

Abstract
Magnetic resonance imaging (MRI) and metabolic nuclear magnetic resonance (NMR) spectroscopy are clinically available but have had little application in the quantification of experimental lung injury. There is a growing and unfulfilled need for predictive animal models that can improve our understanding of disease pathogenesis and therapeutic intervention. Integration of MRI and NMR could extend the application of experimental data into the clinical setting. This study investigated the ability of MRI and metabolic NMR to detect and quantify inflammation-mediated lung injury. Pulmonary inflammation was induced in male B6C3F1 mice by intratracheal administration of IL-1β and TNF-α under isoflurane anesthesia. Mice underwent MRI at 2, 4, 6, and 24 h after dosing. At 6 and 24 h lungs were harvested for metabolic NMR analysis. Data acquired from IL-1β+TNF-α-treated animals were compared with saline-treated control mice. The hyperintense-to-total lung volume (HTLV) ratio derived from MRI was higher in IL-1β+TNF-α-treated mice compared with control at 2, 4, and 6 h but returned to control levels by 24 h. The ability of MRI to detect pulmonary inflammation was confirmed by the association between HTLV ratio and histological and pathological end points. Principal component analysis of NMR-detectable metabolites also showed a temporal pattern for which energy metabolism-based biomarkers were identified. These data demonstrate that both MRI and metabolic NMR have utility in the detection and quantification of inflammation-mediated lung injury. Integration of these clinically available techniques into experimental models of lung injury could improve the translation of basic science knowledge and information to the clinic.
Keywords: pulmonary inflammation, small-animal magnetic resonance imaging, metabolomics
lung injury comprises a broad spectrum of disease from mild interstitial edema without cellular injury to massive fatal destruction of the lung (29). Pulmonary inflammation participates in the pathogenesis of a number of lung diseases that are of significant consequence to human health such as chronic obstructive pulmonary disease (COPD), lung cancer, acute lung injury/acute respiratory distress syndrome (ALI/ARDS), and asthma. Although the biochemical and cellular mechanisms of these diseases are distinct, neutrophils participate in some aspect of the pathogenesis of all of them and pulmonary inflammation is a hallmark regardless of its origin, intensity, or manifestation (7, 10, 16, 24, 31, 43).
Our understanding of the pathogenesis of disease is greatly enhanced by robust and validated experimental models. Importantly, the translation of basic biomedical knowledge to the prevention and treatment of human diseases most often requires the use of animals. However, the utility of animals to model human disease has inherent limitations, some of which cannot be overcome. The integration of new technology and the use of genetic manipulation can enhance the validity of experimental animal models. This can result in new, innovative designs that assist in advancing our knowledge of disease pathogenesis and improving our ability to test the efficacy of new drug therapies. To this end, there is a growing need to develop, improve, and broaden the utility of animal models for the study of inflammatory pulmonary diseases. Furthermore, given the homology between the human and mouse genomes, the fusion of state-of-the-art technology with mouse models could improve the application of experimental data to the clinical setting (14). Magnetic resonance imaging (MRI) and nuclear magnetic resonance (NMR)-based metabolomics are two examples of contemporary technologies that could have significant utility in the development of and improvement in animal models of pulmonary inflammation based on phenotypic and biochemical end points, respectively.
MRI is noninvasive and does not rely on ionizing radiation like computed tomography (CT). In fact, the radiation deposition of CT is quite high (especially in animal microimaging) and may be a limiting factor in the use of this approach for longitudinal studies. Alternatively, MRI, using prolonged scan times, offers submillimeter spatial resolution, giving it an advantage over other imaging methodologies such as positron emission tomography and X-rays (12, 30, 35). It has been demonstrated that, despite various challenges in lung MRI (such as low tissue density and low water content, variations in magnetic susceptibility, and respiratory and cardiac motions of the lung), fast gradient-echo or, alternatively, spin-echo, respiratory-gated MRI can detect submillimeter lung lesions in mice treated with benzo(a)pyrene (2, 12). Most importantly, use of MRI permits the longitudinal acquisition of anatomic and physiological information from the same subject, thereby minimizing the number of animals needed to generate meaningful data. To date, MRI has been used to assess edema and inflammation in the lungs following allergen or endotoxin challenges or hyperoxia, but additional data are needed to more completely assess the ability of MRI to longitudinally detect and quantify pulmonary inflammation and acute injury (3, 5, 45, 48). One approach to accomplish this objective is to determine whether MRI data correlate to established pathological markers of lung injury (29).
The collection of global metabolic data and their interpretation (both spectral and biochemical) with modern spectroscopic techniques and appropriate statistical approaches is called “metabolic profiling” or “metabolomics” (15, 27, 39). 1H-NMR can quantify multiple metabolites from a single sample and can be used for liquid or solid samples with minimal sample preparation (39). Additional information on energy-rich metabolites and membrane composition can be derived from 31P-NMR. NMR-based metabolomics has significant potential as a tool in translational research since it can provide comprehensive information on the degree of organ dysfunction and putative mechanisms of injury and it has applicability in both experimental and clinical science. In addition, metabolic biomarkers discovered in vitro by NMR on tissue specimens can be translated into in vivo magnetic resonance spectroscopy protocols, which represents another strength of this translational technology. Since metabolic derangement is associated with cellular injury and death, NMR-based metabolomics may be a very sensitive and useful tool for the quantification of pulmonary injury. However, to date NMR-based metabolomics has only had limited evaluation in the quantification of metabolic dysfunction associated with pulmonary injury (40).
To test the utility of MRI and NMR-based metabolomics, we hypothesized that they could be used to detect and quantify pulmonary injury in mice following intratracheal (IT) administration of inflammatory cytokines.
MATERIALS AND METHODS
Mice.
The animal protocol was approved by the University of Colorado Health Sciences Center Animal Care and Use Committee and the University of Michigan University Committee on Use and Care of Animals, and the research adhered to the “Principles of Laboratory Animal Care.” Male mice (8–9 wk old; B6C3F1) were purchased from Taconic Farms (Hudson, NY). Pulmonary injury was induced by the simultaneous IT administration of recombinant mouse interleukin (IL)-1β (50 ng; eBioscience, San Diego, CA) and recombinant mouse tumor necrosis factor (TNF)-α (50 ng; eBioscience); an equivalent volume of sterile saline was given to control animals (6, 7, 9, 13). At the time of dosing, IL-1β and TNF-α were drawn into a sterile syringe (1 ml) to which a gavage needle (22-gauge, Hallowell EMC, Pittsfield, MA) was attached. The syringe was prepared so that ∼100 μl of air followed the dose. For IT administration, each mouse was anesthetized under isoflurane (IsoFlo, isoflurane, USP; Abbott Animal Health, North Chicago, IL), after which the vocal cords were visualized with a rodent intubation table (Hallowell EMC) and a pediatric otoscope (Welch-Allyn Medical Products, Skaneateles Falls, NY). The tip of the gavage needle was gently moved past the vocal cords and into the trachea; the dose of either IL-1β +TNF-α or sterile saline was delivered by depression of the syringe's plunger. After IT administration, the mouse was held upright for ∼5 s, after which the mouse was returned to the cage and allowed to recover from anesthesia.
Magnetic resonance imaging.
Mice were scanned by MRI at 2, 4, 6, and 24 h after IT dosing. The animals were weighed and anesthetized by intraperitoneal injection of a cocktail of ketamine (80 mg/kg) and xylazine (10 mg/kg) and placed on a heating pad until the anesthesia was complete. Mice were positioned in a custom Plexiglas animal bed and inserted into a Bruker volume coil (31-mm diameter), tuned to the 1H frequency of 200 MHz, which was used for radio frequency transmission and receiving. The fast spin-echo rapid acquisition with relaxation enhancement (RARE) proton density (PD)-MRI scans were performed with a Bruker PharmaScan animal scanner (Bruker Medical, Billerica, MA) at 4.7 T equipped with ParaVision software (version 3.2). PD parameters were field of view (FOV) 4.0 cm, slice thickness 1.0 mm no gap, with 16 slices, matrix size 256 × 256, repetition time (TR) = 2,000 ms, echo time (TE) = 31.9 ms, number of averages 4, with a saturation slice around the cardiac area that was applied perpendicular to the imaging projection (total scan time 4 min 16 s). All images were acquired on anesthetized living mice. Respiratory activity (visually), body temperature (by touch), and the level of anesthesia (by toe pinch) were assessed immediately before and after image acquisition.
All images were processed with Bruker ParaVision software. For PD-weighted MRI, the total lung and hyperintense volumes were determined by multiplying the pixel volume by the number of pixels within the inflamed area by hand-drawing the region of interest with “track” (a freehand drawing) command from each set of images. The radiology scientist was blinded to the animal group assignment. Four sequential images that represented the mid-lower thorax region were selected from each animal. From each image, a hyperintense-to-total lung volume (HTLV) ratio was determined. The mean (±SE) HTLV ratio was calculated from all HTLV ratio values from each respective group of mice.
Lung lavage and tissue harvesting.
Immediately after the 6- and 24-h scans, animals were anesthetized and bronchoalveolar lavage fluid (BALF) was acquired from half of the animals after tracheostomy and intubation with a 20-gauge luer stub adapter (Becton Dickinson, Sparks, MD). The lungs were gently lavaged twice with sterile PBS (400 μl). A BALF cell count was performed with a hemacytometer, and a cell differential was performed by fluorescence-activated cell sorting (FACS). The BALF was assayed for albumin and protein as indicators of lung injury (29). After thoracotomy, these lungs were perfused blood free with PBS via catheterization of the right ventricle, while the lungs of animals that were not subjected to lavage were perfused with buffered formalin (10%). Buffer-perfused lungs were removed immediately and snap-frozen in liquid nitrogen for NMR-based metabolomics analysis. After formalin perfusion of the lungs, buffered formalin was instilled into the lungs via the tracheal cannula at 20 cmH2O. After 10 min, the lungs were harvested. Lung samples were paraffin embedded, sectioned, and stained (hematoxylin-eosin) by the University of Colorado histology core laboratory.
Lung histology.
Sections (5 μm) from paraffin-embedded lung samples were stained with hematoxylin and eosin for histological evaluation. An assessment of pulmonary injury was made by a veterinary pathologist who was unaware of the treatment assignments (1).
Differentiation of BALF cells by FACS.
To determine the percentage of leukocytes (neutrophils, macrophages, lymphocytes) in each BALF sample, cells were pelleted by centrifugation (10,000 g, 4°C, 5 min) and resuspended in goat serum (2%) in a Krebs-Ringer-phosphate-dextrose buffer for 1 h. Cells were pelleted again and resuspended in 100 μl of staining buffer (1% BSA in PBS) to which the following stains were added: F480 PE (Cal Tag; Burlingame, CA), rat anti-mouse neutrophil-FITC (Cal Tag), anti-CD45 PE-Cy7 (BD Bioscience, San Jose, CA), anti-CD3 APC (BD Bioscience), anti-CD45/B220 APC (BD Bioscience), and Live/Dead Fixable Red Dead Cell stain (Invitrogen, Carlsbad, CA). Cells were stained for 20 min at room temperature. After staining, cells were washed in staining buffer and fluorescence was detected by a Cytomics FC500 (Beckman Coulter, Fullerton, CA) at the University of Colorado Cancer Center Flow Cytometry Core laboratory.
Determination of BALF albumin and protein concentrations.
Protein levels were determined with a micro-bicinchoninic acid (BCA) protein assay (Pierce, Rockford, IL) (41). This assay uses an extended incubation time that results in a very sensitive colorimetric protein assay. The presence of BALF albumin was measured with a modified albumin assay kit (Bethyl Laboratories, Montgomery, TX). Briefly, Immulon II plates were coated with 1 μg of goat anti-mouse albumin and blocked with 1% BSA. Clarified BALF supernatant was diluted (1:1,000, 1:10,000, and 1:20,000), and 100 μl was incubated at room temperature for 1 h. Purified mouse albumin (MP Biomedicals, Solon, OH) was used to create the standard curve (0–250 ng/ml). Bound albumin was detected with a goat anti-mouse albumin-horseradish peroxidase (HRP) conjugate and 3,3′,5,5′-tetramethylbenzidine (TMB) substrate (Pierce). The absorbance data were analyzed with Softmax PRO 4.1 on a ThermoMax microplate reader (Molecular Devices, Sunnyvale, CA).
Lung acid extraction for NMR-based metabolomics.
Lung tissues were extracted with 8% perchloric acid (Sigma-Aldrich, St. Louis, MO) as previously described (37). Briefly, ∼0.12 g of frozen lung was powdered in a mortar in the presence of liquid nitrogen to which 6 ml of ice-cold perchloric acid was added. After centrifugation for 20 min (1,300 g, 4°C), the supernatants were collected and the pellets were redissolved with 2 ml of perchloric acid, vortexed, and centrifuged. Both supernatants, containing the hydrophilic fraction of the extract, were combined, and the mixture was neutralized (pH 7.0) with KOH before centrifuging again to remove potassium perchlorate.
Supernatants with water-soluble metabolites (hydrophilic compounds) were then lyophilized overnight to remove water for NMR experiments. The extracted hydrophilic metabolites were dissolved in 0.45 ml of deuterium oxide (D2O) before 1H-NMR. The pellets from the second centrifugation containing the lipid fraction were dissolved in 4 ml of ice-cold water, adjusted to pH 7.0 with KOH, and then lyophilized overnight to remove water for NMR experiments. The lipids were dissolved in 1 ml of deuterated chloroform-methanol mixture (2:1 vol/vol) before 1H-NMR. All deuterated compounds were purchased from Cambridge Isotope (Andover, MA).
Quantitative NMR metabolomics.
For NMR analysis, the dissolved hydrophilic and lipophilic extracts were transferred into 5-mm NMR glass tubes (Wilmad LabGlass, Buena, NJ). First, the extracts were analyzed by high-resolution 1H-NMR spectroscopy with a 500-MHz high-resolution Bruker DRX system equipped with Bruker TopSpin software (Bruker Biospin, Fremont, CA) (37, 38). An inverse TXI 5-mm probe was used for all experiments. To suppress water residue in the extracts, a standard Bruker water presaturation sequence was used (“zgpr”). An external reference, trimethylsilyl propionic-2,2,3,3-d4 acid (TMSP, 0.5 mmol/l for hydrophilic and 1.2 mmol/l for lipid extracts), was used for metabolite quantification of fully relaxed 1H-NMR spectra and as a 1H chemical shift reference (0 ppm). For metabolite identification in water-soluble and lipid extracts, a two-dimensional 1H,13C-heteronuclear single quantum correlation (HSQC) NMR sequence was used. The 1H-NMR peaks for single metabolites were identified and referred to a metabolite chemical shift library. After Fourier transformation and phase and baseline corrections, each 1H peak was integrated with 1D WINNMR (Bruker Biospin). The absolute concentrations of single metabolites were then referred to the TMSP integral and calculated according to the formula
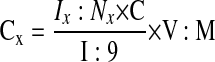 |
where Cx is metabolite concentration, Ix is the integral of the metabolite 1H peak, Nx is the number of protons in the metabolite 1H peak (from CH, CH2, CH3, etc.), C is TMSP concentration, I is the integral of the TMSP 1H peak at 0 ppm (:9 since TMSP has 9 protons), V is volume of the extract, and M is the weight of lung tissue. The final metabolite concentrations were expressed as micromoles per gram of lung tissue.
In addition, the water-soluble (hydrophilic) lung extracts were analyzed by 31P-NMR spectroscopy immediately after 1H-NMR and addition of 100 mmol/l EDTA to chelate divalent ions bound to ATP (28). Phosphorus spectra were obtained on a Bruker 300 MHz Avance spectrometer (31P-NMR frequency: 121.5 MHz) equipped with a 5-mm QNP 31P/13C/19F/1H probe using a composite pulse decoupling (CPD) program. An external standard in a thin capillary, methyldiphosphoric acid (MDP, 2.3 mmol/l D2O, Sigma-Aldrich), was placed into the NMR tube to serve as a reference for both chemical shift (18.6 ppm) and phosphor metabolite quantification (see above).
Statistical analyses.
Treatment and time-dependent changes in HTLV ratios were assessed by a two-way analysis of variance (ANOVA) with StatView (SAS, Cary, NC). Post hoc testing was performed with a Tukey's post hoc test when applicable. BALF cell count/differential and albumin-to-protein ratios from placebo- and IL-1β+TNF-α-treated mice were compared with an unpaired Student's t-test. Absolute individual concentrations of each metabolite obtained from each of the three NMR experiments for each sample (1H-NMR on hydrophilic and hydrophobic extracts as well as 31P-NMR on the hydrophilic extracts) were analyzed by ANOVA followed by Tukey's post hoc test to identify the groups that differed significantly.
Principal component analysis (PCA) was independently applied to the global quantitative data sets (consisting of absolute concentrations of all hydrophilic and lipophilic endogenous metabolites calculated from 1H- and 31P-NMR spectra). The PCA prediction and classification (group clustering or “pattern recognition” visualization) and all mathematical models were built by Bioreactor R package (2.00) on absolute concentrations of endogenous lung metabolites (39). Briefly, after bucket histograms were loaded into the R package software, PCA was applied in order to 1) cluster the samples among placebo- and IL-1β+TNF-α-treated animals (scores ti) and 2) identify markers responsible for this group clustering (plots pi).
RESULTS
MRI of the lungs detected increased water signal (hyperintense edema areas) in mice treated with IT IL-1β+TNF-α that was absent in mice treated with IT sterile saline (placebo) (Fig. 1). This result is substantiated by the difference in HTLV ratio between IL-1β+TNF-α-treated mice and placebo-treated mice and was sustained over a 6-h time course (Fig. 2). By 24 h, the mean (±SE) HTLV ratio of IL-1β+TNF-α-treated mice returned to control values. Although these results alone are not sufficient to demonstrate that the IL-1β+TNF-α-induced pulmonary infiltrates observed on MRI were indicative of lung injury, the histological data and the data acquired from the BALF confirm that this was indeed the case.
Fig. 1.

Representative magnetic resonance imaging (MRI; 4 axial sections, 1 mm thick) of a mouse thorax that 6 h earlier received intratracheal (IT) interleukin (IL)-1β + tumor necrosis factor (TNF)-α (arrows indicate inflamed areas) (A) or received IT placebo (sterile saline) (B). Diffuse infiltrates are apparent in the images of the IL-1β+TNF-α-treated mouse. These changes corresponded to a hyperintense-to-total lung volume (HTLV) ratio of 0.15 compared with the placebo-treated mouse, for which the ratio was 0.04.
Fig. 2.

Time course of HTLV ratio for IL-1β+TNF-α- and placebo-treated mice. MRI was performed at 2, 4, and 6 h in the same mice. At these time points, the mean HTLV volume ratio was greater for IL-1β+TNF-α-treated mice than placebo-treated mice [*P = 0.04 (2 h), P = 0.001 (4 h), P = 0.03 (6 h)]. By 24 h, the mean (±SE) HTLV ratio of IL-1β+TNF-α-treated mice returned to control values (0.05 ± 0.00 and 0.07 ± 0.02, respectively; P = 0.52). Data are means (±SE) for 4 animals/group.
Microscopic examination of the lungs showed significant alveoli damage and edema secondary to the administration of IL-1β+TNF-α that, similar to the HTLV ratio, resolved by 24 h (Fig. 3). The predictiveness of the HTLV ratio was further substantiated by the BALF cell count and differential. The BALF cell differential was determined by FACS. This is a rapid, accurate, and quantitative approach that has been validated previously (8, 47). IT instillation of IL-1β+TNF-α resulted in a profound increase in BALF cell count (Fig. 4A) at 6 h that was attributed to an influx of neutrophils into the airways (Fig. 4B). This IL-1β+TNF-α-induced increase in BALF neutrophils was accompanied by evidence of lung injury, since these animals had a higher mean (±SE) BALF albumin-to-total protein ratio at 6 h compared with saline-treated animals (0.31 ± 0.05 vs. 0.14 ± 0.02, P = 0.012). Both BALF cell count and albumin-to-total protein ratio declined by 24 h but were still greater than control. This suggests that the BALF data lagged behind MRI and histological indicators of lung injury. Nevertheless, collectively, these results show a strong association between MRI-detectable hyperintense areas as a quantitative marker of pulmonary inflammation and the gold standard of measured pathological and histological end points.
Fig. 3.

Representative light micrographs (×20) of hematoxylin and eosin-stained lung sections. A: 6 h after a mouse received IT IL-1β+TNF-α. The micrograph shows evidence of severe lung injury with an acute inflammatory response characterized by mononuclear inflammatory cell infiltrate and innumerable sloughed pneumocytes in the alveoli. The alveolar wall is edematous. Type II pneumocytes lining the alveoli are hypertrophic and hyperplastic. B: 6 h after a mouse received IT sterile saline (placebo). The micrograph shows empty alveolar spaces filled with air. The alveolar walls are lined with flattened epithelial cells. There are no inflammatory cells, no reactive type II pneumocytes, no fibrin, and no evidence of injury. C and D: 24 h after a mouse received either IT IL-1β+TNF-α (C) or IT sterile saline (D). Both micrographs show normal cellular morphology and no evidence of injury.
Fig. 4.

IT instillation of IL-1β+TNF-α resulted in an increase in bronchoalveolar lavage fluid (BALF) cell concentration (A) compared with placebo-treated mice at 6 h (P = 0.011). By 24 h the cell count had declined but was still greater than that in placebo-treated mice (P = 0.002). Data are means (±SE) of 4 animals/group. B and C: representative fluorescence-activated cell sorting (FACS) data showing the distribution (%) of leukocytes in BALF acquired from a single animal 6 h after IL-1β+TNF-α treatment (B) or placebo treatment (C). The mean (±SE) distribution of BALF neutrophils from all lavaged IL-1β+TNF-α-treated mice was 76.8 ± 2.2% compared with 1.6 ± 0.9% for all lavaged placebo-treated mice (P < 0.0001). Macrophages were less prevalent in IL-1β+TNF-α-treated mice than in placebo-treated mice (89.1 ± 2.0% vs. 0.6 ± 0.4%, P < 0.0001). This shift in the differential reflects the acute nature of the inflammation and is further evidenced by the presence of more aggregates, which most likely represent neutrophil-macrophage interactions, in the IL-1β+TNF-α-treated mice.
In addition to these conventional end points, we elected to characterize the relationship between metabolic NMR and MRI data. The lung tissue from control animals and IL-1β+TNF-α-treated mice 6 and 24 h after IT dosing were analyzed by quantitative NMR-based metabolomics. The representative 1H- and 31P-NMR spectra of control lungs are shown in Fig. 5, and 52 endogenous metabolites and/or metabolite ratios from the 1H- and 31P-NMR of the hydrophilic and lipophilic fractions of these lung extracts are depicted in Table 1. These metabolites were included as a quantitative data set for each animal into the PCA. Figure 6A depicts a PCA score of variables 1 and 2 (t1 and t2), which shows a significant group clustering between 6-h IL-1β+TNF-α-treated mice and the control group, while the lungs 24 h after IT IL-1β+TNF-α dosing are between the two groups. Variables 1 and 2 (t1 and t2) are principal components accounting for 75% and 16%, respectively, of the total variance within the quantitative data matrix. By examining the corresponding loading plots (Fig. 6B), which discriminate single metabolic points responsible for group clustering on the PCA scores in Fig. 6A, the high-energy phosphates (ATP, ADP, energy balance, and energy charge) as well as the lactate-to-glucose ratio were identified as candidate biomarkers that differed among the study groups. The PCA demonstrated that a physiological consequence of IL-1β+TNF-α treatment was an ∼50% depletion of ATP at 6 h and a corresponding elevation of the lactate-to-glucose ratio, indicating a shift to anaerobic metabolism during inflammation (Fig. 7). Both of these end points returned to control levels by 24 h. Likewise, the energy balance, which is represented by the ATP-to-ADP ratio, declined in IL-1β+TNF-α-treated mice at 6 h but returned to baseline at 24 h. These data exemplify the reversible nature of the IL-1β+TNF-α-induced lung injury. Collectively, the PCA data indicate that there is a clustering of distinct high-energy metabolites associated with IL-1β+TNF-α administration and that these metabolites are sensitive to temporal changes that mimic the histological, pathological, and MRI data.
Fig. 5.

Representative 1H- and 31P-nuclear magnetic resonance (NMR) spectra of control lung tissue that provide quantitative information on 52 endogenous metabolites and ratios. Peak assignment of major metabolites on 1H- and 31P-NMR spectra: 1, valine, leucine, isoleucine; 2, lactate; 3, alanine; 4, acetate, 5, CH3-acetyl groups; 6–9, glutamate, succinate, glutamine, and total glutathione; 10, creatine; 11, total choline; 12, taurine; 13, glycine; 14, inositol, 15, glucose; 16, ATP; 17, sugar phosphates; 18, NAD+; 19, ADP; 20, phosphodiesters; 21, AMP; 22, phosphomonoesters.
Table 1.
Absolute concentrations of lung endogenous metabolites and their ratios calculated from 1H (water-soluble and lipid fraction)- and 31P-NMR spectra of control lung tissues
| Metabolite | NMR | Concentration, μmol/g lung tissue |
|---|---|---|
| Acetate | 1H-NMR (ws) | 0.11±0.09 |
| CH3-acetyl groups | 1H-NMR (ws) | 0.72±0.08 |
| Adenine | 1H-NMR (ws) | 0.46±0.06 |
| Adenosine | 1H-NMR (ws) | 0.29±0.03 |
| Adenosine diphosphate (ADP) | 31P-NMR | 0.56±0.06 |
| Alanine | 1H-NMR (ws) | 0.12±0.02 |
| Adenosine monophosphate (AMP) | 31P-NMR | 0.04±0.01 |
| Aromatic amino acids | 1H-NMR (ws) | 2.64±2.24 |
| Aspartate | 1H-NMR (ws) | 0.07±0.01 |
| Adenosine triphosphate (ATP) | 31P-NMR | 1.96±0.18 |
| OH-butyrate | 1H-NMR (ws) | 0.04±0.01 |
| Total choline | 1H-NMR (ws) | 0.33±0.10 |
| Total choline (lipids) | 1H-NMR (lipids) | 3.13±0.18 |
| Citrate | 1H-NMR (ws) | 0.09±0.02 |
| Creatine | 1H-NMR (ws) | 0.33±0.08 |
| Cholesterol | 1H-NMR (lipids) | 1.93±0.22 |
| Energy balance | 31P-NMR | 3.54±0.67 |
| Energy charge | 31P-NMR | 0.87±0.02 |
| Total fatty acids | 1H-NMR (lipids) | 27.37±6.28 |
| Glucose | 1H-NMR (ws) | 0.35±0.03 |
| Glutamate | 1H-NMR (ws) | 0.37±0.09 |
| Glutamine | 1H-NMR (ws) | 0.22±0.06 |
| Reduced glutathione | 1H-NMR (ws) | 0.10±0.04 |
| Total glutathione | 1H-NMR (ws) | 0.34±0.13 |
| Glycerol backbone on phospholipids | 1H-NMR (lipids) | 3.91±0.45 |
| Glycine | 1H-NMR (ws) | 0.48±0.04 |
| Inositol | 1H-NMR (ws) | 0.20±0.04 |
| Lactate | 1H-NMR (ws) | 0.55±0.12 |
| [Lactate/glucose] | 1H-NMR (ws) | 1.56±0.27 |
| (CH2)n-lipids | 1H-NMR (lipids) | 285.71±69.71 |
| Total lipids | 1H-NMR (lipids) | 45.03±7.40 |
| Lysine | 1H-NMR (ws) | 0.19±0.09 |
| Monounsaturated fatty acids (MUFA) | 1H-NMR (lipids) | 5.68±0.64 |
| NAD+ | 31P-NMR | 0.15±0.03 |
| Nucleotides | 1H-NMR (ws) | 0.17±0.01 |
| Phosphatidylcholine (PtdCho) | 1H-NMR (lipids) | 2.10±0.01 |
| Phosphatidylethanolamine (PtdEth) | 1H-NMR (lipids) | 0.60±0.04 |
| Phosphatidylinositol (PtdIns) | 1H-NMR (lipids) | 0.80±0.08 |
| Phosphocreatine | 31P-NMR | 0.02±0.01 |
| Phosphodiesters (PDE) | 31P-NMR | 0.24±0.01 |
| Phosphomonoesters (PME) | 31P-NMR | 0.69±0.08 |
| [PME/PDE] | 31P-NMR | 2.86±0.32 |
| Polyols | 1H-NMR (ws) | 0.57±0.06 |
| Polyunsaturated fatty acids (PUFA) | 1H-NMR (lipids) | 13.75±2.98 |
| [PUFA/MUFA] | 1H-NMR (lipids) | 2.40±0.27 |
| Signal X | 1H-NMR (ws) | 0.32±0.08 |
| Sphingomyelin | 1H-NMR (lipids) | 0.28±0.12 |
| Succinate | 1H-NMR (ws) | 0.30±0.07 |
| Sugar phosphates (UDPG) | 31P-NMR | 0.06±0.02 |
| Taurine | 1H-NMR (ws) | 2.32±0.16 |
| Triacylglycerol (TAG) | 1H-NMR (lipids) | 3.91±0.96 |
| Valine, leucine, isoleucine | 1H-NMR (ws) | 0.73±0.07 |
Data are means ± SD. NMR, nuclear magnetic resonance; ws, water soluble. All metabolite ratios are italicized.
Fig. 6.

Principal component analysis (PCA) scores on 52 quantitative metabolic end points of lung extracts. A: PCA scores (ti) based on the global metabolic pattern clusters control for IL-1β+TNF-α-treated and placebo-treated mice. Each triangle represents the quantitative metabolic data set from an individual animal for which 1–4 represent IL-1β+TNF-α-treated mice at 6 h, 5–8 represent IL-1β+TNF-α-treated mice at 24 h, and 9–11 represent placebo-treated mice at 6 h. B: PCA plots (pi) on individual data sets distinguish single putative biomarkers responsible for the clustering pattern observed in A. Each circle represents a specific metabolic end point (from the panel of 52 metabolites included in the PCA).
Fig. 7.

Quantitative comparison of distinguished metabolic biomarkers for IL-1β+TNF-α-treated mice, which shows the reversible nature of the metabolic derangement induced by the cytokines. The energy balance was calculated as ATP-to-ADP ratios and the energy charge as [(ATP + 0.5ADP)/(ATP + ADP + AMP)]. All measured indexes were different from control at 6 h (*P ≤ 0.03), with the exception of ADP. Differences were also evident between the 6 h and 24 h time points (+P < 0.05). By 24 h, ATP levels and the lactate-to-glucose ratio had recovered to control levels (P = 0.11 and P = 0.07, respectively). The amount of ADP increased over the study's time course, most likely because of injury-induced increase in energy production. Data are means ± SE of 3 or 4 animals/group.
DISCUSSION
This study demonstrates that simultaneous IT administration of IL-1β and TNF-α resulted in profound but reversible inflammatory pulmonary injury in free-breathing, normal mice that was detected by MRI and metabolic NMR spectroscopy. Importantly, our data suggest that the MRI-derived HTLV ratio (as a marker of lung edema) and changes in energy metabolism by NMR spectroscopy (as a marker of cell vitality) are potentially viable biomarkers of reversible lung injury since they were both sensitive to temporal changes in lung inflammation as determined by pathological (e.g., BALF albumin-to-protein ratio) and histological end points of lung injury.
To induce pulmonary inflammation, a combination of IL-1β and TNF-α was used. The rationale for this is that these two inflammatory cytokines have pathophysiological relevance in a number of lung diseases such as lung cancer, asthma, COPD, and ALI/ARDS (6, 9, 13, 23, 42). To target delivery of the cytokines to the lungs, we administered them IT. This nonsurgical dose delivery method conserves test material and, as previously demonstrated, results in diffuse delivery to the lower lungs (44). This was evident from the MRI and histological data generated in this study. In addition, the combination of the two, more so than each individually, induced a potent neutrophil-mediated lung injury, a physiological consequence of which was derangement in energy metabolism as detected by NMR-spectroscopy (34, 36).
Imaging technology is growing and developing at a rapid pace, resulting in the introduction of a number of options for small-animal imaging (32, 35). We elected to use MRI because it offers submillimeter spatial resolution (156 μm was the pixel size in this study) and does not require the use of radiation, images are rapidly attainable (<10 min) with no influence of the respiratory cycle, and it is clinically available (12, 30). The clinical applicability of imaging is particularly appealing since it may enhance the translation of experimental data to the human situation. As mentioned previously, the noninvasive nature of MRI allows for time-dependent evaluations of disease progression in the same animal. The advantage of this approach is evident in the present study—while inflammation-related injury was present in the mouse lungs at 2, 4, and 6 h after IL-1β+TNF-α injection, inflammation was reduced at the 24 h time point. The importance of documenting the time course of lung injury has also been previously demonstrated by Beckmann and colleagues (3) in a rat model sensitized and challenged with ovalbumin. An intense, uniform edematous signal was detected in the lungs 24 h after challenge, reached a maximum at 38 h, and resolved after 100 h. The edematous hyperintense areas, present in our MRI scans at 6 h, resolved by 24 h and correlated well with changes in the energy state of the lung tissue by NMR, as well as with the gold standard end points in lung histology. The energy state of the inflamed lung was decreased (as seen by ATP depletion, energy balance, and energy charge levels) with a significantly increased glycolytic activity (elevated lactate-to-glucose ratios). However, the inflammation trigger used in our study was not sufficient enough to induce cell death. Despite ATP depletion of 50%, lung energy homeostasis returned to normal 24 h after IL-1β+TNF-α administration.
A number of studies have been conducted using small-animal imaging to quantify lung injury (3, 11, 21, 45, 49). These studies have generated data from mice and rats with different imaging techniques, but to date only two have utilized MRI to quantify inflammation in mice. Tournebize et al. (46) and Marzola et al. (25) demonstrated the utility of MRI for the detection and quantification of bacteria-induced inflammatory lesions and correlated these data to histological end points. Our study extended these findings by demonstrating the ability of MRI to detect and quantify more diffuse cytokine-induced inflammation and linked these data to metabolic changes as well as histological end points. Collectively, there now appears to be a growing body of evidence that demonstrates the ability of MRI to detect and quantify lung inflammation in mice.
We elected to use mice despite the reported limitations of their use for studying pulmonary physiology (20). However, recent evidence, as well as the importance of genetic manipulation, outweighed these limitations (18, 19, 33). Given the prominence of the mouse model in biomedical research, there is still a paucity of data that pertain to the use of mice as experimental models of pulmonary disease. Despite the genetic advantage of mice, our experiments employed normal B6C3F1 mice because the objective of our work was to determine the ability of MRI and NMR to detect and quantify inflammation-mediated lung injury rather than to distinguish genetic mechanisms of disease. This is a first generation (F1) hybrid strain produced by crossing C57BL/6 females with C3H males. The advantage of an F1 hybrid is genetic and phenotypic uniformity that minimizes animal-to-animal variability. The data generated from this work provide the basis and rationale for planned subsequent work to study more complicated disease-genetic interactions.
The breadth of lung injury spans from mild interstitial edema (reversible damage) to extensive cellular injury (irreversible damage) (29). Specific biomarkers that differentiate the two have not been identified but could represent a powerful experimental and clinical tool to distinguish the spectrum and extent of pulmonary injury. Instillation of IL-1β and TNF-α into the lungs caused significant injury, as evidenced by data acquired from histology and the BALF (17, 22). However, if a single time point had been assessed, then the resolution of inflammation and the reversible nature of the injury would have been missed. This illustrates the relevance and need for longitudinal models that can detect the formation as well as the resolution of injury. In addition, longitudinal study permits the assessment of pathophysiological mechanisms as well as the impact of therapeutic intervention on these processes. Importantly, the use of MRI coupled with metabolic NMR spectroscopy, which showed metabolic derangement (change in energy metabolism) consistent with reversible cellular injury, may have the ability to demarcate the broad range of what constitutes lung injury. In this regard, neither the reversible nature of the injury nor the absence of NMR-detected changes in lipid peroxidation should minimize the consequence of IL-1β+TNF-α-induced injury, since a clinical study demonstrated that a decrease in the ratio of ATP to ADP (energy balance) was associated with increased organ failure and nonsurvival (4). Certainly, the applicability of both MRI and NMR will require more extensive and rigorous testing, but the data presented here introduce the concept that these experimental tools could significantly advance our ability to quantify, characterize, and differentiate the spectrum of experimental pulmonary injury. We do acknowledge that although metabolic NMR spectroscopy is clinically available, the translational aspect of the experimental NMR data presented here is limited by the fact that they were acquired from lung tissue. Pulmonary biopsy is not routinely performed in patients with lung inflammation. In addition, the detection of changes in high-energy phosphates in humans has been done with skeletal muscle (4, 26). Therefore, inevitably, additional work is needed to develop and validate clinical protocols that utilize more readily accessible samples (e.g., blood) before the translational element of this work can be fully realized. Nevertheless, this does not minimize the significance of our findings, because they demonstrate a strong temporal association between MRI-derived HTLV ratio, NMR-based metabolomics, and pathological and histological indexes of lung injury. Thus MRI and metabolic NMR spectroscopy may enhance our ability to develop more robust and predictive longitudinal models of experimental lung injury.
GRANTS
This work was supported, in part, by National Institutes of Health (NIH) Grant R42-HL-071439 (K. A. Stringer). Biomedical MRI/NMR Cancer Center Core was partly supported by the Department of Anesthesiology and the NIH P30-CA-046934 Cancer Center Core Grant (N. J. Serkova). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Heart, Lung, and Blood Institute, the National Cancer Institute, or the National Institutes of Health.
Acknowledgments
We acknowledge and thank Kendra Hasebroock (MRI/CT RT) and Dr. Mark Brown for their assistance during MRI sessions.
The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
REFERENCES
- 1.Ashcroft T, Simpson JM, Timbrell V. Simple method of estimating severity of pulmonary fibrosis on a numerical scale. J Clin Pathol 41: 467–470, 1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Beckmann N, Cannet C, Karmouty-Quintana H, Tigani B, Zurbruegg S, Ble FX, Cremillieux Y, Trifilieff A. Lung MRI for experimental drug research. Eur J Radiol 64: 381–396, 2007. [DOI] [PubMed] [Google Scholar]
- 3.Beckmann N, Tigani B, Ekatodramis D, Borer R, Mazzoni L, Fozard JR. Pulmonary edema induced by allergen challenge in the rat: noninvasive assessment by magnetic resonance imaging. Magn Reson Med 45: 88–95, 2001. [DOI] [PubMed] [Google Scholar]
- 4.Brealey D, Brand M, Hargreaves I, Heales S, Land J, Smolenski R, Davies NA, Cooper CE, Singer M. Association between mitochondrial dysfunction and severity and outcome of septic shock. Lancet 360: 219–223, 2002. [DOI] [PubMed] [Google Scholar]
- 5.Caruthers SD, Paschal CB, Pou NA, Roselli RJ, Harris TR. Regional measurements of pulmonary edema by using magnetic resonance imaging. J Appl Physiol 84: 2143–2153, 1998. [DOI] [PubMed] [Google Scholar]
- 6.Chung KF Cytokines in chronic obstructive pulmonary disease. Eur Respir J Suppl 34: 50s–59s, 2001. [PubMed] [Google Scholar]
- 7.Coussens LM, Werb Z. Inflammation and cancer. Nature 420: 860–867, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dauber JH, Wagner M, Brunsvold S, Paradis IL, Ernst LA, Waggoner A. Flow cytometric analysis of lymphocyte phenotypes in bronchoalveolar lavage fluid: comparison of a two-color technique with a standard immunoperoxidase assay. Am J Respir Cell Mol Biol 7: 531–541, 1992. [DOI] [PubMed] [Google Scholar]
- 9.Engels EA, Wu X, Gu J, Dong Q, Liu J, Spitz MR. Systematic evaluation of genetic variants in the inflammation pathway and risk of lung cancer. Cancer Res 67: 6520–6527, 2007. [DOI] [PubMed] [Google Scholar]
- 10.Foley SC, Hamid Q. Images in allergy and immunology: neutrophils in asthma. J Allergy Clin Immunol 119: 1282–1286, 2007. [DOI] [PubMed] [Google Scholar]
- 11.Ford NL, Martin EL, Lewis JF, Veldhuizen RA, Drangova M, Holdsworth DW. In vivo characterization of lung morphology and function in anesthetized free-breathing mice using micro-computed tomography. J Appl Physiol 102: 2046–2055, 2007. [DOI] [PubMed] [Google Scholar]
- 12.Garbow JR, Zhang Z, You M. Detection of primary lung tumors in rodents by magnetic resonance imaging. Cancer Res 64: 2740–2742, 2004. [DOI] [PubMed] [Google Scholar]
- 13.Goodman RB, Pugin J, Lee JS, Matthay MA. Cytokine-mediated inflammation in acute lung injury. Cytokine Growth Factor Rev 14: 523–535, 2003. [DOI] [PubMed] [Google Scholar]
- 14.Gregory SG, Sekhon M, Schein J, Zhao S, Osoegawa K, Scott CE, Evans RS, Burridge PW, Cox TV, Fox CA, Hutton RD, Mullenger IR, Phillips KJ, Smith J, Stalker J, Threadgold GJ, Birney E, Wylie K, Chinwalla A, Wallis J, Hillier L, Carter J, Gaige T, Jaeger S, Kremitzki C, Layman D, Maas J, McGrane R, Mead K, Walker R, Jones S, Smith M, Asano J, Bosdet I, Chan S, Chittaranjan S, Chiu R, Fjell C, Fuhrmann D, Girn N, Gray C, Guin R, Hsiao L, Krzywinski M, Kutsche R, Lee SS, Mathewson C, McLeavy C, Messervier S, Ness S, Pandoh P, Prabhu AL, Saeedi P, Smailus D, Spence L, Stott J, Taylor S, Terpstra W, Tsai M, Vardy J, Wye N, Yang G, Shatsman S, Ayodeji B, Geer K, Tsegaye G, Shvartsbeyn A, Gebregeorgis E, Krol M, Russell D, Overton L, Malek JA, Holmes M, Heaney M, Shetty J, Feldblyum T, Nierman WC, Catanese JJ, Hubbard T, Waterston RH, Rogers J, de Jong PJ, Fraser CM, Marra M, McPherson JD, Bentley DR. A physical map of the mouse genome. Nature 418: 743–750, 2002. [DOI] [PubMed] [Google Scholar]
- 15.Griffin JL, Shockcor JP. Metabolic profiles of cancer cells. Nat Rev Cancer 4: 551–561, 2004. [DOI] [PubMed] [Google Scholar]
- 16.Gungor N, Godschalk RW, Pachen DM, Van Schooten FJ, Knaapen AM. Activated neutrophils inhibit nucleotide excision repair in human pulmonary epithelial cells: role of myeloperoxidase. FASEB J 21: 2359–2367, 2007. [DOI] [PubMed] [Google Scholar]
- 17.Hardie WD, Prows DR, Leikauf GD, Korfhagen TR. Attenuation of acute lung injury in transgenic mice expressing human transforming growth factor-α. Am J Physiol Lung Cell Mol Physiol 277: L1045–L1050, 1999. [DOI] [PubMed] [Google Scholar]
- 18.Hoymann HG Invasive and noninvasive lung function measurements in rodents. J Pharmacol Toxicol Methods 55: 16–26, 2007. [DOI] [PubMed] [Google Scholar]
- 19.Hoymann HG New developments in lung function measurements in rodents. Exp Toxicol Pathol 57, Suppl 2: 5–11, 2006. [DOI] [PubMed] [Google Scholar]
- 20.Irvin CG, Bates JH. Measuring the lung function in the mouse: the challenge of size. Respir Res 4: 4, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Johnson KA Imaging techniques for small animal imaging models of pulmonary disease: micro-CT. Toxicol Pathol 35: 59–64, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kenyon NJ, van der Vliet A, Schock BC, Okamoto T, McGrew GM, Last JA. Susceptibility to ozone-induced acute lung injury in iNOS-deficient mice. Am J Physiol Lung Cell Mol Physiol 282: L540–L545, 2002. [DOI] [PubMed] [Google Scholar]
- 23.Kim J, Remick DG. Tumor necrosis factor inhibitors for the treatment of asthma. Curr Allergy Asthma Rep 7: 151–156, 2007. [DOI] [PubMed] [Google Scholar]
- 24.Knaapen AM, Gungor N, Schins RP, Borm PJ, Van Schooten FJ. Neutrophils and respiratory tract DNA damage and mutagenesis: a review. Mutagenesis 21: 225–236, 2006. [DOI] [PubMed] [Google Scholar]
- 25.Marzola P, Lanzoni A, Nicolato E, Di Modugno V, Cristofori P, Osculati F, Sbarbati A. 1H MRI of pneumococcal pneumonia in a murine model. J Magn Reson Imaging 22: 170–174, 2005. [DOI] [PubMed] [Google Scholar]
- 26.Meyerspeer M, Kemp GJ, Mlynarik V, Krssak M, Szendroedi J, Nowotny P, Roden M, Moser E. Direct noninvasive quantification of lactate and high energy phosphates simultaneously in exercising human skeletal muscle by localized magnetic resonance spectroscopy. Magn Reson Med 57: 654–660, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nicholson JK, Lindon JC, Holmes E. “Metabonomics”: understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 29: 1181–1189, 1999. [DOI] [PubMed] [Google Scholar]
- 28.Niemann CU, Hirose R, Liu T, Behrends M, Brown JL, Kominsky DF, Roberts JP, Serkova N. Ischemic preconditioning improves energy state and transplantation survival in obese Zucker rat livers. Anesth Analg 101: 1577–1583, 2005. [DOI] [PubMed] [Google Scholar]
- 29.Parker JC, Townsley MI. Evaluation of lung injury in rats and mice. Am J Physiol Lung Cell Mol Physiol 286: L231–L246, 2004. [DOI] [PubMed] [Google Scholar]
- 30.Pautler RG Mouse MRI: concepts and applications in physiology. Physiology (Bethesda) 19: 168–175, 2004. [DOI] [PubMed] [Google Scholar]
- 31.Petty JM, Sueblinvong V, Lenox CC, Jones CC, Cosgrove GP, Cool CD, Rai PR, Brown KK, Weiss DJ, Poynter ME, Suratt BT. Pulmonary stromal-derived factor-1 expression and effect on neutrophil recruitment during acute lung injury. J Immunol 178: 8148–8157, 2007. [DOI] [PubMed] [Google Scholar]
- 32.Pomper MG, Lee JS. Small animal imaging in drug development. Curr Pharm Des 11: 3247–3272, 2005. [DOI] [PubMed] [Google Scholar]
- 33.Reinhard C, Eder G, Fuchs H, Ziesenis A, Heyder J, Schulz H. Inbred strain variation in lung function. Mamm Genome 13: 429–437, 2002. [DOI] [PubMed] [Google Scholar]
- 34.Sample AK, Czuprynski CJ. Priming and stimulation of bovine neutrophils by recombinant human interleukin-1 alpha and tumor necrosis factor alpha. J Leukoc Biol 49: 107–115, 1991. [DOI] [PubMed] [Google Scholar]
- 35.Schuster DP, Kovacs A, Garbow J, Piwnica-Worms D. Recent advances in imaging the lungs of intact small animals. Am J Respir Cell Mol Biol 30: 129–138, 2004. [DOI] [PubMed] [Google Scholar]
- 36.Schutze S, Machleidt T, Kronke M. The role of diacylglycerol and ceramide in tumor necrosis factor and interleukin-1 signal transduction. J Leukoc Biol 56: 533–541, 1994. [DOI] [PubMed] [Google Scholar]
- 37.Serkova N, Fuller TF, Klawitter J, Freise CE, Niemann CU. H-NMR-based metabolic signatures of mild and severe ischemia/reperfusion injury in rat kidney transplants. Kidney Int 67: 1142–1151, 2005. [DOI] [PubMed] [Google Scholar]
- 38.Serkova NJ, Jackman M, Brown JL, Liu T, Hirose R, Roberts JP, Maher JJ, Niemann CU. Metabolic profiling of livers and blood from obese Zucker rats. J Hepatol 44: 956–962, 2006. [DOI] [PubMed] [Google Scholar]
- 39.Serkova NJ, Niemann CU. Pattern recognition and biomarker validation using quantitative 1H-NMR-based metabolomics. Expert Rev Mol Diagn 6: 717–731, 2006. [DOI] [PubMed] [Google Scholar]
- 40.Singleton KD, Serkova N, Banerjee A, Meng X, Gamboni-Robertson F, Wischmeyer PE. Glutamine attenuates endotoxin-induced lung metabolic dysfunction: potential role of enhanced heat shock protein 70. Nutrition 21: 214–223, 2005. [DOI] [PubMed] [Google Scholar]
- 41.Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, Fujimoto EK, Goeke NM, Olson BJ, Klenk DC. Measurement of protein using bicinchoninic acid. Anal Biochem 150: 76–85, 1985. [DOI] [PubMed] [Google Scholar]
- 42.Stringer KA, Hybertson BM, Cho OJ, Cohen Z, Repine JE. Tissue plasminogen activator (tPA) inhibits interleukin-1 induced acute lung leak. Free Radic Biol Med 25: 184–188, 1998. [DOI] [PubMed] [Google Scholar]
- 43.Stringer KA, Tobias M, O'Neill HC, Franklin CC. Cigarette smoke extract-induced suppression of caspase-3-like activity impairs human neutrophil phagocytosis. Am J Physiol Lung Cell Mol Physiol 292: L1572–L1579, 2007. [DOI] [PubMed] [Google Scholar]
- 44.Su X, Looney M, Robriquet L, Fang X, Matthay MA. Direct visual instillation as a method for efficient delivery of fluid into the distal airspaces of anesthetized mice. Exp Lung Res 30: 479–493, 2004. [DOI] [PubMed] [Google Scholar]
- 45.Tigani B, Cannet C, Karmouty-Quintana H, Ble FX, Zurbruegg S, Schaeublin E, Fozard JR, Beckmann N. Lung inflammation and vascular remodeling after repeated allergen challenge detected noninvasively by MRI. Am J Physiol Lung Cell Mol Physiol 292: L644–L653, 2007. [DOI] [PubMed] [Google Scholar]
- 46.Tournebize R, Doan BT, Dillies MA, Maurin S, Beloeil JC, Sansonetti PJ. Magnetic resonance imaging of Klebsiella pneumoniae-induced pneumonia in mice. Cell Microbiol 8: 33–43, 2006. [DOI] [PubMed] [Google Scholar]
- 47.van Rijt LS, Kuipers H, Vos N, Hijdra D, Hoogsteden HC, Lambrecht BN. A rapid flow cytometric method for determining the cellular composition of bronchoalveolar lavage fluid cells in mouse models of asthma. J Immunol Methods 288: 111–121, 2004. [DOI] [PubMed] [Google Scholar]
- 48.Yokoyama T, Tomiguchi S, Nishi J, Yamashita Y, Ichikado K, Gushima Y, Ando M, Takahashi M. Hyperoxia-induced acute lung injury using a pig model: correlation between MR imaging and histologic results. Radiat Med 19: 131–143, 2001. [PubMed] [Google Scholar]
- 49.Zhou Z, Kozlowski J, Schuster DP. Physiologic, biochemical, and imaging characterization of acute lung injury in mice. Am J Respir Crit Care Med 172: 344–351, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]