [DOI] [PubMed] [Google Scholar]
- Barber MA, Welch HCE. PI3K and RAC signaling in leukocyte and cancer cell migration. Bull Cancer. 2006;93:E44–52. [PubMed] [Google Scholar]
- Ben-Hur T, Ben-Menachem O, Furer V, Einstein O, Mizrachi-Kol R, Grigoriadis N. Effects of proinflammatory cytokines on the growth, fate, and motility of multipotential neural precursor cells. Mol Cell Neurosci. 2003a;24:623–631. doi: 10.1016/s1044-7431(03)00218-5. [DOI] [PubMed] [Google Scholar]
- Ben-Hur T, Einstein O, Mizrachi-Kol R, Ben-Menachem O, Reinhartz E, Karussis D, Abramsky O. Transplanted multipotential neural precursor cells migrate into the inflamed white matter in response to experimental autoimmune encephalomyelitis. Glia. 2003b;41:73–80. doi: 10.1002/glia.10159. [DOI] [PubMed] [Google Scholar]
- Bokoch GM. Regulation of innate immunity by Rho GTPases. Trends Cell Biol. 2006;15:163–171. doi: 10.1016/j.tcb.2005.01.002. [DOI] [PubMed] [Google Scholar]
- Catsicas M, Bonness V, Becker D, Mobbs P. Spontaneous Ca2+ transients and their transmission in the developing chick retina. Curr Biol. 1998;8:283–286. doi: 10.1016/s0960-9822(98)70110-1. [DOI] [PubMed] [Google Scholar]
- Degousee N, Stefanski E, Lindsay TF, Ford DA, Shahani R, Andrews CA, Thuerauf DJ, Glembotski CC, Nevalainen TJ, Tischfield J, Rubin BB. p38 MAPK regulates group IIa phospholipase A2 expression in interleukin-1beta -stimulated rat neonatal cardiomyocytes. J Biol Chem. 2001;276:43842–43849. doi: 10.1074/jbc.M101516200. [DOI] [PubMed] [Google Scholar]
- Dobrenis K, Chang HY, Pina-Benabou MH, Woodroffe A, Lee SC, Rozental R, Spray DC, Scemes E. Human and mouse microglia express connexin36, and functional gap junctions are formed between rodent microglia and neurons. J Neurosci Res. 2005;82:306–315. doi: 10.1002/jnr.20650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fawcett JW, Asher RA. The glial scar and central nervous system repair. Brain Res Bull. 1999;49:377–391. doi: 10.1016/s0361-9230(99)00072-6. [DOI] [PubMed] [Google Scholar]
- Fok-Seang J, DiProspero NA, Meiners S, Muir E, Fawcett JW. Cytokine-induced changes in the ability of astrocytes to support migration of oligodendrocyte precursors and axon growth. Eur J Neurosci. 1998;10:2400–2415. doi: 10.1046/j.1460-9568.1998.00251.x. [DOI] [PubMed] [Google Scholar]
- Gehrmann J, Matsumoto Y, Kreutzberg GW. Microglia: intrinsic immuneffector cell of the brain. Brain Research Reviews. 1995;20:269–287. doi: 10.1016/0165-0173(94)00015-h. [DOI] [PubMed] [Google Scholar]
- Grynkiewicz G, Poenie M, Tsien RY. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J Biol Chem. 1985;260:3440–3450. [PubMed] [Google Scholar]
- Imitola J, Raddassi K, Park KI, Mueller FJ, Nieto M, Teng YD, Frenkel D, Li J, Sidman RL, Walsh CA, Snyder EY, Khoury SJ. Directed migration of neural stem cells to sites of CNS injury by the stromal cell-derived factor 1alpha/CXC chemokine receptor 4 pathway. Proc Natl Acad Sci U S A. 2004;101:18117–18122. doi: 10.1073/pnas.0408258102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- John GR, Scemes E, Suadicani SO, Liu JS, Charles PC, Lee SC, Spray DC, Brosnan CF. IL-1beta differentially regulates calcium wave propagation between primary human fetal astrocytes via pathways involving P2 receptors and gap junction channels. Proc Natl Acad Sci U S A. 1999;96:11613–11618. doi: 10.1073/pnas.96.20.11613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- John GR, Chen L, Rivieccio MA, Melendez-Vasquez CV, Hartley A, Brosnan CF. Interleukin-1beta induces a reactive astroglial phenotype via deactivation of the Rho GTPase-Rock axis. J Neurosci. 2004;24:2837–45. doi: 10.1523/JNEUROSCI.4789-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly S, Bliss TM, Shah AK, Sun GH, Ma M, Foo WC, Masel J, Yenari MA, Weissman IL, Uchida N, Palmer T, Steinberg GK. Transplanted human fetal neural stem cells survive, migrate, and differentiate in ischemic rat cerebral cortex. Proc Natl Acad Sci U S A. 2004;101:11839–11844. doi: 10.1073/pnas.0404474101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kokaia Z, Lindvall O. Neurogenesis after ischaemic brain insults. Curr Opin Neurobiol. 2003;13:127–132. doi: 10.1016/s0959-4388(03)00017-5. [DOI] [PubMed] [Google Scholar]
- Komuro H, Kumada T. Ca2+ transients control CNS neuronal migration. Cell Calcium. 2005;37:387–393. doi: 10.1016/j.ceca.2005.01.006. [DOI] [PubMed] [Google Scholar]
- Komuro H, Rakic P. Orchestration of neuronal migration by activity of ion channels, neurotransmitter receptors, and intracellular Ca2+ fluctuations. J Neurobiol. 1998;37:110–130. [PubMed] [Google Scholar]
- Kumada T, Komuro H. Completion of neuronal migration regulated by loss of Ca2+ transients. Proc Natl Acad Sci U S A. 2004;101:8479–8484. doi: 10.1073/pnas.0401000101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y, Rao MS. Glial progenitors in the CNS and possible lineage relationships among them. Biol Cell. 2004;96:279–290. doi: 10.1016/j.biolcel.2004.02.001. [DOI] [PubMed] [Google Scholar]
- Liu Y, Wu Y, Lee JC, Xue H, Pevny LH, Kaprielian Z, Rao MS. Oligodendrocyte and astrocyte development in rodents: an in situ and immunohistological analysis during embryonic development. Glia. 2002;40:25–43. doi: 10.1002/glia.10111. [DOI] [PubMed] [Google Scholar]
- Même W, Calvo CF, Froger N, Ezan P, Amigou E, Koulakoff A, Giaume C. Proinflammatory cytokines released from microglia inhibit gap junctions in astrocytes: potentiation by beta-amyloid. FASEB J. 2006;20:494–496. doi: 10.1096/fj.05-4297fje. [DOI] [PubMed] [Google Scholar]
- Monje ML, Toda H, Palmer TD. Inflammatory blockade restores adult hippocampal neurogenesis. Science. 2003;302:1760–1765. doi: 10.1126/science.1088417. [DOI] [PubMed] [Google Scholar]
- Narcisse L, Scemes E, Zhao Y, Lee SC, Brosnan CF. The cytokine IL-1beta transiently enhances P2X7 receptor expression and function in human astrocytes. Glia. 2005;49:245–258. doi: 10.1002/glia.20110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owens DF, Kriegstein AR. Patterns of intracellular calcium fluctuation in precursor cells of the neocortical ventricular zone. J Neurosci. 1998;18:5374–5388. doi: 10.1523/JNEUROSCI.18-14-05374.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pearson R, Catsicas M, Becker D, Mobbs P. Purinergic and muscarinic modulation of the cell cycle and calcium signaling in the chick retinal ventricular zone. J Neurosci. 2002;22:7569–7579. doi: 10.1523/JNEUROSCI.22-17-07569.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picard-Riera N, Decker L, Delarasse C, Goude K, Nait-Oumesmar B, Liblau R, Pham-Dinh D, Evercooren AB. Experimental autoimmune encephalomyelitis mobilizes neural progenitors from the subventricular zone to undergo oligodendrogenesis in adult mice. Proc Natl Acad Sci U S A. 2002;99:13211–13216. doi: 10.1073/pnas.192314199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pluchino S, Zanotti L, Rossi B, Brambilla E, Ottoboni L, Salani G, Martinello M, Cattalini A, Bergami A, Furlan R, Comi G, Constantin G, Martino G. Neurosphere-derived multipotent precursors promote neuroprotection by an immunomodulatory mechanism. Nature. 2005;436:266–271. doi: 10.1038/nature03889. [DOI] [PubMed] [Google Scholar]
- Rao MS, Mayer-Proschel M. Glial-restricted precursors are derived from multipotent neuroepithelial stem cells. Dev Biol. 1997;188:48–63. doi: 10.1006/dbio.1997.8597. [DOI] [PubMed] [Google Scholar]
- Scemes E. Modulation of astrocyte P2Y1 receptors by the carboxyl terminal domain of the gap junction protein Cx43. Glia. 2008;56:145–153. doi: 10.1002/glia.20598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scemes E, Duval N, Meda P. Reduced expression of P2Y1 receptors in connexin43-null mice alters calcium signaling and migration of neural progenitor cells. J Neurosci. 2003;23:11444–11452. doi: 10.1523/JNEUROSCI.23-36-11444.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Striedinger K, Meda P, Scemes E. Exocytosis of ATP from astrocyte progenitors modulates spontaneous Ca2+ oscillations and cell migration. Glia. 2007;55:652–662. doi: 10.1002/glia.20494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soulet C, Hechler B, Gratacap MP, Plantavid M, Offermanns S, Gachet C, Payrastre B. A differential role of platelet ADP receptors P2Y1 and P2Y12 in Rac activation. J Thromb Haemost. 2005;3:2296–2306. doi: 10.1111/j.1538-7836.2005.01588.x. [DOI] [PubMed] [Google Scholar]
- Wang X, Arcuino G, Takano T, Lin J, Peng WG, Wan P, Li P, Xu Q, Liu QS, Goldman SA, Nedergaard M. P2X7 receptor inhibition improves recovery after spinal cord injury. Nat Med. 2004;10:821–827. doi: 10.1038/nm1082. [DOI] [PubMed] [Google Scholar]
- Webb SE, Miller AL. Calcium signalling during embryonic development. Nat Rev Mol Cell Biol. 2003;4:539–551. doi: 10.1038/nrm1149. [DOI] [PubMed] [Google Scholar]
- Weissman TA, Riquelme PA, Ivic L, Flint AC, Kriegstein AR. Calcium waves propagate through radial glial cells and modulate proliferation in the developing neocortex. Neuron. 2004;43:647–661. doi: 10.1016/j.neuron.2004.08.015. [DOI] [PubMed] [Google Scholar]
- Widera D, Mikenberg I, Elvers M, Kaltschmidt C, Kaltschmidt B. Tumor necrosis factor alpha triggers proliferation of adult neural stem cells via IKK/NF-kappaB signaling. BMC Neurosci. 2006;7:64. doi: 10.1186/1471-2202-7-64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeung D, Zablocki K, Lien CF, Jiang T, Arkle S, Brutkowski W, Brown J, Lochmuller H, Simon J, Barnard EA, Gorecki DC. Increased susceptibility to ATP via alteration of P2X receptor function in dystrophic mdx mouse muscle cells. FASEB J. 2006;20:610–620. doi: 10.1096/fj.05-4022com. [DOI] [PubMed] [Google Scholar]
- Zhang X, Zhang M, Laties AM, Mitchell CH. Stimulation of P2X7 receptors elevates Ca2+ and kills retinal ganglion cells. Invest Ophthalmol Vis Sci. 2005;46:2183–2191. doi: 10.1167/iovs.05-0052. [DOI] [PubMed] [Google Scholar]