

Abstract
The multitransmembrane protein Patched (PTCH) is the receptor for Sonic Hedgehog (Shh), a secreted molecule implicated in the formation of embryonic structures and in tumorigenesis. Current models suggest that binding of Shh to PTCH prevents the normal inhibition of the seven-transmembrane-protein Smoothened (SMO) by PTCH. According to this model, the inhibition of SMO signaling is relieved after mutational inactivation of PTCH in the basal cell nevus syndrome. Recently, PTCH2, a molecule with sequence homology to PTCH, has been identified. To characterize both PTCH molecules with respect to the various Hedgehog proteins, we have isolated the human PTCH2 gene. Biochemical analysis of PTCH and PTCH2 shows that they both bind to all hedgehog family members with similar affinity and that they can form a complex with SMO. However, the expression patterns of PTCH and PTCH2 do not fully overlap. While PTCH is expressed throughout the mouse embryo, PTCH2 is found at high levels in the skin and in spermatocytes. Because Desert Hedgehog (Dhh) is expressed specifically in the testis and is required for germ cell development, it is likely that PTCH2 mediates its activity in vivo. Chromosomal localization of PTCH2 places it on chromosome 1p33–34, a region deleted in some germ cell tumors, raising the possibility that PTCH2 may be a tumor suppressor in Dhh target cells.
Hedgehog proteins, a family of secreted molecules first identified by a genetic screen in Drosophila (1), are involved in many patterning processes during development (for review see refs. 2 and 3). Three mammalian hedgehog homologues have been identified: Sonic (Shh), Desert (Dhh), and Indian (Ihh). Shh acts to establish cell fate in the developing limb, somites, and neural tube (4–13). Ihh is involved specifically in chondrocyte development (14), and Dhh plays a key role in germ cell development (15). With the exception of the gut, in which both Ihh and Shh are expressed, the expression patterns of the hedgehog family members do not overlap (16).
At the cell surface, Shh function appears to be mediated by a multicomponent receptor complex involving Patched (PTCH) and Smoothened (SMO), two multitransmembrane proteins initially identified as segment polarity genes in Drosophila and later characterized in vertebrates (17–22). Both genetic and biochemical evidence supports the existence of a receptor complex in which PTCH is the ligand-binding subunit and SMO, a G protein-coupled receptor-like molecule, is the signaling component (22–24). Upon binding of Shh to PTCH, the normal inhibitory effect of PTCH on SMO is relieved, allowing SMO to transduce the Shh signal across the plasma membrane. It remains to be established whether the PTCH/SMO receptor complex mediates the action of all three mammalian hedgehogs or whether ligand-specific components exist. Interestingly, a second murine PTCH gene, PTCH2, was isolated recently (25) but its function as a hedgehog receptor has not been established. To characterize PTCH2 and compare it with PTCH with respect to the biological function of the various hedgehog family members, we isolated the human PTCH2 gene. Binding analysis shows that both PTCH and PTCH2 bind to all three hedgehog ligands with similar affinity. Furthermore PTCH2 interacts with SMO, suggesting that it can form a functional multicomponent hedgehog receptor complex similar to PTCH-SMO. Tissue distribution analysis indicates that PTCH2 is preferentially expressed in the skin and in testis, where it is likely to mediate the action of Dhh, which is required for germ cell development.
MATERIALS AND METHODS
cDNA Cloning.
The LIFESEQ(R) database (Incyte Pharmaceuticals, Palo Alto, CA) was screened for the presence of sequences related to PTCH. Two expressed sequence tag sequences homologous to PTCH were identified, and a lambda gt10 testis cDNA library was screened with an overlapping oligo probe derived from one of them: (5′-CTGCGGCGCTGCTTCCTGCTGGCCGTCTGCATCCTGCTGGTGTGC-3′) and (5′-AGAGCACAGACGAGGAAAGTGCACACCAGCAGGATGCAGACGGCC-3′). Filters were hybridized overnight at 42°C in 50% formamide/5× SSC/10× Denhardt’s/0.05 M sodium phosphate, pH 6.5/0.1% sodium pyrophosphate/50 μg/ml of sonicated salmon sperm DNA, and then washed in 0.1× SSC/0.1% SDS and exposed to Kodak x-ray films.
In Situ Hybridization.
E18.5 mouse embryos were immersion-fixed overnight in 4% paraformaldehyde, cryoprotected overnight in 15% sucrose, embedded in paraffin, and sectioned at 10 μm. Adult mouse testes were embedded in optimal temperature cutting compound, frozen on liquid nitrogen, sectioned at 16 μm, and processed for in situ hybridization by a method described previously (26). 33P-UTP-labeled RNA probes were generated as described (27). Sense and antisense probes were synthesized from the 3′ noncoding region of mouse PTCH or PTCH2 and from a mouse Fused cDNA fragment corresponding to the region encoding amino acid 317–486 of the human sequence using T3 and T7, respectively.
Immunoprecipitation and Binding Analysis.
Human 293 embryonic kidney cells were transiently transfected with expression constructs encoding Flag-tagged PTCH or PTCH2 and Myc-tagged SMO by the calcium phosphate method. The SMO-PTCH complex was detected by coimmunoprecipitation from dual-transfected cells. Immunoprecipitation was performed with antibodies to the Flag epitope followed by protein A Sepharose, and then separated on a denatured 6% polyacrylamide gel. Proteins were transferred to nitrocellulose and detected by probing with antibodies to the Flag or Myc epitope, using the enhanced chemiluminescence detection system (Amersham). For binding analysis, 293 cells overexpressing PTCH or PTCH2 were incubated with 100 pM 125I-Shh (bacterially expressed N-terminal fragment) and various concentrations of unlabeled Shh, Dhh, and Ihh.
Immunofluorescence.
Cos-7 cells were transiently transfected with Flag-tagged PTCH or PTCH2. Cells were incubated with Shh-IgG or Dhh-IgG (1 μg/ml, 2 h at 4°C) 48 h after transfection, washed, fixed for 10 min in 4% paraformaldehyde, permeabilized (5 min in 0.1% Triton X-100), and stained for ligand binding (biotin/anti-human IgG followed by cy3-streptavidin) and receptor expression (anti-Flag M2 antibody followed by Cy2/anti-mouse).
RESULTS
cDNA Cloning of Human PTCH2.
To characterize PTCH2 and compare it with PTCH with respect to the biological function of the various hedgehog family members, we have screened expressed sequence tag (EST) databases with the PTCH protein and identified two EST candidates corresponding to a novel human PTCH gene. Northern blot analysis (see below) revealed that the testes were the primary site of expression, and a full-length cDNA encoding human PTCH2 was cloned from a human testis cDNA library. The initiator ATG defines a 3,612-nt ORF encoding a 1,204-aa-long protein with a predicted molecular mass of approximately 131 kDa. The overall identity between human PTCH and PTCH2 is 54% (Fig. 1), while the identity between human PTCH2 and the recently described mouse PTCH2 (25) is 90%. The most obvious structural difference between the two human PTCH proteins is a truncated C-terminal cytoplasmic domain in PTCH2. In addition, only one of the two glycosylation sites present in PTCH is conserved in PTCH2. The gene encoding PTCH2 was mapped by fluorescence in situ hybridization and by PCR using a radiation hybrid panel to human chromosome 1p33–34 (data not shown).
Figure 1.

Amino acid sequence homology between human PTCH and PTCH2. The predicted amino acid sequence of human PTCH2 is aligned with PTCH (31). Gaps introduced for optimal alignment are shown by dashes. Identical amino acids are boxed. The transmembrane domains are highlighted. The conserved glycosylation site is indicated by asterisks.
Binding of Hedgehog Ligands to PTCH and PTCH2.
To determine whether PTCH2 is a hedgehog receptor and whether the two PTCH molecules are capable of discriminating between the various hedgehog ligands through specific binding, we transfected COS-7 cells or human 293 embryonic kidney cells with PTCH or PTCH2 expression constructs and analyzed the cells for binding of the N-terminal biologically active domain of Shh, Dhh, and Ihh. Immunofluorescence analysis of transiently transfected COS-7 cells indicates that both PTCH and PTCH2 are capable of binding Shh-IgG and Dhh-IgG (Fig. 2a). Interestingly, even though both PTCH and PTCH2 were expressed at similar levels as determined by using an antibody directed to the Flag epitope tag inserted at the C terminus of these molecules, the staining was weaker for ligand binding to PTCH2 as compared with PTCH and not all cells expressing PTCH2 were labeled with Shh-IgG or Dhh-IgG, indicating that PTCH2 may not be accessible to the immunoadhesin on the cell surface. Because of this, no detectable binding of 125I-Shh to cells transiently transfected with PTCH2 could be detected.
Figure 2.
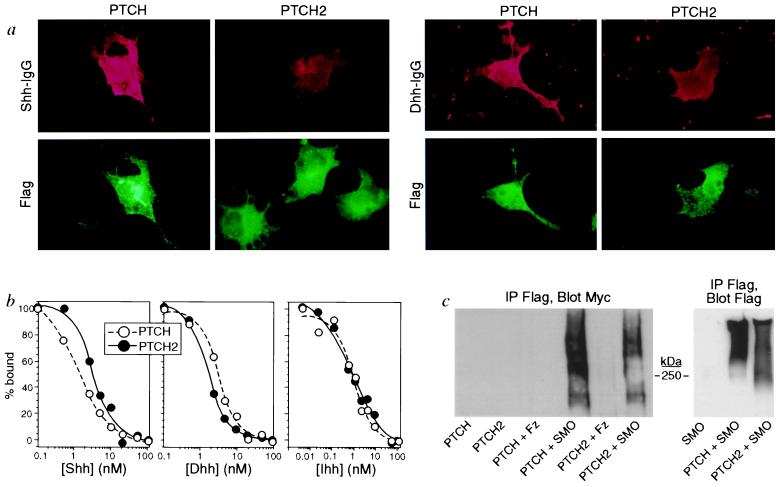
PTCH2 interacts with hedgehogs and SMO. (a) Immunostaining of COS-7 cells transfected with PTCH or PTCH2 and labeled with Shh-IgG or Dhh-IgG (red), and antibody directed against the Flag epitope (green). No labeling was detected in cells transfected with a control vector (data not shown). (b) Competition binding of recombinant murine 125I-Shh to 293 cells overexpressing hPTCH or hPTCH2. There was no detectable binding to mock-transfected cells (data not shown). (c) Coimmunoprecipitation of epitope-tagged PTCH or PTCH2 with epitope-tagged SMO. Immunoprecipitation was performed with antibodies to the Flag-tagged PTCH and analyzed by Western blotting on a 6% acrylamide gel with antibodies to the Myc-tagged SMO. Protein complexes can be detected with both PTCH and PTCH2. PTCH and PTCH2 express at similar levels as shown by immunoprecipitation by using antibodies to the Flag tag and by Western blot by using the same anti-Flag antibody.
To further analyze binding of the various hedgehog proteins to PTCH and PTCH2, we have established stable 293 cells lines expressing PTCH or PTCH2 and selected for receptor-expressing clones by PCR analysis and cell surface 125I-Shh binding. As shown in Fig. 2b, binding of 125I-Shh to these cell lines can be competed with an excess of Shh, Dhh, or Ihh. Scatchard analysis of the displacement curves indicates that all hedgehogs have a similar affinity for PTCH (Shh, 1.0 nM; Dhh, 2.6 nM; Ihh, 1.0 nM) and PTCH2 (Shh, 1.8 nM; Dhh, 0.6 nM; Ihh, 0.4 nM), indicating that both PTCH and PTCH2 can serve as physiological receptors for the three mammalian hedgehog proteins.
PTCH and PTCH2 Form a Complex with SMO.
We next determined whether, like PTCH, PTCH2 can physically associate with SMO, the signaling component of the hedgehog receptor. Expression constructs for Flag-tagged PTCH or PTCH2 were cotransfected into 293 cells with Myc-tagged SMO. As described previously (22), in cells expressing PTCH and SMO, PTCH can be immunoprecipitated with antibodies against the epitope-tagged SMO (Fig. 2c). Similarly, PTCH2 can be immunoprecipitated with antibodies against the epitope-tagged SMO (Fig. 2c) when the two proteins are coexpressed in 293 cells while a distantly related serpentine receptor, rat Frizzled 5, did not associate with PTCH or PTCH2 (Fig. 2c). These data indicate that PTCH2 can associate with SMO to form a multicomponent hedgehog receptor complex similar to the one described for PTCH (22).
Tissue Distribution of PTCH2.
To investigate whether PTCH2 could mediate the function of a specific hedgehog molecule based on a defined expression profile, we have compared the expression pattern of PTCH and PTCH2. First, Northern blot analysis using probes specific for PTCH and PTCH2 revealed high levels of PTCH2 mRNA only in the testis where PTCH was not detected (Fig. 3; ref. 18). By this method, PTCH2 expression was not detected in embryonic tissue in which PTCH is expressed. In situ hybridization of E18 mouse embryos revealed transcripts for PTCH2 in tissues such as the gut and teeth, where it is coexpressed with PTCH, although at lower levels (Fig. 4 a and b). On the contrary, higher levels of PTCH2 expression compared with PTCH were detected in epithelial cells of the skin (Fig. 4 c and d). To determine whether PTCH2 expression was up-regulated in response to hedgehog signaling, we have analyzed skin sections of transgenic mice expressing a constitutively active form of SMO (SMO-M2) in basal cells under the control of the keratin 5 promoter (28). As shown in Fig. 4 e and f, PTCH is strongly up-regulated in the basal cells of SMO-M2 transgenic mice while the levels of PTCH2 appear to be only slightly increased compared with wild-type levels. Similarly, we could not detect up-regulation of PTCH2 in cell lines such as C3H10T1/2 and Ref52, where PTCH is up-regulated in response to Shh stimulation (data not shown). Since PTCH2 has been reported to be up-regulated in response to Shh in newt animal caps (29), the regulation mechanism of PTCH and PTCH2 may be cell type-specific.
Figure 3.

Northern blot analysis of mouse PTCH and PTCH2. Northern blots containing RNA from multiple mouse tissues, as indicated, were hybridized with a cDNA fragment corresponding to the 3′ untranslated region of mPTCH or mPTCH2.
Figure 4.

In situ hybridization analysis of PTCH, PTCH2, and Fused. Emulsion autoradiographs after in situ hybridization of sagittal sections through embryonic day 18 (E18) mouse for PTCH (a) or PTCH2 (b) probes, transverse sections through E18 wild-type or SMO-M2 transgenic mouse skin for PTCH (c and e) and PTCH2 (d and f), and sections of adult mouse testis for PTCH (g), PTCH2 (h and k), Fused (i and l), and Dhh (j). [Bar = 2.1 mm (a and b); 0.065 mm (c– f, j); 0.12 mm (h and i); 1.0 mm (k and l).] bn, bone; gt, gut; sk, skin; th, teeth; vib, vibrissae; ep, epidermis; de, dermis; fol, hair follicle.
More detailed analysis of the expression pattern of PTCH and PTCH2 in the testis shows that high levels of PTCH2 are expressed inside the seminiferous tubules, on the primary and secondary spermatocytes (Fig. 4 h and k) while only low levels of PTCH can be detected on the Leydig cells located in the interstitium of the seminiferous tubules (Fig. 4g). The primary and secondary spermatocytes are in close contact with the supporting Sertoli cells, the source of Dhh in the testis (Fig. 4j) (15). To determine which one of the two receptors is a likely mediator of Dhh activity in the testis, we have analyzed the expression profile of a mouse homologue of the Fused kinase (M. Murone, J. Zhang, A.R., and F.J.d.S., unpublished data), a component of the hedgehog signaling pathway (30). Consistent with the idea that PTCH2 mediates the signal of Dhh in the testis, we found that Fused is expressed only in germ cells in which it colocalizes with PTCH2 (Fig. 4 i and l).
DISCUSSION
The secreted hedgehog proteins have emerged as a family of molecules involved in the development of the nervous system, limbs, bones, skin, and germ cells. Recently, they also have been implicated in a number of pathologies, including basal cell carcinoma, medulloblastoma, glioblastoma, and holoproencephaly (31–35). The function of the hedgehog ligands appears to be mediated at the cell surface by the PTCH-SMO receptor complex. Current models suggest that binding of Shh to PTCH prevents the normal inhibition of the seven-transmembrane-protein Smoothened (SMO) by PTCH. According to this model, the inhibition of SMO signaling is relieved after mutational inactivation of PTCH in the basal cell nevus syndrome. The recent identification of PTCH2, a molecule with sequence homology to PTCH, raises the possibility that multiple receptors mediate the effects of the hedgehog ligands. To investigate this possibility, we have isolated the human PTCH2 gene. We demonstrate here that both PTCH and PTCH2 are genuine hedgehog receptors capable of recognizing the various hedgehog ligands (Sonic, Desert, and Indian) with similar affinity. Interestingly, transient transfection of 293 or Cos-7 cells with PTCH2 resulted in a lower number of cells displaying weaker Shh-IgG or Dhh-IgG binding at the cell surface compared with PTCH even though both molecules appeared to be expressed at similar levels. These results may indicate that PTCH2 requires additional, cell-specific, folding or transport protein to reach the surface. The existence of such cell-specific molecules has been reported, for example, for the odorant receptor in C. elegans (36) and for the calcitonin-receptor-like receptor (37). In addition, the presence of accessory molecules could alter the specificity of the receptor for one ligand versus another (37) and could modify the affinity of PTCH molecules toward one of the hedgehog ligands.
Coimmunoprecipitation experiments demonstrate that, like PTCH, PTCH2 physically associates with SMO, suggesting that PTCH2 forms a multicomponent hedgehog receptor complex with SMO similar to the one described for PTCH (22). Interestingly, these results also demonstrate that the long C-terminal tail that is missing in PTCH2 is not required for the interaction with SMO, as was already suggested by the analysis of a truncated PTCH (22). However, it remains possible that the absence of a C-terminal domain affects the capacity of PTCH2 to block SMO signaling or leads to other differences in signaling by PTCH compared with PTCH2.
Because the two patched receptors do not seem to discriminate the various mammalian hedgehog ligands based on their affinity, we investigated whether they could mediate the function of specific hedgehog ligands based on their expression profile. Interestingly, the tissue distribution of these two receptors does not fully overlap. While PTCH is expressed broadly, high levels of PTCH2 expression was detected only in the skin and in the testis. These results suggest that in vivo, while PTCH may be the primary mediator of the broad range of Shh activities, PTCH2 may have a more restricted function. In particular, PTCH2 is expressed on spermatocytes, where it colocalizes with other hedgehog signaling molecules such as Fused and members of the Gli family of zinc finger transcription factors (38). PTCH2 therefore is likely to be involved in mediating the function of Dhh in germ cells. Dhh is required for proper differentiation of germ cells, and male Dhh-deficient mice are sterile because of lack of mature sperm (15). Confirmation of the role in spermatogenesis of PTCH2 on germ cells as well as the function of PTCH expressed in the absence of Fused and Gli on testosterone-producing Leydig cells is likely to be elucidated by knockout studies.
Loss of heterozygosity for PTCH was reported to occur at high frequency in familial as well as sporadic basal cell carcinoma (31–34), suggesting that it functions as a tumor suppressor. According to the receptor model described above, loss of PTCH function may result in aberrant signaling by SMO, leading to hyperproliferation of the skin basal cells. If, as suggested above, PTCH2 mediates the function of Dhh, loss of PTCH2 may lead to tumor formation in tissues where SMO activity is controlled by PTCH2. Interestingly, recent analysis of recurrent chromosomal abnormalities in testicular tumors, including seminomas, revealed a deletion of the region 1p32–36 (39), where the PTCH2 gene is localized. Loss of this region encompassing the PTCH2 locus was consistent in 36% of the germ cell tumor cases. These data raise the possibility that, as with PTCH in basal cell carcinoma and medulloblastoma, PTCH2 may be a tumor suppressor in Dhh target cells such as spermatocytes, further implicating hedgehog signaling in cancer.
Acknowledgments
We thank D. Yansura, S. Spencer, and G. Bennett for expression, purification, and labeling of Shh, Dhh, and Ihh, the Genentech DNA sequencing and DNA synthesis labs, and W. Anstine for preparing the figures.
ABBREVIATIONS
- SMO
Smoothened
- PTCH
Patched
- Shh
Sonic hedgehog
- Dhh
Desert hedgehog
- Ihh
Indian hedgehog
Footnotes
Data deposition: The sequence reported in this paper has been deposited in the GenBank database [accession no. AF091501 (human PTCH2)].
References
- 1.Nusslein-Volhard C, Wieschaus E. Nature (London) 1980;287:795–801. doi: 10.1038/287795a0. [DOI] [PubMed] [Google Scholar]
- 2.Perrimon N. Cell. 1995;80:517–520. doi: 10.1016/0092-8674(95)90503-0. [DOI] [PubMed] [Google Scholar]
- 3.Hammerschmidt M, Brook A, McMahon A P. Trends Genet. 1997;13:14–21. doi: 10.1016/s0168-9525(96)10051-2. [DOI] [PubMed] [Google Scholar]
- 4.Echelard Y, Epstein D J, St. Jacques B, Shen L, Mohler J, McMahon J A, McMahon A P. Cell. 1993;75:1417–1430. doi: 10.1016/0092-8674(93)90627-3. [DOI] [PubMed] [Google Scholar]
- 5.Riddle R D, Johnson R L, Laufer E, Tabin C. Cell. 1993;75:1401–1416. doi: 10.1016/0092-8674(93)90626-2. [DOI] [PubMed] [Google Scholar]
- 6.Roelink H, Augsburger A, Heemskerk J, Korzh V, Norlin S, Ruiz i Altaba A, Tanabe Y, Placzek M, Edlund T, Jessell T M, et al. Cell. 1994;76:761–775. doi: 10.1016/0092-8674(94)90514-2. [DOI] [PubMed] [Google Scholar]
- 7.Marti E, Bumcrot D A, Takada R, McMahon A P. Nature (London) 1995;375:322–325. doi: 10.1038/375322a0. [DOI] [PubMed] [Google Scholar]
- 8.Ericson J, Muhr J, Placzek M, Lints T, Jessell T M, Edlund T. Cell. 1995;81:747–756. doi: 10.1016/0092-8674(95)90536-7. [DOI] [PubMed] [Google Scholar]
- 9.Hynes M, Porter J A, Chiang C, Chang D, Tessier-Lavigne M, Beachy P A, Rosenthal A. Neuron. 1995;15:35–44. doi: 10.1016/0896-6273(95)90062-4. [DOI] [PubMed] [Google Scholar]
- 10.Krauss S, Concordet J P, Ingham P W. Cell. 1993;75:1431–1444. doi: 10.1016/0092-8674(93)90628-4. [DOI] [PubMed] [Google Scholar]
- 11.Laufer E, Nelson C E, Johnson R L, Morgan B A, Tabin C. Cell. 1994;79:993–1003. doi: 10.1016/0092-8674(94)90030-2. [DOI] [PubMed] [Google Scholar]
- 12.Fan C M, Tessier-Lavigne M. Cell. 1994;79:1175–1186. doi: 10.1016/0092-8674(94)90009-4. [DOI] [PubMed] [Google Scholar]
- 13.Johnson R L, Laufer E, Riddle R D, Tabin C. Cell. 1994;79:1165–1173. doi: 10.1016/0092-8674(94)90008-6. [DOI] [PubMed] [Google Scholar]
- 14.Vortkamp A, Lee K, Lanske B, Segre G V, Kronenberg H M, Tabin C J. Science. 1996;273:613–622. doi: 10.1126/science.273.5275.613. [DOI] [PubMed] [Google Scholar]
- 15.Bitgood M J, Shen L, McMahon A P. Curr Biol. 1996;6:298–304. doi: 10.1016/s0960-9822(02)00480-3. [DOI] [PubMed] [Google Scholar]
- 16.Bitgood M J, McMahon A P. Dev Biol. 1995;172:126–138. doi: 10.1006/dbio.1995.0010. [DOI] [PubMed] [Google Scholar]
- 17.Nakano Y, Guerrero I, Hildalgo A, Taylor A, Whittle J R S, Ingham P W. Nature (London) 1989;341:508–513. doi: 10.1038/341508a0. [DOI] [PubMed] [Google Scholar]
- 18.Goodrich L V, Johnson R L, Milenkovic L, McMahon J A, Scott M P. Genes Dev. 1996;10:301–312. doi: 10.1101/gad.10.3.301. [DOI] [PubMed] [Google Scholar]
- 19.Marigo V, Scott M P, Johnson R L, Goodrich L V, Tabin C J. Development (Cambridge, UK) 1996;122:1225–1233. doi: 10.1242/dev.122.4.1225. [DOI] [PubMed] [Google Scholar]
- 20.van den Heuvel M, Ingham P W. Nature (London) 1996;382:547–551. doi: 10.1038/382547a0. [DOI] [PubMed] [Google Scholar]
- 21.Alcedo J, Ayzenzon M, Von Ohlen T, Noll M, Hooper J E. Cell. 1996;86:221–232. doi: 10.1016/s0092-8674(00)80094-x. [DOI] [PubMed] [Google Scholar]
- 22.Stone D M, Hynes M, Armanini M, Swanson T A, Gu Q, Johnson R L, Scott M P, Pennica D, Goddard A, Phillips H, et al. Nature (London) 1996;384:129–134. doi: 10.1038/384129a0. [DOI] [PubMed] [Google Scholar]
- 23.Marigo V, Davey R A, Zuo Y, Cunningham J M, Tabin C J. Nature (London) 1996;384:176–179. doi: 10.1038/384176a0. [DOI] [PubMed] [Google Scholar]
- 24.Chen Y, Struhl G. Cell. 1996;87:553–563. doi: 10.1016/s0092-8674(00)81374-4. [DOI] [PubMed] [Google Scholar]
- 25.Motoyama J, Takabatake T, Takeshima K, Hui C-C. Nat Genet. 1998;18:104–106. doi: 10.1038/ng0298-104. [DOI] [PubMed] [Google Scholar]
- 26.Phillips H S, Hains J M, Laramee G R, Rosenthal A, Winslow J W. Science. 1990;250:290–294. doi: 10.1126/science.1688328. [DOI] [PubMed] [Google Scholar]
- 27.Melton D A, Krieg P A, Rebagliati M R, Maniatis T, Zinn K, Green M R. Nucleic Acids Res. 1984;12:7035–7052. doi: 10.1093/nar/12.18.7035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Xie J, Murone M, Luoh S M, Ryan A, Gu Q, Zhang C, Bonifas J M, Lam C W, Hynes M, Goddard A, et al. Nature (London) 1998;391:90–92. doi: 10.1038/34201. [DOI] [PubMed] [Google Scholar]
- 29.Takabatake T, Ogawa M, Takahashi T C, Mizuno M, Okamoto M, Takeshima K. FEBS Lett. 1997;410:485–489. doi: 10.1016/s0014-5793(97)00645-5. [DOI] [PubMed] [Google Scholar]
- 30.Preat T, Therond P, Lamour-Isnard C, Limbourg-Bouchon B, Tricoire H, Erk I, Mariol M C, Busson D. Nature (London) 1990;347:87–89. doi: 10.1038/347087a0. [DOI] [PubMed] [Google Scholar]
- 31.Johnson R L, Rothman A L, Xie J, Goodrich L V, Bare J W, Bonifas J M, Quinn A G, Myers R M, Cox D R, Epstein E H, Jr, Scott M P. Science. 1996;272:1668–1671. doi: 10.1126/science.272.5268.1668. [DOI] [PubMed] [Google Scholar]
- 32.Hahn H, Wicking C, Zaphiropoulous P G, Gailani M R, Shanley S, Chidambaram A, Vorechovsky I, Holmberg E, Unden A B, Gillies S, et al. Cell. 1996;85:841–851. doi: 10.1016/s0092-8674(00)81268-4. [DOI] [PubMed] [Google Scholar]
- 33.Gailani M R, Stahle-Backdahl M, Leffell D J, Glynn M, Zaphiropoulos P G, Pressman C, Unden A B, Dean M, Brash D E, Bale A E, Toftgard R. Nat Genet. 1996;14:78–81. doi: 10.1038/ng0996-78. [DOI] [PubMed] [Google Scholar]
- 34.Xie J, Johnson R L, Zhang X, Bare J W, Waldman F M, Cogen P H, Menon A G, Warren R S, Chen L C, Scott M P, Epstein E H., Jr Cancer Res. 1997;57:2369–2372. [PubMed] [Google Scholar]
- 35.Roessler E, Belloni E, Gaudenz K, Jay P, Berta P, Scherer S W, Tsui L C, Muenke M. Nat Genet. 1996;14:357–360. doi: 10.1038/ng1196-357. [DOI] [PubMed] [Google Scholar]
- 36.Dwyer N D, Troemel E R, Sengupta P, Bargmann C I. Cell. 1998;93:455–466. doi: 10.1016/s0092-8674(00)81173-3. [DOI] [PubMed] [Google Scholar]
- 37.McLatchie L M, Fraser N J, Main M J, Wise A, Brown J, Thompson N, Solari R, Lee M G, Foord S M. Nature (London) 1998;393:333–339. doi: 10.1038/30666. [DOI] [PubMed] [Google Scholar]
- 38.Persengiev S P, Kondova I I, Millette C F, Kilpatrick D L. Oncogene. 1997;14:2259–2264. doi: 10.1038/sj.onc.1201068. [DOI] [PubMed] [Google Scholar]
- 39.Summersgill B, Goker H, Weber-Hall S, Huddart R, Horwich A, Shipley J. Br J Cancer. 1998;77:305–313. doi: 10.1038/bjc.1998.47. [DOI] [PMC free article] [PubMed] [Google Scholar]