

Abstract
The genotypic proportions for major histocompatibility complex loci, HLA-A and HLA-B, of progeny in families in 23 South Amerindian tribes in which segregation for homozygotes and heterozygotes could occur are examined. Overall, there is a large deficiency of homozygotes compared with Mendelian expectations (for HLA-A, 114 observed and 155.50 expected and for HLA-B 110 observed and 144.75 expected), consistent with strong balancing selection favoring heterozygotes. There is no evidence that these deficiencies were associated with particular alleles or with the age of the individuals sampled. When these families were divided into four mating types, there was strong selection against homozygotes, averaging 0.462 for three of the mating types over the two loci. For the other mating type in which the female parent is homozygous and shares one allele with the heterozygous male parent, there was no evidence of selection against homozygotes. A theoretical model incorporating these findings surprisingly does not result in a stable polymorphism for two alleles but does result in an excess of heterozygotes and a minimum fitness at intermediate allele frequencies. However, for more than two alleles, balancing selection does occur and the model approaches the qualities of the symmetrical heterozygote advantage model as the number of alleles increases.
Keywords: heterozygote advantage, maternal–fetal interaction, major histocompatibility complex, polymorphism, reciprocal matings
There is a near-unanimous consensus that variation at the major histocompatibility complex (MHC; HLA in humans) is maintained by balancing selection (1–3). The evidence supporting the importance of balancing selection is of several types. First, there are data resulting from the cumulative effects of generations, such as a more uniform frequency of alleles than expected under neutrality (4) and the higher rate of nonsynonymous than synonymous substitution (5, 6) and higher heterozygosity (7) at functionally important amino acid positions. Second, there are data that must be the result of selection acting in the present generation, such as an excess of heterozygotes compared with Hardy–Weinberg proportions (8, 9) or an excess of heterozygotes in families compared with Mendelian expectations (8).
In spite of this universal recognition of balancing selection at MHC loci, there are quite different opinions about the magnitude of these selective effects. For example, it has been suggested that the selective difference (s) between homozygotes and heterozygotes may be relatively small, between 0.01 and 0.1 (1). Further, from sequence data, it has been estimated that in humans s ranges from 0.0007 (for locus DPB1) to 0.042 (HLA-B) with a mean over seven loci of 0.013 (ref. 10; see ref. 11). On the other hand, to explain the population deficiency of homozygotes for HLA-A and HLA-B in the Havasupai, a remote tribe from Arizona (9), an s value of 0.35 is necessary. The reduced proportion of HLA-A and HLA-B homozygotes observed in population samples of South Amerindians (8) needs an s value of 0.425 to explain the deficiency (12). These results may not necessarily be in conflict if by chance the observations in refs. 8 and 9 are high values that would average away with lower values in the past. In fact, there has probably been strong selection in recent generations, if not the present one (3, 13).
Here, we present data showing a large excess of heterozygotes (a large deficiency of homozygotes) compared with Mendelian expectations in 510 offspring from parents with known genotypes. The matings are divided into four general categories based on the parental genotypes. The magnitude of selection against homozygotes from three of these mating types is quite large. For the other mating type, in which the mother is homozygous and the father is heterozygous and shares one allele with the mother, there does not appear to be selection against homozygotes. A theoretical model consistent these data has the surprising property of neutrality with two alleles (see ref. 14) but leads to balancing selection with multiple alleles as shown below.
MATERIALS AND METHODS
Sample.
The sample consists of 510 offspring, 0–53 years old, whose parents were known. The individuals are from 23 tribes in the Amazon and Orinoco basins (Fig. 1, Table 1). The sample is broadly representative of the indigenous population of these lowlands both geographically and linguistically (15) and, with a few recent exceptions, marriages appear to have been within tribes. Persons with contemporary non-Indian admixture were excluded at the time of typing, and five other individuals found to have foreign alleles in the course of typing have also been excluded. The sample may not be completely representative of the whole population at either end of the age scale because there is not a representative number of babies and only persons whose parents survived were included.
Figure 1.

Map of the lower Amazon and Orinoco basins, showing the location of the 23 tribal groups included in the study. The numbers refer to the tribes listed in Table 1.
Table 1.
The 23 tribes (with the numbers used in Fig. 1), the number of children (N) with parents of known HLA genotypes, and the observed (O) and expected (E) numbers of homozygotes in children from parents that are potentially segregating for homozygotes
| Tribe | N |
HLA-A
|
HLA-B
|
||||
|---|---|---|---|---|---|---|---|
| NA | O | E | NB | O | E | ||
| 1. Warao | 14 | 14 | 4 | 5.25 | 13 | 4 | 5.25 |
| 2. Palikur | 3 | 2 | 1 | 0.50 | 3 | 0 | 1.00 |
| 3. Tiriyo | 32 | 20 | 3 | 7.00 | 28 | 5 | 9.25 |
| 4. Molokopote | 5 | 5 | 1 | 2.25 | 5 | 0 | 1.50 |
| 5. Waiápi | 55 | 49 | 12 | 16.50 | 38 | 10 | 12.25 |
| 6. Apalai | 41 | 41 | 7 | 14.75 | 27 | 7 | 8.25 |
| 7. Kaxuwana | 3 | 3 | 2 | 1.25 | 2 | 1 | 0.50 |
| 8. Urubu-Kaapor | 53 | 36 | 12 | 14.50 | 37 | 9 | 13.50 |
| 9. Asurini Trocara | 53 | 27 | 10 | 12.00 | 22 | 8 | 9.25 |
| 10. Asurini Koatinema | 6 | 1 | 0 | 0.25 | 4 | 1 | 1.50 |
| 11. Arara | 21 | 16 | 2 | 6.25 | 20 | 4 | 6.75 |
| 12. Paracaná Novo | 11 | 11 | 2 | 2.25 | 11 | 7 | 4.75 |
| 13. Paracaná Velho | 46 | 46 | 16 | 17.25 | 45 | 13 | 17.25 |
| 14. Arawete | 1 | 0 | 0 | 0.00 | 1 | 0 | 0.50 |
| 15. Paratintins | 2 | 2 | 0 | 0.50 | 2 | 1 | 1.00 |
| 16. Jamamadi | 19 | 14 | 2 | 7.00 | 19 | 3 | 9.50 |
| 17. Xikrin | 22 | 20 | 4 | 6.50 | 15 | 6 | 6.75 |
| 18. Mundurucu | 45 | 36 | 6 | 10.75 | 19 | 4 | 7.75 |
| 19. Kayapo | 14 | 12 | 6 | 6.75 | 13 | 5 | 5.25 |
| 20. Karitiana | 34 | 33 | 10 | 15.00 | 33 | 13 | 15.00 |
| 21. Surui | 4 | 4 | 3 | 2.00 | 4 | 3 | 2.00 |
| 22. Uru-eu-uau-uau | 2 | 2 | 2 | 0.50 | 2 | 2 | 0.50 |
| 23. Cinta Larga | 24 | 17 | 9 | 6.50 | 13 | 4 | 5.50 |
| Total | 510 | 411 | 114 | 155.50 | 376 | 110 | 144.75 |
NA and NB indicate the number of children potentially segregating for HLA-A and HLA-B, respectively.
Methods.
The parents and their children were typed for HLA-A and HLA-B using traditional serotyping protocols (15, 16). At HLA-A, four different alleles were observed—02, 24, 31, and 68—and for HLA-B, five different alleles were observed—05, 15, 35, 39, and 40. There is no evidence of serological blank alleles in these samples for either HLA-A or HLA-B or in the larger population samples from these groups (F.L.B., unpublished data), suggesting that if blank alleles were present, they were at quite low frequency. The heterozygosities from the population samples averaged approximately 0.7 for both loci, so there was a substantial degree of polymorphism in the populations. However, there was much lower serotypic variation than observed in European or African populations, which may have 15–18 alleles for HLA-A and 25–30 alleles for HLA-B (4).
From the studies summarized in ref. 3, it appears that three of the serotypic alleles at HLA-A—A*2402, A*31012, and A*68012—are probably homogeneous for their nucleotide sequence, as they are in other South Amerindian tribes (the first two digits indicate the serotype and the remaining digits indicate a specific allelic sequence). The other allele, 02, shows heterogeneity, so we do not know at present which 02 allele or alleles are in our samples. The picture for HLA-B is quite different, with every serotype found in the 11 populations showing heterogeneity in South Amerindians. There may have been only one founding allele for three of these serotypes—namely, B*1501, B*3501, and B*39011—whereas both 40 and 51 had two founding alleles (3). Assuming a similar difference between loci in this sample, families are less likely to have undocumented variation for HLA-A than for HLA-B.
There are four basic types of matings in which there is segregation for homozygotes and heterozygotes. First, there are three types of matings in which half the progeny are expected to be heterozygotes and half are expected to be homozygotes, AiAi × AiĀi and AiĀi × AiAi, which are reciprocal matings in which the first individual is a female and second a male (the overbar indicates some allele other than Ai) and AiAj × AiAj in which the two parents share both alleles. If we let the observed number of homozygotes AiAi and heterozygotes AiĀi from these matings be Nii and Ni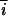 , respectively, then a nearly unbiased estimate of the selective difference between heterozygotes (with a fitness of 1) and homozygotes (with a fitness of 1 − s) is
, respectively, then a nearly unbiased estimate of the selective difference between heterozygotes (with a fitness of 1) and homozygotes (with a fitness of 1 − s) is
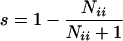 |
1 |
(17).
There is also a mating in which 3/4 of the progeny are expected to be heterozygotes and 1/4 are expected to be homozygotes–i.e., AiAj × AiAk where each parent has one copy of allele Ai and the other alleles, Aj and Ak, are different from Ai and from each other. In this case, a nearly unbiased estimate of s is
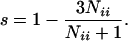 |
2 |
We note that progeny from families of known parents can be aggregated over tribes without resulting in a deficiency of heterozygotes that may occur when population data are aggregated over tribes.
Segregation of homozygotes and heterozygotes for both loci was also examined for each allele and for four age classes (0–4 years old, 5–9 years old, 10–14 years old, and >14 years old. Significance of homogeneity over allele or over age classes was determined by using simulation of 10,000 random samples. In these tests for the given expected number of homozygotes in different allele or age classes, random samples using the overall deficiency of homozygotes for the locus were drawn and the observed variation in deficiency over classes was compared with the distribution of randomly generated values.
RESULTS
Table 1 gives the number of children (N), 510 over all the tribes, with known HLA genotypes from parents with known genotypes and the observed and expected number of homozygotes based on expected Mendelian proportions. The proportion of the total sample segregating for homozygotes and heterozygotes for the two loci (see NA and NB totals in Table 1) is 80% and 74% for HLA-A and HLA-B, respectively. Most parents in the excluded proportion are segregating only for different heterozygous progeny with a smaller number homozygous for the same allele.
For both loci HLA-A and HLA-B, there is a substantial deficiency of homozygotes (excess of heterozygotes) over that expected, 26.7% for HLA-A and 24.0% for HLA-B. In the 15 tribes with 10 or more progeny, there was a deficiency of homozygotes in 14 of the tribes for both HLA-A and HLA-B.
Table 2 gives the observed and expected number of heterozygotes and homozygotes for the four types of matings described above summed over the 23 tribes. The observed numbers are different from expected for mating type AiAj × AiAk for HLA-A but not for HLA-B and significant for the mating types AiĀi × AiAi and AiAj × AiAj for both loci. The average estimate of s for these three matings types over both loci is 0.462.
Table 2.
The total observed (O) and expected (E) numbers of heterozygotes and homozygotes for the four types of matings that potentially segregate for homozygotes and the estimated selective difference between heterozygotes and homozygotes
| Locus | Female × Male | Heterozygotes
|
Homozygotes
|
χ2 | s | ||
|---|---|---|---|---|---|---|---|
| O | E | O | E | ||||
| HLA-A | AiAj × AiAk | 169 | 151.50 | 33 | 50.50 | 8.08** | 0.418 |
| AiAi × AiĀi | 39 | 38.50 | 38 | 38.50 | 0.01 | (0.050) | |
| AiĀi × AiAi | 34 | 26.50 | 19 | 26.50 | 4.25* | 0.457 | |
| AiAj × AiAj | 56 | 40.00 | 24 | 40.00 | 12.80*** | 0.579 | |
| Total | 298 | 256.50 | 114 | 155.50 | |||
| HLA-B | AiAj × AiAk | 140 | 132.75 | 37 | 44.25 | 1.58 | (0.213) |
| AiAi × AiĀi | 36 | 33.00 | 30 | 33.00 | 0.54 | (0.189) | |
| AiĀi × AiAi | 37 | 26.00 | 15 | 26.00 | 9.31** | 0.605 | |
| AiAj × AiAj | 55 | 41.50 | 28 | 41.50 | 8.78** | 0.500 | |
| Total | 268 | 233.25 | 110 | 144.75 | |||
Values of s are given in parentheses when χ2 was not significant. ∗, P < 0.05; ∗∗, P < 0.01; ∗∗∗, P < 0.001.
On the other hand, when the female is homozygous and the male shares one allele with her, mating AiAi × AiĀi, the observed numbers are not significantly different from the expected numbers for either locus. Because the reciprocal mating (AiĀi × AiAi) does give progeny numbers that differ from expected, it appears that some type of maternal–fetal interaction is of significance in influencing the genotypes of the progeny in these matings.
To determine whether the deficiency of homozygous progeny is caused by homozygosity at particular alleles, we calculated the observed and expected numbers of homozygotes for each allele separately (Table 3). There is some variation in the ratio of observed to expected for different alleles, but this variation is not significantly different from that observed by chance (P = 0.444 and P = 0.273 for HLA-A and HLA-B), suggesting that the deficiency is not associated with particular alleles at either locus.
Table 3.
The observed and expected numbers of homozygotes for each of the alleles at HLA-A and HLA-B and the ratio of observed to expected (O/E) for each allele
| Locus | Allele | Observed | Expected | O/E |
|---|---|---|---|---|
| HLA-A | 02 | 41 | 53.00 | 0.774 |
| 24 | 32 | 44.25 | 0.723 | |
| 31 | 34 | 46.00 | 0.739 | |
| 68 | 7 | 12.25 | 0.571 | |
| HLA-B | 15 | 25 | 32.00 | 0.781 |
| 35 | 46 | 61.75 | 0.745 | |
| 39 | 21 | 24.50 | 0.857 | |
| 40 | 11 | 17.00 | 0.647 | |
| 51 | 7 | 9.50 | 0.737 |
If the deficiency of homozygotes were associated with cumulative mortality with age, then the deficiency should increase in older age classes. Table 4 gives the deficiency for four age classes, individuals in the first three 5-year intervals and those over 14. There is some variation in deficiency over the age classes, with slightly greater deficiency for both loci in the >14 class. However, this variation is no different than expected at random (P = 0.429 and P = 0.404 for HLA-A and HLA-B). As a result, there does not appear to be an association of homozygote deficiency with age.
Table 4.
The observed and expected numbers of homozygotes for individuals of different age classes for HLA-A and HLA-B and the ratio of observed to expected (O/E) for each age group
| Locus | Age class | Observed | Expected | O/E |
|---|---|---|---|---|
| HLA-A | 0–4 | 30 | 40.00 | 0.750 |
| 5–9 | 35 | 47.25 | 0.741 | |
| 10–14 | 26 | 32.75 | 0.794 | |
| >14 | 23 | 35.50 | 0.648 | |
| HLA-B | 0–4 | 28 | 38.75 | 0.723 |
| 5–9 | 33 | 41.50 | 0.795 | |
| 10–14 | 23 | 27.75 | 0.829 | |
| >14 | 26 | 36.75 | 0.707 |
MODEL
Two Alleles.
A two-allele model to describe the findings in Table 2—i.e., selection against homozygotes in all matings with segregation except when the mother is homozygous and the father is heterozygous and shares an allele with the mother—is given in Table 5 (14). Alleles A1 and A2 are assumed to have frequencies of p1 and p2 and genotypes A1A1, A1A2, and A2A2 frequencies of P11, P12, and P22, respectively. The mean fitness is w̄ = 1 − (sP12/2) and the expected change in the frequency of A1 is
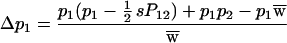 |
3 |
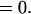 |
Quite surprisingly, there is no change in allelic frequency for any value of p1. The equilibrium frequency of heterozygotes is calculated by letting P12.e = P12 so that
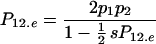 |
4 |
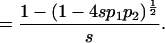 |
Therefore, the mean fitness at equilibrium is w̄ = 1 − (sP12.e/2).
Table 5.
A two-allele selection model to describe the segregation observed in families for HLA-A and HLA-B
| Parents
|
Progeny | |||
|---|---|---|---|---|
| Female × Male | Frequency | A1A1 | A1A2 | A2A2 |
| A1A1 × A1A1 | P112 | 1 | — | — |
| A1A1 × A1A2 | P11P12 | 1 | 1 | — |
| A1A1 × A2A2 | P11P22 | — | 1 | — |
| A1A2 × A1A1 | P11P12 | 1 − s | 1 | — |
| A1A2 × A1A2 | P122 | 1 − s | 1 | 1 − s |
| A1A2 × A2A2 | P12P22 | — | 1 | 1 − s |
| A2A2 × A1A1 | P11P22 | — | 1 | — |
| A2A2 × A1A2 | P12P22 | — | 1 | 1 |
| A2A2 × A2A2 | P222 | — | — | 1 |
| p1(p1 − ½ sP12) | 2p1p2 | p2(p2 − ½ sP12) | ||
| 1 − ½ sP12 | 1 − ½ sP12 | 1 − ½ sP12 | ||
The mean fitness is a function of allelic frequencies with a minimum at p1 = p2 and a maximum when p1 = 0 or 1, a pattern reminiscent of selection against heterozygotes (18). The fixation index F = 1 − (sP12.e/2p1p2) is negative, a pattern reminiscent of selection favoring heterozygotes (18).
Multiple Alleles.
The above model can be extended to multiple alleles by using the approach outlined above to better describe the highly polymorphic situation in our HLA data and that in other populations. For multiple alleles, the frequencies of the homozygotes and heterozygotes are
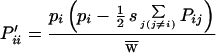 |
5a |
and
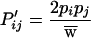 |
5b |
where
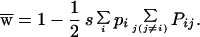 |
5c |
When there are three or more alleles, the unusual properties of the two-allele model disappear and it becomes a model of balancing selection. In particular, given two alleles and the generation of a third rarer allele by mutation, this new allele has a selective advantage and will increase. For situations with three or more alleles, such as exist for most MHC genes, this model predicts that selection will maintain a stable polymorphism for multiple alleles.
To determine how effective this model is in maintaining polymorphism, the change in allelic frequency for different numbers of alleles, selection coefficients, and allelic frequencies were numerically compared with a symmetric heterozygote advantage model in which all homozygotes have a fitness of 1 − s and all heterozygotes a fitness of 1. For three alleles, the change in allelic frequency was generally between 40% and 50% that of the heterozygote advantage model with the same s. As the number of alleles increases, this model quickly approaches that of heterozygote advantage and, for example, with eight alleles, the change in allelic frequency is generally between 80% and 90% that of the heterozygote advantage model with the same s.
The basis of this similarity is apparent if it is assumed for illustration that genotypes are close to Hardy–Weinberg proportions. With this assumption, expressions 5a and 5c become
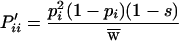 |
and
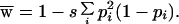 |
As the number of alleles increases so that in general pi decreases, 1 − pi approaches unity, and these expressions approach the comparable expressions for heterozygote advantage.
DISCUSSION
From these data, it is clear that there is strong selection against homozygotes in the present generation in these South Amerindian families. The amount of selection approaches 50% over three of the mating types in which there is segregation for homozygotes and heterozygotes. This selection is much larger than that suggested by the authors of refs. 1 and 10. Because their estimates are based on selection over many generations, their estimates could be expected to be less if, for example, selection was lower in the past and is greater in the present generation.
Because of the difference between reciprocal matings in segregation proportions (no deficiency of homozygotes when the mother is homozygous and father heterozygous and a large deficiency when the mother is heterozygous and father homozygous), it appears that an interaction between the mother and the fetus is important in causing the strong selection against homozygotes. The exact mechanistic basis of this interaction is not known at present, but it appears to be different from previously described patterns of maternal–fetal interaction. A model to describe these data has surprising findings for two alleles but approaches a heterozygote advantage model with increasing numbers of alleles, making it consistent with previous models of balancing selection at MHC.
The higher heterogeneity within serotype for HLA-B than HLA-A in South Amerindians (3) suggests that a number of HLA-B serotypic homozygotes may in fact be heterozygotes at the sequence level. If variation within serotype made a selective difference, then one would expect that the deficiency of homozygotes would be less for HLA-B than HLA-A. However, the level of homozygote deficiency is very similar for HLA-A and HLA-B, suggesting that the within-serotypic heterozygotes may be similar in their selective effects to sequence homozygotes.
Several modes of balancing selection have been proposed as important in the maintenance of variation at the MHC: namely, parasite resistance, maternal–fetal interaction, and negative-assortative mating (2). Of special interest here is that a number of studies indicate that a significant number of couples with a history of spontaneous abortions share antigens for HLA loci (19, 20). The immunological explanation for these observations is that an immune response occurring when the mother and fetus differ at HLA loci is necessary for proper implantation and fetal growth.
Hedrick and Thomson (21) developed a model consistent with the observation of higher spontaneous abortion associated with higher sharing of HLA alleles between parents. This model has the potential to maintain many alleles at a single locus and to result in substantial gametic disequilibrium between linked loci; however, this model is quite different from the model given in Table 5. For example, in the single-locus version of the model of ref. 21, if a progeny shared one or two alleles with its mother, then the fitness of the progeny was 1 or 1 − s, respectively, independent of parental mating type.
A major function of MHC histocompatibility molecules is to present foreign antigens (peptides) to T cells to stimulate an immune response against invading parasites (22). Histocompatibility alleles have been shown to differ in their ability to create an immune response to a variety of infectious agents, suggesting that the epidemic diseases of the human past may have played a central role in determining HLA allelic frequencies observed in human populations today, particularly in South Amerindians (13). It is possible that the results presented here could occur because homozygous progeny had lower survival relative to heterozygotes (e.g., lower resistance to infectious diseases) when their mothers were heterozygous but that there was no difference in relative survival between genotypes when the mother was homozygous. On the other hand, the deficiency of homozygotes observed in this study does not appear to increase significantly with age as expected if there were increased relative mortality of homozygotes due to increased exposure to infectious diseases with age. In fact, the lack of association of homozyote deficiency and age supports the importance of immune reactions between mother and fetus.
Finally, there have been studies in laboratory mice demonstrating mating preference based on MHC type (23). However, there is no published evidence of nonrandom mating in humans for HLA although Wedekind et al. (24) claim to have shown female HLA preferences for T-shirts worn by males (however, see refs. 25 and 26). Most importantly for the present study, there was no evidence for nonrandom mating with respect to HLA-A or HLA-B for the parents of the progeny in this study for the 11 tribes with the largest number of known matings (26).
Acknowledgments
We appreciate the comments of William Klitz.
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
Abbreviation: MHC, major histocompatibility complex.
References
- 1.Nei M, Hughes A. In: Evolution at the Molecular Level. Selander R K, Clark A G, Whittam T S, editors. Sunderland, MA: Sinauer; 1991. pp. 222–247. [Google Scholar]
- 2.Hedrick P W. Am Nat. 1994;143:945–964. [Google Scholar]
- 3.Parham P, Ohta T. Science. 1996;272:67–74. doi: 10.1126/science.272.5258.67. [DOI] [PubMed] [Google Scholar]
- 4.Hedrick P W, Thomson G. Genetics. 1983;104:449–456. doi: 10.1093/genetics/104.3.449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hughes A L, Nei M. Nature (London) 1988;335:167–179. doi: 10.1038/335167a0. [DOI] [PubMed] [Google Scholar]
- 6.Hughes A L, Nei M. Proc Natl Acad Sci USA. 1989;86:958–962. doi: 10.1073/pnas.86.3.958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hedrick P W, Whittam T S, Parham P. Proc Natl Acad Sci USA. 1991;88:5897–5901. doi: 10.1073/pnas.88.13.5897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Black F L, Salzano F M. Am J Hum Genet. 1981;33:894–899. [PMC free article] [PubMed] [Google Scholar]
- 9.Markow T, Hedrick P W, Zuerlin K, Danilovs J, Martin J, Vyvial T, Armstrong C. Am J Hum Genet. 1993;53:943–952. [PMC free article] [PubMed] [Google Scholar]
- 10.Satta S, O’Huigin C, Takahata N, Klein J. Proc Natl Acad Sci USA. 1994;91:7184–7188. doi: 10.1073/pnas.91.15.7184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hedrick P W, Kim T J. In: Evolutionary Genetics from Molecules to Morphology. Singh R S, Krimbas C K, editors. New York: Cambridge Univ. Press; 1997. , in press. [Google Scholar]
- 12.Hedrick P W. Hum Hered. 1990;40:213–220. doi: 10.1159/000153933. [DOI] [PubMed] [Google Scholar]
- 13.Black F L. Perspect Biol Med. 1994;37:292–307. doi: 10.1353/pbm.1994.0030. [DOI] [PubMed] [Google Scholar]
- 14.Hedrick P W. Nature (London) 1997;387:138. doi: 10.1038/387138a0. [DOI] [PubMed] [Google Scholar]
- 15.Bhatia K K, Black F L, Smith T A, Prasad M L, Koki G N. Am J Hum Anthropol. 1995;97:291–305. doi: 10.1002/ajpa.1330970304. [DOI] [PubMed] [Google Scholar]
- 16.Black F L, Berman L L, Gabbay Y. Tissue Antigens. 1980;16:368–376. doi: 10.1111/j.1399-0039.1980.tb00317.x. [DOI] [PubMed] [Google Scholar]
- 17.Haldane J B S. J Genet. 1956;54:294–296. [Google Scholar]
- 18.Hedrick P W. Genetics of Populations. Boston: Jones and Bartlett; 1985. [Google Scholar]
- 19.Thomas M L, Harger J H, Wagener D K, Rabin B S, Gill T J. Am J Obstet Gynecol. 1985;151:1053–1058. doi: 10.1016/0002-9378(85)90379-5. [DOI] [PubMed] [Google Scholar]
- 20.Alberts S C, Ober C. Yearb Phys Anthropol. 1993;36:71–89. [Google Scholar]
- 21.Hedrick P W, Thomson G. Genetics. 1988;119:205–212. doi: 10.1093/genetics/119.1.205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zinkernagel R M. Annu Rev Microbiol. 1979;33:201–213. doi: 10.1146/annurev.mi.33.100179.001221. [DOI] [PubMed] [Google Scholar]
- 23.Brown J L, Eklund A. Am Nat. 1994;143:435–461. [Google Scholar]
- 24.Wedekind C, Seebeck R, Bettens F, Paeke A J. Proc R Soc London. 1995;260:245–249. doi: 10.1098/rspb.1995.0087. [DOI] [PubMed] [Google Scholar]
- 25.Hedrick P W, Loeschcke V. Trends Ecol Evol. 1996;11:24. doi: 10.1016/0169-5347(96)80237-0. [DOI] [PubMed] [Google Scholar]
- 26.Hedrick, P. W. & Black, F. L. (1997) Am. J. Hum. Genet. 61, in press. [DOI] [PMC free article] [PubMed]