

Abstract
Leptin is an important regulator of appetite and energy expenditure in adulthood, although its role as a nutritional signal in the control of growth and metabolism before birth is poorly understood. This study investigated the effects of leptin on growth, carbohydrate metabolism and insulin signalling in fetal sheep. Crown–rump length-measuring devices and vascular catheters were implanted in 12 sheep fetuses at 105–110 days of gestation (term 145 ± 2 days). The fetuses were infused i.v. either with saline (0.9% NaCl; n = 6) or recombinant ovine leptin (0.5–1.0 mg kg−1 day−1; n = 6) for 5 days from 125 to 130 days when they were humanely killed and tissues collected. Leptin receptor mRNA and protein were expressed in fetal liver, skeletal muscle and perirenal adipose tissue. Throughout infusion, plasma leptin in the leptin-infused fetuses was 3- to 5-fold higher than in the saline-infused fetuses, although plasma concentrations of insulin, glucose, lactate, cortisol, catecholamines and thyroid hormones did not differ between the groups. Leptin infusion did not affect linear skeletal growth or body, placental and organ weights in utero. Hepatic glycogen content and activities of the gluconeogenic enzymes glucose-6-phosphatase and phosphoenolpyruvate carboxykinase in the leptin-infused fetuses were lower than in the saline-infused fetuses by 44, 48 and 36%, respectively; however, there were no differences in hepatic glycogen synthase activity or insulin signalling protein levels. Therefore, before birth, leptin may inhibit endogenous glucose production by the fetal liver when adipose energy stores and transplacental nutrient delivery are sufficient for the metabolic needs of the fetus. These actions of leptin in utero may contribute to the development of neonatal hypoglycaemia in macrosomic babies of diabetic mothers.
In adult animals, leptin has an important role in the control of appetite and energy expenditure. It is primarily secreted by white adipose tissue, and therefore, acts as an endocrine indicator of energy stores to the hypothalamus and other tissues in the coordination of appetite and metabolism with nutrient availability (Ahima & Flier, 2000; Margetic et al. 2002). Leptin has a variety of actions both in the long term control of energy balance and body size and in the short term control of metabolism and other endocrine responses during fasting (Ahima & Flier, 2000; Margetic et al. 2002). The role of leptin as a nutritional signal before birth, however, is less clear when the fetus normally receives a continuous transplacental supply of glucose and other nutrients for metabolism and growth.
Leptin is detected in the fetal circulation of many animal species, including man and sheep (Jaquet et al. 1998; Forhead et al. 2002), and the widespread abundance of leptin receptors in fetal tissues, especially in developing cartilage and bone, has led to the suggestion that leptin is involved in the control of growth in accordance with nutrient availability in utero (Hoggard et al. 1997; Lin et al. 2000; Buchbinder et al. 2001). In human infants at delivery, umbilical leptin concentration has been correlated with body weight and length, head circumference, ponderal index, adiposity and bone mineral content and density (Hassink et al. 1997; Varvarigou et al. 1999; Javaid et al. 2005). Furthermore, macrosomic offspring of diabetic mothers have high blood leptin concentration at delivery (Persson et al. 1999), although the extent to which leptin acts, in association with insulin, to promote tissue growth in these babies is unknown.
In fetal life, leptin may also act as an important regulator of metabolic pathways, especially in preparation for the nutritional changes that occur at birth. For most of gestation and in normal conditions, the fetus has a sufficient and continuous supply of nutrients and does not undergo endogenous glucose production (Hay et al. 1981). At the time of delivery, however, glycogenolytic and gluconeogenic pathways must be stimulated to maintain a glucose supply in the immediate neonatal period, before nutritive suckling is established. A number of maturational changes occur in the fetal liver near term, including deposition of glycogen and activation of gluconeogenic enzymes, which are induced by the prepartum rise in plasma glucocorticoids (Fowden et al. 1993). In fetal sheep, plasma leptin concentration increases close to term and in response to endogenous and synthetic glucocorticoid treatment (Forhead et al. 2002; O'Connor et al. 2007). Therefore, the rise in plasma leptin observed in the fetus near term may mediate some of the maturational effects of glucocorticoids on hepatic carbohydrate metabolism. Leptin has been shown to influence carbohydrate metabolism in adult life, often interacting with insulin signalling pathways (Harris, 2000; Margetic et al. 2002). In studies on adult tissue, however, data on the effects of leptin on carbohydrate metabolism and insulin signalling pathways are conflicting and appear to depend on the dose and duration of leptin administered, the tissue type and species investigated and whether the study was carried out in vitro or in vivo. Furthermore, the role of leptin as a regulator of carbohydrate metabolism before birth is unknown.
Therefore, the present study investigated the effects of leptin on (a) body and organ growth and (b) aspects of carbohydrate metabolism and tissue insulin signalling pathways in sheep fetuses during late gestation. This was achieved by exogenous i.v. infusion of recombinant ovine leptin to chronically catheterized sheep fetuses for 5 days. The study hypothesized that increased leptin exposure in utero would promote growth of the fetus and activate maturational changes in hepatic glycogen deposition and gluconeogenic enzyme activity.
Methods
Animals
All surgical and experimental procedures were in accordance with the UK Animals (Scientific Procedures) Act 1986 and were approved by the local animal ethics committee. Twelve Welsh Mountain singleton sheep fetuses of known gestational age were used in this study. There were six male and six female fetuses. The ewes were housed in individual pens, and were maintained on 200 g kg−1 concentrates with free access to hay, water and a salt-lick block. Food, but not water, was withheld for 18–24 h before surgery.
Surgical procedures
At between 105 and 110 days of pregnancy (term 145 ± 2 days), the ewes were anaesthetized with halothane (1.5% in O2–N2O) and positive pressure ventilation. Using techniques previously described, a nylon microfilament device was implanted subcutaneously in the fetus to measure crown–rump length (CRL; Fowden et al. 1996), and intravascular catheters were inserted into the femoral artery and vein of the fetus and the femoral artery of the ewe (Comline & Silver, 1972). At the time of surgery, the actual CRL of the fetus was measured. All catheters, including that containing the nylon microfilament, were exteriorized through the flank of the ewe and secured in a plastic bag sutured to the skin. The vascular catheters were flushed daily with heparinized saline solution (100 i.u. heparin (ml 0.9% saline (w/v))−1) from the day after surgery. At surgery, all fetuses were administered 100 mg ampicillin i.v. (Penbritin, Beecham Animal Health, Brentford, UK) and 2 mg gentamycin i.v. (Frangen-100, Biovet, Mullingar, Ireland). The ewes were given antibiotics i.m. (procaine penicillin, Depocillin, Mycofarm, Cambridge, UK) on the day of surgery and for 3 days thereafter.
Experimental procedures
From the day after surgery to the end of the study, increments in CRL were measured daily. The fetuses were divided into two groups with three male and three female fetuses in each group. Starting between 122 and 126 days of gestation (mean 125 days), one group was infused i.v. with saline (0.9% NaCl, n = 6) while the other received recombinant ovine leptin (0.5–1.0 mg kg−1 day−1; n = 6; Protein Laboratories Rehovot, Israel; Gertler et al. 1998) for 5 days. Both treatments were administered at a rate of 3 ml day−1 using portable infusion pumps. Arterial blood from the fetus and ewe (2–3 ml) was collected daily from 2 days before and throughout infusion.
On the fifth day of infusion (mean 130 days of gestation), the fetuses were delivered by Caesarean section under maternal general anaesthesia (20 mg kg−1 sodium pentobarbitone i.v.). After administration of a lethal dose of barbiturate (200 mg kg−1 sodium pentobarbitone) to the ewe and fetus, the fetus was weighed and the integrity of the CRL-measuring device was checked. In all fetuses, actual CRL, body weight, and fore and hind limb lengths, were measured, and a number of tissues including the placenta were collected and weighed. Samples of liver, perirenal adipose tissue (PAT) and skeletal muscle from the hind limb of the fetus were either immediately fixed in 4% paraformaldehyde or frozen in liquid nitrogen and stored at −80°C until analysis.
Biochemical analyses
Blood analyses
All arterial blood samples were analysed immediately for pH and the partial pressure of oxygen 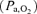 and carbon dioxide
and carbon dioxide 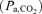 using an ABL330 Radiometer analyser corrected for fetal body temperature, and for haemoglobin content and O2 saturation using an OSM2 Hemoximeter (Radiometer, Copenhagen, Denmark). Blood O2 content (mm) was calculated as haemoglobin content (g dl−1) × O2 saturation (%) × 0.00062, assuming the insignificance of O2 dissolved in plasma. Blood glucose and lactate concentrations were measured using an autoanalyser (2300 Statplus, Yellow Springs, USA).
using an ABL330 Radiometer analyser corrected for fetal body temperature, and for haemoglobin content and O2 saturation using an OSM2 Hemoximeter (Radiometer, Copenhagen, Denmark). Blood O2 content (mm) was calculated as haemoglobin content (g dl−1) × O2 saturation (%) × 0.00062, assuming the insignificance of O2 dissolved in plasma. Blood glucose and lactate concentrations were measured using an autoanalyser (2300 Statplus, Yellow Springs, USA).
After blood gas and metabolite analysis, the samples were placed into EDTA-containing tubes and centrifuged for 5 min at 1000 g and 4°C. The plasma aliquots were stored at −20°C until analysis. Plasma leptin concentration was measured by radioimmunoassay using ovine leptin standards as previously described (Blache et al. 2000); the lower limit of detection was 0.09 ng ml−1 and the interassay coefficient of variation was 5%. Total plasma cortisol concentration was measured by radioimmunoassay (Robinson et al. 1983) where the lower limit of detection was 1.0–1.5 ng ml−1 and the interassay coefficient of variation was 12%. Total plasma T3 and T4 concentrations were also measured by radioimmunoassay using a commercial kit (Fowden & Silver, 1995; ICN Biomedicals, Thame, UK). The lower limits of detection were 0.07 ng ml−1 for T3 and 7.6 ng ml−1 for T4, and the interassay coefficients of variation were 10% for both assays. All radioimmunoassays were validated for use with ovine plasma. Plasma insulin concentration was measured by ELISA (Mercodia, Uppsala, Sweden) using ovine insulin as standards. The lower limit of detection was 0.05 ng ml−1, and the interassay coefficient of variation was 11%. Plasma catecholamine (adrenaline and noradrenaline) concentrations were measured by ELISA (Biosource Europe, Nivelles, Belgium) where all samples were measured within one assay; the lower limits of detection were 5 pg ml−1 and 20 pg ml−1 for adrenaline and noradrenaline, respectively.
Tissue analyses
Gene and protein expression of the leptin receptor were localized in two samples of fetal liver, PAT and skeletal muscle from all animals using in situ hybridization and immunohistochemical methods as previously described (O'Connor et al. 2007). Oligonucleotide probes to all forms of ovine leptin receptor were commercially synthesized (Sigma-Genosys, UK); the antisense and sense probes were 5′-GTT ATG TCT GTG CTC TCA GCC TCA GAG AAC TCA GCC TGC ACT GGT C-3′ and 5′-GAC CAG TGC AGC AGT GCT GAG TTC TCT GAG GCT GAG AGG CAC AGA CAT AAC-3′, respectively (GenBank accession number U63719). A rabbit polyclonal antibody against the long form of the human leptin receptor (Linco Research, USA) was used at a dilution of 1 : 100.
Glycogen content in fetal liver and skeletal muscle, and hepatic activities of the gluconeogenic enzymes glucose-6-phosphatase (G6Pase) and phosphoenolpyruvate carboxykinase (PEPCK) were measured using biochemical methods previously described (Fowden et al. 1993; Franko et al. 2007). Hepatic glycogen synthase (GS) was measured as the rate of incorporation of 3H-uridine diphosphate glucose (UDPG) into glycogen using a method adapted from Thomas et al. (1968). Tissues were homogenized in an extraction buffer at pH 7.5 containing 10 mm Tris, 150 mm potassium fluoride, 15 mm EDTA, 600 mm sucrose, 50 mm mercaptoethanol, 1 mm phenylmethanesulphonyl fluoride, 1 mm benzamidine and 3.5 μg ml−1 leupeptin, and were centrifuged at 600 g for 10 min. The supernatants were incubated with an assay mixture at pH 7.8 containing 50 mm Tris, 20 mm EDTA, 25 mm potassium fluoride, 1% glycogen, 6.5 mm UDPG and 6 μCi ml−13H-UDPG, with or without 10 mm glucose-6-phosphate to determine total and active GS activity, respectively. Samples were incubated at 12.5 mg ml−1 for 15 min (total GS) or 300 mg ml−1 for 30 min (active GS) with assay mixture at 37°C. After incubation, the samples were added to 4 cm2 pieces of filter paper and washed in ice-cold 66% ethanol for 2 h. The filter papers were air-dried and the radioactivity was determined using a β-counter (1216 Rackbeta, LKB Wallac, Turku, Finland). Hepatic GS activity was expressed as nanomoles of UDPG converted to glycogen per minute per milligram of protein, where tissue protein was measured by the Lowry method (Lowry et al. 1951).
Protein levels of the insulin receptor β-subunit (IRβ), insulin-like growth factor I receptor (IGFIR), phosphatidylinositol-3-kinase (PI3K) p85 subunit and phosphokinase C zeta (PKCζ) were determined in samples of fetal liver, skeletal muscle and PAT by Western blot using equal amounts of total protein as previously described (Ozanne et al. 1997). In addition, protein levels of glucose transporter 4 (GLUT4) in skeletal muscle and PAT, and PI3K p110β subunit in liver and PAT, were measured using the same method. In samples of skeletal muscle, protein levels of p-mTOR (mammalian target of rapamyosin), p-S6 kinase, ubiquitin and calpastatin were also determined by Western blot and normalized using β-actin as previously described (Zhu et al. 2004). Polyclonal antibodies to IRβ, IGFIR, p110β and PKCζ were obtained from Santa Cruz Biotechnology (Santa Cruz, USA), to p85 from Upstate Biotechology (Millipore, Billerica, USA), to p-mTOR, p-S6 kinase and ubiquitin from Cell Signalling Technology (Danvers, USA), and to calpastatin from Abcam (Cambridge, UK). In each fetus, mean myofibre diameter was determined from measurements made in 100 fibres as previously described (Zhu et al. 2006).
Statistical analyses
All data are presented as mean values ±s.e.m. Data obtained from the two groups of fetuses on each day of the study were compared by two-way ANOVA with repeated measures followed by Tukey's test. Data obtained from the two groups on the day of surgery, or on the fifth day of infusion, were compared by unpaired t test. Differences where P was < 0.05 were regarded as significant.
Results
Circulating hormone and metabolite concentrations
In the two groups of fetuses, there was no significant difference in any of the circulating parameters measured before the infusion of either saline or leptin (Fig. 1 and Table 1). An i.v. infusion of recombinant ovine leptin to the sheep fetus for 5 days caused a significant increase in leptin concentration in the fetal, but not maternal, circulation (Fig. 1). Compared to the pre-treatment baseline, a significant rise in plasma leptin was observed on all days of the infusion in the leptin-treated fetuses (Fig. 1A). Furthermore, within 24 h of the start of the infusion and on each day thereafter, plasma leptin concentration in the leptin-infused fetuses was 3- to 5-fold higher than in those infused with saline (3.32 ± 0.23 versus 0.77 ± 0.09 ng ml−1 on day 5, P < 0.05, Fig. 1A). There was no significant effect of fetal leptin infusion on plasma leptin concentration in the ewe (Fig. 1B).
Figure 1.
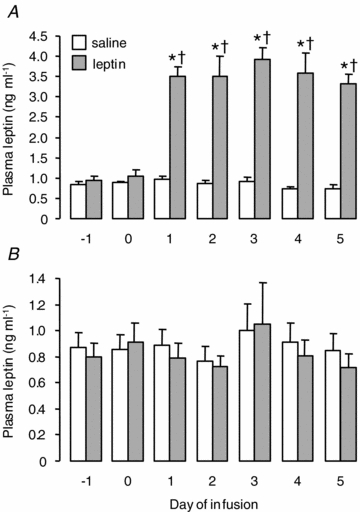
Mean (±s.e.m.) plasma leptin concentration in fetuses (A) and ewes (B) before and during an i.v. infusion of saline (n = 6) or leptin (n = 6) to the fetus for 5 days. *Significant difference from pre-infusion baseline, P < 0.05. †Significant difference from saline-infused fetuses on the same day, P < 0.05.
Table 1.
Mean (±S.E.M.) levels of circulating hormone, metabolite and gas variables in the saline and leptin-infused fetuses before (pre-infusion: mean values for days −1 and 0) and on the fifth day of infusion (day 5)
| Saline (n = 6) | Leptin (n = 6) | |||
|---|---|---|---|---|
| Pre-infusion | Day 5 | Pre-infusion | Day 5 | |
| Blood pH | 7.37 ± 0.01 | 7.37 ± 0.01 | 7.36 ± 0.01 | 7.36 ± 0.01 |
Blood 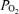 (mmHg) (mmHg) |
20.0 ± 0.2 | 19.7 ± 0.7 | 18.5 ± 1.1 | 18.8 ± 0.7 |
Blood 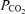 (mmHg) (mmHg) |
52.0 ± 2.0 | 52.8 ± 1.7 | 55.7 ± 0.8 | 55.3 ± 0.9 |
| Blood O2 saturation (%) | 60.0 ± 2.7 | 53.2 ± 3.7 | 54.2 ± 3.7 | 52.6 ± 1.0 |
| Blood haemoglobin (g dl−1) | 10.1 ± 0.4 | 10.1 ± 0.8 | 10.5 ± 0.2 | 10.6 ± 0.3 |
| Blood O2 content (mm) | 3.75 ± 0.12 | 3.28 ± 0.10 | 3.53 ± 0.25 | 3.46 ± 0.09 |
| Plasma glucose (mm) | 0.69 ± 0.07 | 0.66 ± 0.08 | 0.71 ± 0.06 | 0.60 ± 0.07 |
| Plasma lactate (mm) | 1.30 ± 0.12 | 1.36 ± 0.12 | 1.49 ± 0.10 | 1.37 ± 0.06 |
| Plasma insulin (ng ml−1) | 0.19 ± 0.02 | 0.19 ± 0.01 | 0.26 ± 0.03 | 0.17 ± 0.03 |
| Plasma cortisol (ng ml−1) | 9.2 ± 2.7 | 11.9 ± 2.8 | 11.8 ± 2.6 | 11.6 ± 3.6 |
| Plasma T3 (ng ml−1) | 0.19 ± 0.02 | 0.19 ± 0.03 | 0.15 ± 0.03 | 0.16 ± 0.02 |
| Plasma T4 (ng ml−1) | 88.2 ± 7.2 | 89.7 ± 8.7 | 92.1 ± 11.5 | 95.3 ± 8.9 |
| Plasma adrenaline (pg ml−1) | 62.7 ± 15.4 | 89.8 ± 10.8 | 67.1 ± 6.6 | 94.6 ± 23.6 |
| Plasma noradrenaline (pg ml−1) | 712 ± 126 | 623 ± 119 | 1065 ± 271 | 939 ± 176 |
Exogenous leptin infusion for 5 days had no significant effect on blood pH, haemoglobin content, partial pressures of oxygen and carbon dioxide, or oxygen saturation and content in the fetuses (Table 1). Plasma concentrations of glucose, lactate, insulin, cortisol, thyroid hormones (T3 and T4) and catecholamines (adrenaline and noradrenaline) also remained unchanged throughout the infusion of leptin (data obtained pre-infusion and on day 5 are shown in Table 1).
Fetal growth
Actual CRL at the time of surgery for insertion of the growth-measuring device was similar between the two groups of fetuses (saline, 33.3 ± 0.6 cm versus leptin, 35.1 ± 0.9 cm). Over the whole period of the study, there was no significant difference in the rate of CRL increment between the saline and leptin-infused fetuses (Fig. 2). The mean daily rate of growth from 108 to 112 days of gestation, 14 days before the start of the infusion, to tissue collection at 127–130 days of gestation was similar in both groups of fetuses (saline, 4.62 ± 0.31 versus leptin, 4.30 ± 0.35 mm day−1, Fig. 2). Furthermore, no significant differences in mean daily CRL increment were observed between the saline and leptin-infused fetuses before (saline, 4.89 ± 0.41 versus leptin, 4.37 ± 0.48 mm day−1) or during the infusion (saline, 3.80 ± 0.21 versus leptin, 4.10 ± 0.43 mm day−1, Fig. 2). Within each treatment group, the mean rates of daily CRL increment were not significantly different between the pre-infusion and infusion periods.
Figure 2.
Mean (±s.e.m.) cumulative crown–rump length (CRL) in saline and leptin-infused fetuses (n = 6 in each group) from the day after insertion of the CRL-measuring device to the fifth day of infusion. The period of infusion is indicated by the bar.
On the fifth day of infusion, there were no significant differences in fetal body weight, placental weight, the ratio of fetal to placental weights, CRL, limb lengths, or absolute and relative organ weights between the saline and leptin-treated fetuses (Table 2).
Table 2.
Mean (±S.E.M.) fetal body and placental weights, fetal : placental weight ratio, crown–rump and limb lengths, and tissue weights, in the saline and leptin-infused fetuses on the fifth day of infusion
| Saline (n = 6) | Leptin (n = 6) | ||
|---|---|---|---|
| Fetal body weight (FBW, g) | 2533 ± 137 | 2680 ± 151 | |
| Placental weight (PW, g) | 266.8 ± 26.4 | 298.5 ± 21.9 | |
| FBW : PW ratio | 9.90 ± 0.96 | 9.05 ± 0.23 | |
| Crown–rump (cm) | 42.3 ± 0.7 | 43.2 ± 0.9 | |
| Lower fore limb (cm) | 12.7 ± 0.2 | 12.5 ± 0.4 | |
| Middle fore limb (cm) | 10.8 ± 0.3 | 10.3 ± 0.3 | |
| Upper fore limb (cm) | 9.1 ± 0.2 | 8.8 ± 0.4 | |
| Lower hind limb (cm) | 15.5 ± 0.4 | 15.3 ± 0.2 | |
| Middle hind limb (cm) | 13.5 ± 0.3 | 13.3 ± 0.5 | |
| Upper hind limb (cm) | 11.2 ± 0.3 | 10.6 ± 0.4 | |
| Liver | (g) | 62.0 ± 6.5 | 75.9 ± 7.7 |
| (%)a | 2.43 ± 0.15 | 2.81 ± 0.18 | |
| PAT | (g) | 9.3 ± 0.9 | 10.1 ± 0.9 |
| (%) | 0.37 ± 0.05 | 0.38 ± 0.04 | |
| Heart | (g) | 14.0 ± 0.9 | 15.7 ± 2.1 |
| (%) | 0.56 ± 0.04 | 0.57 ± 0.05 | |
| Lungs | (g) | 72.4 ± 3.1 | 63.2 ± 4.1 |
| (%) | 2.90 ± 0.19 | 2.38 ± 0.17 | |
| Kidneys | (g) | 17.8 ± 1.1 | 18.3 ± 1.2 |
| (%) | 0.70 ± 0.03 | 0.67 ± 0.04 | |
| Brain | (g) | 36.9 ± 1.7 | 36.1 ± 1.3 |
| (%) | 1.49 ± 0.05 | 1.35 ± 0.04 | |
| Pancreas | (g) | 2.25 ± 0.12 | 2.48 ± 0.15 |
| (%× 102) | 8.97 ± 0.58 | 9.29 ± 0.31 | |
| Adrenal | (mg) | 234 ± 23 | 252 ± 21 |
| (%× 103) | 9.30 ± 0.87 | 9.11 ± 0.50 | |
PAT, perirenal adipose tissue.
Tissue weights are expressed both as absolute weight and weight relative to fetal body weight (%).
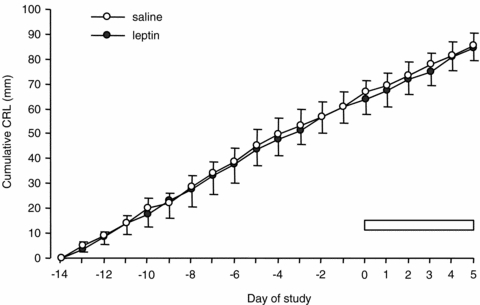
Localization of leptin receptor protein mRNA and protein
At 130 days of gestation, mRNA and protein of the leptin receptor were expressed in fetal liver, skeletal muscle and PAT (Fig. 3). For each tissue type, similar patterns of leptin receptor mRNA and protein expression were observed in samples from all animals.
Figure 3.
Localization of leptin receptor mRNA and protein in fetal PAT, liver and skeletal muscle from a saline-infused fetus at 130 days of gestation. A–C, leptin receptor mRNA localization in fetal PAT, liver and skeletal muscle, respectively; D–F, control hybridization with sense probes in fetal PAT, liver and skeletal muscle, respectively; G–I, leptin receptor protein localization in fetal PAT, liver and skeletal muscle, respectively; J–L, negative control for leptin receptor protein in fetal PAT, liver and skeletal muscle, respectively. Scale bars indicate 50 μm.
Tissue carbohydrate metabolism
On the fifth day of infusion, hepatic glycogen content in the leptin-infused fetuses was significantly lower than in the fetuses infused with saline (P < 0.05, Fig. 4). The 44% decrement in hepatic glycogen content observed in the leptin-treated fetuses was associated with significantly lower activities of the gluconeogenic enzymes, G6Pase (by 48%) and PEPCK (by 36%, P < 0.05 in both cases, Fig. 4). However, in the livers of the two groups of fetuses, there were no significant differences in total GS activity (saline, 15.7 ± 2.2 versus leptin, 12.7 ± 1.1 nmol UDPG min−1 (mg protein)−1), active GS activity (saline, 0.56 ± 0.10 versus leptin, 0.48 ± 0.05 nmol UDPG min−1 (mg protein)−1) or active GS as a percentage of total GS (saline, 3.71 ± 0.47 versus leptin, 3.76 ± 0.26%). On the fifth day of infusion, hepatic protein levels of IRβ, IGFIR, p85, p110β and PKCζ were similar between the two groups of infused fetuses (Fig. 5).
Figure 4.
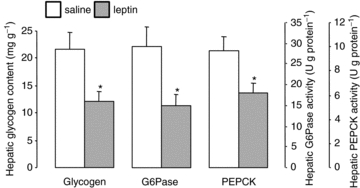
Mean (±s.e.m.) glycogen content, and G6Pase and PEPCK activities, in liver from sheep fetuses on the fifth day of either saline (n = 6) or leptin (n = 6) infusion. *Significant difference from saline-infused fetuses, P < 0.05.
Figure 5.
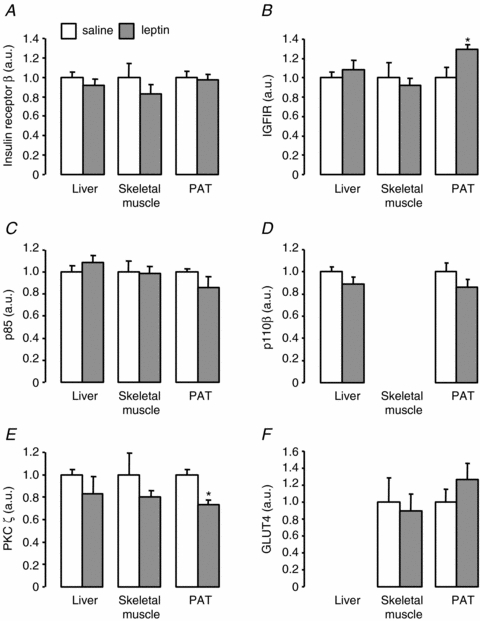
Mean (±s.e.m.) protein levels of insulin receptor β-subunit (A), IGFIR (B), p85 (C), p110β (D), PKCζ (E) and GLUT4 (F) in liver, skeletal muscle and PAT from sheep fetuses on the fifth day of either saline (n = 6) or leptin (n = 6) infusion. *Significant difference from saline-infused fetuses, P < 0.05. a.u., arbitrary units.
In skeletal muscle, there were no significant effects of leptin infusion on glycogen content (saline, 50.56 ± 3.21 versus leptin, 45.78 ± 3.47 mg g−1) or the protein levels of IRβ, IGFIR, p85, PKCζ, GLUT4, p-mTOR, p-S6 kinase, ubiquitin or calpastatin (Figs 5 and 6). Mean myofibre diameter did not differ between the saline and leptin-infused fetuses (10.32 ± 0.83 versus 10.63 ± 0.52 μm).
Figure 6.
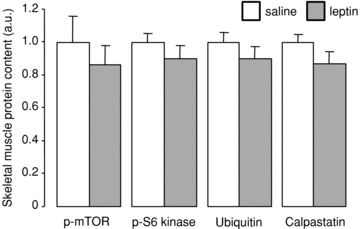
Mean (±s.e.m.) protein levels of p-mTOR, p-S6 kinase, ubiquitin and calpastatin in skeletal muscle from sheep fetuses on the fifth day of either saline (n = 6) or leptin (n = 6) infusion.
In PAT, a significant 30% increase in IGFIR (P < 0.05), and a significant 26% decrease in PKCζ (P < 0.005), were observed in the leptin-infused fetuses compared to those treated with saline (Fig. 5). Leptin infusion had no effect on protein levels of IRβ, p85, p110β or GLUT4 in PAT (Fig. 5).
Discussion
The findings of the present study do not support the hypotheses that leptin stimulates fetal growth or mediates the developmental and glucocorticoid-dependent activation of glycogen storage and gluconeogenic enzyme activity in the fetal liver. However, this is the first study to demonstrate that leptin has actions on carbohydrate metabolism in utero. Administration of recombinant ovine leptin to sheep fetuses caused reductions in hepatic glycogen content and gluconeogenic enzyme activities. These findings suggest that leptin may have a role in suppression of hepatic glucose production in normal conditions when the fetus has adequate energy stores and the transplacental nutrient supply is sufficient to meet its energy requirements.
Leptin and fetal growth
In the present study, leptin infusion over 5 days had no effect on the rate of linear skeletal growth, or body, organ (absolute and relative) and placental weights, in sheep fetuses at 0.85–0.90 of gestation. These findings agree with previous studies in fetal sheep where elevation of plasma leptin concentration by 4- to 5-fold, by an i.v. leptin infusion for 4 days, had no effect on CRL, body or relative organ weights at 140 days of gestation (Yuen et al. 2003). Furthermore, clinical and experimental studies have reported that human and murine neonates with genetic leptin deficiency are morphologically normal (Montague et al. 1997; Mounzih et al. 1998). In the hind limb skeletal muscle of fetal sheep, leptin had no influence on myofibre diameter or on signalling proteins involved in the control of protein anabolism (p-mTOR and p-S6 kinase) or catabolism (calpastatin and ubiquitin). In ob/ob mice, however, a reduction in brain weight and abnormalities in neuronal proteins have been observed at 4 weeks of postnatal life which can be normalized by subsequent leptin replacement (Ahima et al. 1999). The extent to which leptin regulates brain development in species where this largely occurs before birth remains to be established.
In other animal species, leptin derived from the placenta may be an important regulator of placental and, hence, fetal growth. Leptin is synthesized by the human and rodent placenta and is transported across placental cell layers (Hoggard et al. 1997; Masuzaki et al. 1997; Smith & Waddell, 2003; Wyrwoll et al. 2005). Placental leptin concentration is elevated in pregnant mice heterozygous for a mutation in the leptin receptor, and this is associated with an increase in both placental and fetal weights at term (Yamashita et al. 2001). Leptin has also been shown to stimulate activity of the amino acid transporter system A in human placental villous cells in vitro (Jansson et al. 2003). Placental expression of leptin mRNA in the sheep, however, is low during late gestation (Thomas et al. 2001; O'Connor et al. 2007). Receptors for leptin are present in the ovine placenta (Thomas et al. 2001), but the effects of leptin on materno-fetal transfer, and consumption, of nutrients by the placenta are unknown in this species. Leptin may therefore alter the quantity and relative proportion of specific nutrients supplied to the fetus by the placenta but, in the short term, this appears to have little effect on the overall growth of the sheep fetus or its major organs. More detailed analyses of tissue structure and function at the cellular and molecular levels are required to determine fully the role of leptin in feto-placental development.
Leptin and carbohydrate metabolism in utero
The findings that leptin reduced glycogen content, and the activities of G6Pase and PEPCK, in ovine fetal liver are consistent with previous studies in adult rodents. Hepatic glycogen stores are augmented in leptin-deficient ob/ob mice, and depleted in transgenic skinny mice with overexpression of leptin, and in wild-type mice after central or peripheral treatment with leptin (Kamohara et al. 1997; Ogawa et al. 1999; Turner et al. 2005). Furthermore, leptin suppresses PEPCK activity and gluconeogenesis in perfused liver and cultured hepatocytes from adult rats (Ceddia et al. 1999; Anderwald et al. 2002). In contrast, central and peripheral leptin administration has been shown to up-regulate hepatic G6Pase and PEPCK mRNA abundance and to cause an increase in the gluconeogenic, and a decrease in the glycogenolytic, contribution to hepatic glucose production in adult rats (Liu et al. 1998). In addition, leptin increases PEPCK gene expression in adult human hepatocytes in vitro (Cohen et al. 1996).
Overall, the inhibitory effects of leptin on hepatic glycogenic and gluconeogenic pathways observed in the present study are in opposition to the stimulatory actions of glucocorticoids. Leptin is therefore unlikely to be responsible for the developmental and glucocorticoid-induced maturation of carbohydrate metabolism in the fetal liver near term. Preliminary evidence, however, suggests that gene expression of the leptin receptor in ovine fetal liver decreases near term and in response to cortisol treatment (A. J. Forhead, D M. O'Connor & A. L. Fowden, unpublished data). Therefore, glucocorticoids may reduce the inhibitory effects of leptin on hepatic carbohydrate metabolism by decreasing tissue sensitivity to leptin near term.
The mechanisms by which leptin acts to alter hepatic glucogenic capacity in utero are unclear and may be complex. Leptin may influence fetal hepatocytes directly as the presence of leptin receptor mRNA and protein in liver, skeletal muscle and PAT suggests that these tissues are responsive to leptin during late gestation. Suppression of hepatic glycogen content by leptin is likely to occur secondary to the reduction in the activities of the gluconeogenic enzymes, G6Pase and PEPCK, as glycogen deposition in the ovine liver depends primarily on gluconeogenesis both before and after birth (Levitsky et al. 1988). Alternatively, leptin may act indirectly via the autonomic nervous system and/or other endocrine pathways involved in glucoregulation. Previous studies in adult animals have demonstrated that there are interactions between leptin and the PI3K intracellular signalling pathway activated by insulin (Hegyi et al. 2004; Fruhbeck, 2006). Indeed, the inhibitory actions of leptin on gluconeogenic enzyme activity in the ovine fetal liver resemble those observed previously in response to insulin in fetuses of other species (McCormick et al. 1979; Porterfield, 1979). However, in the present study, leptin infusion did not appear to influence the circulating concentration of insulin or the protein levels of the insulin signalling molecules in the fetal liver. Nor did it affect the circulating concentrations of any of the other glucoregulatory hormones measured in this study. Furthermore, intracerebroventricular infusion of recombinant ovine leptin over 5 days in sheep fetuses, which caused a 3-fold increase in plasma leptin, had no effect on the insulin response to a glucose challenge near term (Howe et al. 2002). Taken together, present and previous findings provide little evidence for leptin action via changes in the circulating concentrations of other glucoregulatory hormones. However, there may be changes in the metabolism and/or receptor abundance of these hormones at the cellular level that may account for the actions of leptin on hepatocytes in utero and which remain to be determined.
Clinical implications
The findings of the present study may have significance for the understanding and management of the macrosomic offspring of diabetic mothers. In pregnant women with type 1 or gestational diabetes, poor glucose control leads to hyperinsulinaemia and excessive adipose tissue deposition in the fetus. Offspring of diabetic mothers are at risk of neonatal hypoglycaemia that may be due, in part, to immaturity in hepatic glycogen storage and gluconeogenic enzyme activity at birth. The present results suggest that this may be a consequence of hyperleptinaemia, as well as hyperinsulinaemia, in these babies. Indeed, umbilical leptin concentration in offspring of diabetic mothers is 3- to 4-fold greater than in normal babies (Persson et al. 1999) which is similar to the magnitude of hyperleptinaemia induced experimentally in the present study. Clinically, hyperleptinaemia may arise from insulin-induced leptin synthesis within adipocytes as well as from an overall growth of adipose tissue. Furthermore, although no significant change in PAT mass was observed in response to leptin in the present study, up-regulation of IGFIR gene expression in PAT of the leptin-infused sheep fetuses may be one mechanism that contributes to the growth of fetal adipose tissue in human diabetic pregnancy. Indeed, in fetal sheep, leptin infusion causes a shift in the relative proportions of unilocular and multilocular PAT (Yuen et al. 2003). Further studies are required to determine the extent to which chronic exposure to leptin is responsible for some of the metabolic and growth-promoting actions of insulin in the fetuses of diabetic mothers.
In summary, the findings of the present study do not support the hypothesis that leptin is a growth-promoting hormone in utero, at least not during the latter stage of gestation or at the circulating concentrations investigated. Leptin may, however, act as an endocrine signal of adipose tissue mass to other physiological systems in fetal, as in adult, life. In normal conditions, leptin may be responsible partly for inhibition of endogenous glucose production in utero when adipose energy stores are increasing towards term and transplacental nutrient delivery is sufficient to meet the energy requirements of the fetus. In addition, since leptin concentration in the fetus varies in response to a range of endocrine and metabolic stimuli (Buchbinder et al. 2001; Devaskar et al. 2002; Ducsay et al. 2006; O'Connor et al. 2007), it may also act as a signal of changing nutrient availability more generally, particularly in the control of fetal glucogenic capacity during late gestation. During hyperleptinaemia, however, these actions may become detrimental at birth if glycogenolytic and gluconeogenic capacities are impaired, and may, in turn, contribute to the neonatal hypoglycaemia observed in macrosomic babies of diabetic mothers.
Acknowledgments
The authors are grateful to Margaret Blackberry, University of Western Australia, for the measurement of plasma leptin, and to the members of the Department of Physiology, Development and Neuroscience who provided technical assistance in this study. The study was supported by the Biotechnology and Biological Sciences Research Council.
References
- Ahima RS, Bjorbaek C, Osei S, Flier JS. Regulation of neuronal and glial proteins by leptin: implications for brain development. Endocrinology. 1999;140:2755–2762. doi: 10.1210/endo.140.6.6774. [DOI] [PubMed] [Google Scholar]
- Ahima RS, Flier JS. Leptin. Ann Rev Physiol. 2000;62:413–437. doi: 10.1146/annurev.physiol.62.1.413. [DOI] [PubMed] [Google Scholar]
- Anderwald C, Muller G, Koca G, Furnsinn C, Waldhausl W, Roden M. Short-term leptin-dependent inhibition of hepatic gluconeogenesis is mediated by insulin receptor substrate-2. Mol Endocrinol. 2002;16:1612–1628. doi: 10.1210/mend.16.7.0867. [DOI] [PubMed] [Google Scholar]
- Blache D, Tellam RL, Chagas LM, Blackberry MA, Vercoe PE, Martin GB. Level of nutrition affects leptin concentrations in plasma and cerebrospinal fluid in sheep. J Endocrinol. 2000;165:625–637. doi: 10.1677/joe.0.1650625. [DOI] [PubMed] [Google Scholar]
- Buchbinder A, Lang U, Baker RS, Khoury JC, Mershon J, Clark KE. Leptin in the ovine fetus correlates with fetal and placental size. Am J Obstet Gynecol. 2001;185:786–791. doi: 10.1067/mob.2001.117313. [DOI] [PubMed] [Google Scholar]
- Ceddia RB, Lopes G, Souza HM, Borba-Murad GR, William WN, Bazotte RB, Curi R. Acute effects of leptin on glucose metabolism of in situ perfused livers and isolated hepatocytes. Int J Obes Relat Metab Disord. 1999;23:1207–1212. doi: 10.1038/sj.ijo.0801095. [DOI] [PubMed] [Google Scholar]
- Cohen B, Novick D, Rubinstein M. Modulation of insulin activities by leptin. Science. 1996;274:1185–1188. doi: 10.1126/science.274.5290.1185. [DOI] [PubMed] [Google Scholar]
- Comline RS, Silver M. The composition of foetal and maternal blood during parturition in the ewe. J Physiol. 1972;222:248–256. doi: 10.1113/jphysiol.1972.sp009795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devaskar SU, Anthony R, Hay W. Ontogeny and insulin regulation of fetal ovine white adipose tissue leptin expression. Am J Physiol Regul Integr Comp Physiol. 2002;282:R431–R438. doi: 10.1152/ajpregu.2002.282.2.R431. [DOI] [PubMed] [Google Scholar]
- Ducsay CA, Hyatt K, Mlynarczyk M, Kaushal KM, Myers DA. Long-term hypoxia increases leptin receptors and plasma leptin concentrations in the late-gestation ovine fetus. Am J Physiol Regul Integr Comp Physiol. 2006;291:R1406–R1413. doi: 10.1152/ajpregu.00077.2006. [DOI] [PubMed] [Google Scholar]
- Forhead AJ, Thomas L, Crabtree J, Hoggard N, Gardner DS, Giussani DA, Fowden AL. Plasma leptin concentration in fetal sheep during late gestation: ontogeny and effect of glucocorticoids. Endocrinology. 2002;143:1166–1173. doi: 10.1210/endo.143.4.8762. [DOI] [PubMed] [Google Scholar]
- Fowden AL, Mijovic J, Silver M. The effects of cortisol on hepatic and renal gluconeogenic enzyme activities in the sheep fetus during late gestation. J Endocrinol. 1993;137:213–222. doi: 10.1677/joe.0.1370213. [DOI] [PubMed] [Google Scholar]
- Fowden AL, Silver M. The effects of thyroid hormones on oxygen and glucose metabolism in the sheep fetus during late gestation. J Physiol. 1995;482:203–213. doi: 10.1113/jphysiol.1995.sp020510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fowden AL, Szemere J, Hughes P, Gilmour RS, Forhead AJ. The effects of cortisol on the growth rate of the sheep fetus during late gestation. J Endocrinol. 1996;151:97–105. doi: 10.1677/joe.0.1510097. [DOI] [PubMed] [Google Scholar]
- Franko KL, Giussani DA, Forhead AJ, Fowden AL. Effects of dexamethasone on the glucogenic capacity of fetal, pregnant, and non-pregnant adult sheep. J Endocrinol. 2007;192:67–73. doi: 10.1677/joe.1.07063. [DOI] [PubMed] [Google Scholar]
- Fruhbeck G. Intracellular signalling pathways activated by leptin. Biochem J. 2006;393:7–20. doi: 10.1042/BJ20051578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gertler A, Simmons J, Keisler DH. Large-scale preparation of biologically active recombinant ovine obese protein (leptin) FEBS Lett. 1998;422:137–140. doi: 10.1016/s0014-5793(97)01613-x. [DOI] [PubMed] [Google Scholar]
- Harris RB. Leptin – much more than a satiety signal. Ann Rev Nutr. 2000;20:45–75. doi: 10.1146/annurev.nutr.20.1.45. [DOI] [PubMed] [Google Scholar]
- Hassink SG, de Lancey E, Sheslow DV, Smith-Kirwin SM, O'Connor DM, Considine RV, Opentanova I, Dostal K, Spear ML, Leef K, Ash M, Spitzer AR, Funanage VL. Placental leptin: an important new growth factor in intrauterine and neonatal development? Pediatrics. 1997;100:E1. doi: 10.1542/peds.100.1.e1. [DOI] [PubMed] [Google Scholar]
- Hay WW, Sparks JW, Quissell BJ, Battaglia FC, Meschia G. Simultaneous measurements of umbilical uptake, fetal utilization rate, and fetal turnover rate of glucose. Am J Physiol Endocrinol Metab. 1981;240:E662–E668. doi: 10.1152/ajpendo.1981.240.6.E662. [DOI] [PubMed] [Google Scholar]
- Hegyi K, Fulop K, Kovacs K, Toth S, Falus A. Leptin-induced signal transduction pathways. Cell Biol Int. 2004;28:159–169. doi: 10.1016/j.cellbi.2003.12.003. [DOI] [PubMed] [Google Scholar]
- Hoggard N, Hunter L, Duncan JS, Williams LM, Trayhurn P, Mercer JG. Leptin and leptin receptor mRNA and protein expression in the murine fetus and placenta. Proc Natl Acad Sci U S A. 1997;94:11073–11078. doi: 10.1073/pnas.94.20.11073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howe DC, Gertler A, Challis JRG. The late gestation increase in circulating ACTH and cortisol in the fetal sheep is suppressed by intracerebroventricular infusion of recombinant ovine leptin. J Endocrinol. 2002;174:259–266. doi: 10.1677/joe.0.1740259. [DOI] [PubMed] [Google Scholar]
- Jansson N, Greenwood SL, Johansson BR, Powell TL, Jansson T. Leptin stimulates the activity of the system A amino acid transporter in human placental villous fragments. J Clin Endocrinol Metab. 2003;88:1205–1211. doi: 10.1210/jc.2002-021332. [DOI] [PubMed] [Google Scholar]
- Jaquet D, Leger J, Levy-Marchal C, Oury JF, Czernichow P. Ontogeny of leptin in human fetuses and newborns: effect of intrauterine growth retardation on serum leptin concentrations. J Clin Endocrinol Metab. 1998;83:1243–1246. doi: 10.1210/jcem.83.4.4731. [DOI] [PubMed] [Google Scholar]
- Javaid MK, Godfrey KM, Taylor P, Robinson SM, Crozier SR, Dennison EM, Robinson JS, Breier BR, Arden NK, Cooper C. Umbilical cord leptin predicts bone mass. Calcif Tissue Int. 2005;76:341–347. doi: 10.1007/s00223-004-1128-3. [DOI] [PubMed] [Google Scholar]
- Kamohara S, Burcelin R, Halaas JL, Friedman JM, Charron MJ. Acute stimulation of glucose metabolism in mice by leptin treatment. Nature. 1997;389:374–377. doi: 10.1038/38717. [DOI] [PubMed] [Google Scholar]
- Levitsky LL, Paton JB, Fisher DE. Precursors to glycogen in ovine fetuses. Am J Physiol Endocrinol Metab. 1988;255:E743–E747. doi: 10.1152/ajpendo.1988.255.5.E743. [DOI] [PubMed] [Google Scholar]
- Lin J, Barb CR, Matteri RL, Kraeling RR, Chen X, Meinersmann RJ, Rampacek GB. Long form leptin receptor mRNA expression in the brain, pituitary, and other tissues in the pig. Domest Anim Endocrinol. 2000;19:53–61. doi: 10.1016/s0739-7240(00)00064-3. [DOI] [PubMed] [Google Scholar]
- Liu L, Karkanias GB, Morales JC, Hawkins M, Barzilai N, Wang J, Rossetti L. Intracerebroventricular leptin regulates hepatic but not peripheral glucose fluxes. J Biol Chem. 1998;273:31160–31167. doi: 10.1074/jbc.273.47.31160. [DOI] [PubMed] [Google Scholar]
- Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ. Protein measurement with the folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- McCormick KL, Susa JB, Widness JA, Singer DB, Adamsons K, Schwartz R. Chronic hyperinsulinemia in the fetal rhesus monkey: effects on hepatic enzymes active in lipogenesis and carbohydrate metabolism. Diabetes. 1979;28:1064–1068. doi: 10.2337/diab.28.12.1064. [DOI] [PubMed] [Google Scholar]
- Margetic S, Gazzola C, Pegg GG, Hill RA. Leptin: a review of its peripheral actions and interactions. Int J Obes Relat Metab Disord. 2002;26:1407–1433. doi: 10.1038/sj.ijo.0802142. [DOI] [PubMed] [Google Scholar]
- Masuzaki H, Ogawa Y, Sagawa N, Hosoda K, Matsumoto T, Mise H, Nishimura H, Yoshimasa Y, Tanaka I, Mori T, Nakao K. Nonadipose tissue production of leptin: leptin as a novel placenta-derived hormone in humans. Nat Med. 1997;3:1029–1033. doi: 10.1038/nm0997-1029. [DOI] [PubMed] [Google Scholar]
- Montague CT, Farooqi IS, Whitehead JP, Soos MA, Rau H, Wareham NJ, Sewter CP, Digby JE, Mohammed SN, Hurst JA, Cheetham CH, Earley AR, Barnett AH, Prins JB, O'Rahilly S. Congenital leptin deficiency is associated with severe early-onset obesity in humans. Nature. 1997;387:903–908. doi: 10.1038/43185. [DOI] [PubMed] [Google Scholar]
- Mounzih K, Qiu J, Ewart-Toland A, Chehab FF. Leptin is not necessary for gestation and parturition but regulates maternal nutrition via a leptin resistance state. Endocrinology. 1998;139:5259–5262. doi: 10.1210/endo.139.12.6523. [DOI] [PubMed] [Google Scholar]
- O'Connor DM, Blache D, Hoggard N, Brookes E, Wooding FBP, Fowden AL, Forhead AJ. Developmental control of plasma leptin and adipose leptin mRNA in the ovine fetus during late gestation: role of glucocorticoids and thyroid hormones. Endocrinology. 2007;148:3750–3757. doi: 10.1210/en.2007-0310. [DOI] [PubMed] [Google Scholar]
- Ogawa Y, Masuzaki H, Hosoda K, Aizawa-Abe M, Suga J, Suda M, Ebihara K, Iwai H, Matsuoka N, Satoh N, Odaka H, Kasuga H, Fujisawa Y, Inoue G, Nishimura H, Yoshimasa Y, Nakao K. Increased glucose metabolism and insulin sensitivity in transgenic skinny mice overexpressing leptin. Diabetes. 1999;48:1822–1829. doi: 10.2337/diabetes.48.9.1822. [DOI] [PubMed] [Google Scholar]
- Ozanne SE, Nave BT, Wang CL, Shepherd PR, Prins J, Smith GD. Poor fetal nutrition causes long-term changes in expression of insulin signaling components in adipocytes. Am J Physiol Endocrinol Metab. 1997;273:E46–E51. doi: 10.1152/ajpendo.1997.273.1.E46. [DOI] [PubMed] [Google Scholar]
- Persson B, Westgren M, Celsi G, Nord E, Ortqvist E. Leptin concentrations in cord blood in normal newborn infants and offspring of diabetic mothers. Horm Metab Res. 1999;31:467–471. doi: 10.1055/s-2007-978776. [DOI] [PubMed] [Google Scholar]
- Porterfield SP. The effects of growth hormone, thyroxine and insulin on the activities of reduced nicotinamide adenine dinucleotide phosphate dehydrogenase, glucose-6-phosphatase and glycogen phosphorylase in fetal rat liver. Horm Metab Res. 1979;11:444–448. doi: 10.1055/s-0028-1092757. [DOI] [PubMed] [Google Scholar]
- Robinson PM, Comline RS, Fowden AL, Silver M. Adrenal cortex of fetal lamb: changes after hypophysectomy and effects of Synacthen on cytoarchitecture and secretory activity. Q J Exp Physiol. 1983;68:15–27. doi: 10.1113/expphysiol.1983.sp002697. [DOI] [PubMed] [Google Scholar]
- Smith JT, Waddell BJ. Leptin distribution and metabolism in the pregnant rat: transplacental leptin passage increases in late gestation but is reduced by excess glucocorticoids. Endocrinology. 2003;144:3024–3030. doi: 10.1210/en.2003-0145. [DOI] [PubMed] [Google Scholar]
- Thomas JA, Schlender KK, Larner J. A rapid filter paper assay for UDPglucose-glycogen glucosyltransferase, including an improved biosynthesis of UDP-14C-glucose. Anal Biochem. 1968;25:486–499. doi: 10.1016/0003-2697(68)90127-9. [DOI] [PubMed] [Google Scholar]
- Thomas L, Wallace JM, Aitken RP, Mercer JG, Trayhurn P, Hoggard N. Circulating leptin during ovine pregnancy in relation to maternal nutrition, body composition and pregnancy outcome. J Endocrinol. 2001;169:465–476. doi: 10.1677/joe.0.1690465. [DOI] [PubMed] [Google Scholar]
- Turner SM, Linfoot PA, Neese RA, Hellerstein MK. Sources of plasma glucose and liver glycogen in fasted ob/ob mice. Acta Diabetol. 2005;42:187–193. doi: 10.1007/s00592-005-0201-3. [DOI] [PubMed] [Google Scholar]
- Varvarigou A, Mantzoros CS, Beratis NG. Cord blood leptin concentrations in relation to intrauterine growth. Clin Endocrinol. 1999;50:177–183. doi: 10.1046/j.1365-2265.1999.00630.x. [DOI] [PubMed] [Google Scholar]
- Wyrwoll CS, Mark PJ, Waddell BJ. Directional secretion and transport of leptin and expression of leptin receptor isoforms in human placental BeWo cells. Mol Cell Endocrinol. 2005;241:73–79. doi: 10.1016/j.mce.2005.05.003. [DOI] [PubMed] [Google Scholar]
- Yamashita H, Shao J, Ishizuka T, Klepcyk PJ, Muhlenkamp P, Qiao L, Hoggard N, Friedman JE. Leptin administration prevents spontaneous gestational diabetes in heterozygous Leprdb/+ mice: effects on placental leptin and fetal growth. Endocrinology. 2001;142:2888–2897. doi: 10.1210/endo.142.7.8227. [DOI] [PubMed] [Google Scholar]
- Yuen BSJ, Owens PC, Muhlhausler BS, Roberts CT, Symonds ME, Keisler DH, McFarlane JR, Kauter KG, Evens Y, McMillen IC. Leptin alters the structural and functional characteristics of adipose tissue before birth. FASEB J. 2003;17:1102–1104. doi: 10.1096/fj.02-0756fje. [DOI] [PubMed] [Google Scholar]
- Zhu M-J, Ford SP, Nathanielsz PW, Du M. Effect of maternal nutrient restriction in sheep on the development of fetal skeletal muscle. Biol Reprod. 2004;71:1968–1973. doi: 10.1095/biolreprod.104.034561. [DOI] [PubMed] [Google Scholar]
- Zhu MJ, Ford SP, Means WJ, Hess BW, Nathanielsz PW, Du M. Maternal nutrient restriction affects properties of skeletal muscle in offspring. J Physiol. 2006;575:241–250. doi: 10.1113/jphysiol.2006.112110. [DOI] [PMC free article] [PubMed] [Google Scholar]