

Abstract
Interaction between the cerebral hemispheres may allow both hemispheres to contribute their processing resources in order to cope efficiently with complex tasks (Banich, 1998). The current study investigated whether the benefits of interhemispheric interaction arise because of top-down knowledge about the task built up over the course of a block of trials or because of the processing demands present in a single trial. Participants performed a less computationally complex physical identity task and a more complex adding task on within-visual field and across-visual field trials. Task differences in interhemispheric interaction were compared between the blocked and mixed conditions to investigate whether frequent task switches altered the pattern of interhemispheric resource recruitment. A similar interaction between task difficulty and trial type (across-or within- visual field presentation) was obtained for both the blocked and mixed condition. The degree of task-dependency of interhemispheric interaction was not altered in the mixed condition. This finding supports the view that interhemispheric interaction becomes beneficial in response to the processing demands of an individual trial rather than as a result of top-down task knowledge.
Keywords: attention, corpus callosum, task complexity
Understanding the conditions and mechanisms by which the cerebral hemispheres interact may shed light on how the brain’s processing power is used. Interaction between the hemispheres may allow humans to cope efficiently with complex tasks by allowing both hemispheres to contribute their processing resources as needed (Banich, 1998). The current study investigated the flexibility of the recruitment of interhemispheric resources in order to determine whether adjustments in interhemispheric processing can occur rapidly as task demands change. Knowledge of this type is important in order to determine whether the benefits of interhemispheric interaction arise because of top-down task knowledge or because of processing demands that arise as the task is performed.
The Banich paradigm (Banich & Shenker, 1994; Belger & Banich, 1998; Weissman & Banich, 2000) is a well-known measure of interhemispheric interaction. In this paradigm, task complexity and trial type (across or within hemisphere) are manipulated. Participants typically perform a less complex task that requires only simple perceptual matching in one block and a more complex task that requires at least one additional computational step in another block of trials. For both tasks, half of the trials are within-hemisphere trials in which both relevant items are presented to the same visual field and half are across-hemisphere trials in which one relevant item is presented to each visual field. Producing a correct response on within-hemisphere trials does not require interhemispheric interaction as one hemisphere receives all the relevant information. For across-hemisphere trials, in contrast, information must be transferred between the hemispheres for a correct response to be made.
According to Banich (1998), there are both costs and benefits to interhemispheric interaction. Because processing must be coordinated and integrated, there can be a cost to transferring information between the hemispheres. However, this cost can be offset by the benefit of allowing the two hemispheres to process relevant information in parallel. Typically, task complexity and trial type interact such that performance is better on across-hemisphere trials only for the more complex task; for the less complex task, performance is either equivalent across trial types or better for within-hemisphere trials (Belger & Banich, 1998; Koivisto, 2000; Rueter-Lorenz, Stanczak, & Miller, 1999; Weissman & Banich, 2000). The authors interpret these results by suggesting that the benefits of interhemispheric interaction outweigh the costs only when the task is sufficiently computationally complex. For less complex tasks, the benefit of parallel processing is not sufficient to overcome the cost of transferring information. For the more complex task, the computational demands are great enough that the parallel processing afforded by across-hemisphere presentation outweighs the cost of transfer.
Functional neuroimaging studies have shown that computationally complex tasks elicit a greater degree of bilateral activation than simpler tasks (Braver et al, 1997; Klingberg, O’Sullivan, & Roland, 1997). A functional imaging study using a variation of the Banich paradigm demonstrated that a more complex letter name task led to more bilateral activation in the fusiform and lateral occipital gyri than a less complex letter shape task (Pollmann, Zaidel, & Cramon, 2003). These findings are consistent with the view that interhemispheric resource sharing increases with increasing task demands.
Previous studies have presented the more and less computationally complex tasks in separate blocks of trials (Belger & Banich, 1998; Koivisto, 2000; Rueter-Lorenz, Stanczak, & Miller, 1999; Weissman & Banich, 2000). While this approach protects against the possibility that participants will perform the same processing for both tasks, it also builds expectancies such that participants are fully aware of the task that will be performed well in advance of the actual stimulus presentation. Because of this design, previous studies have not addressed the question of whether the benefits of interhemispheric interaction arise because of top-down knowledge about the task or because of the processing demands that arise on a single trial. The current study aimed to determine whether recruitment of resources through interhemispheric interaction is flexible enough that task differences will appear when task switches occur frequently and unpredictably.
One possibility is that top-down factors influence the degree to which interhemispheric interaction will occur. It is possible that knowledge about the features of the task biases the system towards across-hemisphere processing for complex tasks and towards within-hemisphere processing for less complex tasks. One proposed task property relating to the degree of benefit provided by interhemispheric interaction is the degree of hemispheric specialization for the task (Yazgan, Wexler, Kinsbourne, Peterson, & Leckman, 1995). Another feature of the task proposed to underlie degree of interhemispheric interaction is computational complexity (Weissman & Banich, 2000). If repeated experience through the course of a block of trials controls the extent to which interhemispheric interaction will be beneficial, mixed presentation should reduce or eliminate the interaction between task complexity and trial type.
An alternate explanation of how an across-hemisphere advantage arises is that the resources of the other hemisphere are automatically recruited whenever the resources of one hemisphere are insufficient to complete the task (Weissman & Banich, 2000). According to this explanation, no top-down knowledge about the task to be performed or expectations regarding the upcoming task are necessary in order for a differential benefit of interhemispheric interaction to be shown for tasks of differing complexity. Instead, a property of the individual trial controls the extent to which interhemispheric interaction is beneficial. If degree to which interhemispheric interaction is beneficial is related to the properties of an individual trial, a similar pattern of results should be shown in the mixed condition as in the blocked condition.
This study, then, investigated whether efficient allocation of hemispheric resources results from top-down processing relying on task expectations or from automatic processing relying on properties of individual trials. Whether interhemispheric resource allocation is controlled by task expectations or trial properties might have implications for other aspects of this process. Automatic processes are generally fast and do not require attention; in contrast, controlled processes are slower but more flexible (Schneider and Shiffrin, 1977). If the degree of interhemispheric interaction is determined by task expectations, it might also be expected to be a slower, attentionally demanding process. If the degree of interhemispheric interaction is determined by stimulus properties, it might be expected to be a faster, more flexible process.
Some support for the hypothesis that processing mode selection is flexible, and not fully determined by task properties, comes from previous studies examining the effects of practice with a task on interhemispheric interaction (Cherbuin & Brinkman, 2005; Liederman, Merola, & Martinez, 1985; Maertens & Pollman, 2005; Weissman & Compton, 2003). In each of these studies, performance on within-hemisphere trials showed greater improvement over the course of the experiment than performance on across-hemisphere trials. The result was that an advantage for across-hemisphere trials present early in a task was reduced or eliminated with more experience with the task. These findings indicate that the degree of benefit from interhemispheric interaction elicited by a task is not a stable property of the task, but can vary as a function of processing demands that change over the course of an experiment. However, practice effects build up gradually over the course of an experiment. To more directly investigate the flexibility of recruitment of interhemispheric resources, the current study included a condition in which task demands changed much more rapidly.
The current study investigated whether experience with a task built up over the course of a block of trials is necessary for the differential benefit of interhemispheric interaction for complex tasks to occur. Participants performed a less difficult digit-matching task and a more difficult adding task. Digit tasks were used so that exactly the same stimuli could appear in the more difficult and less difficult tasks; in the more typical letter matching tasks, target letters are uppercase for one task and lowercase in the other. Performance in a blocked condition analogous to previous studies was compared to performance in a mixed condition in which the task switched every 1, 2, or 3 trials. Participants were cued as to which task to perform by a letter at fixation that appeared 150 ms before the stimulus array. If experience with a task is necessary in order for interhemispheric interaction to be differentially beneficial for more complex tasks, the degree of across-hemisphere advantage would be reduced in the mixed condition relative to the blocked condition.
In addition, for the mixed condition, performance on trials in which the task was the same as that performed on the previous trial could be compared to performance on trials on which a task switch occurred. These comparisons are a test of whether recent experience with a task impacts the degree to which interhemispheric interaction will be beneficial. If recent experience biases the system to process information similarly on subsequent trials, the task effect on interhemispheric interaction would be reduced or eliminated for trials immediately following a task switch.
Method
Participants
120 students (60 female) from the University of California, Riverside participated in the experiment and were compensated with course credit or $7. All participants were right-handed as assessed by a five-item preference questionnaire (Bryden, 1982) resulting in a scale ranging from −1 (most left-handed) to +1 (most right-handed). The mean handedness rating was 0.90. Participants all had normal or corrected-to-normal vision.
Apparatus and Stimuli
Participants were seated in front of Macintosh computers equipped with Psyscope software (Cohen, MacWhinney, Flatt, & Provost, 1993) to control stimulus presentation, regulate timing of events, and record participants’ responses. Stimuli were displayed in black on a white background. Throughout the experiment each participant’s head was kept a constant 60 cm from the screen through the use of a headrest.
The stimuli were the digits 0–9. Following Banich (1998; Weissman & Banich, 2000), the stimuli consisted of a triangular array of digits surrounding a central fixation point. The two probe digits were presented 2.8° lateral to and 1.4° above fixation. The target digit appeared 1.4° lateral to and 1.4° below fixation. Each digit subtended between .29° and .48° horizontally and .67° vertically.
On each trial, participants completed one of two tasks cued by a letter at fixation (“A” for the adding task or “S” for the physical identity task). For the physical identity task, participants were asked to decide whether the bottom digit matched either of the top two digits. For the adding task, participants were asked to decide whether the sum of the bottom digit and one of the top two digits was exactly ten. Half of the 384 items were “yes” trials in which the target digit matched with or summed to ten with one of the probe digits and the other half were “no” trials. For half of the “yes” trials (96 trials), the bottom item appeared on the same side of the fixation point as the probe item it matched (within-hemisphere trials). For the other half of the “yes” trials, the bottom item appeared on the opposite side of the fixation point from the item it matched (across-hemisphere trials). The target digit appeared to the left of the fixation point in half of the within-hemisphere trials, half of the across-hemisphere trials, and half of the mismatch trials and to the right of the fixation point in the other half1. Fifteen practice trials having the same composition as experimental trials were constructed for each task.
For half of the participants, the task was blocked so that participants completed all of the physical identity trials, followed by all of the adding trials. This task ordering is typical for experiments employing the Banich paradigm (Belger & Banich, 1998; Koivisto, 2000; Rueter-Lorenz, Stanczak, & Miller, 1999; Weissman & Banich, 2000) and prevents participants from carrying over a strategy from the more complex task to the less complex task. For the other half of the participants, the task was switched randomly every one, two, or three trials2. For half of these trials, the task was the same as the task performed on the previous trial. For the other half, the task was different from that performed on the previous trial. Switches occurred equally often after one, two, or three repetitions of the same task.
Procedure
Participants were instructed to fixate on the fixation cross at all times and to respond as quickly and accurately as possible. On each trial of each task, a fixation cross was displayed in the center of the screen. After 300 msec, the fixation cross was replaced by the cue letter for 50 ms in order to instruct the participant as to which task to do. The fixation cross then re-appeared and remained during the presentation of targets and probes. The target and probe digits appeared simultaneously 150 ms after the cue letter and remained on the screen for 140 ms. The intertrial interval was 1500 ms. Participants responded by pressing the “.” or “0” key on the numeric keypad with the index finger or middle finger of their right hand. Response-key mappings were counterbalanced across participants. Participants received a practice block of 15 trials of each task before beginning the experimental trials. The 384 experimental items for each task were divided into four blocks of 96 trials.
Results
2×2×2 mixed design analyses of variance (ANOVAs) were performed on mean correct response times (RTs) and mean error rates for match trials with the following variables: task condition (blocked or mixed), task (physical identity or adding) and trial type (across or within VF). Task condition was the only between-subjects factor. Correct trials on which the RT was 2.5 or more standard deviations from the subject’s mean were trimmed to the subject’s mean ± 2.5 SD. Because this procedure takes into account each individual’s mean RT, it is able to identify outliers specific to a particular participant. Trimming affected 2.7 % of responses overall. Mean RTs and error rates are given in Table 1.
Table 1.
Mean Error Rate and RT (Standard Deviation) by Task Condition, Task, and Trial Type
| Error Rate (%) | Reaction Time (ms) | |||
|---|---|---|---|---|
| Within-VF | Across-VF | Within-VF | Across-VF | |
| Blocked | ||||
| Physical Identity | 2.4 (2.8) | 3.3 (3.5) | 621 (110) | 634 (103) |
| Adding | 2.9 (3.6) | 1.9 (2.9) | 708 (120) | 686 (103) |
| Mixed | ||||
| Physical Identity | 5.7 (5.7) | 5.8 (6.2) | 793 (188) | 788 (181) |
| Adding | 4.6 (4.5) | 3.7 (4.4) | 866 (199) | 828 (177) |
To further examine the effects of task switching on interhemispheric interaction, correct trials in the mixed condition were submitted to a 2×2×2 ANOVA with the following within-subjects variables: switch (switch or no switch), task (physical identity or adding) and trial type (across or within VF). Mean RTs and error rates are given in Table 2.
Table 2.
Mean Error Rate and RT (Standard Deviation) by Switch Condition, Task, and Trial Type in Mixed Condition
| Error Rate (%) | Reaction Time (ms) | |||
|---|---|---|---|---|
| Within-VF | Across-VF | Within-VF | Across-VF | |
| No Switch Trials | ||||
| Physical Identity | 4.5 (5.9) | 5.4 (7.0) | 788 (200) | 791 (193) |
| Adding | 3.7 (4.3) | 2.8 (4.1) | 871 (208) | 849 (199) |
| Switch Trials | ||||
| Physical Identity | 6.9 (8.4) | 6.2 (7.6) | 800 (185) | 788 (179) |
| Adding | 5.5 (5.8) | 4.7 (6.7) | 865 (198) | 811 (167) |
Error Rate Results
There was a main effect of task condition F (1, 118) = 15.90 p < .001, reflecting a lower error rate when trials were blocked by task (2.6%) than when switches were present (5.0%). There was also a main effect of task F (1, 118)=7.25, p < 0.01, with a lower error rate for the physical identity task (3.3%) than for the adding task (4.3%). This main effect was tempered by the expected significant two-way interaction between trial type and task F (1, 118)=6.87, p<0.01. For the physical identity trials, there was no significant effect of trial type p > 0.25. For the adding trials, the effect of trial type was significant F (1, 119)=6.32, p < 0.05, reflecting a higher error rate for within-VF trials (3.7%) than for across-VF trials (2.9%). No other significant interactions were obtained. These results indicate that a significant across-hemisphere advantage was present for adding trials but not for physical identity trials, and that this effect was not altered by mixed presentation.
To further examine the effects of task and task condition on the across-hemisphere advantage, the across-hemisphere advantage as a proportion of total correct responses was calculated. An ANOVA was performed on this percentage with task and task condition as variables. There was a significant main effect of task F (1,118)=6.69, p < .05, reflecting a larger across-hemisphere advantage for the adding task (0.95%) than the physical identity task (−0.45%). The main effect of task condition and the interaction of task condition and task were not significant (p > .25). The task difference in the across hemisphere advantage was comparable in the mixed and blocked conditions, with a task difference of 0.92 % in the mixed condition and 1.89 % in the blocked condition.
For the additional analyses in the mixed condition, there was a main effect of switch F (1, 59) = 14.12, p < 0.001, reflecting a lower error rate when the task was the same as the previous trial (4.1%) than when a task switch occurred (5.8%). No other significant main effects or interactions were obtained.
RT Results
There was a main effect of task condition F (1, 118) = 35.76, p <0.0001 reflecting a faster RT when trials were blocked by task (663 ms) than when switches were present (819 ms). There was a main effect of task F (1, 118)=61.93, p < 0.0001, with a faster RT for the physical identity task (709 ms) than for the adding task (773 ms). There was also a main effect of trial type, F (1, 118)=11.55, p < 0.001, reflecting a faster RT for across-VF trials (735 ms) than for within-VF trials (747 ms). These main effects were tempered by the expected significant two-way interaction between trial type and task F (1, 118)=35.26, p<0.0001. For the physical identity trials, there was no significant effect of trial type, p > 0.10. For the adding trials, the effect of trial type was significant F (1, 119)=32.70, p < 0.0001, reflecting a longer RT for within-VF trials (788 ms) than for across-VF trials (758 ms) (See Figure 1). Unexpectedly, the interaction between task condition and trial type was also significant F(1,118)=4.97, p < .05. For the blocked condition, the effect of trial type was not significant, p > .10. For the mixed condition, the overall effect of trial type was significant, F (1,59)=9.51, p < 0.01, reflecting a longer RT for within-VF trials (828 ms) than for across-VF trials (808 ms). This overall across-hemisphere advantage, regardless of task, in the mixed condition may reflect the fact that this condition was more difficult than the blocked condition. The crucial three-way interaction between trial type, condition, and task was not significant, p > 0.20, indicating that the task difference in the across hemisphere advantage was comparable in the mixed and blocked conditions.
Figure 1.
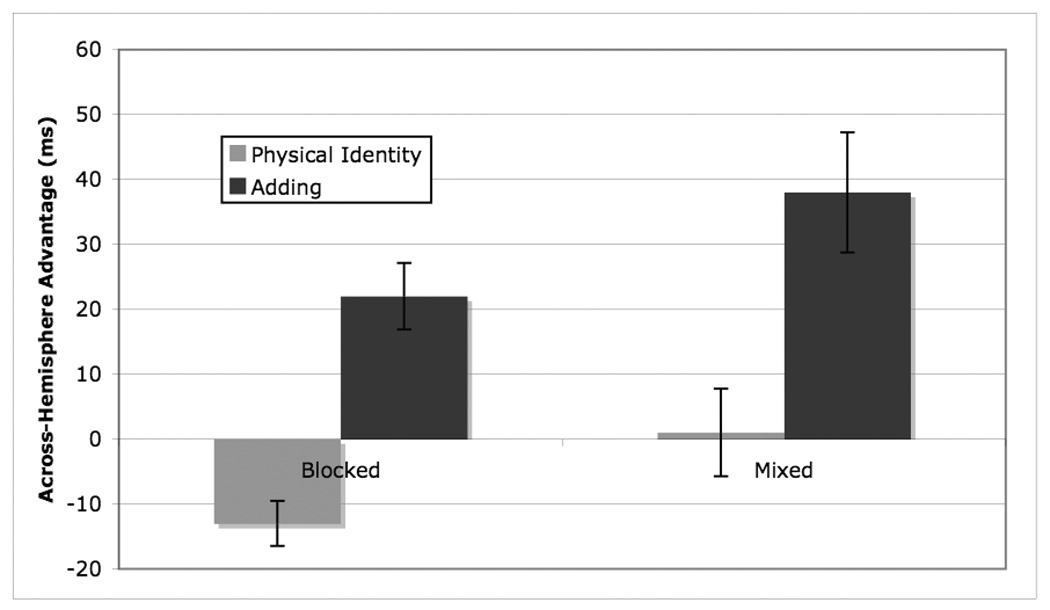
Across hemisphere advantage in RT (RT for within-VF trials – RT for across-VF trials) for each task by task condition
To further examine the effects of task and task condition on the across-hemisphere advantage, the across-hemisphere advantage as a proportion of mean RT was calculated for each participant. An ANOVA was performed on this percentage with task and task condition as variables. There was a significant main effect of task F (1,118)=41.01, p < .0001, reflecting a larger across-hemisphere advantage for the adding task (3.43%) than the physical identity task (−1.10%). There was also a significant main effect of task condition F(1, 118)= 5.42, p < .05, reflecting a greater across-hemisphere advantage, collapsed across task, in the mixed condition (2.08%) than in the blocked condition (0.25%). The interaction between the two variables was not significant, p > .25. The task difference in the across hemisphere advantage was comparable in the mixed and blocked conditions, with a task difference of 4.15% in the mixed condition and 5.02% in the blocked condition.
A further set of analyses examined the effect of switching in the mixed condition. Within the mixed condition, there was a main effect of trial type, F(1, 59)=10.00, p < 0.01, reflecting a faster RT to across-VF trials (810 ms) than within-VF trials (831 ms). There was a main effect of task, F(1,59)=18.35, p < 0.0001, reflecting a faster RT for physical identity trials (792 ms) than for adding trials (849 ms). These effects were tempered by a significant interaction between trial type and task F(1, 59)=8.01, p <.001. For the physical identity task, the effect of trial type was not significant, p < 0.25. For the adding task, the effect of trial type was significant F(1, 59)=15.84, reflecting a longer RT to within-VF trials (868 ms) than to across-VF trials (830 ms) (See Figure 2). There was also a significant interaction between trial type and switch condition. For the No Switch trials, the effect of trial type was not significant, p > 0.10. For the Switch trials, the effect of trial type was significant F(1,59)=13.70, p < 0.001, reflecting a longer RT to within-VF trials (831 ms) than to across-VF trials (798 ms).
Figure 2.
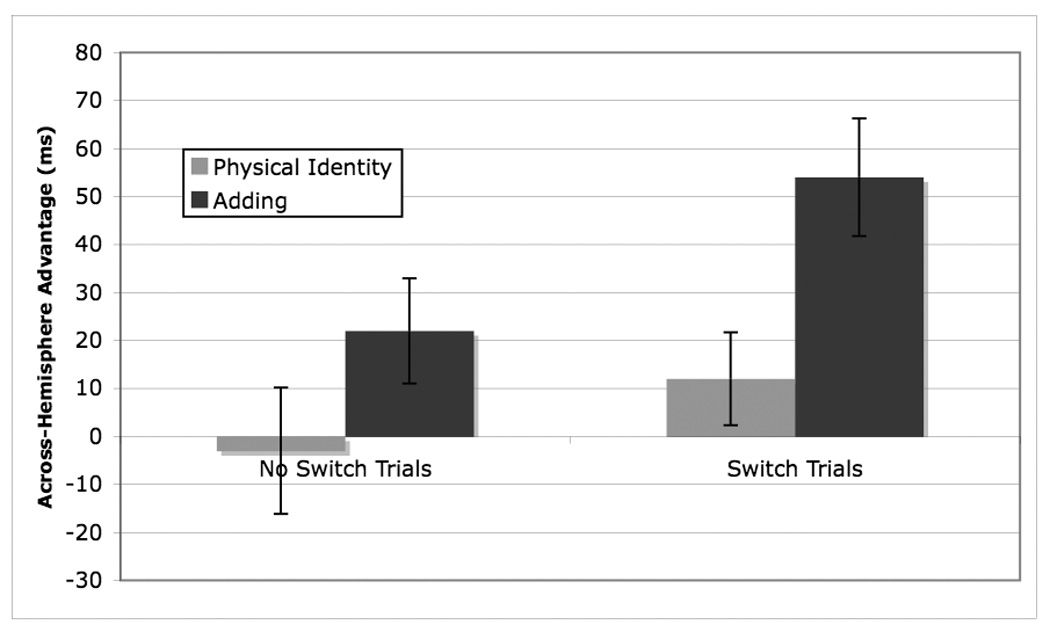
Across hemisphere advantage in RT (RT for within-VF trials – RT for across-VF trials) for each task by switch in mixed condition
This overall across-hemisphere advantage for Switch trials may reflect the increased difficulty of these trials. The crucial three-way interaction between trial type, condition, and task was not significant, p > .20, indicating that the task difference in across hemisphere advantage was comparable for the Switch and No Switch trials.
Discussion
In accordance with previous findings (Belger & Banich, 1998; Koivisto, 2000; Rueter-Lorenz, Stanczak, & Miller, 1999; Weissman & Banich, 2000) a robust across-hemisphere advantage was seen for the more computationally complex adding task but not for the less complex physical identity task. Unexpectedly, there was an interaction between trial type and experimental condition, such that in the mixed condition only, there was an overall across-hemisphere advantage. A similar interaction was found between trial type and Switch condition, in which only switch trials produced an overall across-hemisphere advantage. One potential explanation for these unexpected findings is that the mixed condition and switch trials were more difficult. Indeed, reaction times were longer and error rates higher in these conditions. This increased overall difficulty may have increased the processing demands of both tasks, shifting the baseline across-hemisphere advantage relative to the less complex blocked condition.
Despite this shift in baseline across hemisphere advantage, the tasks difference in the degree to which across-hemisphere processing was advantageous was preserved in the mixed condition. Crucially, the three-way interaction between task condition, task, and trial type was not significant. The key interaction between task and trial type was significant in both the blocked and mixed conditions, reflecting a comparable task dependence in across-hemisphere advantage in the blocked and mixed conditions. These findings support the view that interhemispheric interaction occurs in response to the processing demands of an individual trial, rather than arising from top-down knowledge of the task to be performed. Previous experience built up through a block of trials was not necessary to induce an across-hemisphere advantage selectively for the more computationally complex task. These findings support the view that interhemispheric interaction becomes beneficial when the processing demands overtax the resources of one hemisphere within a trial rather than the view that either an across- or within-hemisphere processing mode is selected in advance because of top-down task knowledge. This suggests that selection of the most efficient processing mode does not require extensive past experience with a task, and instead results from the online processing demands that characterize the task. This would allow for efficient processing even when a situation is new or rarely encountered.
The current findings are inconsistent with the view that the processing mode beneficial on the previous task would be carried over to subsequent trials. When the task to be performed differed from that of the previous trial, the task dependency of the across-hemisphere advantage was preserved. This finding provides further evidence that the degree to which interhemispheric interaction will be beneficial is determined within a single trial, rather than arising from previous experience. This flexibility might allow an efficient processing mode to be selected under conditions in which the tasks and processing demands facing an individual change quickly.
The finding that recruitment of interhemispheric resources is flexible, rather than a static property of the task, is in keeping with previous studies of practice effects (Cherbuin & Brinkman, 2005; Liederman, Merola, & Martinez, 1985; Maertens & Pollman, 2005; Weissman & Compton, 2003). These experiments showed that as the processing demands changed over the course of the experiment, the degree to which interhemispheric interaction was beneficial changed as well. The current study extended these findings to a task-switching situation in which the change in processing demands occurred more rapidly. The ability to adapt to rapidly changing task demands would seem necessary to cope with rapidly changing processing demands present in the real world.
This study had several limitations. First, the cues were presented 150 ms before the stimuli, allowing participants to prepare for the task. Future studies should manipulate time interval between the cue and the stimuli to investigate the degree to which this preparation impacts the degree of across-hemisphere advantage. Future studies might also eliminate the need for cues by using two tasks that can be identified by their stimuli alone (e.g., letter-matching and digit-adding). Additionally, this study involved an indirect measure of interhemispheric interaction. Future studies employing functional imaging technology should investigate the dynamics of interhemispheric interaction.
In summary, the results of this study support the view put forth by Banich and Nicholas (1998) that interhemispheric interaction is “a flexible and dynamic process” (p. 66). Rather than arising from top-down task knowledge or experience on previous trials, the degree to which interhemispheric interaction will benefit performance appears to arise from the processing demands of a single trial. This flexibility may be a mechanism for maximizing the processing power of the brain (Weissman & Banich, 2000).
Authors’ Notes
This research was partially supported by NSF grant BCS0079456 and NIH grant RO1DC006957-01 to the second author. We thank Fong Chen, Jessica Johnston, Elizabeth Cox, Janelle Julagay, and Evan Patterson for their assistance with data collection.
Footnotes
To examine the degree of hemispheric lateralization for the tasks, ANOVAs were performed on RTs and error rates from within-VF with task and VF as variables. These analyses revealed no significant main effect of VF or task by VF interaction. Because there was no clear pattern of lateralization, all analyses were collapsed across the VF of target presentation.
Analyses indicated that there were no main effects or interactions with the number of task repetitions preceding a switch. For this reason, all analyses were collapsed across number of repetitions.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Banich M. The missing link: the role of interhemispheric interaction in attentional processing. Brain and Cognition. 1998;36:128–157. doi: 10.1006/brcg.1997.0950. [DOI] [PubMed] [Google Scholar]
- Banich M, Shenker Investigations of interhemispheric processing: methodological considerations. Neuropsychology. 1994;8:263–277. [Google Scholar]
- Banich MT, Nicholas CD. Integration of reading processing between the hemispheres. In: Beeman M, Chiarello C, editors. Right Hemisphere language comprehension: Perspectives from cognitive neuroscience. Hillsdale, N.J.: Lawrence Erlbaum Associates; 1998. pp. 51–77. [Google Scholar]
- Belger A, Banich MT. Costs and benefits of integrating information between the cerebral hemispheres: a computational perspective. Neuropyschology. 1998;12(3):380–398. doi: 10.1037//0894-4105.12.3.380. [DOI] [PubMed] [Google Scholar]
- Braver TS, Cohen JD, Nystrom LE, Jonides J, Smith EE, Noll DC. A parametric study of prefrontal cotex involvement in human working memory. NeuroImage. 1997;5:49–62. doi: 10.1006/nimg.1996.0247. [DOI] [PubMed] [Google Scholar]
- Bryden MP. Laterality: Functional asymmetry in the normal brain. New York: Academic Press; 1982. [Google Scholar]
- Cherbuin N, Brinkman C. Practice makes two hemispheres almost perfect. Cognitive Brain Research. 2005;24(3):413–422. doi: 10.1016/j.cogbrainres.2005.02.010. [DOI] [PubMed] [Google Scholar]
- Cohen JD, MacWhinney B, Flatt M, Provost J. PsyScope: A new graphic interactive environment for designing psychology experiments. Behavioral Research Methods, Instruments, and Computers. 1993;25:257–271. [Google Scholar]
- Klingberg T, O’Sullivan BT, Roland PE. Bilateral activation of fronto-parietal networks by incrementing demand in a working memory task. Cerebral Cortex. 1997;7:465–471. doi: 10.1093/cercor/7.5.465. [DOI] [PubMed] [Google Scholar]
- Koivisto M. Interhemispheric interaction in semantic categorization of pictures. Cognitive Brain Research. 2000;9(1):45–51. doi: 10.1016/s0926-6410(99)00042-7. [DOI] [PubMed] [Google Scholar]
- Liederman J, Merola J, Martinez S. Interhemispheric collaboration in response to simultaneous bilateral input. Neuropsychologia. 1985;23(5):673–683. doi: 10.1016/0028-3932(85)90068-5. [DOI] [PubMed] [Google Scholar]
- Maertens M, Pollmann S. Interhemispheric resource sharing: Decreasing benefit with increasing processing efficiency. Brain and Cognition. 2005;58(2):183–192. doi: 10.1016/j.bandc.2004.11.002. [DOI] [PubMed] [Google Scholar]
- Pollmann S, Zaidel E, Cramon DY. The neural basis of the bilateral distribution advantage. Experimental Brain Research. 2003;153(3):322–333. doi: 10.1007/s00221-003-1551-0. [DOI] [PubMed] [Google Scholar]
- Reuter-Lorenz PA, Stanczak L, Miller AC. Neural recruitment and cognitive aging: Two hemispheres are better than one, especially as you age. Psychological Science. 1999;10(6):494–500. [Google Scholar]
- Schneider W, Shiffrin RM. Controlled and automatic human information processing: 1. Detection, search, and attention. Psychological Review. 1977;84:1–66. [Google Scholar]
- Weissman DH, Banich MT. The cerebral hemispheres cooperate to perform complex but not simple tasks. Neuropsychology. 2000;14(1):41–59. doi: 10.1037//0894-4105.14.1.41. [DOI] [PubMed] [Google Scholar]
- Weissman DH, Compton RJ. Practice makes a hemisphere perfect: The advantage of interhemispheric recruitment is eliminated with practice. Laterality: Asymmetries of Body, Brain, and Cognition. 2003;8(4):361–375. doi: 10.1080/13576500342000103. [DOI] [PubMed] [Google Scholar]
- Yazgan MY, Wexler BE, Kinsbourne M, Peterson B, Leckman JF. Functional significance of individual variations in callosal area. Neuropsychologia. 1995;33(6):769–779. doi: 10.1016/0028-3932(95)00018-x. [DOI] [PubMed] [Google Scholar]