

Abstract
Nonhuman animals reliably select the largest of two or more sets of discrete items, particularly if those items are food items. However, many studies of these numerousness judgments fail to control for confounds between amount of food e.g., mass or volume) and number of food items. Stimulus dimensions other than number of items also may play a role in how animals perceive sets and make choices. Four chimpanzees (Pan troglodytes) completed a variety of tasks that involved comparisons of food items (graham crackers) that varied in terms of their number, size, and orientation. In Experiment 1, chimpanzees chose between two alternative sets of visible cracker pieces. In Experiment 2, the experimenters presented one set of crackers in a vertical orientation (stacked) and the other in a horizontal orientation. In Experiment 3, the experimenters presented all food items one-at-a-time by dropping them into opaque containers. Chimpanzees succeeded overall in choosing the largest amount of food. They did not rely on number or contour length as cues when making these judgments but instead primarily responded to the total amount of food in the sets. However, some errors reflected choices of the set with the smaller total amount of food but the individually largest single food item. Thus, responses were not optimal because of biases that were not related to the total amount of food in the sets.
Keywords: Chimpanzees, Pan troglodytes, Perception, Quantity Judgment, Numerosity, Representation
Many nonhuman animals perceive and respond to the quantitative properties of stimulus sets (e.g., Beran & Beran 2004; Brannon & Terrace 2000; Call 2000; Emmerton et al. 1997; Hauser et al. 2003; Pepperberg 1994; Roberts et al. 2000). In some cases, animals encode the ordinal properties of stimulus sets and make judgments between two or more sets on the basis of the quantity of stimuli in each set (e.g., Beran et al. 2005; Biro & Matsuzawa 2001; Boysen et al. 1993; Brannon et al. 2006; Brannon & Terrace 2000; Olthof et al., 1997; Pepperberg, 2006). In nature, a variety of animals may use this capacity to choose among food patches comprised of different quantities of food items, or to assess the relative quantity of allies and competitors in a potentially aggressive encounter (Kitchen, 2004; Wilson, Hauser, & Wrangham, 2001). Although the primates are well-studied, crows (Smirnova, Lazareva, & Zorina, 2000), dogs (Ward & Smuts, 2007; West & Young, 2002), salamanders (Uller, Jaeger, Guidry, & Martin, 2003), lions (McComb, Packer, & Pusey, 1994), ferrets (Davis, 1996), dolphins (Jaakola, Fellner, Erb, Rodriguez, & Guarino, 2005) and fish (Agrillo, Dadda, & Bisazza, 2007) all make quantity judgments.
In the laboratory, animals are tested for these abilities in several different situations. Some tests involve spontaneous judgments of quantity, while others involve trained responses. Typically, animals choose between alternative sets of food items or other naturally relevant stimuli (e.g., Beran 2001, 2004; Call 2000; Hauser et al. 2000; Rumbaugh et al. 1987; Uller et al., 2003). Often, in these scenarios, animals select the greater of two quantities, because they realize naturally that ‘more’ is better than ‘less’ (especially with respect to food items). Conversely, judgments between alternative sets of less naturally relevant stimuli (e.g. plastic blocks or digital arrays) require some degree of training, because animals have no inherent reason to discriminate between such items. However, when investigators train an animal to respond to arbitrary stimuli, they can control the rule that the animals must use to profit from a response (e.g., ‘choose more,’ ‘choose less,’ ‘choose three’; Beran 2007; Brannon & Terrace 2000; Judge et al. 2005).
Depending on the test situation, the quantity judgment of interest may not necessarily be a numerical judgment. Several nonnumeric stimulus dimensions co-vary with number, and these include surface area, mass, volume, density, or contour length (Clearfield & Mix 1999, 2001; Davis & Perusse 1988). For example, if an animal is given a choice between two sets of uniformly sized food items, the overall mass of food in each set will correlate with the number of individual items in each set. In order to investigate the use of number as the sole cue in quantity judgments, researchers use artificial stimulus arrays to control for other influential dimensions (e.g., Beran 2007; Brannon & Terrace 2000; Judge et al. 2005).
However, in scenarios in which stimulus sets are not controlled with regard to these nonnumeric properties, we do not know whether animals use number as a cue. After all, when animals choose between two food sources that are comprised of different quantities of identical items, number and amount will correlate perfectly (e.g., Beran 2001, 2004). In this situation, we do not know whether the animal makes a quantity judgment based on the difference in number, the difference in some estimate of amount, or some combination of numeric and nonnumeric properties. Further, we do not know how animals respond in spontaneous judgment tasks when number and one of these other properties conflict (e.g., in a situation in which one set of items is greater in number and, at the same time, smaller in overall amount than the other set). If the choice is between alternative sets of food items, animals should attend mostly to differential amount, because animals naturally prefer more food compared to less food. However, investigators have not thoroughly studied cues relevant to spontaneous quantity judgments of this type in the laboratory.
Contour length may contribute to judgments of quantity. Contour length is defined as the amount of edge or perimeter that is visible around an item, and this property sometimes plays a prominent role in the responses of children (e.g., Clearfield & Mix 1999, 2001; Feigenson, Carey, & Spelke., 2002). The contour length of a stimulus set is simply the contour length of every item in the set added together. When alternative sets contain different sized items, individual item size may also play a role in the responses of animals. Thus, we must account for these properties when we present nonhuman animals with choice tasks using discrete food items if we want to understand the basis for the responses made by animals.
We provided chimpanzees (Pan troglodytes) with several food-choice scenarios, in which number, contour length, and amount (i.e., mass) varied systematically across trials. We tested how chimpanzees would respond when nonnumeric dimensions were presented in conflict with the numeric dimension. We presented chimpanzees with the choice between two sets of the same food graham cracker pieces) across 3 experiments. In Experiment 1 and Experiment 2 we presented choices between static visible sets of different sized cracker pieces in both horizontal and vertical arrangements. In Experiment 3, we provided the chimpanzees with similar choices between sets of cracker pieces, but we used a sequential presentation method (i.e., one by one addition of items to an opaque container). We hypothesized that the chimpanzees would respond on the basis of amount when amount, number, and contour length were presented in conflict. However, we did not know how strongly each of these stimulus dimensions contour length, number of items, size of items, and the visibility of items) might contribute to chimpanzees’ responding. Our hypothesis was that none of these other dimensions would strongly distract the chimpanzees from maximizing intake (i.e., using amount of food as a guide) given how well chimpanzees usually perform in comparing slight differences in food amount (e.g., Menzel 1961; Menzel & Davenport 1962). However, we expected that the number of food items and the presentation of individual food pieces that were larger than all other items in the two sets might slightly affect chimpanzees’ ability to choose the larger amount of food.
We based our last prediction on data from a previous study in which chimpanzees showed an overall bias to point to sets of food items that had the larger individually-sized items, even when those items were not in the set containing the larger amount of food (Boysen et al., 2001). This suggested that item size produced a strong response bias for chimpanzees. In addition, the first author (Beran) once observed a chimpanzee repeatedly choose to receive a whole banana instead of 3 halves of bananas, suggesting that individual element size might play a role in choice responding. In that case, the chimpanzee failed to maximize its intake because of this bias to choose the whole item. Thus, we wanted to assess whether presenting comparisons between alternative sets, in which items within each set could vary in size, would bias chimpanzees toward selecting the set with the largest individual item. We predicted that item size would contribute to responding.
EXPERIMENT 1: JUDGMENTS OF VISIBLE AMOUNTS
Methods
Participants
Four chimpanzees, Lana (female, 35 years of age), Sherman (male, 32 years of age), Panzee (female, 20 years of age), and Mercury (male, 19 years of age) participated in the experiment. These chimpanzees had extensive testing histories in a variety of cognitive tasks including tests relevant to quantity judgment (e.g., Beran 2001, 2004; Beran & Beran 2004; Beran et al. 2005; Rumbaugh et al. 1987).
Apparatus
We positioned food arrays at opposite ends of a wooden bench (48 cm high, 67 cm wide, and 36 cm deep). An experimenter pushed forward a shelf that was mounted on a drawer slide at the top of the bench. This movement presented both sets of food items to the chimpanzee at the same time.
Design and Procedure
At the start of each trial, we placed an opaque barrier between the chimpanzee and the experimental apparatus. Behind that barrier, an experimenter placed two sets of food items on each of the two sides of the shelf. These sets consisted of graham cracker pieces. Each piece of cracker was one of four sizes: a whole cracker, ½ cracker, ¼ cracker, or 1/8 of a cracker. The experimenter arranged the individual pieces with no specific pattern or arrangement (i.e., as pseudo-randomly as possible), but no two pieces touched each other. Thus, the arrangement produced two sets that clearly consisted of a group of discrete graham cracker pieces.
Next, the barrier was removed. Experimenter 1 closed his eyes and pushed the shelf forward. At this point, the chimpanzee reached through its cage mesh and touched one of the two ends of the shelf. This constituted the response, and it was a well-established behavior for these animals. A second experimenter, out of view of the chimpanzee, announced the selection, and the first experimenter then handed the chimpanzee that set of food items before removing the other set. Thus, the experimenter could not cue the chimpanzee because he did not know what selection was made until it was announced.
The two sets of food items varied on a number of different dimensions. The two sets differed in the total graham cracker amount on every trial, and we considered this property to be the relevant property for determining correct and incorrect responses. However, the two sets of food items also could differ in the number of food items in each set, the size of items in those sets, the total contour length of each set, and whether or not one set had a cracker piece that was the overall largest piece. This created a number of different trial types of interest. We examined performance in choosing the larger total amount of food as a function of each of these properties to determine how number, contour length and item size might influence the chimpanzees’ responses. There were three ways that each of these properties could be related to the total food amount property. In the Equal conditions, the two alternative sets of food items had the same number of cracker pieces, the same overall contour length, or largest pieces of the same size. Thus, in these conditions, a number, contour, or item size cue could not be used to differentiate the two sets. Alternatively, in the Congruent conditions the set of food items with the greatest amount also had the largest number of items, the longest contour, or the largest individual cracker piece. Therefore, chimpanzees could use those cues to obtain the larger amount of food in those conditions. In the Incongruent conditions, the set of food items that had the greatest total amount had the smaller number of items, the shorter contour, or the smaller individual cracker pieces. Thus, if chimpanzees used those cues in the Incongruent conditions, they would not obtain the larger total amount of food.
These conditions were not mutually exclusive trial categories. In other words, a given trial could belong to one category with respect to number of items and a different category with respect to individual item size. We presented alternative sets from these multiple conditions in a completely random order. Each chimpanzee completed approximately 20 trials each day and a total of 100 trials in the experiment.
Results
Overall, each chimpanzee’s performance in choosing the set of food items with the larger amount of food exceeded chance levels of responding as assessed with a binomial sign test. Lana responded correctly on 96% of the trials (P < 0.01). Mercury responded correctly on 91% of the trials (P < 0.01). Panzee responded correctly on 83% of the trials (P < 0.01). Sherman responded correctly on 92% of the trials (P < 0.01). Chimpanzees chose the larger amount of food equally often in the first 10 trials compared to the last 10 trials (90% compared to 92.5%). Therefore, the chimpanzees chose the larger amount of food in most trials, and they did so from the very beginning of the experiment.
When chimpanzees and other animals choose between two alternative sets of items on the basis of an established quantitative rule, performance (i.e., percentage correct) and the ratio between the two sets (the value obtained by dividing the smaller set size by the larger set size) usually correlate highly. This relationship reflects the typical distance and magnitude effects in which larger differences between sets makes discrimination easier, and smaller overall magnitudes also makes discrimination easier (e.g., Brannon & Roitman 2003; Gallistel & Gelman 2000). Figure 1 presents the data from Experiment 1 as a function of the ratio between sets either in terms of the number of pieces in the two sets or in terms of the total amount of food in the two sets. The chimpanzees made psychophysical judgments on the basis of the amount of food, r3 = −.96, and not the number of food items, r11 = .39. This suggests that the number of items in the two sets influenced chimpanzees’ selections minimally.
Figure 1.
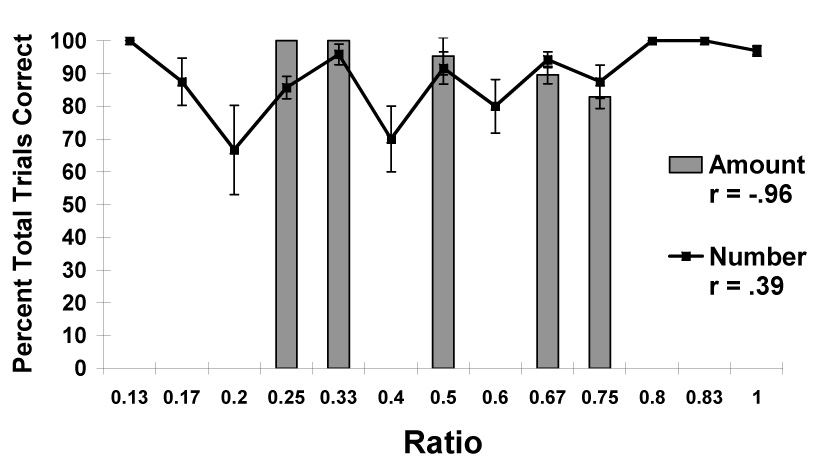
Overall performance in choosing the larger amount in Experiment 1. Data are shown on the basis of the numerical ratio between sets and the ratio between the set with the smaller total amount of food compared to the set with the larger total amount of food. Error bars represent standard errors of means.
Our main purpose was to learn how visual cues other than total amount controlled the chimpanzees’ choices. Figure 2 presents the data from each condition with respect to each visible cue. Here, the frequency data (number of responses) are combined from all chimpanzees. In all conditions, performance in choosing the larger amount was better than chance (P < 0.01, sign test). For each possible cue type, we also conducted a one-way Repeated Measures Analysis of Variance on the chimpanzees’ percentage of correct responses with condition as the factor. For trials in which the number of items differed between sets, performance was not significantly different among the conditions, F (2, 6) = 3.58, P = 0.09. Performance was significantly different across conditions when contour length differed, F (2, 6) = 10.64, P = 0.011, ηp2 = 0.78. However, post hoc paired comparisons indicated that performance in choosing the larger amount was significantly higher in the equal contour condition compared to the other two contour conditions. Performance was significantly different across largest-item conditions (when one alternative set had a larger item than all items in the other set), F (2, 6) = 34.41, P = 0.001, ηp2 = 0.92. Paired comparisons indicated that performance was significantly lower in the incongruent largest-item condition (i.e., when the largest item was in the set with the smaller total amount of food) compared to the other two largest-item conditions (Equal and Congruent).
Figure 2.
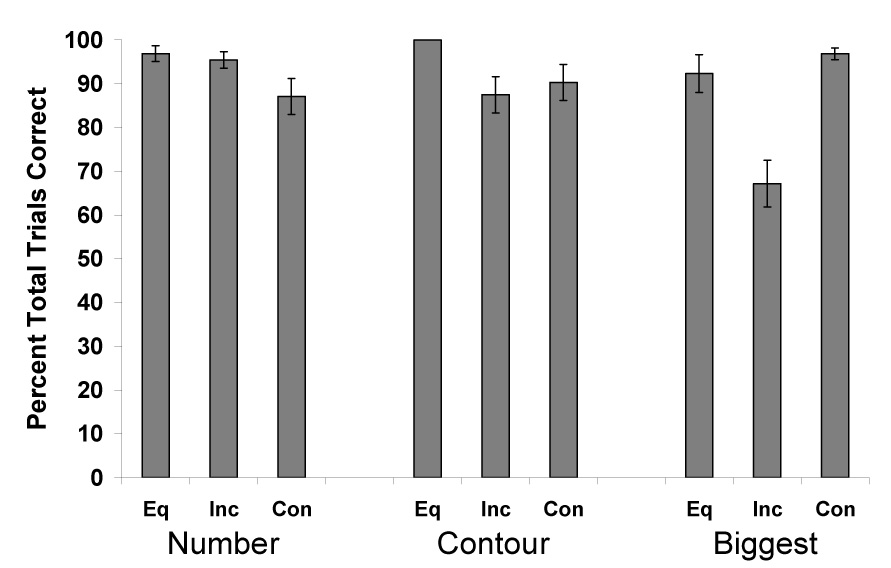
Overall performance in choosing the larger amount for each of the trial types in Experiment 1. Data are separated on the basis of assuming the number, contour, or the location of the biggest individual piece of food acted as the primary cue. Data also are separated on the basis of how well those cues would have operated in helping the chimpanzee choose the larger amount of food. Equal trials (Eq.) were those in which the cue was equated in both set. Congruent trials (Con.) were those in which the cue indicated the set with the larger total amount. Incongruent (Inc.) trials were those in which the cue indicated the set with the smaller total amount of food. Error bars represent standard errors of means.
Discussion
Overall, and as expected, the chimpanzees selected food sets with relatively larger amounts of food in most trials. Thus, the chimpanzees used the amount of food to guide their responses. The chimpanzees were not distracted the number of items in the two sets, and they did not use number as the primary cue in this task. These data provide an important reminder that any interpretation of a judgment task as being a numerical comparison task is premature if amount covaries with number in the procedure. This is especially true when the discrimination is between alternative sets of edible items.
Our observation that chimpanzees use amount, rather than number or contour length, makes adaptive sense. Amount directly affects intake rate, and so choosing the larger amount promotes survival and reproduction. In contrast, the effect of item size would seem to lead to suboptimal decisions. Thus, we were surprised by the large effect that individual item size had on the chimpanzees’ responses.
The chimpanzees performed most poorly when the set with the smaller total amount of food also had the largest individual food piece within it. Chimpanzees’ performance in choosing the larger amount still exceeded chance levels in this condition, but this discrepancy between amount and individual item size did negatively impact the overall amount of food obtained by the chimpanzees. Given the previous research indicating chimpanzees’ very fine discrimination between individual food items (e.g., Menzel 1961; Menzel & Davenport 1962) and sets of food items (e.g., Beran 2001, 2004; Rumbaugh et al. 1987), this bias toward individual items of largest size within an array is intriguing. However, it is not unprecedented (Boysen et al. 2001). Sometimes, sets of larger items have been chosen over alternative sets of small items independent of overall amount of food. However, we expected this bias to decrease when all items varied in size, making the perceptual variation between sets (and item sizes) less evident. In Experiment 2, we investigated whether this bias would also occur when we used a different spatial arrangement from that used in Experiment 1.
EXPERIMENT 2: COMPARISONS BETWEEN VERTICAL AND HORIZONTAL SETS
We again presented two visible sets. However, we arranged one of the two sets differently. All items in one set were identically sized and were stacked so that they presented not only width and depth cues but also a height cue. Here, the chimpanzees had to compare the amount of food in a horizontal, non-overlapping array of items with a vertically stacked array.
Methods
Participants and Apparatus
We tested the same chimpanzees using the same apparatus as in Experiment 1.
Design and Procedure
We made two modifications to the procedure of Experiment 1. First, we randomly assigned the chimpanzees to two observation groups. One group observed as the experimenter arranged the food items in the two alternative sets (Panzee and Sherman) whereas the other group (Lana and Mercury) did not view this arrangement (as in Experiment 1). We varied this presentation form to determine whether chimpanzees benefited from seeing items being stacked into a vertical set. The second change from Experiment 1 was that we arranged one of the two sets differently: we stacked identically sized graham cracker pieces on top of each other (from 2 to 6 pieces). We also gave the chimpanzees some trials in which both sets were horizontal, as in Experiment 1. We presented some trials in which the two sets contained equal amounts of food because these trials provided an important control condition. Whenever we presented one vertical set and one horizontal set, the horizontal set always consisted of either one or two cracker pieces, and the vertical set always had an equal or a larger number of pieces compared to the horizontal set.
We presented five unique conditions on those trials in which the amount of food differed between the alternative sets. In the condition Biggest Item Invalid Cue, we presented a vertical set that contained the larger amount of food, and a horizontal set that possessed the largest individual piece of cracker. In the condition Equal Item Size, we presented a vertical set containing a larger amount of food and items of equal size to the horizontal set. In the condition Number As Valid Cue, we presented two horizontal sets containing different numbers of identically sized items. In the condition Biggest Item Valid Cue – Both Horizontal Sets, we presented two horizontal sets, and one contained the larger total amount of food as well as the largest individual piece of cracker. In the condition Biggest Item Valid Cue – Horizontal and Vertical Sets, we presented a horizontal set that contained a larger amount of food, as well as the largest individual piece of food, in comparison to a vertical set. In the condition Equal Food Amounts, we presented a horizontal set and a vertical set with equal food amounts, but we presented the largest individual piece of food in the horizontal set and the larger number of food items in the vertical set. In this experiment, each chimpanzee completed 56 trials.
Results
Chimpanzees chose the larger total amount of food in a very high percentage of trials (range 83.3%–97.6% correct). Chimpanzees’ performance was equally high in the first 10 trials compared to the last 10 trials (92.5% compared to 95%). Chimpanzees performed similarly regardless of the type of view of the trial set-up, X21 = 3.57, P > 0.05. Thus, it did not matter whether chimpanzees saw the cracker pieces being arranged or did not see the arrangement.
Figure 3 presents the chimpanzees’ performance in the five conditions in which the total amount of food differed between the two sets. Chimpanzees selected the larger amount of food very frequently in all conditions. We again conducted a one-way Repeated Measures Analysis of Variance that indicated no difference in performance on the basis of condition, F (4, 12) < 1.00, P = 0.56, ηp2 = 0.20. For the Equal Food Amounts condition, the chimpanzees selected the horizontal set that contained the largest individual piece on 51.8% of the trials. This did not differ from random levels of responding, P > 0.05, as assessed with a binomial test.
Figure 3.
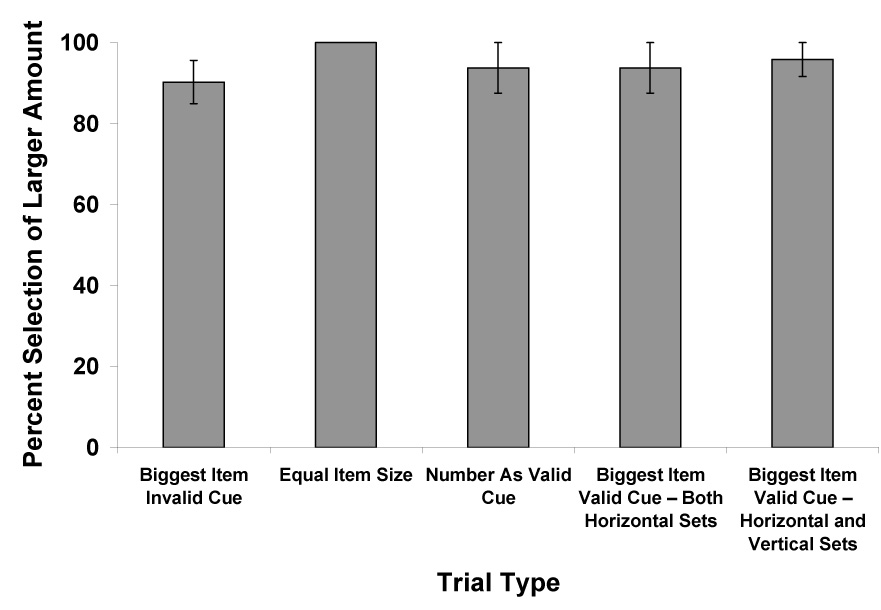
Overall performance in choosing the larger amount for each condition in Experiment 2 for which one set contained more food than the other. See text for descriptions of each trial type. Error bars represent standard errors of means.
Discussion
The chimpanzees again chose the larger amount of food even when we presented one set vertically. This was a more complicated discrimination because the chimpanzees now had to accommodate the potential influence of array height, in addition to previously seen array properties, when judging the relative amount of food in the arrays. They did so easily, however. The chimpanzees also showed no bias toward sets that contained the largest individual item. They performed just as well when we presented the largest item in the alternative set with the larger total amount of food compared to when we presented it in the set with the smaller total amount of food (95.3% and 90.3% correct, respectively). In fact, no matter what the relation between the two sets, the chimpanzees performed very well in choosing the larger food amount (Fig. 3). Thus, the usual effects of ratio between sets or quantitative difference in the number of items were not present because of this high performance. Put simply, this was a very easy task for these chimpanzees.
Perhaps most interesting, the chimpanzees were indifferent in choosing horizontal or vertical sets when each contained an equal amount of food. So, in comparisons between horizontal and vertical arrangements of stimuli, the chimpanzees’ large-item bias from Experiment 1 disappeared. We propose that this occurred because the vertical set presented another potential size cue. The vertical set produced the largest magnitude cue on a third dimension (height) whereas the largest single piece in the horizontal set offered the largest magnitude cue on the length and width dimensions. Thus, the chimpanzees may have abandoned their bias toward sets projecting a particular magnitude cue because both alternative sets now offered such cues. This result suggested that other presentation forms might also disrupt the chimpanzees’ use of this largest-item bias. In Experiment 3 we examined the influence of a sequential presentation method on chimpanzee perception of food amount.
EXPERIMENT 3: COMPARISONS OF SEQUENTIALLY PRESENTED, NON-VISIBLE SETS
We next presented alternative sets by sequentially placing individual food items into opaque containers, similar to the presentation method of previous studies involving identical food items (e.g., Beran 2001, 2004). Thus, the chimpanzees never viewed either set in its entirety. We re-evaluated the relative contributions of the stimulus/set dimensions examined in Experiment 1 in this task in which items were not visible at response time. We know that chimpanzees can make relative judgments of stimuli in situations in which items do not remain in view (e.g., Beran 2001, 2004; Menzel & Draper 1965), but we wanted to determine what effect nonvisibility of arrays at choice time would have on overall performance and on the bias to respond to alternative sets with the largest individual item.
The sequential item presentation method also required the chimpanzees to employ their working memory to form and maintain representations of the food arrays over time (i.e. to sum the sequentially presented items). We expected these manipulations to make the task more difficult, and perhaps alter the likelihood of perceptual biases (such as the largest item bias) contributing to the chimpanzees’ responding.
Methods
Participants and Apparatus
We tested the same chimpanzees using the same apparatus.
Design and Procedure
The experimenter showed food items to the chimpanzee one-at-a-time, and then placed them into an opaque container. Thus, the chimpanzees only saw one food item at a given time, and once it was placed into the opaque container it remained out of view. The experimenter presented one set in its entirety before moving on to the next. After the experimenter presented both sets, the chimpanzees touched one of the two containers to make a selection. We presented the exact same trials as in Experiment 1, but in a new random order. This created the same trial types of interest as in Experiment 1. Each chimpanzee again completed a total of 100 trials.
Results
Overall, each chimpanzee chose the larger amount of food at above-chance levels as assessed with a binomial sign test. Lana responded correctly on 72% of the trials (P < 0.01). Mercury responded correctly on 80% of the trials (P < 0.01). Panzee responded correctly on 74% of the trials (P < 0.01). Sherman responded correctly on 92% of the trials (P < 0.01). All of the chimpanzees chose the larger food amount equally often in the first 10 trials compared to the last 10 trials (75% in both cases).
Figure 4 presents the data as a function of the ratio between alternative sets, either in terms of the number of pieces in the two sets, or in terms of the total amount of food in the two sets. As in Experiment 1, chimpanzees’ selection of sets with larger food amounts correlated highly with the ratio between alternative sets, r3 = −0.91, whereas their selection of sets with larger numbers of food items did not, r11 = 0.48.
Figure 4.
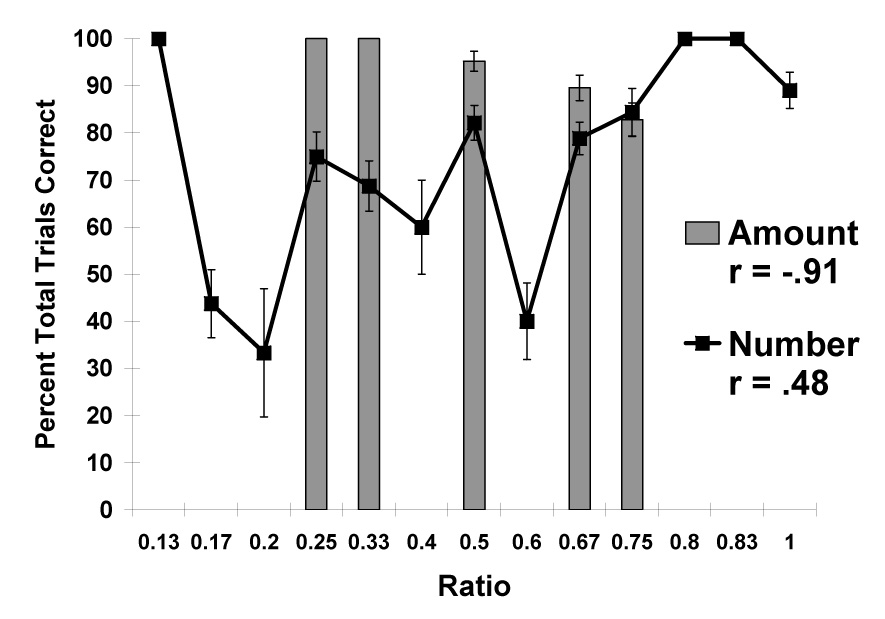
Overall performance in choosing the larger amount in Experiment 3. Data are shown on the basis of the numerical ratio between sets and the ratio between the set with the smaller total amount of food compared to the set with the larger total amount of food. Error bars represent standard errors of means.
Figure 5 presents the data from each of the conditions outlined in Experiment 1. Chimpanzees selected the larger food amount more often than chance (p < .01, sign test) in all conditions except the incongruent condition for the variable of largest individual item. For each possible cue type, we again conducted a one-way Repeated Measures Analysis of Variance on percentage of correct responses with condition as the factor. When the number of items differed between sets, chimpanzees’ choice of the larger amount varied among the conditions, F (2, 6) = 8.32, P = 0.019, ηp2 = 0.72. Paired comparisons indicated that performance was significantly higher in the incongruent number condition compared to the congruent number condition. This means that the chimpanzees performed better when number was not a valid cue. Chimpanzees’ performed similarly across conditions when contour length differed, F (2, 6) = 2.81, P = 0.14. Chimpanzees performed significantly different across largest-item conditions, F (2, 6) = 32.36, P = 0.001, ηp2 = 0.91. Paired comparisons indicated chimpanzees performed significantly lower in the incongruent largest-item condition (when the largest item was in the set with the smaller total amount of food) compared to the other two largest-item conditions.
Figure 5.
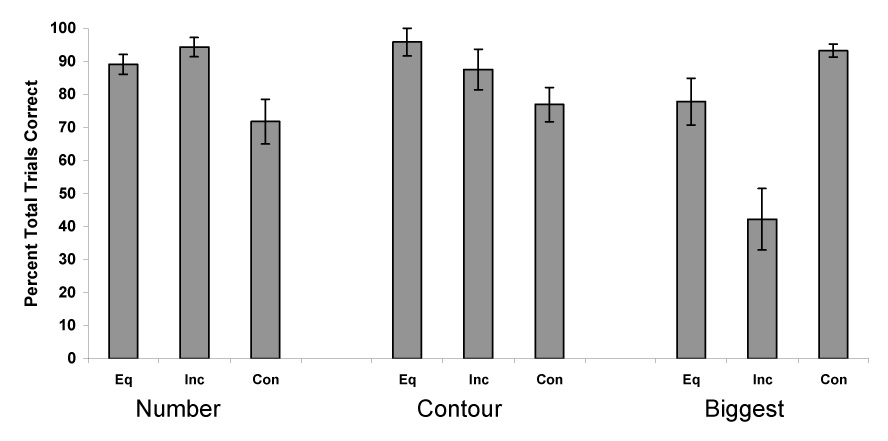
Overall performance in choosing the larger amount for each of the trial types in Experiment 3. Data are shown as in Figure 2. Error bars represent standard errors of means.
Discussion
We expected the sequential presentation method to dampen chimpanzees’ bias to select sets containing the largest individual item. However, it did not. The chimpanzees failed to maximize their intake when the biggest item was in the set with the smaller total amount. They remained handicapped in that situation compared to other trial types, which was similar to the outcome of Experiment 1.
GENERAL DISCUSSION
It seems intuitive that chimpanzees would attempt to maximize the amount of food that they could obtain. The experimental procedure we used is designed for probing many abilities related to quantity judgment including memory, estimation, enumeration, summation, ordinal learning, and perhaps numerosity representation. However, when identical food items make up the alternative sets being compared, it is important to restrict any claims made about animals’ use of the actual number of items (e.g., Anderson et al. 2005; Beran 2001, 2004; Beran & Beran 2004; Call 2000; Rumbaugh et al. 1987). In these so-called relative numerousness tasks, number and amount of food are perfectly correlated, and so we do not know what cue an animal is using. The data from this study suggest that number is not the primary cue when it is presented in conflict with cues regarding total food amount. Across all of these experiments, the chimpanzees were not comparing the number of items in each set. Therefore, number played a secondary role in their judgments (Breukelaar & Dalrymple-Alford 1998; Davis & Memmott 1982). This is not to suggest that numerosity is not a salient aspect of the animals’ environment. Other studies, using non-edible stimuli, show that number can control responding (e.g., Beran 2007; Boysen & Berntson 1989; Brannon & Terrace 2000; Cantlon & Brannon 2006; Judge et al. 2005). However, food judgments most likely do not activate numerical representations because the (presumed) goal of animal is not to maximize the number of obtained food items but rather to maximize the overall mass of obtained food. Like item number, contour length did not influence chimpanzee responding significantly in these experiments. This contrasts with the behavior of human children tested in similar situations (Clearfield & Mix 1999, 2001; Feigenson et al. 2002).
It was somewhat more surprising to see the chimpanzees’ bias to select the set with the biggest individual item. In two of three experiments, the chimpanzees’ chose the food array containing the largest individual item even when that array contained overall less food. This was particularly true when chimpanzees could not view the alternative sets in their entirety (i.e., when items were seen one-at-a-time). Large items contributed psychologically somehow to the representation of the total set food amount beyond what they contributed physically. Boysen et al. (2001) also found that chimpanzees showed a bias to point to the largest items, not the largest amounts of food. This bias is very interesting because it is difficult to understand why any species would prefer bigger individual items at the expense of overall intake. Certainly, if a chimpanzee were only allowed to choose one food item, it would choose the biggest item (e.g., Menzel 1961; Menzel & Davenport 1962). Our chimpanzees have completed thousands of trials over many years in which they choose between alternative sets of food items, so they already learned that they would receive the entire set. In addition, when they must perform summation before selecting a food array, they are not biased by individual set sizes (Beran 2001; Rumbaugh et al. 1987). Thus, something else must be going on in the present situation.
One possibility is that chimpanzees’ responses are biased toward the largest item because those items produce the highest consummatory impulses. However, we know little about the order in which the chimpanzees consume multiple items of the same type but differing sizes. In an informal experiment, we gave the chimpanzees bowls of identical food types of differing sizes and measured the order in which they ate the items. Each chimpanzee completed three trials with four different sized pieces of banana and four trials with four different sized pieces of graham cracker. Table 1 presents the data. Chimpanzees exhibited no tendency to eat the largest item first from these arrays. In fact, the only statistically significant correlations obtained (in 3 of 8 cases) showed that chimpanzees consumed larger food items later within a trial. Overall, the chimpanzees ate the biggest food item last on 64% of these trials. This provides counter-evidence to the idea that chimpanzees chose the alternative set with the largest individual item because they wanted to eat that item first. However, the data do indicate that chimpanzees do notice the relative size of food items as they consume items of different sizes in a non-random order.
Table 1.
Order of consumption of food items of identical type but differing size
| Food Type | Lana | Mercury | Panzee | Sherman |
|---|---|---|---|---|
| Banana (df = 10) | r = .39 | r = 0.0 | r = .33 | r = .60* |
| Graham cracker (df = 12) | r = .15 | r =.60* | r = 0.0 | r = .50* |
Note. Correlations are between selection order and food item size. Positive correlations indicate that larger-sized food items were eaten later within feeding bouts.
p < .05.
Perhaps chimpanzees have difficulty combining multiple stimulus properties to form accurate representations of quantity. Beran et al. (2005) reported that chimpanzees learned to order arbitrary stimuli (plastic eggs) on the basis of the number of food items consistently found in eggs of differing colors. The chimpanzees compared these eggs to visible sets of food items and even changed responses on the basis of the number of eggs in each set. For example, when chimpanzees compared a pink egg (with five food items inside) to seven visible food items, the chimpanzees chose the visible food items. However, when the chimpanzees compared two pink eggs (10 items total) to seven food items, the chimpanzees then chose the egg set. However, chimpanzees had difficulty comparing two sets of eggs (with differing numbers of eggs in each set). The chimpanzees seemed to focus only on the relative values of the colors of the eggs in the two sets, but they did not take into account the relative difference in the numbers of eggs in the two sets (e.g., picking one red egg with five items over two blue eggs with eight total items). Perhaps a similar problem occurs in the present tasks. The chimpanzees may use the relative size of the largest food item as the dominant cue, and they may not take into account that other items in that set are smaller. Perhaps the chimpanzees were not representing sets on the basis of each item’s size but were instead using the largest sized item as a template for all of the items in the set. If this were true, it might help explain the chimpanzees’ poorer performance in the sequential task, in which visual feedback was not available from each of the items in a set. Thus, chimpanzees had more difficulty tracking the contributions of the relatively smaller items to the total amount.
Animals may not evaluate all possible aspects of each problem they face but instead they may rely on a short-cut during decision making if that shortcut typically produces a positive outcome (e.g., Gigerenzer & Goldstein, 1996). Rather than calculating the best response for maximizing overall intake based on all available information, heuristics may be employed that allow for more rapid responding. Gigerenzer and colleagues have suggested one model of heuristic decision making, the Take the Best Algorithm (Gigerenzer & Goldstein, 1996; Todd & Gigerenzer, 2007). This algorithm works by ranking all possible cues and using only the best available cue. If that cue does not indicate a specific response, the next best cue is used. What constitutes the best cue is its ecological validity, or its ability to produce positive outcomes in natural environments (Goldstein & Gigerenzer, 2002). Within foraging situations, this best cue could be the largest piece of food because, at a minimum, that item is most highly valued over other, smaller items. In fact, a bias for the largest piece of food would seldom lead to suboptimal decisions in nature, and may even be the result of natural selection (Hutchinson & Gigerenzer, 2005) because large items equate to more food generally. For chimpanzees, competition for food is also a natural part of travel and foraging bouts, and this might mean that only a limited number of items can be confiscated by a given animal. Here, again, choosing the biggest item in such a situation would be an optimal strategy in almost all situations.
To summarize, chimpanzees do not rely on any one property of food items when choosing between alternative sets. Multiple properties play a role in decision-making. As expected, the total amount of food in the sets plays a dominant role. Number of items plays a more subordinate role. Individual items within heterogeneous sets of stimuli also appear to command attention and bias responding when they are larger than all other elements. Thus, perception of individual food items and perception of the overall quantity in discrete alternative sets of items by chimpanzees is complex, and the responses of these animals are not perfectly efficient in maximizing food intake.
ACKNOWLEDGMENTS
Michael J. Beran, Language Research Center, Georgia State University; Theodore A. Evans, Language Research Center, Georgia State University; Emily H. Harris, Department of Psychology and Language Research Center, Georgia State University.
This research project was supported by Grant HD-38051 from the National Institute of Child Health and Human Development and by Grant BCS-0634662 from the National Science Foundation.
The authors thank John Kelley and Sarah Hunsberger for their care of chimpanzees and their assistance with data collection.
Correspondence concerning this article should be addressed to Michael J. Beran, Language Research Center, Georgia State University, University Plaza, Atlanta, GA 30302. E-mail may be sent to mjberan@yahoo.com
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Lay Summary Animals try to obtain the most food that they can, but we do not know what physical properties of food items attract more or less attention during these choices. We examined how chimpanzees used different properties of food items to make choices of one set over another. The chimpanzees compared two sets of graham crackers pieces, and the sets differed in the number of pieces, the size of the pieces, and the arrangement of the pieces. The chimpanzees did not rely on the number of crackers or the contour length, but they did rely heavily on the total amount of food in the sets. The chimpanzees also were biased to pick sets that had the largest individual food item within them, even when this meant that the chosen set had the smaller total amount of food. Thus, responses were not always optimal in terms of total food obtained.
REFERENCES
- Agrillo C, Dadda M, Bisazza A. Quantity discrimination in female mosquitofish. Animal Cognition. 2007;10:63–70. doi: 10.1007/s10071-006-0036-5. [DOI] [PubMed] [Google Scholar]
- Anderson US, Stoinski TS, Bloomsmith MA, Marr MJ, Smith AD, Maple TS. Relative numerousness judgment and summation in young and old Western Lowland gorillas. Journal of Comparative Psychology. 2005;119:285–295. doi: 10.1037/0735-7036.119.3.285. [DOI] [PubMed] [Google Scholar]
- Beran MJ. Summation and numerousness judgments of sequentially presented sets of items by chimpanzees (Pan troglodytes) Journal of Comparative Psychology. 2001;115:181–191. doi: 10.1037/0735-7036.115.2.181. [DOI] [PubMed] [Google Scholar]
- Beran MJ. Chimpanzees (Pan troglodytes) respond to nonvisible sets after one-by-one addition and removal of items. Journal of Comparative Psychology. 2004;118:25–36. doi: 10.1037/0735-7036.118.1.25. [DOI] [PubMed] [Google Scholar]
- Beran MJ. Rhesus monkeys (Macaca mulatta) enumerate sequentially presented sets of items using analog numerical representations. Journal of Experimental Psychology: Animal Behavior Processes. 2007;33:42–54. doi: 10.1037/0097-7403.33.1.42. [DOI] [PubMed] [Google Scholar]
- Beran MJ, Beran MM. Chimpanzees remember the results of one-by-one addition of food items to sets over extended time periods. Psychological Science. 2004;15:94–99. doi: 10.1111/j.0963-7214.2004.01502004.x. [DOI] [PubMed] [Google Scholar]
- Beran MJ, Beran MM, Harris EH, Washburn DA. Ordinal judgments and summation of nonvisible sets of food items by two chimpanzees (Pan troglodytes) and a rhesus macaque (Macaca mulatta) Journal of Experimental Psychology: Animal Behavior Processes. 2005;31:351–362. doi: 10.1037/0097-7403.31.3.351. [DOI] [PubMed] [Google Scholar]
- Biro D, Matsuzawa T. Use of numerical symbols by the chimpanzee (Pan troglodytes): Cardinals, ordinals, and the introduction of zero. Animal Cognition. 2001;4:193–199. doi: 10.1007/s100710100086. [DOI] [PubMed] [Google Scholar]
- Boysen ST, Berntson GG. Numerical competence in a chimpanzee (Pan troglodytes) Journal of Comparative Psychology. 1989;103:23–31. doi: 10.1037/0735-7036.103.1.23. [DOI] [PubMed] [Google Scholar]
- Boysen ST, Berntson GG, Mukobi KL. Size matters: Impact of item size and quantity on array choice by chimpanzees (Pan troglodytes) Journal of Comparative Psychology. 2001;115:106–110. doi: 10.1037/0735-7036.115.1.106. [DOI] [PubMed] [Google Scholar]
- Boysen ST, Berntson GG, Shreyer TA, Quigley KS. Processing of ordinality and transitivity by chimpanzees (Pan troglodytes) Journal of Comparative Psychology. 1993;107:206–215. doi: 10.1037/0735-7036.107.2.208. [DOI] [PubMed] [Google Scholar]
- Brannon EM, Roitman JD. Nonverbal representations of time and number in animals and human infants. In: Meck WH, editor. Functional and neural mechanisms of interval timing. Boca Raton, FL: CRC Press; 2003. pp. 143–182. [Google Scholar]
- Brannon EM, Terrace HS. Representation of the numerosities 1–9 by rhesus macaques (Macaca mulatta) Journal of Experimental Psychology: Animal Behavior Processes. 2000;26:31–49. doi: 10.1037//0097-7403.26.1.31. [DOI] [PubMed] [Google Scholar]
- Brannon EM, Cantlon JF, Terrace HS. The role of reference points in ordinal numerical comparisons by rhesus macaques (Macaca mulatta) Journal of Experimental Psychology: Animal Behavior Processes. 2006;32:120–134. doi: 10.1037/0097-7403.32.2.120. [DOI] [PubMed] [Google Scholar]
- Breukelaar JWC, Dalrymple-Alford JC. Timing ability and numerical competence in rats. Journal of Experimental Psychology: Animal Behavior Processes. 1998;24:84–97. doi: 10.1037//0097-7403.24.1.84. [DOI] [PubMed] [Google Scholar]
- Call J. Estimating and operating on discrete quantities in orangutans (Pongo pygmaeus) Journal of Comparative Psychology. 2000;114:136–147. doi: 10.1037/0735-7036.114.2.136. [DOI] [PubMed] [Google Scholar]
- Cantlon JF, Brannon EM. Shared system for ordering small and large numbers in monkeys and humans. Psychological Science. 2006;17:401–406. doi: 10.1111/j.1467-9280.2006.01719.x. [DOI] [PubMed] [Google Scholar]
- Clearfield MW, Mix KS. Number versus contour length in infants' discrimination of small visual sets. Psychological Science. 1999;10:408–411. [Google Scholar]
- Clearfield MW, Mix KS. Amount versus number: Infants' use of area and contour length to discriminate small sets. Journal of Cognition and Development. 2001;2:243–260. [Google Scholar]
- Davis H. Numerical competence in ferrets (Mustela putorius furo) International Journal of Comparative Psychology. 1996;9:51–64. [Google Scholar]
- Davis H, Memmott J. Counting behavior in animals: A critical evaluation. Psychological Bulletin. 1982;92:547–571. [Google Scholar]
- Davis H, Perusse R. Numerical competence in animals: Definitional issues, current evidence, and new research agenda. Behavioral and Brain Sciences. 1988;11:561–615. [Google Scholar]
- Emmerton J, Lohmann A, Niemann J. Pigeons' serial ordering of numerosity with visual arrays. Animal Learning and Behavior. 1997;25:234–244. [Google Scholar]
- Feigenson L, Carey S, Spelke E. Infants' discrimination of number vs. continuous extent. Cognitive Psychology. 2002;44:33–66. doi: 10.1006/cogp.2001.0760. [DOI] [PubMed] [Google Scholar]
- Gallistel CR, Gelman R. Non-verbal numerical cognition: From reals to integers. Trends in Cognitive Sciences. 2000;4:59–65. doi: 10.1016/s1364-6613(99)01424-2. [DOI] [PubMed] [Google Scholar]
- Gigerenzer G, Goldstein DG. Reasoning the fast and frugal way: Models of bounded rationality. Psychological Review. 1996;103:650–669. doi: 10.1037/0033-295x.103.4.650. [DOI] [PubMed] [Google Scholar]
- Goldstein DG, Gigerenzer G. Models of ecological rationality: The recognition heuristic. Psychological Review. 2002;109:75–90. doi: 10.1037/0033-295x.109.1.75. [DOI] [PubMed] [Google Scholar]
- Hauser MD, Carey S, Hauser LB. Spontaneous number representation in semi-free-ranging rhesus monkeys. Proceedings of the Royal Society of London B. 2000;267:829–833. doi: 10.1098/rspb.2000.1078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauser MD, Tsao F, Garcia P, Spelke ES. Evolutionary foundations of number: Spontaneous representation of numerical magnitudes by cotton-top tamarins. Proceedings of the Royal Society of London B. 2003;270:1441–1446. doi: 10.1098/rspb.2003.2414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchinson JMC, Gigerenzer G. Simple heuristics and rules of thumb: Where psychologists and behavioural biologists might meet. Behavioural Processes. 2005;69:97–124. doi: 10.1016/j.beproc.2005.02.019. [DOI] [PubMed] [Google Scholar]
- Jaakkola K, Fellner W, Erb L, Rodriguez M, Guarino E. Understanding of the concept of numerically "less" by bottlenose dolphins (Tursiops truncatus) Journal of Comparative Psychology. 2005;119:286–303. doi: 10.1037/0735-7036.119.3.296. [DOI] [PubMed] [Google Scholar]
- Judge PG, Evans TA, Vyas DK. Ordinal representation of numeric quantities by brown capuchin monkeys (Cebus apella) Journal of Experimental Psychology: Animal Behavior Processes. 2005;31:79–94. doi: 10.1037/0097-7403.31.1.79. [DOI] [PubMed] [Google Scholar]
- Kitchen DM. Alpha male black howler monkeys responses to loud calls: Effect of numeric odds, male companion behaviour and reproductive investment. Animal Behaviour. 2004;67:125–139. [Google Scholar]
- McComb K, Parker C, Pusey A. Roaring and numerical assessment in contests between groups of female lions, Pantera leo. Animal Behaviour. 1994;47:379–387. [Google Scholar]
- Menzel EW. Perception of food size in the chimpanzee. Journal of Comparative and Physiological Psychology. 1961;54:588–591. [Google Scholar]
- Menzel EW, Davenport RK. The effects of stimulus presentation variables upon chimpanzee's selection of food by size. Journal of Comparative and Physiological Psychology. 1962;55:235–239. [Google Scholar]
- Menzel EW, Draper WA. Primate selection of food by size: Visible versus invisible rewards. Journal of Comparative and Physiological Psychology. 1965;59:231–239. doi: 10.1037/h0021833. [DOI] [PubMed] [Google Scholar]
- Olthof A, Iden CM, Roberts WA. Judgments of ordinality and summation of number symbols by squirrel monkeys (Saimiri sciureus) Journal of Experimental Psychology: Animal Behavior Processes. 1997;23:325–339. doi: 10.1037//0097-7403.23.3.325. [DOI] [PubMed] [Google Scholar]
- Pepperberg IM. Numerical competence in an African Grey parrot (Psittacus erithacus) Journal of Comparative Psychology. 1994;108:36–44. [Google Scholar]
- Pepperberg IM. Ordinality and inferential abilities of a Grey Parrot (Psittacus erithacus) Journal of Comparative Psychology. 2006;120:205–216. doi: 10.1037/0735-7036.120.3.205. [DOI] [PubMed] [Google Scholar]
- Roberts WA, Coughlin R, Roberts S. Pigeons flexibly time or count on cue. Psychological Science. 2000;11:218–222. doi: 10.1111/1467-9280.00244. [DOI] [PubMed] [Google Scholar]
- Rumbaugh DM, Savage-Rumbaugh ES, Hegel MT. Summation in the chimpanzee (Pan troglodytes) Journal of Experimental Psychology: Animal Behavior Processes. 1987;13:107–115. [PubMed] [Google Scholar]
- Smirnova AA, Lazareva OF, Zorina ZA. Use of number by crows: Investigation by matching and oddity learning. Journal of the Experimental Analysis of Behavior. 2000;73:163–176. doi: 10.1901/jeab.2000.73-163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd PM, Gigerenzer G. Environments that make us smart. Current Directions in Psychological Science. 2007;16:167–171. [Google Scholar]
- Uller C, Jaeger R, Guidry G, Martin C. Salamanders (Plethodon cinereus) go for more: Rudiments of number in an amphibian. Animal Cognition. 2003;6:105–112. doi: 10.1007/s10071-003-0167-x. [DOI] [PubMed] [Google Scholar]
- Ward C, Smuts BB. Quantity-based judgments in the domestic dog (Canis lupus familiaris) Animal Cognition. 2007;10:71–80. doi: 10.1007/s10071-006-0042-7. [DOI] [PubMed] [Google Scholar]
- West RE, Young RJ. Do domestic dogs show any evidence of being able to count? Animal Cognition. 2002;5:183–186. doi: 10.1007/s10071-002-0140-0. [DOI] [PubMed] [Google Scholar]
- Wilson ML, Hauser MD, Wrangham RW. Does participation in intergroup conflict depend on numerical assessment, range location, or rank for wild chimpanzees? Animal Behaviour. 2001;61:1203–1216. [Google Scholar]
- Xu F. Numerosity discrimination in infants: Evidence for two systems of representations. Cognition. 2003;89:B15–B25. doi: 10.1016/s0010-0277(03)00050-7. [DOI] [PubMed] [Google Scholar]