

Abstract
In motion standstill, a quickly moving object appears to stand still, and its details are clearly visible. It is proposed that motion standstill can occur when the spatiotemporal resolution of the shape and color systems exceeds that of the motion systems. For moving red-green gratings, the first- and second-order motion systems fail when the grating is isoluminant. The third-order motion system fails when the green/red saturation ratio produces isosalience (equal distinctiveness of red and green). When a variety of high-contrast red-green gratings, with different spatial frequencies and speeds, were made isoluminant and isosalient, the perception of motion standstill was so complete that motion direction judgments were at chance levels. Speed ratings also indicated that, within a narrow range of luminance contrasts and green/red saturation ratios, moving stimuli were perceived as absolutely motionless. The results provide further evidence that isoluminant color motion is perceived only by the third-order motion system, and they have profound implications for the nature of shape and color perception.
Categories of Motion Perception.
Three qualitatively different categories are traditionally distinguished in the perception of moving objects: (i) For extremely fast movements, direction of movement may be obvious, although the moving object is perceived only as a blur. What is critical here is not the absolute speed of motion [degrees (deg)/sec] but the speed relative to the size of details within the object (deg/cycle). This ratio defines the temporal wavelength—how long a detail is present at a point in visual space before it is replaced by a new one. It is more common to give the reciprocal of wavelength, the temporal frequency. Thus, we say that, when the temporal frequency of the physical movement exceeds the temporal resolution of the human visual system, only a blur is perceived. (ii) When the temporal frequency of the physical movement is well within the resolution of the human visual system, smooth movement of a single object is perceived. (iii) When the motion signal is below the detectability of the human motion system, e.g., as in the slow movement of the moon across the sky, no motion or a stationary object is perceived. Motion standstill is a fourth, little studied category of movement perception. It is observed not for extremely slow movements but for motions that are too fast or too weak for the motion system to resolve, in which the moving object appears to stand still.
Motion Standstill in Stereoscopic Displays.
Motion standstill was first reported by Julesz and Payne (1) in viewing a dynamic, random-dot stereogram. In stereoscopic viewing, a vertical bar was made to appear in front of a flat background, and it alternated between two positions, one left, the other to the right. At alternation rates of one or two cycles per second, observers perceived apparent movement between the two locations. At alternations of 12 Hz, observers perceived two standing bars and no apparent movement. At alternation rates of ≈6 Hz, only one bar was perceived, and it did not appear to move. The authors called this “motion standstill”; they did not have an explanation.
Motion Standstill in Isoluminant Displays.
In viewing isoluminant color gratings, slower motion than the actual physical motion is commonly reported. More to the point, there have been occasional reports of motion standstill (2–6), in which the moving grating appeared to stand still for a time and subsequently was seen at a new location, without any perception of movement.
Perceptual standstill of moving colored gratings is an astounding phenomenon because a high-contrast, relatively rapidly moving display appears to stand still under circumstances that would seem optimal for the perception of movement. In Julesz's display, excessive speed seemed to cause standstill, but this is not necessary for standstill in the movement of isoluminant chromatic gratings. Despite its intrinsic interest, motion standstill has not been parametrically studied in chromatic displays, owing perhaps to the haphazard circumstances that have produced it thus far.
Previous attempts to explain motion standstill in chromatic displays were concerned primarily with the slow response of the color system. The first theory to deal with motion standstill more generally (7) proposed that motion standstill occurred when a motion signal that was visible only to the third-order motion system was so reduced in amplitude that third-order motion failed. This theory requires a definition of third-order motion.
Motion Systems: Photons, Features, Salience.
A motion system is a computation, presumably carried out in a brain nucleus, that takes as input a space–time representation of the visual field and produces a motion flowfield as output. In a motion flowfield, each space–time neighborhood of the visual field is represented by a vector that indicates the direction and magnitude of motion in that neighborhood.
The input to the first-order motion system is a simple function of the luminance in the neighborhood of each point (8–11). The input to the second-order motion system is a representation of the texture in the neighborhood of each point (12). Thus, one can describe the first-order system as computing the motion of photons and the second-order system as computing the motion of features.
At a higher level, a visual image can be segmented into areas perceived as figure, and other areas perceived as background, or simply “ground.” Because figure–ground is not an absolute dichotomy, it is more useful to define a variable, which we call salience. For specificity, we let salience take the value 1.0 for figure, 0 for ground, intermediate values for ambiguous regions, and values >1.0 for selectively attended figures. The third-order motion system takes salience as input, and its flowfield output therefore represents the motion of areas of the visual field designated as figure (13–16).
Concurrent Motion and Shape Computations.
Both succeed.
The modular organization of visual computation in the brain has long been recognized (17–21). All visual information is initially received by the visual receptors, rod and cones. Subsequently, color, motion, shape, and perhaps stereo depth and texture are segregated and processed separately at intermediate stages. Eventually, the results of these intermediate computations are recombined to form complex perceptions.
A good example to illustrate conflict between motion and shape systems is pedestaled motion, in which a small-amplitude moving sinewave grating is superimposed on a large-amplitude stationary grating (7, 9, 14, 22). The first-order motion system can report the direction of motion of the moving grating equally well whether the stationary grating is present or absent.§ At temporal frequencies >8 Hz, the temporal resolution of the third-order motion system is exceeded, and it reports “no motion” (14). The shape system reports the pedestal grating. Here, there are three relevant outputs from shape and from motion modules: the first-order motion system reports linear motion, the third-order motion system reports no-motion, and the shape system reports a grating. The resulting perception combines all three outputs including the conflict: motionless grating plus left-to-right motion, which is not attached to any object (11).
Motion Standstill: Concurrent Motion and Shape Computations.
Motion fails, shape succeeds.
We propose a simple theory of motion standstill that has wide-reaching implications, not merely for motion systems, but also for the mechanisms of shape perception. A remarkable fact of human shape perception is that it succeeds in spite of large image movements on the retina. For small image movements relative to the shape being perceived, success of shape perception is not surprising. However, the jitter of the retinal image during normal walking and similar activities is very large relative to the resolution of human vision (24). The shape system has evolved to extract shape in spite of random image movement. When the shape system succeeds, its output is a shape or pattern. When the motion systems also succeed, the perception is that of a single moving object of a specified shape. However, when the motion systems fail, and the shape system still reports a shape, we have the percept of motion standstill—the spatiotemporal resolution of the shape system exceeds the resolution of the motion systems.
A Snapshot Process for Extracting Shape Invariants from Moving Images.
Our proposed explanation of motion standstill requires an answer to a fundamental question: How can a shape system report a static invariant shape when the image is moving? Indeed, the experiments herein will demonstrate that motion standstill can occur for stimuli moving with temporal frequencies of up to 5 Hz, and we have informally observed standstill at higher temporal frequencies. Suppose the shape system resolved a grating moving at 5 Hz by taking snapshots of motion during short intervals of time. (We use the word “snapshot” rather than the more conventional “sample” to indicate that the input is averaged over a period of time—the snapshot duration—rather than sampled instantaneously.) What temporal resolution would such a process require?
To represent a sinewave grating, a snapshot that averaged over as much as 1/2 of a temporal cycle would involve a great deal of cancellation of one part of the grating by another, depending on the phase of the grating. For example, averaging over 1/2 of a cycle yields a zero output for certain phases. For a faithful representation of the stimulus, averaging over 1/4 of a cycle or less seems desirable. Thus, snapshots by the spatial system of a 5-Hz moving grating should have an effective duration of 50 msec or less. If snapshots were consecutive, there would be 20 snapshots during a 1-sec display; there would be >20 if there were temporal overlap. The simple average of 20 snapshots of a moving stimulus in 20 different phases would not be useful. We must assume that the shape system has evolved a better combination rule than averaging, perhaps a winner-take-all rule, with hysteresis (25) to perpetuate a winner until there is a significant input perturbation. Another, compatible, possibility is that the shape system represents primarily (invariant) relational properties. Hubel and Wiesel (26) originally proposed that complex cells might be involved in the extraction of features independent of position. The snapshot model is an elaboration of how such a complex process might begin. These suggestions are presented so that the reader may appreciate what mechanisms might enable the shape (and color) systems to occasionally exceed the motion system in temporal resolution.
Outline.
From an empirical point of view, motion standstill offers a remarkable opportunity to study the temporal resolution of the shape system—the ability of the shape system to report the presence of a shape—when the motion system (which would normally report the movement of this shape) has been silenced. Here, we concentrate on a particular class of moving displays, isoluminant chromatic gratings, for which a wide range of perceptual qualities, from motion standstill to smooth motion, has been reported (2–6, 27–45). Specifically, the displays are gratings composed of alternating red and green stripes in which the component colors have been so carefully equated in luminance that they stimulate only color-sensitive motion mechanisms but not luminance-sensitive motion mechanisms. We show how, by manipulating luminance and salience, it is possible to produce motion standstill in such isoluminant gratings under a variety of conditions.
Salience Modulation.
In a previous publication (7), we showed that the relevant variable for determining the quality of isoluminant motion is salience. Salience refers to the tendency of an area to be perceived as figure rather than ground, and it can be precisely measured (13, 16). For the isoluminant red-green gratings used here, salience is proportional to the color saturation of red and green areas, relative to the neutral gray background. After isoluminance between red and green areas was established, the relative salience of the red and the green areas was manipulated by keeping the saturation of the red areas constant and changing the saturation of the green areas. In preliminary observations (7), we observed that, by increasing the ratio of green/red contrast from very small to very large values, we were able to produce high-contrast, easily visible, isoluminant displays that exhibited the full gamut of motion responses: from normal, easily perceived motion at small ratios to motion standstill at red-green ratios near to 1, and then again to easily perceived motion for large green/red ratios. Here, we formalize these observations for the motion of red-green gratings.
In Experiment 1, observers make objective motion–direction judgments (which may be correct or incorrect) to determine the conditions for motion standstill. Standstill is indicated by the inability to judge motion direction (because the grating appears to be motionless). In Experiment 2, observers make subjective speed estimates, matching the speeds of isoluminant red-green gratings to speeds of ordinary black-white luminance gratings. The luminance and saturation parameters that define red and green areas of the color grating are manipulated so that red and green are simultaneously isoluminant and isosalient and thereby produce zero apparent speed (motion standstill). We measure how perceived speed increases as the red-green grating departs from isoluminance and isosalience.
Because of the extreme sensitivity of foveal color vision, even tiny shifts in retinal sensitivity to color saturation and luminance made it difficult to combine data from successive sessions although each individual session was consistent. Therefore, the observations were made from 3.5 to 4.7 deg parafoveally.
Methods
Apparatus.
The experiments were controlled by a 7500/100 PowerPC Macintosh computer running matlab programs based on PsychToolBox (46). A special 30-bit Radius Thunder 1600/30 video graphics card was used to display images on an Apple 1710 multisync color monitor at a refresh rate of 60 frames/sec. The 30-bit resolution (1,024 intensity levels for each of red, green, and blue) is critical for accurate control of luminance and color in these experiments. By using a Tektronix J17 photometer with a J1820 chromaticity head, the monitor was calibrated with standard procedures (47) to generate the initial directions and linear scales of the three cardinal axis in the Derrington-Lennie-Krauskopf (DLK) color space (48, 49). Subsequent calibration is described below.
Stimuli.
All of the chromatic gratings were modulated sinusoidally along the long–medium (L-M) wavelength axis in the DLK color space, with a gray background at chromaticity (for a CIE standard observer) (x, y) = (0.296, 0.314) and luminance of 25.4 cd/m2. On this L-M axis, the chromaticity of 0.08 (red) is (x, y) = (0.333, 0.301); the chromaticity of −0.08 (green) is (x, y) = (0.241, 0.358). To minimize chromatic aberration (50–52), all of the gratings were horizontal (Fig. 1a).
Figure 1.
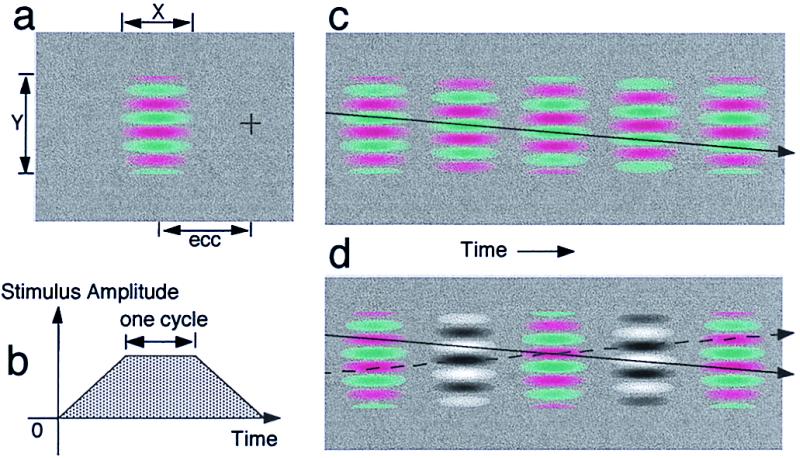
Experimental and calibration stimuli. (a) Viewing arrangement. Fixation cross (+), eccentricity in deg (ecc), width and height (x, y). (b) The temporal display sequence, showing the on-ramp, steady display, and off-ramp of the moving grating; each component is one full period (4 frames). (c) Five frames of a moving grating. (d) Five frames of the calibration display. Frames 2 and 4 are luminance amplifier frames.
In both Experiment 1 and 2, the moving stimuli were displayed at an eccentricity, ecc, defined as the distance in degrees of visual angle between the center of the grating and the fixation point (Fig. 1a). When eccentricity = 0, the display is centered at fovea. One temporal cycle (T sec) of a moving sinewave grating consisted of four frames with a consistent phase shift of 90 deg between successive frames (Fig. 1c). The temporal frequency of a moving display is defined as 1/T Hz. Moving stimuli of different temporal frequencies were generated by varying frame duration. The stimuli were ramped linearly in time (Fig. 1b) at both onset and offset to avoid auxiliary temporal frequencies. The duration of the onset ramp, plateau, and offset ramp was one temporal cycle. To avoid edge effects, the stimuli were also Gaussian-windowed in space, with standard deviations σx and σy in the horizontal and vertical dimensions. The spatial frequencies in different conditions were produced by changing the period of the grating on the screen and the viewing distance (see individual experiments for details).
Motion Calibration.
To ensure that all of the moving “isoluminant” chromatic displays only activate color-sensitive motion mechanisms, not luminance-sensitive (first-order) motion mechanisms, we performed calibration in two phases: first, we followed the standard procedures to define the isoluminant L-M axis in the DLK color space (47) for static displays; second, for each particular isoluminant display, we applied a dynamic calibration procedure (53, 54) to remove any residual luminance contamination.
Fig. 1d depicts the motion calibration display. For a given candidate moving isoluminant display, the calibration display is constructed by replacing color gratings in the even frames with luminance sinewave gratings (the amplifier gratings). Except for being modulated along the L+M (luminance) direction in the DLK color space, these luminance sinewave gratings were similar to the corresponding, replaced color gratings: i.e., same contrast (in DLK space), same phase (with red ≈ white, green ≈ black), same spatial frequency, same Gaussian windowing, and same ramp.
Typically, the amplitudes of the amplifier luminance sinewaves were 3–4× their own motion thresholds. Second, small amounts of “correction” luminance sinewave gratings were added to the color gratings. Third, the amplitude of the correction luminance sinewave grating was varied (in the method of constant stimuli) to determine the amount of luminance correction for the particular moving chromatic display—the amount of luminance added to the color gratings such that the observer is at chance in judging its direction of motion when the calibration display moves either up or down randomly across trials. For a given isoluminant display, two different luminance sinewave phases (in-phase: white aligned with red, black with green; or out-of-phase: white aligned with green, black with red) and five different luminance amplitudes were used. Forty motion trials were conducted at each phase and amplitude.
Definition of Isoluminance.
There is random variation at every level of the visual system, so that what is isoluminant for one neuron is not perfectly isoluminant for any other. However, the calibration procedure is designed so that all deviations from isoluminance exactly cancel; their net contribution to the decision “did it move up or down?” is zero. Of course, zero contamination is impossible to achieve, but because of the amplifier principle in motion perception (54), luminance contamination can be made very small. In our displays, the strength of apparent motion is proportional to the product of the modulation amplitude of even and odd frames. When the even frames are 3× threshold, a luminance modulation 1/3 of threshold in the odd frames will be visible (because 3 × 1/3 = 1). Thereby, the maximum possible luminance contamination is reduced to <1/3 of threshold amplitude.
In principle, isoluminance is a property of a particular display condition, viewed by a particular observer. When any factor changes (i.e., saturation, luminance, temporal frequency, fixation, display geometry, or viewing distance), a new calibration must be carried out. Because first-order (luminance) motion is a monocular computation (15), binocular viewing would require separate luminance calibrations for each eye. Therefore, viewing was monocular throughout. The importance of context in motion calibration cannot be overemphasized: The calibration procedure must be as close as possible to the actual isoluminant displays. Our calibration procedures guarantee that each chromatic motion display is devoid of perceivable luminance contamination.
Observers.
Three naive observers and the second author participated in various parts of the experiments. All of the observers had corrected-to-normal vision. Viewing was monocular. Before each experimental session, the observer adapted to the background for ≈3 minutes.
Experiment 1: Relative Salience Determines Motion Strength
Procedure.
The purpose of this experiment is to test the prediction that the relative salience between colored areas determines motion strength. Keeping the entire grating at isoluminance, the relative salience between the red and green areas was varied by keeping the saturation of red areas constant and increasing or decreasing the saturation of the green areas (Fig. 2). A typical display is shown in Fig. 1a.
Figure 2.

Five stimuli illustrating different |G|/|R| green-red saturation ratios.
The experiment was conducted in four parts, differing in terms of the spatial and temporal frequency of the sinewave gratings and the viewing eccentricity. The spatial and temporal frequencies and eccentricity of the conditions are 0.875 cycles per degree (c/d), 4 Hz, 4.7 deg; 0.5 c/d, 2 Hz, 3.5 deg; 0.5 c/d, 4 Hz, 4.5 deg; and 1.30 c/d, 4 Hz, 4.7 deg. The width σx and height σy (Fig. 1a) were 1.25 and 2.5 deg, respectively.
Let the peak saturations of red and green areas of a grating be |R| and |G|. For a given condition, we first generated and performed motion calibration on a set of red-green gratings with various saturation ratios |G|/|R|, with |R| = constant. |R| was fixed at 90% of the maximum achievable, and |G| was varied because the apparatus afforded a greater range for |G|. The gratings used the maximum contrasts available on the apparatus. The method of constant stimuli was then used to study how the |G|]/|R| ratio would affect motion direction judgments. Forty trials were run at each ratio in each condition.
Results.
Fig. 3 shows percent correct in motion direction judgment as a function of the |G|/|R| ratio for all four display conditions. For each observer in each display condition, a V-shaped function was observed. The critical ratio rc = |G|/|R| at the bottom of the V determines performance that is at or very near chance. rc varies widely between observers and display conditions; the range was from 0.9 to 1.7. For |G|/|R| ≠ rc, performance could be very good (≈85–90%); in fact, along any radius, performance increases monotonically with the distance from rc.
Figure 3.

Accuracy of up/down motion–direction judgments of isoluminant gratings as a function of the green/red saturation ratio, |G|/|R|. Data are shown for four subjects (HT, HK, LL, SL) and four conditions.
Summary and Conclusions.
Motion strength in isoluminant red-green gratings is determined by the |G|/|R| ratio and goes to zero for a critical ratio rc that varies widely between observers. At rc, the red and green areas of the grating are both isoluminant and isosalient. There is no luminance modulation (so the first-order motion system is silent), and no salience modulation (so the third-order motion system is silent). Consequently, no motion is perceived, and motion–direction judgments fall to chance levels. This is true for a variety of red-green gratings chosen from a range of spatial and temporal frequencies optimal for perceiving isoluminant chromatic motion.
The spatial and temporal frequencies for which motion standstill occurred are typically ideal for seeing isoluminant motion. It has previously been shown (7) that the third-order system (and not the first- or second-order system) sees isoluminant red-green gratings. When third-order is silenced at isosalience, and motion standstill ensues, it means that third-order is the only mechanism for perceiving isoluminant color motion.
Experiment 2: Salience and Luminance Jointly Determine Perceived Speed
Procedure.
Experiment 2 uses a magnitude estimation procedure to investigate the factors that determine the perceived speed of moving chromatic displays. We wish to demonstrate that, at isoluminance and isosalience, observers are unable to judge motion direction (as in Experiment 1) because they actually experience motion standstill, rather than perceiving motion in random directions from trial to trial. We further wish to determine the actual speed of ordinary luminance gratings that would be judged equal to the perceived speeds of the chromatic gratings. The experiment consisted of three parts.
In Part I of the experiment, the observers practiced making reliable speed magnitude estimates. A 0.5 c/d, luminance sinewave grating moving at 4 deg/sec with a contrast of 8% was used as the standard; its speed was assigned a magnitude of 10. A luminance sinewave grating moving at a speed chosen randomly from 4, 2, 1, 0.5, and 0.25 deg/sec was shown to the observer on each trial at the same eccentricity as that of the chromatic gratings. All luminance (and subsequent chromatic) gratings had a spatial frequency of 0.5 c/d and a duration of 12 frames at 60 frames/sec, with the first and the last four frames being linearly ramped (Fig. 1b). Different speeds were generated by varying the amount of phase shift between frames, with the same amount of phase shift between successive frames for a given speed. Ten trials of each speed were sufficient for the observers to establish very consistent responses.
Part II of the experiment determined the isoluminant point for a moving chromatic grating at 0.5 c/d, 2 Hz, with equal red and green modulation amplitude (8% in DLK units) for a given eccentricity. This was necessary because, in Part III, observers were to estimate perceived speeds near isoluminance.
In Part III of the experiment, the speed estimation procedure was used to match the perceived speed of chromatic gratings to the speed of luminance gratings. Tests were conducted at many |G|/|R| ratios near and away from isoluminance. All chromatic gratings had spatial and temporal frequencies, respectively, of 0.5 c/d and 2 Hz, resulting in a speed of 4 deg/sec. The red modulation amplitude was fixed (at 0.08, in DLK space, near the maximum achievable with the apparatus) and green modulation amplitudes were varied from trial to trial. To each of these chromatic gratings was added a luminance sinewave grating; its amplitude varied from trial to trial. The amplitudes of most of the luminance gratings were close to the amplitude of luminance correction determined in Part II, so that most stimuli were near the isoluminant and isosalient point. A few “extreme” luminance amplitudes and |G|/|R| ratios were also tested to determine the perceived speed far from isoluminance and isosalience.
The chromatic gratings, physically moving at 4 deg/sec, along with luminance gratings moving at 4 deg/sec, 2 deg/sec, 1 deg/sec, 0.5 deg/sec, and 0.25 deg/sec (as described in Part I), were shown to the observer in parafoveal viewing in random order. The chromatic gratings were identical to the “standard” luminance grating, which moved at 4 deg/sec, in all respects except color.
The observer was required to estimate the magnitude of the speed of the grating presented on each trial. Luminance and color gratings were intermixed, so that the judged speed of color gratings could be compared directly to similarly rated luminance gratings. All gratings were tested in a random order. The entire sequence was repeated twice more in different random orders, so that three speed judgments were obtained for each grating.
Analysis.
The data from Part III were segregated into two groups: those for the luminance gratings and those for the chromatic gratings. Speed judgments of luminance gratings yielded a function relating judged magnitude M to physical speed S. For both observers, the function M = kSγ, where k is a constant and γ = 0.60 and 0.90 for observers LL and HK, respectively, described the luminance data very well (r2 = 0.98). The relation between S and M was then used to convert the judged speed of chromatic gratings to the speed of the equivalently moving luminance grating.
Results.
Fig. 4 shows the perceived speed of chromatic gratings as a function of deviations from isoluminance (x axis) and deviations from isosalience (y axis). The perceived speed is the speed of the matched luminance grating. The actual speed (4 deg/sec) of all of the color and of the standard luminance grating is defined as 100. For example, a perceived speed of 50 means that the color grating was matched to a luminance grating that moved at 1/2 the actual speed of the color grating. (It is important to remember that both the color and the 100% matching luminance grating had the same estimated cone contrast and moved at the same objective speed.) The data are the average of three judgments.
Figure 4.

Speed ratings of red-green gratings as a function of green/red saturation ratio |G|/|R| and the contrast of an added luminance grating. The dotted lines indicate the points at which the added luminance grating produced isoluminance in the calibration procedure. The moving grating was 0.5 cycles/deg, 2 Hz, speed 4 deg/sec, 8% contrast in DLK space, and was viewed at eccentricities of 3.5 and 4.5 deg (observers LL and HK). The rated speed of a matched, 8% contrast black-white grating is taken as 100%. The numbers in the graph indicate the speed of the black-white grating to which the speed of the color grating was matched. Each point is the average of three judgments made in consecutive sessions. 0 indicates absolute standstill on all trials.
Motion Standstill.
For each observer there are at least two combinations of added luminance and |G|/|R| that result in a perceived speed of zero (on a scale 0–100) on all three trials. These are points of isoluminance and isosalience. Nearby, there are combinations of luminance and salience that result in very low perceived speeds, often because one of the three trials was not perceived as stationary. In Fig. 4, the slant of observer LL's isoluminant locus demonstrates that his isoluminant axis (the L+M dimension) is not completely orthogonal to his red-green (L-M) axis as determined in our calibration.
Second, moving away from isoluminance and isosalience, in either direction (luminance, |G|/|R| ratio), increases perceived speed. Subjects are not aware of any difference in perceived movement produced by added (moving) luminance gratings or movement produced by |G|/|R| ratios different from isosalience, although they are aware, of course, of the colors of the stimuli.
Even at the extremes of Fig. 4, none of the color gratings is perceived to move as fast as the standard, although every grating represented in the graph has precisely the same physical speed. The change in perceived speed from motion standstill to good apparent motion as luminance is added to a (previously) isoluminant grating occurs within a very small range of added luminances. Adding a luminance component of ≈3% to an isoluminant color grating (the left and right extrema in Fig. 4) increases the speed from zero at isoluminance and isosalience to 48–83% (mean = 65%). Because 3 and 8% luminance gratings (without color) have almost the same perceived speeds, the presence of the moving color grating the presence of the moving color grating seems to slow the luminance grating. This is consistent with previous reports (3, 6, 55).
The extreme |G|/|R| ratios at the top and bottom of Fig. 4 produce perceived speeds of 34–70% (mean = 55%) of the actual speed. The perceived speed resulting from a large salience modulation in an isoluminant grating is slightly smaller than the speed from added luminance but nevertheless produces high-quality apparent motion. We have shown (7) that high-quality apparent motion is also obtained when gratings with very large or very small |G|/|R| ratios have been individually calibrated to isoluminance.
The symmetry of the data along the two axes of Fig. 4 indicates that there are two contributions to the strength of apparent motion: luminance and |G|/|R| ratio, corresponding to first-order and third-order motion computations. We consider the possibility that the third-order motion system also has some sensitivity to luminance, especially when it is added to a color to produce a perceived color change: e.g., from red to pink. A similar phenomenon (third-order motion sensitivity to a variable aimed at a lower-order motion system) was noted by Ho and Sperling (56). They observed sensitivity of the third-order motion system to texture contrast—which was intended as a second-order motion stimulus. Third-order motion sensitivity to luminance-produced color changes may account for the fact that the point of minimum motion does not lie exactly on isoluminance line (Fig. 4) as determined in the calibration procedure.
Summary and Conclusions
Objective Motion–Direction Judgments.
Red-green grating stimuli were chosen with high-contrast and optimal spatial and temporal parameters for perceiving chromatic motion. In Experiment 1, observers were unable to judge the motion direction of such gratings when they were both isoluminant and isosalient: i.e., when added luminance exactly canceled luminance contamination and when the critical ratio of green saturation to red saturation rc = |G|/|R| produced isosalience. Values of rc varied from 0.9 to 1.7, depending on observers and conditions.
Subjective Speed Ratings.
In Experiment 2, stimuli similar to the ones of Experiment 1 for which observers were unable to judge motion direction, as well as a wide range of related stimuli that varied in the amount of luminance correction and |G|/|R|, were rated for their apparent speed. For a critical range of luminance corrections, and |G|/|R| ratios, reliable motion standstill was observed. That is, when speed was rated on a scale of 0–100, these stimuli received three consecutive zeros; i.e., they were consistently judged to be absolutely motionless.
When a grating was perceived to be motionless, the grating pattern itself was perfectly visible with normal color. This is a striking instance of the shape system delivering a high-quality image of a moving grating, and the color system providing an accurate representation of the grating's color, while the motion system is failing. Adding a ±3% luminance sinewave to the motionless isoluminant, isosalient red-green green grating restored perceived speed to ≈65% of the actual speed, which is normal for 3% contrast. Increasing or decreasing the |G|/|R| ratio by a factor of 4 brought the speed to ≈56%, even with a minimal contribution from the first-order (luminance) system. Either the first-order or third-order motion system alone can produce good apparent motion.
The Mechanism of Isoluminant Motion Perception.
In our conditions, all stimulus components move in the same direction; i.e., there is no motion competition. When motion standstill occurs, it means there is no successful motion computation. Previously, it has been shown that chromatic motion can be perceived by third-order but not by first- or second-order motion systems (7). That motion standstill occurs with high contrast red-green gratings when the luminance component (first-order) and salience component (third-order) have been balanced, indicates that there are no other mechanisms available to sense the motion of these stimuli. That is, isoluminant color motion is perceived only by the third-order motion system.
Conclusions.
Motion standstill can be produced in red-green chromatic stimuli by a delicate adjustment of luminance contrast to cancel residual luminance contamination and by an adjustment of the |G|/|R| saturation ratio to produce isosalience. That the shape and color of the apparently motionless moving grating are clearly visible indicates good performance of the shape and color systems under conditions in which the motion system fails.
Acknowledgments
This work was supported by the U.S. Air Force Office of Scientific Research, Life Sciences, Visual Information Processing Program.
Abbreviations
- deg
degrees
- DLK
Derrington-Lennie-Krauskopf
- L-M
long–medium
- c/d
cycles per degree
Footnotes
To demonstrate this requires small amplitude modulations (low-contrast stimuli) to avoid nonlinear distortions that result from gain-control and similar processes before the motion computation (22). For continuous displays, it requires an integer number of cycles. For temporally sampled displays, it requires 90-deg phase shifts between successive frames and 4n + 1 frames per display, where n is a positive integer (23).
References
- 1.Julesz B, Payne R. Vision Res. 1968;8:433–444. doi: 10.1016/0042-6989(68)90111-9. [DOI] [PubMed] [Google Scholar]
- 2.Moreland J D. In: Colour Vision Deficiencies. Verriest B G, editor. Bristol, U.K.: Hilger; 1980. pp. 189–191. [Google Scholar]
- 3.Cavanagh P, Tyler C W, Favreau O E. J Opt Soc Am A. 1984;1:893–899. doi: 10.1364/josaa.1.000893. [DOI] [PubMed] [Google Scholar]
- 4.Livingstone M S, Hubel D H. J Neurosci. 1987;7:3416–3468. doi: 10.1523/JNEUROSCI.07-11-03416.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Teller D Y, Lindsey D T. J Opt Soc Am A. 1993;10:1324–1331. doi: 10.1364/josaa.10.001324. [DOI] [PubMed] [Google Scholar]
- 6.Mullen K T, Boulton J C. Vision Res. 1992;32:483–488. doi: 10.1016/0042-6989(92)90240-j. [DOI] [PubMed] [Google Scholar]
- 7.Lu Z-L, Lesmes L A, Sperling G. Proc Natl Acad Sci USA. 1999;96:8289–8294. doi: 10.1073/pnas.96.14.8289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Watson A B, Ahumada Jr. A J. In: Motion: Perception and Representation. Tsotsos J K, editor. New York: Assoc. Computing Machinery; 1983. pp. 1–10. [Google Scholar]
- 9.van Santen J P, Sperling G. J Opt Soc Am A. 1984;1:451–473. doi: 10.1364/josaa.1.000451. [DOI] [PubMed] [Google Scholar]
- 10.Adelson E H, Bergen J K. J Opt Soc Am A. 1985;2:284–299. doi: 10.1364/josaa.2.000284. [DOI] [PubMed] [Google Scholar]
- 11.Sperling G, Lu Z-L. In: High-Level Motion Processing. Watanabe T, editor. Cambridge, MA: MIT Press; 1998. pp. 153–183. [Google Scholar]
- 12.Chubb C, Sperling G. J Opt Soc Am A. 1988;5:1986–2006. doi: 10.1364/josaa.5.001986. [DOI] [PubMed] [Google Scholar]
- 13.Lu Z-L, Sperling G. Nature (London) 1995;377:237–239. doi: 10.1038/377237a0. [DOI] [PubMed] [Google Scholar]
- 14.Lu Z-L, Sperling G. Vision Res. 1995;35:2697–2722. doi: 10.1016/0042-6989(95)00025-u. [DOI] [PubMed] [Google Scholar]
- 15.Lu Z-L, Sperling G. Curr Dir Psychol Sci. 1996;5:44–53. [Google Scholar]
- 16.Blaser E, Sperling G, Lu Z-L. Proc Natl Acad Sci USA. 1999;96:11681–11686. doi: 10.1073/pnas.96.20.11681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zeki S. Nature (London) 1980;284:412–418. doi: 10.1038/284412a0. [DOI] [PubMed] [Google Scholar]
- 18.Ungerleider L G, Mishkin M. In: Analysis of Visual Behavior. Ingle D J, Goodale M A, Mansfield R J, editors. Cambridge, MA: MIT Press; 1982. pp. 549–580. [Google Scholar]
- 19.Livingstone M, Hubel D. Science. 1988;240:740–749. doi: 10.1126/science.3283936. [DOI] [PubMed] [Google Scholar]
- 20.DeYoe E A, Van Essen D C. Trends Neurosci. 1988;11:219–226. doi: 10.1016/0166-2236(88)90130-0. [DOI] [PubMed] [Google Scholar]
- 21.Schiller P H, Logothetis N K, Charles E R. Nature (London) 1990;343:68–70. doi: 10.1038/343068a0. [DOI] [PubMed] [Google Scholar]
- 22.Lu Z-L, Sperling G. J Opt Soc Am A. 1996;13:2305–2318. doi: 10.1364/josaa.13.002305. [DOI] [PubMed] [Google Scholar]
- 23.Sperling G, Lu Z-L. Invest Ophthalmol Visual Sci ARVO Supplement. 1998;39:461. [Google Scholar]
- 24.Steinman R M, Collewijn H. Vision Res. 1980;20:415–429. doi: 10.1016/0042-6989(80)90032-2. [DOI] [PubMed] [Google Scholar]
- 25.Sperling G. Am J Psychol. 1970;83:461–534. [Google Scholar]
- 26.Hubel D H, Wiesel T N. J Physiol (London) 1959;148:574–591. [Google Scholar]
- 27.Ramachandran V S, Gregory R L. Nature (London) 1978;275:55–56. doi: 10.1038/275055a0. [DOI] [PubMed] [Google Scholar]
- 28.Anstis S M. Philos Trans R Soc London B. 1980;290:153–168. doi: 10.1098/rstb.1980.0088. [DOI] [PubMed] [Google Scholar]
- 29.Cavanagh P, MacLeod D I, Anstis S M. J Opt Soc Am A. 1987;4:1428–1438. doi: 10.1364/josaa.4.001428. [DOI] [PubMed] [Google Scholar]
- 30.Lee J, Stromeyer C F., III J Physiol (London) 1989;413:563–593. doi: 10.1113/jphysiol.1989.sp017669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Palmer J, Mobley L A, Teller D Y. J Opt Soc Am A. 1993;10:1353–1362. doi: 10.1364/josaa.10.001353. [DOI] [PubMed] [Google Scholar]
- 32.Albright T D. Trends Neurosci. 1991;14:266–269. doi: 10.1016/0166-2236(91)90134-g. [DOI] [PubMed] [Google Scholar]
- 33.Cavanagh P. Science. 1992;257:1563–1565. doi: 10.1126/science.1523411. [DOI] [PubMed] [Google Scholar]
- 34.Gorea A, Papathomas T V, Kovacs I. Proc Natl Acad Sci USA. 1993;90:11197–11201. doi: 10.1073/pnas.90.23.11197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Culham J C, Cavanagh P. Vision Res. 1994;34:2701–2706. doi: 10.1016/0042-6989(94)90226-7. [DOI] [PubMed] [Google Scholar]
- 36.Gegenfurtner K R, Hawken M J. Trends Neurosci. 1996;19:394–401. doi: 10.1016/S0166-2236(96)10036-9. [DOI] [PubMed] [Google Scholar]
- 37.Cavanagh P, Favreau O E. Vision Res. 1985;25:1595–1601. doi: 10.1016/0042-6989(85)90129-4. [DOI] [PubMed] [Google Scholar]
- 38.Derrington A M, Badcock D R. Vision Res. 1985;25:1879–1884. doi: 10.1016/0042-6989(85)90011-2. [DOI] [PubMed] [Google Scholar]
- 39.Krauskopf J, Farell B. Nature (London) 1990;348:328–331. doi: 10.1038/348328a0. [DOI] [PubMed] [Google Scholar]
- 40.Papathomas T V, Gorea A, Julesz B. Vision Res. 1991;31:1883–1891. doi: 10.1016/0042-6989(91)90183-6. [DOI] [PubMed] [Google Scholar]
- 41.Kooi F L, de Valois K K. Vision Res. 1992;32:657–668. doi: 10.1016/0042-6989(92)90182-i. [DOI] [PubMed] [Google Scholar]
- 42.Chichilnisky E-J, Heeger D, Wandell B A. Vision Res. 1993;33:2113–2125. doi: 10.1016/0042-6989(93)90010-t. [DOI] [PubMed] [Google Scholar]
- 43.Cropper S J, Derrington A M. Nature (London) 1996;379:72–74. doi: 10.1038/379072a0. [DOI] [PubMed] [Google Scholar]
- 44.Webster M A, Mollon J D. Vision Res. 1997;37:1479–1498. doi: 10.1016/s0042-6989(96)00289-1. [DOI] [PubMed] [Google Scholar]
- 45.Burr D C, Fiorentini A, Morrone C. Vision Res. 1998;38:3681–3690. doi: 10.1016/s0042-6989(98)00056-x. [DOI] [PubMed] [Google Scholar]
- 46.Brainard D. Spatial Vision. 1997;10:443–446. [Google Scholar]
- 47.Brainard D. In: Human Color Vision. Kaiser P K, Boynton R M, editors. Washington DC: Opt. Soc. Am.; 1996. pp. 563–579. [Google Scholar]
- 48.MacLeod D I A, Boynton R M. J Opt Soc Am A. 1979;69:1183–1186. doi: 10.1364/josa.69.001183. [DOI] [PubMed] [Google Scholar]
- 49.Derrington A M, Krauskopf J, Lennie P. J Physiol (London) 1984;357:241–265. doi: 10.1113/jphysiol.1984.sp015499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Howarth P A. Ophthalmic Physiol Opt. 1984;4:223–226. doi: 10.1111/j.1475-1313.1984.tb00359.x. [DOI] [PubMed] [Google Scholar]
- 51.Simonet P, Campbell M C. Ophthalmic Physiol Opt. 1990;10:271–279. [PubMed] [Google Scholar]
- 52.Zhang X, Bradley A, Thibos L N. J Opt Soc Am A. 1993;10:213–220. doi: 10.1364/josaa.10.000213. [DOI] [PubMed] [Google Scholar]
- 53.Anstis S, Cavanagh P. In: Colour Vision. Mollon J D, Sharpe E T, editors. New York: Academic; 1983. pp. 155–166. [Google Scholar]
- 54.Lu Z-L, Sperling G. Invest Opthalmol Vision Sci. 1999;40:S199. [Google Scholar]
- 55.Farell B. Vision Res. 1999;39:2633–2647. doi: 10.1016/s0042-6989(99)00039-5. [DOI] [PubMed] [Google Scholar]
- 56.Ho C E, Sperling G. Invest Ophthalmol Vision Sci. 1999;40:S425. [Google Scholar]