

Abstract
A cell’s ability to effectively communicate with a neighboring cell is essential for tissue function and ultimately for the organism to which it belongs. One important mode of intercellular communication is the release of soluble cyto- and chemokines. Once secreted, these signaling molecules diffuse through the surrounding medium and eventually bind to neighboring cell’s receptors whereby the signal is received. This mode of communication is governed both by physicochemical transport processes and cellular secretion rates, which in turn are determined by genetic and biochemical processes. The characteristics of transport processes have been known for some time, and information on the genetic and biochemical determinants of cellular function is rapidly growing. Simultaneous quantitative analysis of the two is required to systematically evaluate the nature and limitations of intercellular signaling. The present study uses a solitary cell model to estimate effective communication distances over which a single cell can meaningfully propagate a soluble signal. The analysis reveals that: (i) this process is governed by a single, key, dimensionless group that is a ratio of biological parameters and physicochemical determinants; (ii) this ratio has a maximal value; (iii) for realistic values of the parameters contained in this dimensionless group, it is estimated that the domain that a single cell can effectively communicate in is ≈250 μm in size; and (iv) the communication within this domain takes place in 10–30 minutes. These results have fundamental implications for interpretation of organ physiology and for engineering tissue function ex vivo.
Keywords: cell signaling/bioreactor design, tissue engineering
Study of the molecular biology of signal transduction is advancing rapidly. The key genes, their role, and their interrelatedness are being deciphered and established. In fact, it is expected that, within only a few years, the entire genomes of key organisms will be completely sequenced, ORFs will be established, and gene assignment will be made (1–3). This avalanche of information has lead to a rapidly growing field known as “bioinformatics” (4, 5). It is thus expected, in the foreseeable future, that all of the molecular determinants of cellular function will become known and characterized.
What lies beyond bioinformatics? It is becoming clear that systemic and kinetic analysis of this molecular information and the cellular processes that it underlies is needed (6). In particular, it is important to quantitatively characterize and study the nature of cellular signaling processes. One important mode of intercellular communication is the release of soluble cyto- and chemokines. Once secreted, these signaling molecules diffuse through the surrounding medium and eventually bind to neighboring cell’s receptors whereby the signal is received. This mode of communication is governed both by physicochemical transport processes and cellular secretion rates, which in turn are determined by genetic and biochemical processes. Estimates for the time scale on which signaling takes place and the effective communication distances are needed. These estimates will be important for the interpretation of cell and organ physiology and for effective reconstruction of tissue function ex vivo.
Cellular therapies promise to become an important therapeutic modality in the near future. Many such therapeutic applications rely on the growth of primary cells and tissues ex vivo (7, 8). Tissues are comprised of “functional units,” such as the nephron in the kidney and the villi in the small intestine. The functional units of tissues contain many different cell types that continuously communicate with one another through a variety of mechanisms. A key mode of communication is the release of soluble cyto- and chemokines. These signals lead to the induction of organogenic processes, such as cell proliferation, differentiation, and motion (9, 10). These organogenic processes in turn influence the development of a tissue culture, including the cell types and numbers that are produced.
When primary cells are aspirated or biopsied and placed in culture, they are removed from their normal physiological environment. They respond by trying to reconstitute tissue function in the environment in which they are placed. A cell’s ability to propagate signals to neighboring cells and receive signals from them is critical in this process. The extracellular propagation of the signal is by random molecular motion of the soluble cyto- and chemokines. Although the cells cannot control diffusion, they can regulate their biological processes such as protein production and secretion rates. In culture, this communication process can be influenced by initial conditions, such as innocula density, that are important in determining how the tissue culture will evolve (11, 12).
The present study focused on the fundamentals of the intercellular signaling processes with the goal of defining characteristic length and time scales. The definition of key, dimensionless groupings of parameters was sought also.
METHODS
Theoretical.
Fundamental models for diffusion processes are used (13, 14) for which analytical solutions are available (15). It is also possible to obtain solutions to the diffusion models by using solutions to the analogous heat conduction problems. Comprehensive compendium of such solutions is available (16).
Numerical.
Solutions were obtained by evaluating the derived analytical expressions using standard and readily available software packages: Microsoft excel Ver. 5.0 for spreadsheet computations and mathematica Ver. 3.0 for symbolic calculations. Symbolic arithmetic and series expansions were evaluated using mathematica Ver. 3.0 (17).
ANALYSIS AND RESULTS
Cyto/Chemokine Signaling by a Single “Solitary” Cell.
We considered a single solitary cell and examined how far it could effectively propagate secreted cyto/chemokine signals. Propagation of signals from a suspended solitary cell can be described as a single spherical surface source (Fig. 1). The diffusion equation that describes the time dependent mass transport from a spherical source is (13):
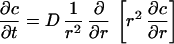 |
1 |
where c is the concentration of the cyto/chemokine and D is the diffusion coefficient. The appropriate boundary conditions are a constant flux at the cell surface and complete dilution very far from the cell; mathematically stated as:
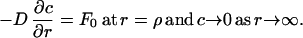 |
where F0 is the cyto/chemokine production rate (molecules/area⋅time) and ρ is the cell radius. It is assumed that the secreted molecule is not reconsumed by the solitary cell and that F0 is uniform over the surface of the cell. However, it is possible that secretion is asymmetric and is restricted to specific regions of the cell’s membrane or that cytokines are concentrated and stored for later release, resulting in the ejection of “plumes” from the cell (18). Analysis of the secretion of such plumes follows the same procedure as presented below.
Figure 1.

A schematic illustrating a solitary cell secreting a soluble cyto/chemokine and the concentration gradient that results from the secretion.
The concentration of cyto/chemokine in the medium surrounding the cell is assumed to be 0 before the start of secretion, leading to the initial condition: c = 0 for t ≤ 0.
Solution and Scaling.
We can express the solution to the above stated problem as [equation 10.4 (11) in ref. 16]:
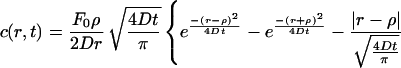 |
2 |
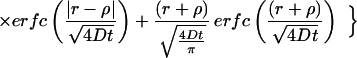 |
where erfc is the complementary error function. Thus, the concentration of the cyto/chemokine can be calculated as a function of spatial location and time.
The behavior of this solution can be characterized in terms of characteristic time and length scales. To define these scales, we need to nondimensionalize the solution. The reference length scale is chosen to be the cell radius ρ, and the reference time scale is chosen to be the corresponding diffusion time tdiff = ρ2/4D. This casts the process in terms of cellular dimensions.
The dimensionless length and time variables are:
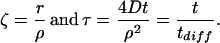 |
3 |
The solution can be written as:
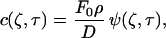 |
4 |
with a dimensionless function of length and time, Ψ, defined by:
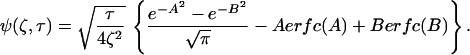 |
5 |
where
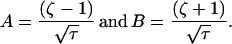 |
6 |
We can now define and estimate values for the characteristic time and length scales that characterize the intercellular signaling process.
Definition and Estimation of a Characteristic Time Constant.
It can be readily shown, for long times, as the process reaches a steady state, that
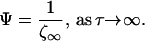 |
7 |
This limiting behavior represents the maximum concentration buildup at a given distance from the secreting cell. However, reaching this limit requires an infinite amount of time. We are therefore interested in determining a time constant that characterizes the concentration buildup in relation to physiologically relevant events. We can define a characteristic time constant, τf, as the time at which the function Ψ will be some fraction, f, of the steady state value as:
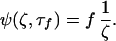 |
8 |
The solution to Eq. 8 may be evaluated numerically, and it is shown in Fig. 2. The nature of this solution for large ζ (ζ > ≈4–5) suggests a relation between τf and ζ of the type:
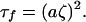 |
9 |
Upon substituting this suggested solution back into Eq. 8, expanding Ψ in a series, and evaluating the limit for large ζ, a relationship between a and f is found:
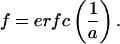 |
10 |
Therefore, a characteristic time constant can be directly determined for chosen values of f at any distance ζ away from the secreting solitary cell. We chose a value for f of 0.5, or the time it takes for the signal to reach half of its maximum strength. This results in the definition of a time constant, τ0.5:
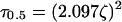 |
11 |
Measurements of the diffusion constant, D, have shown it to range from 1⋅10−6 to 1⋅10−7 cm2/s for signaling molecules in the 10–100 kDa range (19). Signaling proteins typically are small, and thus the expected diffusion coefficient for them is toward the upper end of the numerical range given. Using these physical constants, dimensional values of the time constant can be tabulated (Table 1).
Figure 2.

The numerical solution to the location-dependent time constant. This time constant is defined as the time it takes to reach half of the ultimate concentration at a particular distance. The dashed line represents τ0.5 = (2.097ζ)2
Table 1.
Time constants for intercellular communication at different distances from the secreting cell
| r, μm |
t, min
|
|
|---|---|---|
| D = 10−6 (fast protein) | D = 10−7 (slow protein) | |
| 75 | 1 | 10 |
| 150 | 4 | 40 |
| 235 | 10 | 100 |
| 330 | 20 | 200 |
| 570 | 60 | 600 |
Definition and Estimation of Effective Signal Propagation Distance.
Having a definition for the time it takes to reach a given concentration at a particular distance away from the secreting cell, we now turn to the question of how far can a meaningful signal can be propagated. The concentration of a signaling molecule is meaningful to the responding cell relative to its Km value for that particular protein. We can then evaluate how a meaningful signal propagates with time by equating c to this critical concentration Km:
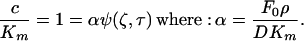 |
12 |
At a given time, this equation can tell us how far a meaningful signal has propagated. For a given value of α, it can be solved for ζ, the distance from the secreting cell that the leading edge of the signal has propagated to as a function of time (Fig. 3).
Figure 3.

The propagation distance ζ of soluble cyto/chemokine signal over time. As indicated in the Inset, this distance is defined as the location where c/Km is unity.
The maximal distance that a signal can propagate is at steady-state. Upon substitution of Eq. 7 into Eq. 12, we obtain:
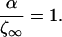 |
13 |
Therefore, the maximum signal propagation distance ζ∞ is equal to the parameter α. This distance is only reached after an infinite amount of time.
However, cells do not have an infinite length of time to propagate a signal. What is required is a definition of an “effective” communication distance. We can define the effective communication distance as the distance that a signal can travel in one time constant, τ0.5. We can illustrate this definition by plotting c/Km as a function of ζ for a fixed value of α and τ as shown in Fig. 4. Whenever c/Km exceeds unity, the signal is received by a neighboring cell. The effective communication distance ζ0.5 is indicated where the c/Km curve at time τ0.5 drops below unity, which occurs at:
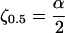 |
14 |
Thus, the effective communication distance is directly proportional to the dimensionless group α. Analogously, the maximum communication distance ζ∞ occurs at a similar point on the curve for the steady-state solution.
Figure 4.

Schematic representation of communication distances, illustrating the definition of the effective communication distance ζ0.5 and the maximum communication distance ζ∞. Eq. 12 is plotted for α = 100, using the definition for the location dependent time constant as well as for the steady state solution.
At first glance, one might expect that faster diffusion would lead to longer communication distances. However, this analysis reveals just the opposite, with slower diffusion resulting in a higher numerical value for α. This permits the propagation of a more intense signal before random molecular motion dilutes the secreted molecules.
Interpretation of α and Estimation of its Numerical Value.
The estimate for the effective communication distance is directly proportional to the parameter α. This key dimensionless group is proportional to the ratio of two time constants, those for cyto/chemokine diffusion and secretion:
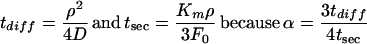 |
15 |
The diffusion coefficient, D, is purely a physicochemical property, a constraint that the signaling cell has to operate under. Conversely, the secretion time constant contains biologically determined parameters: the secretion rate F0 and the target cell’s receptor binding constant Km. F0 can be regulated by the secreting cell, but it has an upper bound (20). Km, on the other hand, has a minimum value required to elicit a response in the target cell (21). Therefore, the secretion rate constant will have a minimum value, resulting in a maximum achievable value for α. Let us now examine the numerical values of these parameters and their implications.
A minimum value for Km corresponds to a high receptor affinity for the signaling molecule. The high end receptor chemical affinity for a wide range of human cytokines is 10–50 pM (22). Functional studies of cellular response to these growth factors (21, 23–25) show that the cytokine concentration required for a 50% of maximum growth rate falls in the same 10–50 pM range.
A key biologic parameter is the secretion rate of the signaling protein from the signaling cell. An estimate is provided by Savinell et al. (20), whose analysis includes such variables as mRNA half-life and the velocity at which RNA polymerase transcribes the gene of interest. Maximum secretion rates fall into the range of 2300–8000 molecules/cell⋅s. These values are then converted from a per cell production rate to a secretion rate per unit membrane area, F0.
Based on these values, as well as those previously stated for D and ρ, the maximum value for α is estimated to be on the order of 100. Therefore, the effective intercellular distances are ≈50 cell radii or 25 cell diameters. From Eq. 11 and Table 1, one can deduce that the characteristic time constant for intercellular communication by soluble cyto/chemokines is on the order of 10–30 minutes.
Signal Propagation from a Single Adherent Cell.
The domain that the signaling cell secretes its cyto/chemokine molecules into influences the effective communication distance. The above results are derived for a suspended solitary cell. Similar analyses can be performed for single cells sitting on a surface, by an edge of a culture well, or in the corner of a culture flask.
If a cell adheres to a growth surface, it secretes into a hemispherical fluid domain. The maximum number of signaling molecules that are derived from a single cell is a constant. Thus, F0 doubles and so does α, and therefore the effective communication distance is twice as long. If the cell was located next to a wall in the culture device, the fluid domain would be reduced to one–fourth compared with a suspended cell. Therefore, the effective communication distance would increase by a factor of 4 compared with a suspended cell. Likewise, a cell in a corner would have an effective communication distance that is eight times longer than a suspended cell. For primary cell cultures in which microenvironmental factors are important, the location of a cell relative to the walls of the culture device is therefore critical.
DISCUSSION
The present study used a solitary cell model to estimate the effective communication distance over which a single cell can meaningfully propagate a cyto/chemokine-mediated signal. The analysis revealed that: (i) the intercellular signaling process is governed by a single, key, dimensionless group, α, that is comprised of biological parameters as well as physicochemical determinants; (ii) α has a maximal value; (iii) for realistic values of the parameters contained in α, it is estimated that the domain within which a solitary cell can effectively communicate is ≈250 μm in size; and (iv) the communication within this domain takes place in 10–30 minutes.
The definition of key, dimensionless property ratios and their interpretation has played an important role in the development of engineering science (13), and dimensionless ratios such as α are likely to become fundamental quantities in the emerging area of cellular bioengineering (21). The dimensionless property α is a ratio of two time constants: for secretion and for diffusion. It measures the relative rates at which a cyto/chemokine builds up close to the membrane of the secreting cells vs. how quickly it diffuses away. If the time constant for secretion is slow compared with diffusion, then insignificant concentration will build up near the signaling cell, and virtually no effective signaling takes place. Conversely, if the diffusion process is slow compared with the secretion of the cyto/chemokine, then a significant concentration can build up around the signaling cell. Because diffusion times are determined by physics, effective signaling depends on the cell’s ability to rapidly secrete the signaling molecule.
The secretion time constant is shown to have a minimum value due to the maximal constraints on the achievable secretion rate and the minimal values for the binding constant of the receiving cells. The maximal secretion rate of a single gene has been estimated to be on the order of a few thousand molecules per cell per second for constitutive secretion (20). The secreting cell can increase its secretion rate by gene amplification. Secretion rates can also be increased, albeit temporarily, by release of cytoplasmic reserves of a stored signaling molecule. There is some evidence that hematopoietic cells take advantage of the second mechanism (18). In many instances, the signaling molecule adsorbs to binding sites in the extracellular matrix. If this binding is irreversible, such binding leads to a loss of signaling molecules, thus shortening the signal propagation distance over a given time period. There has been analysis of this effect, its biological implications have been discussed (26). Similarly, other biological processes may shorten the effective communication distance, including desensitization of receptor proteins (27) (leading to lower Km values) or ligand-induced receptor modification (28). Finally, any convective flow will influence the communication distance. It has been shown recently that the propagation of antibodies in the brain can be significantly augmented by modest convective flows (29, 30).
It is important to note that the relative secretion rate and diffusion rates also determine the concentrations at the cell surface and thus the efficacy of auto- and juxtacrine mechanisms. The dimensionless group α must exceed unity to allow for autocrine and juxtacrine signaling to occur. The maximal local concentration can be calculated from Eq. 4 to be on the order of 10–50 ng/ml. This result may have important implications for the interpretation of the biological effects that are being described for the ultra high concentrations of growth factor required for the stimulation of hematopoietic stem cells (31). This study reports that over 300 ng/ml are needed to yield maximum stimulation of hematopoietic stem cell self-renewal by the flt-3 ligand. Such a high concentration would be hard, if not impossible, to accomplish and sustain by a secreted mechanisms. An alternatively spliced and membrane-displayed form of the flt-3 ligand by a neighboring cell would, on the other hand, enable strong juxtacrine signaling (32). Thus, the analysis presented would argue that the biological effects of such high growth factor concentrations are accomplished by direct cell–cell contact, which is consistent with the well documented importance of accessory cells on hematopoietic stem cell behavior.
These considerations lead to interesting revelations about the limitations on how effectively cells can communicate with their neighbors. The nature of the diffusion and binding processes require high expression rates of the signaling molecule. Numerous studies focus on the measurement of mRNA for cyto/chemokines as indicators of whether a cell is signaling with a particular molecule of interest. The results herein indicate that only mRNA levels lead to high secretion rates and are indicative of an physiologically active signal.
The maximal estimates of α are such that the effective paracrine signaling distances are limited to ≈25 cell diameters. Of interest, this length scale is similar to those of functional sub units in tissues and that of developing embryos, such as that of Drosophila melanogaster. Concentration gradients are known to be important in these situations, and it has been shown experimentally that increasing the gene dosage of signaling molecules alters the signaling distances that govern morphogenesis (33, 34).
The time constant for paracrine communications within this domain is estimated to be 10–30 min. for small protein. Of interest, it has been determined that the turnover times of receptors falls into this range (21, 35). Thus, if the receiving cell is internalizing a bound signaling molecule at this rate, it must be supplied at an equal or higher rate for the signal to be transduced. The estimate arrived at for the time constant of signal delivery overlaps with this experimentally determined receptor recycling time constant.
There are several published experimental observations that are consistent with our communication distance estimates. A time-lapse study of the motility characteristics of bovine pulmonary artery endothelial cells (36) showed that cells appear to sense the presence of one another at distances of ≈40 μm, perhaps by communicating using soluble cyto/chemokines. Furthermore, it has been shown that hematopoietic progenitor cells are aware of each other at and below separation distances of ≈100 μm (37). Unpublished time-lapse results from our laboratory clearly show that hematopoietic progenitors (CD34+ cells) communicate without direct contact and that these communication distances are on the order of 100 μm.
The analysis presented is of fundamental importance to the emerging area of tissue engineering (8, 38–41), namely, the reconstruction of tissue function ex vivo. Similar analysis of diffusion distances in cell cultures, in which cells are grown on microspheres, has shown clearly the existence of critical communication distances leading to minimum inoculation densities in cultures of pure production cell lines (25). Furthermore, the differences in cell behavior close to and far away from an edge in hematopoietic bioreactors has been observed experimentally and has lead to the design of a radial flow device to eliminate such edge effects (42). Effective cellular inoculum densities will be influenced by the results presented, and, perhaps even more importantly, the design of any device to carry out a complex tissue culture will have to take these intercellular communication distances into account.
The definition, estimation, and understanding of the effective intercellular communication distances are of fundamental import in our understanding of organ physiology, in the significance of signaling pathways, and for effective tissue engineering. The study presented here defines, interprets, and evaluates a dimensionless quantity that is fundamental to these endeavors. The present analysis thus exemplifies and extends the influential analysis of the constraints that diffusion places on biochemical function at the cellular level (43).
Acknowledgments
This work was supported by grant NAG9–652 from the National Aeronautics and Space Administration.
References
- 1.Tatusov R L, Mushegian A R, Bork P, Brown N P, Hayes W S, Brodovsky M, Rudd K E, Koonin E V. Curr Biol. 1996;6:279–291. doi: 10.1016/s0960-9822(02)00478-5. [DOI] [PubMed] [Google Scholar]
- 2.Mushegian A R, Koonin E V. Proc Natl Acad Sci USA. 1996;93:10268–10273. doi: 10.1073/pnas.93.19.10268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Koonin E V, Mushegian A R, Rudd K E. Curr Biol. 1996;6:404–416. doi: 10.1016/s0960-9822(02)00508-0. [DOI] [PubMed] [Google Scholar]
- 4.Robinson, C., ed. (1996) Trends Biotechnol. 14, 1–483.
- 5.Walker, R. T., ed. (1996) Nucleic Acids Res. 24, 1–252.
- 6.Palsson B O. Nat Biotechnol. 1997;15:3–4. doi: 10.1038/nbt0197-3. [DOI] [PubMed] [Google Scholar]
- 7.McAdams T A, Miller W M, Papoutsakis E T. Trends Biotechnol. 1996;14:341–349. doi: 10.1016/0167-7799(96)10047-0. [DOI] [PubMed] [Google Scholar]
- 8.Palsson B O, Hubbell J A. In: The Biomedical Engineering Handbook. Bronzino J D, editor. Boca Raton, FL: CRC; 1995. pp. 1583–1824. [Google Scholar]
- 9.Edelman G M. Topobiology: An Introduction to Molecular Embryology. New York: Harper Collins; 1988. [Google Scholar]
- 10.Lauffenburger D A, Horwitz A F. Cell. 1996;84:359–369. doi: 10.1016/s0092-8674(00)81280-5. [DOI] [PubMed] [Google Scholar]
- 11.Levee M G, Lee G, Paek S, Palsson B O. Biotechnol Bioengineer. 1994;43:734–739. doi: 10.1002/bit.260430807. [DOI] [PubMed] [Google Scholar]
- 12.Caplan A I. Sci Am. 1984;251:84–87. doi: 10.1038/scientificamerican1084-84. , 90–94. [DOI] [PubMed] [Google Scholar]
- 13.Bird R B, Stewart W E, Lightfoot E N. Transport Phenomena. New York: Wiley; 1960. [Google Scholar]
- 14.Lightfoot E N. Transport Phenomena and Living Systems. New York: Wiley; 1974. [Google Scholar]
- 15.Crank J. The Mathematics of Diffusion. New York: Oxford Univ. Press; 1975. [Google Scholar]
- 16.Carslaw H S, Jaeger J C. Conduction of Heat in Solids. New York: Oxford Univ. Press; 1959. [Google Scholar]
- 17.Wolfram S. The Mathematica Book. Cambridge, U.K.: Cambridge Univ. Press; 1996. [Google Scholar]
- 18.Griffiths G M. Trends Cell Biol. 1996;6:329–332. doi: 10.1016/0962-8924(96)20031-5. [DOI] [PubMed] [Google Scholar]
- 19.Young M E, Carroad P A, Bell R L. Biotechnol Bioengineer. 1980;22:947–955. [Google Scholar]
- 20.Savinell J M, Lee G M, Palsson B O. Bioprocess Engineering. 1989;4:231–234. [Google Scholar]
- 21.Lauffenburger D A, Linderman J J. Receptors: Models for Binding, Trafficking, and Signaling. New York: Oxford Univ. Press; 1993. [Google Scholar]
- 22.Aggarwal B B, Gutterman J U. Human Cytokines: Handbook for Basic and Clinical Research. Boston: Blackwell Scientific; 1992. [Google Scholar]
- 23.Knauer D J, Wiley H S, Cunningham D D. J Biol Chem. 1984;259:5623–5631. [PubMed] [Google Scholar]
- 24.Gronthos S, Simmons P J. Blood. 1995;85:929–940. [PubMed] [Google Scholar]
- 25.Lauffenburger D A, Cozens C. Biotechnol Bioengineer. 1989;33:1365–1378. doi: 10.1002/bit.260331102. [DOI] [PubMed] [Google Scholar]
- 26.Chu L, Wiley H S, Lauffenburger D A. Tissue Engineering. 1996;2:17–38. doi: 10.1089/ten.1996.2.17. [DOI] [PubMed] [Google Scholar]
- 27.Devreotes P. Science. 1989;245:1054–1058. doi: 10.1126/science.2672337. [DOI] [PubMed] [Google Scholar]
- 28.Walz D, Caplan S R. Proc Natl Acad Sci USA. 1987;84:6152–6156. doi: 10.1073/pnas.84.17.6152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bobo R H, Laske D W, Akbasak A, Morrison P F, Dedrick R L, Oldfield E H. Proc Natl Acad Sci USA. 1994;91:2076–2080. doi: 10.1073/pnas.91.6.2076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Morrison P F, Laske D W, Bobo H, Oldfield E H, Dedrick R L. Am J Physiol. 1994;266:R292–R305. doi: 10.1152/ajpregu.1994.266.1.R292. [DOI] [PubMed] [Google Scholar]
- 31.Zandstra P W, Conneally E, Petzer A L, Piret J M, Eaves C J. Proc Natl Acad Sci USA. 1997;94:4698–4703. doi: 10.1073/pnas.94.9.4698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lyman S D, James L, VandenBos T, deVries P, Brasel K, Gliniak B, Hollingsworth C T, Picha K S, McKenna H J, Splett R R. Cell. 1993;75:1157–1167. doi: 10.1016/0092-8674(93)90325-k. [DOI] [PubMed] [Google Scholar]
- 33.Driever W, Nusslein-Volhard C. Cell. 1988;54:83–93. doi: 10.1016/0092-8674(88)90182-1. [DOI] [PubMed] [Google Scholar]
- 34.Driever W, Nusslein-Volhard C. Cell. 1988;54:95–104. doi: 10.1016/0092-8674(88)90183-3. [DOI] [PubMed] [Google Scholar]
- 35.Wiley H S. Receptors: Topology, Dynamics and Regulation. 5A. Greenwich, CN: JAI; 1992. pp. 113–142. [Google Scholar]
- 36.Lee Y, McIntire L V, Zygourakis K. Biotechnol Bioengineer. 1994;43:622–634. doi: 10.1002/bit.260430712. [DOI] [PubMed] [Google Scholar]
- 37.Mobest D, Strobel E, von Kleist S, Ries S, Dangel M, Mertelsmann R, Henschler R. Blood. 1996;88:534. (abstr.). [PubMed] [Google Scholar]
- 38.Langer R, Vacanti J P. Science. 1993;260:920–926. doi: 10.1126/science.8493529. [DOI] [PubMed] [Google Scholar]
- 39.Hubbell, J. A., Palsson, B. O. & Papoutsakis, E. T., eds. (1994) Biotechnol. Bioengineer. 43. [DOI] [PubMed]
- 40.Bell E. J Cell Biochem. 1991;45:239–241. doi: 10.1002/jcb.240450302. [DOI] [PubMed] [Google Scholar]
- 41.Miller, W. M. & Peshwa, M. V., eds. (1996) Biotechnol. Bioengineer. 50. [DOI] [PubMed]
- 42.Peng C, Palsson B O. Biotechnol Bioengineer. 1996;50:479–492. doi: 10.1002/(SICI)1097-0290(19960605)50:5<479::AID-BIT2>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- 43.Weisz P B. Science. 1973;179:433–440. doi: 10.1126/science.179.4072.433. [DOI] [PubMed] [Google Scholar]