

Abstract
Thymus-derived regulatory T lymphocytes of CD4+CD25+ phenotype regulate a large variety of beneficial and deleterious immune-responses and can inhibit lethal graft-versus-host disease in rodents. In vitro, CD4+CD25+ T cells require specific MHC/peptide ligands for their activation, but once activated they act in an antigen-non-specific manner. In vivo, regulatory T cells are also activated in an antigen-specific fashion, but nothing is known about antigen-specificity of their suppressor effector function. Here we show that CD4+CD25+ regulatory T lymphocytes isolated from naive mice and activated in vitro with allogeneic APC, induced specific long-term tolerance to bone-marrow grafts disparate for major and minor histocompatibility antigens : Whereas “target” bone-marrow was protected, third-party bone-marrow was rejected. Importantly, in mice injected with a mix of target and third-party bone-marrows, protection and rejection processes took place simultaneously. These results indicate that CD4+CD25+ regulatory T cells can act in an antigen-specific manner in vivo. Our results suggest that CD4+CD25+ regulatory T cells could in the future be used in clinical settings to induce specific immunosuppression.
Keywords: Animals, Bone Marrow Cells, immunology, Bone Marrow Transplantation, immunology, CD4-Positive T-Lymphocytes, chemistry, cytology, immunology, CD8-Positive T-Lymphocytes, cytology, immunology, Cells, Cultured, Epitopes, Graft Rejection, immunology, Haplotypes, Histocompatibility Antigens, genetics, Immune Tolerance, immunology, Mice, Mice, Inbred C57BL, Mice, Inbred DBA, Receptors, Interleukin-2, analysis, Spleen, cytology, immunology, Transplantation, Homologous
Introduction
Due to the random rearrangements of genes encoding T and B-lymphocyte antigen-receptors, a significant number of autospecific and potentially autoreactive lymphocytes develop in primary lymphoid organs 1–3. Central tolerance (i.e. induced in primary lymphoid organs) eliminates (by deletion) or functionally inactivates (by induction of anergy) such dangerous lymphocytes. In absence of central tolerance-induction, a strongly self-reactive T cell repertoire develops 4,5. However, when central tolerance is partially defective, self-tolerance can be maintained by peripheral mechanisms 6–8. Several types of peripheral tolerance mechanisms control lymphocytes having escaped central-tolerance and are known to play a crucial role in preventing autoimmunity (for reviews see refs. 6,9).
One of these peripheral tolerance mechanisms was discovered using the day 3 thymectomy model of multi-organ autoimmunity in mice 10. The pathology can be prevented by injection of CD4+CD25+ lymphocytes, which appear after day 3 of life in the peripheral lymphoid organs of normal mice. CD4+CD25+ regulatory T cells do not only inhibit autoimmunity; they can also inhibit experimental inflammatory bowel disease induced by injection of CD4+CD45RBhigh cells into immunodeficient SCID mice or RAG-deficient animals 11. Moreover, they contribute to the fine control of immunity to infectious agents such as parasites and viruses 12,13. An undesired side-effect of the activity of CD4+CD25+ regulatory T lymphocytes is the occasional incapacity of the immune system to eliminate tumor cells 14,15. Therefore, regulatory T lymphocytes play a crucial role in the pathophysiological maintenance of immunological tolerance (reviewed in refs. 11,16,17,18).
Natural (i.e. thymus-derived) CD4+CD25+ regulatory T cells can induce transplantation-tolerance in mice. Lethal graft-versus-host disease (GvHD) can be significantly reduced by injection of freshly isolated or ex vivo cultured regulatory T cells 19–22. In one report, freshly isolated CD4+CD25+ regulatory T cells from naive animals were shown to induce limited tolerance to minor-histocompatibility-antigen disparate skin grafts 23. Intrathymic injection of donor-strain antigen results in a tolerant state which was shown to be due to development of CD4+CD25+ regulatory thymocytes 24.
Regulatory T cells with a CD4+CD25+ phenotype are also known to be involved in experimental systems in which tolerance to alloantigens is induced in vivo with antibodies to T cell surface-antigens (e.g. CD4, CD8, or CD154, refs. 23,25,26–29) or with the active form of vitamin D3 and mycophenolate mofetil 30. In these systems, however, it is not clear if thymus-derived regulatory T cells (that can be found in naive animals) are involved or if these cells are induced in the periphery 31,32.
Like all other TCRαβ-expressing T lymphocytes, thymus-derived CD4+CD25+ regulatory T lymphocytes are antigen-specific, at least during their activation-phase. These cells have been shown to proliferate in an antigen-specific manner in vivo 33–35. Myelin basic protein-specific CD4+CD25+ regulatory T cells protect better against experimental autoimmune encephalomyelitis than regulatory T cells with a restricted (but non-myelin basic protein-specific) TCR-repertoire 36. Regulatory T cells activated in vitro with host-type APC inhibit GvHD more potently than cells activated with third-party APC 21,22. Also experimentally induced CD4+CD25+ regulatory T cells act in a specific manner and protect target but not third-party allografts 27,29. In vitro, however, CD4+CD25+ regulatory T cells only require interaction with specific MHC/peptide complexes during their activation phase. Once activated, their suppressor effector function is completely non antigen-specific 37. The antigen-specificity observed in vivo could therefore potentially be explained by the hypothesis that regulatory T cells are activated only in hosts presenting antigens for which they are specific. In mice in which these antigens are not expressed, the regulatory T cells are not activated and immunosuppression does not take place.
It remains therefore entirely unclear if CD4+CD25+ regulatory T cells are antigen-specific during their effector phase in vivo. Such antigen-specificity is required for a potential future use of these cells in induction of specific immunosuppression in patients. Here we present an experimental model in which transplanted bone marrow, disparate for major and minor histocompatibility antigens, is protected from rejection by host T cells through injection of CD4+CD25+ regulatory T cells cultured ex vivo. Using this model we show that CD4+CD25+ regulatory T cells can act in an alloantigen-specific manner during their effector phase. The model described here should prove useful in deciphering the in vivo mechanisms of specific immunosuppression mediated by CD4+CD25+ regulatory T lymphocytes. Moreover, our data suggest a clinical potential of these cells in protocols aimed at induction of specific tolerance to allogeneic grafts.
Methods
Mice
All mice were used at 6–10 weeks of age. C57BL/6 (“B6”: H-2b), (C57BL/6xDBA/2)F1 (“B6D2F1”, H-2bd) and (C57BL/6xCBA/J)F1 (“B6CBAF1”, H-2bk) mice were purchased from Janvier (Le Genest St. Isle, France). All experiments involving animals were performed in compliance with the relevant laws and institutional guidelines (approval no. 31–13) and have been approved by the local ethics committee (ref. MP/01/31/10/03).
Purification of T-cell subsets
Erythrocyte-depleted (Lympholite-M, Cedarlane Laboratories, Hornby, Canada) splenocytes were enriched in CD4+ T cells by magnetic depletion with sheep anti-rat antibody-coated Dynabeads (Dynal Biotech, Oslo, Norway) after incubation with a cocktail of the following rat mAb: anti-NK1.1 (PK136), anti-CD8 (53.6.7), anti-FcγRIII (2.4G2) and anti-MHC class II molecules (M5). The resulting population was then labeled with PE-labeled anti-CD25 mAb PC61 (BD PharMingen, Heidelberg, Germany; eBioscience, San Diego, CA) and CD4+CD25+ cells enriched with anti-PE microbeads using the AutoMACS (Miltenyi Biotec, Paris, France). Cell purity was checked by flow-cytometry on a FACSCalibur (BD Biosciences, San Jose, CA) using PE-labeled anti-CD25 mAb PC61 and FITC-labeled anti-CD4 (GK1.5). Positively sorted CD4+CD25+ T cells were routinely >95% pure.
CD4+ or CD8+ T cells were similarly isolated using negative selection with Dynabeads (PK136, 2.4G2, M5, and 53.6.7 or GK1.5) followed by positive selection on AutoMACS (with PE-labeled anti-CD4 mAb GK1.5 or anti-CD8 mAb 53.6.7 (BD Pharmingen; eBioscience)). Purity of populations routinely exceeded 98%.
Ex vivo culture of CD4+CD25+ T Cells
2×103 CD4+CD25+ T cells/well from B6 mice were co-cultured with 5×105 γ-irradiated (1700 Rad) total splenocytes from B6D2F1 or B6CBAF1 mice in 96-well round-bottom plates for 14 days. Cells were cultured in RPMI 1640 (Eurobio, Les Ulis, France) supplemented with 10% PCS, 2 mM L-glutamine, Penicillin, Streptomycin, 10 mM Hepes, 50 μM 2-ME, 1 mM non essential amino acids, 1 mM sodium pyruvate and 100 U/ml IL-2 (supernatant of PMA-stimulated EL4.IL-2 cells; ATCC, Manassas, VA). At day 7, 100μl of fresh medium was added and cells were cultured for another 7 days.
Bone marrow chimeras
Bone marrow from femurs and tibias was collected in DMEM (Eurobio, Les Ulis, France) supplemented with 10% FCS, 2 mM L-glutamine, Penicillin, Streptomycin. Single cell suspensions were washed in complete medium. Thy1+ and NK1.1+ cells were eliminated using AT83 and PK136 Abs, respectively, and rabbit complement (Saxon Europe, Suffolk, UK). 5×106 cells from each donor were then injected i.v. into γ-irradiated hosts (850 Rad, 137 Cs source, 630 Rad/minute) that were kept on antibiotic-containing water (0.28% pediatric suspension of Bactrim, Roche, Basel, Switzerland) for the complete duration of the experiment. Effector and/or regulatory T cells were co-injected with bone-marrow preparations.
FACS analysis
For analysis of bone-marrow from femurs and tibias, mice were sacrificed 15 days after bone-marrow and T cell transfer. For PBMC-analysis, blood samples were taken at indicated timepoints. Cells were labeled with anti-H-2Kb-FITC and anti-H-2Kd-PE or with anti-H-2Kk-FITC and anti-H-2Kb-PE (BD Pharmingen; eBioscience). The relative percentages of semiallogeneic cells shown in the figures are calculated as follows: (% of semiallogeneic cells in experimental mouse ÷ mean % of semiallogeneic cells in mice injected with bone-marrow only) × 100.
Results
Ex vivo expanded CD4+CD25+ regulatory T lymphocytes inhibit CD4+ and CD8+ T cell-mediated rejection of allogeneic bone-marrow
We have adapted our previously described mouse model for bone-marrow transplantation in which B6 (H-2b) hosts are lethally irradiated and reconstituted with a 1/1 mixture of syngeneic (B6) and semi-allogeneic B6D2F1 (H-2bd) bone marrow 4,8. Equivalent percentages of syngeneic and allogeneic cells were found in the bone-marrow two weeks post-transfer (Fig. 1A). When host-type CD4+ T cells were co-injected, the semi-allogeneic bone-marrow was eliminated and the retained syngeneic bone marrow allowed for survival of the host (Fig. 1B). Since we have been unable to induce tolerance to semi-allogeneic bone-marrow with freshly isolated regulatory T cells (data not shown), we evaluated the regulatory potential of CD4+CD25+ T cells activated and expanded in vitro. Purified B6 CD4+CD25+ splenic T cells were cultured in vitro with B6D2F1 APC in presence of IL-2. In 2-week cultures a 10 to 20-fold expansion was reproducibly obtained (data not shown). When such in vitro-cultured regulatory T cells were co-injected with B6 CD4+ T lymphocytes, rejection of semi-allogeneic bone-marrow was inhibited very efficiently, even at a ratio of one regulatory to ten CD4+ effector T cells (Figs. 1B and C).
Figure 1. CD4+CD25+ regulatory T cells preactivated in vitro induce CD4+ and CD8+ T cell-tolerance to semiallogeneic bone-marrow.
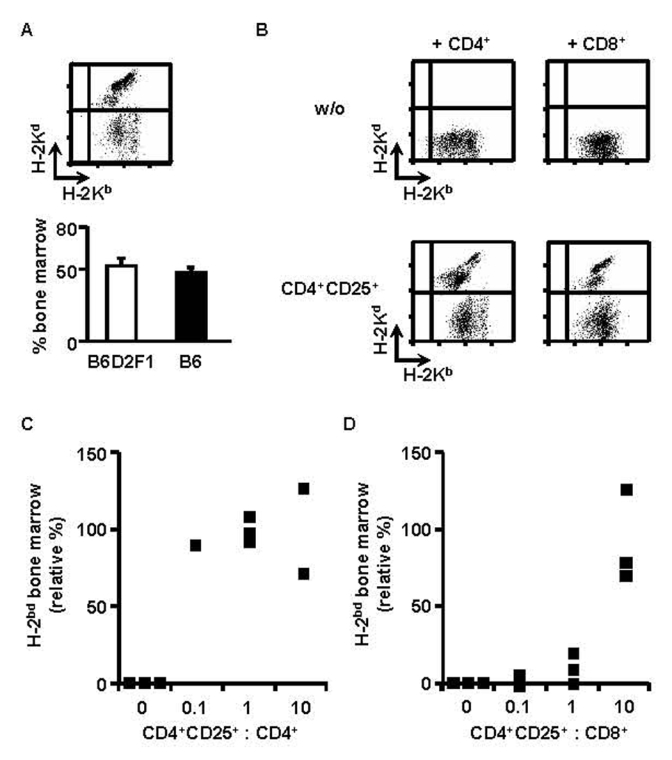
B6 mice were lethally irradiated and grafted with a 1/1 mixture of B6 (H-2b) and B6D2F1 (H-2bd) bone-marrow (5×106 cells each). Two weeks later equivalent percentages of syngeneic and semi-allogeneic bone-marrow were observed in bone marrow (A). Irradiated mice were co-injected with the two types of bone-marrow and 3×105 CD4+ (B left, C) or CD8+ T cells (B right, D) with (B bottom) or without (B top) CD4+CD25+ regulatory T cells from B6 mice in vitro preactivated with B6D2F1 APC (B, regulatory to effector T cell ratio = 10, C, D, as indicated). Mice were analyzed two weeks later. (A top, B) FACS analysis of bone-marrow, (C)(D) Relative percentage (see Material and Methods) of semiallogeneic bone marrow. Squares indicate individual mice from a representative experiment of 4 performed.
Rejection of allogeneic grafts is also mediated by CD8+ T lymphocytes. It was therefore important to assess if regulatory T cells could inhibit CD8+ T cells in vivo. Lethally irradiated B6 mice were reconstituted with a mixture of syngeneic and semi-allogeneic bone-marrow cells. They were simultaneously injected with purified CD8+ T lymphocytes with or without preactivated regulatory T cells. The data shown in figures 1B and 1D indicate that CD8+ T lymphocytes fully rejected semi-allogeneic bone-marrow and that co-injection of in vitro expanded regulatory T cells (at a ratio of 10 CD4+CD25+ to 1 CD8+ T cell) could inhibit this rejection. The higher ratios of regulatory T lymphocytes required to inhibit graft-rejection by CD8+ (as compared to CD4+) T cells are probably due to the much higher efficiency of CD8+ cells in graft-rejection in our system (data not shown). In conclusion, CD4+CD25+ regulatory T lymphocytes are capable of inhibiting in vivo alloreactivity of CD4+ as well as of CD8+ T cells.
Regulatory T cells induce durable tolerance of total splenocytes to allogeneic bone-marrow
We then assessed if regulatory T lymphocytes could also inhibit semi-allogeneic bone-marrow rejection by total splenocytes. Lethally irradiated B6 hosts were reconstituted with a mixture of syngeneic and semi-allogeneic bone marrow cells, injected with B6 splenocytes, and co-injected (or not) with titrated numbers of B6 CD4+CD25+ T lymphocytes that had been preactivated with B6D2F1 APC. As shown in figures 2A and B, co-injected regulatory T cells inhibited semi-allogeneic bone-marrow rejection in a dose-dependent fashion. Already at a CD4+CD25+ regulatory T cell to splenocyte ration of 1/2 significant protection of semi-allogeneic bone-marrow was observed.
Figure 2. Preactivated CD4+CD25+ regulatory T cells durably prevent semiallogeneic bone-marrow rejection by total splenocytes.
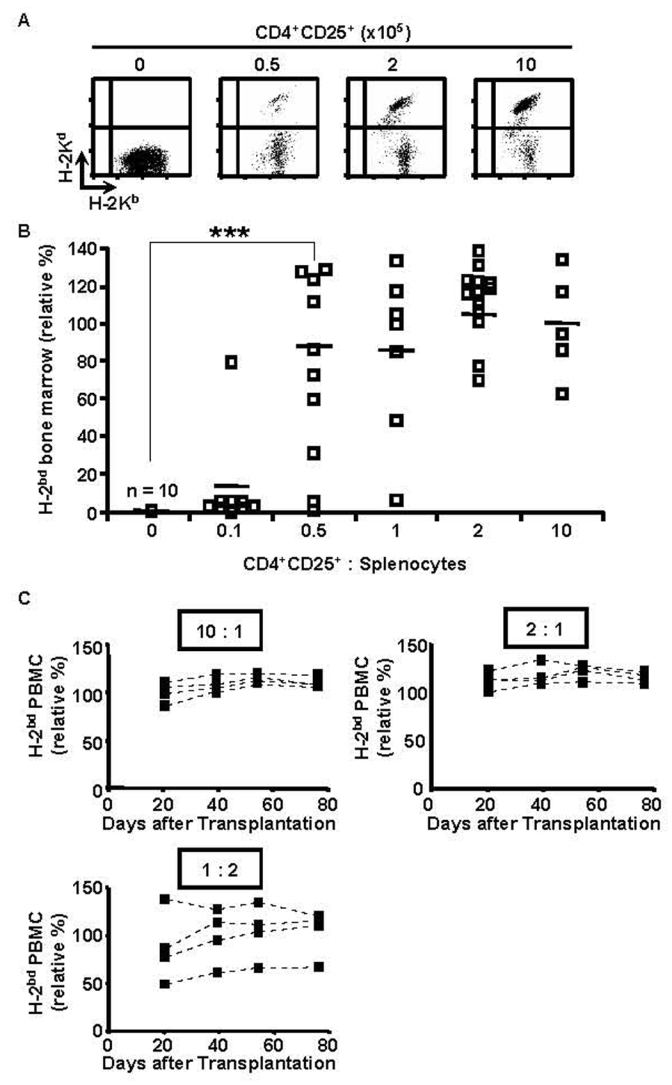
B6 mice were lethally irradiated and grafted with a 1/1 mixture of B6 (H-2b) and B6D2F1 (H-2bd) bone-marrow (5×106 cells each) and co-injected with 105 B6 splenocytes and regulatory T cells preactivated with B6D2F1 APC in vitro (regulatory to effector T cell ratio as indicated). Bone marrow was analyzed 2 weeks later (A, B, combined data from 4 independent experiments), PBMC at indicated timepoints (C, representative individual mice). (A) FACS analysis of bone-marrow, (B) (C) Relative percentage of semiallogeneic cells (see Material and Methods). *** p<0.001 (Student’s t-test).
In these experiments, bone-marrow engraftment was analyzed after 2 weeks. To assess if the tolerance was durable, we reconstituted lethally irradiated B6 mice with syngeneic and semi-allogeneic bone-marrow, co-injected them with B6 splenocytes and preactivated B6 regulatory T cells, and analyzed peripheral blood mononuclear cells from 1 to 11 weeks post-reconstitution. Figure 2C shows that the tolerance was durable: up to 11 weeks after transfer no signs of rejection were observed at regulatory T cell/splenocyte ratios of 2 and 10. At a regulatory T cell/splenocyte ratio of 1:2 levels of protection were quite variable among the four animals analyzed. Interestingly, once established, the percentage of allogeneic cells in PBMC did not significantly change during the period in which the animals were analyzed. Therefore, even partial protection appears to be stable in time. We are currently investigating the underlying mechanisms.
Pre-activated regulatory T cells protect allogeneic bone-marrow in an MHC-haplotype specific manner
We next evaluated if the protection of allogeneic grafts by activated regulatory T cells was specific. B6 CD4+CD25+ T cells were pre-activated in vitro with B6D2F1 (H-2bd) APC in presence of IL-2. These cells were then co-injected with B6 splenocytes, B6 bone marrow, and B6D2F1 (H-2bd) or B6CBAF1 (H-2bk) bone-marrow, into lethally irradiated B6 hosts. The results in figure 3A show that B6D2F1 bone marrow was more efficiently protected than B6CBAF1 cells, most notably at lower regulatory T cell to splenocyte ratios.
Figure 3. Prevention of semiallogeneic bone-marrow rejection mediated by CD4+CD25+ regulatory T cells is MHC-haplotype specific.
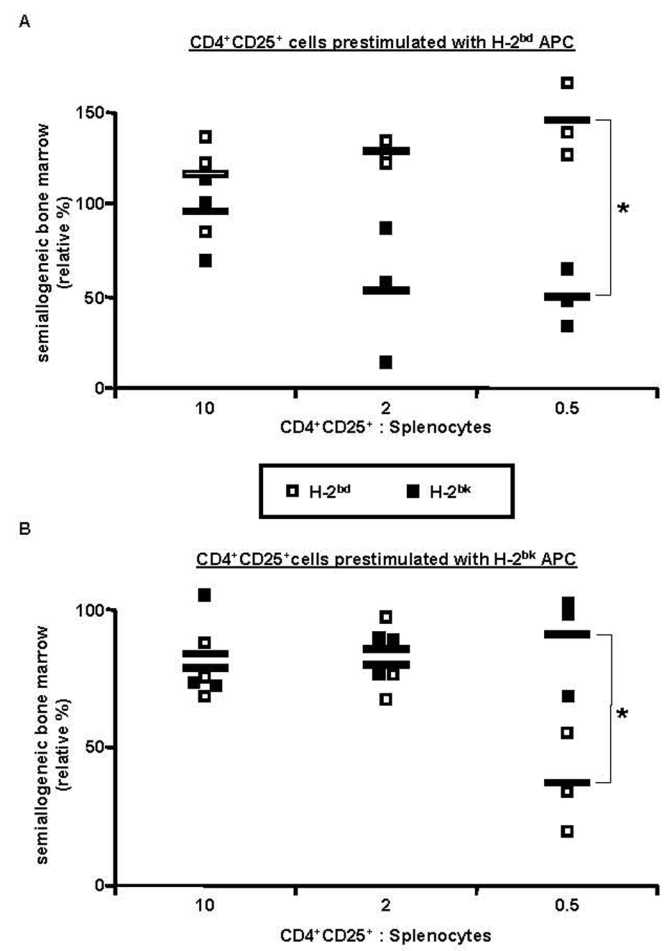
B6 mice were lethally irradiated, grafted with a 1/1 mixture of B6 (H-2b) plus B6D2F1 (H-2bd) or B6CBAF1 (H-2bk) bone-marrow (5×106 cells each), and co-injected with 105 B6 splenocytes and with regulatory T cells from B6 mice preactivated with B6D2F1 (A) or B6CBAF1 (B) APC in vitro (regulatory to effector T cell ratios as indicated). Bone marrow was analyzed 2 weeks later. Relative percentage of semiallogeneic H-2bd (□) or H-2bk (■) cells in the individual mice is indicated (see Material and Methods). * p< 0.05 (Student’s t-test). Combined data from 3 independent experiments are shown.
Since this specific protection might hypothetically be due to differences in the kinetics and/or potency of rejection of the two types of bone-marrow, we also performed the reciprocal experiment and tested if B6 regulatory T cells activated in vitro with B6CBAF1 APC acted in a specific manner (Fig. 3B). At a regulatory T cell to splenocyte ratio of 0.5, B6CBAF1 bone marrow was more efficiently protected than B6D2F1 cells. When using higher regulatory T cell to splenocyte ratios the specificity was lost.
Regulatory T cells protect target bone marrow while allowing third party alloreactivity to develop simultaneously
Since the observed specificity in the protection of allogeneic bone-marrow might be due to several factors other than specificity in the effector function of regulatory T cells (e.g. differential survival, homeostatic expansion, or activation in the distinct experimental mice), we then tested if the specificity was maintained when both types of semi-allogeneic bone marrow (i.e. B6D2F1 and B6CBAF1) were transferred (together with syngeneic bone-marrow) into the same host. When the three types of bone-marrow were injected without effector T cells, practically equivalent percentages of syngeneic (B6) and the two types of allogeneic (B6D2F1 and B6CBAF1) cells were found in bone marrow two weeks later (Fig. 4A). As shown in figures 4B (top) and 4C, addition of syngeneic effector splenocytes resulted in the clearance of both allogeneic populations, and regulatory T cells preactivated with B6D2F1 APC preferentially inhibited rejection of B6D2F1 (as compared to B6CBAF1) bone marrow, especially at lower regulatory to effector T cell ratios. At higher regulatory to effector T cell ratios, specificity was gradually lost, and cells of both semi-allogeneic origins were efficiently preserved.
Figure 4. Simultaneous protection of target bone marrow and rejection of third-party cells.

B6 mice were lethally irradiated, grafted with a 1/1/1 mixture of B6 (H-2b), B6D2F1 (H-2bd), and B6CBAF1 (H-2bk) bone-marrow (5×106 cells each), and co-injected with 105 B6 splenocytes and regulatory T cells preactivated with B6D2F1 or B6CBAF1 APC in vitro (as indicated, regulatory T cell to splenocyte ratios as indicated). Bone marrow was analyzed 2 weeks later. (A) Percentages of the three types of bone marrow recovered from control mice (i.e. mice that had not received splenocytes). (B) H-2Kd and H-2Kk expression by bone-marrow cells recovered from individual mice that had received bone-marrow, splenocytes, and titrated numbers of preactivated regulatory T cells, as indicated. (C) Summary of all data depicted as ratios of relative percentages of H-2bk to H-2bd bone-marrow cells, (□) Mice injected with regulatory T cells preactivated with H-2bd APC, (■) Mice injected with regulatory T cells activated with H-2bk APC. ** p< 0.01 (Wilcoxon test). Combined data from 3 independent experiments are shown.
Again, to exclude the hypothetical possibility that the observed specificity was due to differences in the kinetics and/or potency of rejection of the two types of bone-marrow, we then performed the reciprocal experiment. Regulatory T cells preactivated with B6CBAF1 cells preferentially protected B6CBAF1 (as compared to B6D2F1) bone marrow, again most notably at lower regulatory to effector T cell ratios (Figs. 4B (bottom) and C). These results definitively show that preactivated regulatory T cells can act in an antigen specific manner during their effector function in vivo.
Discussion
Like all other T lymphocytes, CD4+CD25+ regulatory T cells are antigen-specific and are thought to act in an antigen-specific manner in vivo. In vitro, however, antigen-specificity of these cells is limited to the activation phase while the effector phase is completely antigen-non-specific. While in several in vivo models regulatory T cells have been shown to act in an antigen-specific manner, in none of these systems has a distinction been made between antigen-specific activation and antigen-specific suppressor effector function.
Using a novel experimental system in which allogeneic bone marrow is durably protected from rejection by in vitro activated regulatory T cells, we show here that regulatory T cells can act in an antigen-specific manner during their effector phase in vivo. Our results suggest that CD4+CD25+ regulatory T cells could become a powerful tool to induce specific tolerance to allogeneic grafts in clinical settings.
CD4+CD25+ regulatory T cells have been used in a variety of systems to inhibit immunopathology. All these models have in common that injection of T lymphocytes must be followed by homeostatic proliferation. In the colitis model, disease is induced in lymphopenic mice 11. GvHD models rely on lethally irradiated recipients, which are therefore lymphopenic 19–21,38. The induction of transplantation-tolerance (GvHD or graft-rejection) with experimentally induced CD4+CD25+ regulatory T cells has also been studied in lymphopenic mice 23,25–29. In the experimental models of autoimmunity induced by day 3 thymectomy or in nude mice, homeostatic expansion of injected T lymphocytes also certainly occurs 10. Since regulatory T cells are known to regulate homeostatic expansion and to expand themselves in immunodeficient mice 39,40, it has been suggested that regulatory T cells could outcompete pathogenic T cells during the expansion-phase and thus inhibit immunopathology 41. Such a scenario could also explain the regulatory T cell-induced tolerance we observed when mice were injected with only one type of allogeneic bone-marrow (target or third party). In experiments in which the mice were co-injected with two types of allogeneic bone-marrow, however, target bone marrow was preferentially protected (notably at low regulatory to effector T cell ratios). Therefore, in contrast to the abovementioned reports, in our system tolerance cannot be explained by differential homeostatic expansion of alloreactive versus innocuous regulatory T cells.
T-cell tolerance induced by CD4+CD25+ regulatory T lymphocytes has never before been shown to be antigen-specific in the effector phase. In GvHD and transplantation models, tolerance was studied in separate hosts 21,22,27,29. Since homeostatic proliferation of regulatory T cells depends upon MHC class II expression 40 and potentially even upon interaction with specific antigen 33, antigen-specificity in vivo may be due to lack of homeostatic proliferation in animals lacking the “target” tissue. In these models, antigen-specificity is probably also due to the absence of specific ligands capable of activating the suppressor-effector function of regulatory T-lymphocytes. These hypothetical explanations may also apply to the specific tolerance we observed when target and third party bone marrows were injected (together with splenocytes and regulatory T cells) into separate hosts. However, we also observed preferential protection of target bone marrow in mice in which third party cells were simultaneously rejected. One could argue that the specific protection was due to differences in the kinetics and/or potency of rejection of the two types of bone marrow but such objections can be ruled out since identical results were obtained in reciprocal experiments. Since target bone marrow was protected in these mice, regulatory T cells had clearly been activated, but protected third-party bone marrow much less efficiently. Therefore, our data directly show that CD4+CD25+ regulatory T cells can act in an antigen-specific manner during their effector phase.
Several reason(s) could explain why protection is not entirely antigen-specific, especially at higher regulatory to effector T cell ratios. Since this lack of specificity was also observed in mice in which target bone-marrow was absent, regulatory T cells are sufficiently cross-reactive with third-party antigens to cause partial protection. Such a cross-reactivity may in part be due to “indirect antigen presentation” by host-type MHC molecules that are expressed by both types of semi-allogeneic bone marrows. Direct cross-reactivity towards allogeneic MHC molecules most likely also plays an important role 3. In contrast to the C57BL/6 hosts we used, both donor strains (B6D2F1 and B6CBAF1) present several mouse mammary tumor virus-encoded endogenous superantigens 42. Therefore, superantigen-reactivity of injected regulatory T cells may cause very significant cross-reactivity. In the mice in which we injected both types of allogeneic bone-marrow a contribution of non antigen-specific “bystander” mechanisms formally cannot be excluded. In any case, our data directly indicate that immunosuppression mediated by CD4+CD25+ regulatory T cells can act in an antigen-specific manner, not only in the activation but very importantly also during the in vivo effector phase. It will be important to assess if the antigen-specific suppressor effector function of CD4+CD25+ T cells will allow for clearance of infection of donor-type cells (e.g. by viruses).
Allogeneic bone-marrow or hematopoietic stem cell transplantation is extensively used to correct hereditary defects such as primary immunodeficiencies, metabolic diseases, β-thalassemia major, and sickle cell anemia (for review see ref. 43). These protocols require very frequent transfusions and/or immunosuppression to obtain and maintain hematopoietic chimerism. Since immunosuppressive drugs do not act in an antigen specific manner, their use is associated with opportunistic infections and increased tumor-incidence. Patients suffering from the above-mentioned congenital diseases would benefit from the availability of specific immunosuppressive protocols. The results we have obtained with our experimental model indicate that specific immunosuppression can be achieved with CD4+CD25+ regulatory T lymphocytes. They should open the way to development of clinical approaches aimed at induction of specific and durable tolerance to bone-marrow and hematopoietic stem cell allografts. Moreover, regulatory T cells could potentially also be used to induce specific and durable immunological tolerance to organ-allografts, and we are currently investigating this intriguing possibility.
In conclusion, the data presented here indicate that regulatory T cells can induce antigen-specific immunosuppression. It will be interesting to assess how antigen-specific in vivo tolerance is achieved. The model we describe here may prove particularly useful in addressing this and other questions concerning the in vivo functioning of CD4+CD25+ regulatory T cells. Moreover, our results suggest the feasibility of development of clinical protocols in which CD4+CD25+ regulatory T lymphocytes would be used to induce specific tolerance to allogeneic hematopoietic stem cell or organ-grafts.
Acknowledgments
The authors would like to acknowledge the staff of the IFR 30 animal facility for excellent animal-care, Carine Chanut for expert technical assistance, Drs. Jean-Charles Guéry, Etienne Joly, and Abdelhadi Saoudi for critical reading of the manuscript, Drs. Ronald N. Germain and Guy Laurent for helpful discussions, and Dr. Clemens Utzny for advice on statistical analysis.
Research grants: This work was supported by grants from the “Etablissement Français des Greffes” and the “Région Midi-Pyrénées”.
Abbreviations
- GvHD
Graft-versus-host disease
- B6
C57BL/6
- B6CBAF1
(C57BL/6xCBA/J)Fi
- B6D2F1
(C57BL/6xDBA/2)F1
References
- 1.Wardemann H, Yurasov S, Schaefer A, Young JW, Meffre E, Nussenzweig MC. Predominant Autoantibody Production by Early Human B Cell Precursors. Science. 2003;301:1374–1377. doi: 10.1126/science.1086907. [DOI] [PubMed] [Google Scholar]
- 2.van Meerwijk JPM, Marguerat S, Lees RK, Germain RN, Fowlkes BJ, MacDonald HR. Quantitative impact of thymic clonal deletion on the T cell repertoire. J Exp Med. 1997;185:377–383. doi: 10.1084/jem.185.3.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zerrahn J, Held W, Raulet DH. The MHC reactivity of the T cell repertoire prior to positive and negative selection. Cell. 1997;88:627–636. doi: 10.1016/s0092-8674(00)81905-4. [DOI] [PubMed] [Google Scholar]
- 4.Capone M, Romagnoli P, Beermann F, MacDonald HR, van Meerwijk JPM. Dissociation of thymic positive and negative selection in transgenic mice expressing major histocompatibility complex class I molecules exclusively on thymic cortical epithelial cells. Blood. 2001;97:1336–1342. doi: 10.1182/blood.v97.5.1336. [DOI] [PubMed] [Google Scholar]
- 5.Laufer TM, DeKoning J, Markowitz JS, Lo D, Glimcher LH. Unopposed positive selection and autoreactivity in mice expressing class II MHC only on thymic cortex. Nature. 1996;383:81–85. doi: 10.1038/383081a0. [DOI] [PubMed] [Google Scholar]
- 6.Sprent J, Kosaka H, Gao E-K, Surh CD, Webb SR. Intrathymic and extrathymic tolerance in bone marrow chimeras. Immunol Rev. 1993;133:151–176. doi: 10.1111/j.1600-065x.1993.tb01515.x. [DOI] [PubMed] [Google Scholar]
- 7.van Meerwijk JPM, MacDonald HR. In vivo T lymphocyte tolerance in the absence of thymic clonal deletion mediated by haematopoietic cells. Blood. 1999;93:3856–3862. [PubMed] [Google Scholar]
- 8.Hudrisier D, Feau S, Bonnet V, Romagnoli P, van Meerwijk JPM. In vivo unresponsiveness of T lymphocyte induced by thymic medullary epithelium requires antigen presentation by radioresistant cells. Immunol. 2003;108:24–31. doi: 10.1046/j.1365-2567.2003.01546.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Stockinger B. T lymphocyte tolerance: from thymic deletion to peripheral control mechanisms. Adv Immunol. 1999;71:229–265. doi: 10.1016/s0065-2776(08)60404-6. [DOI] [PubMed] [Google Scholar]
- 10.Asano M, Toda M, Sakaguchi N, Sakaguchi S. Autoimmune disease as a consequence of developmental abnormality of a T cell subpopulation. J Exp Med. 1996;184:387–396. doi: 10.1084/jem.184.2.387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Singh B, Read S, Asseman C, et al. Control of intestinal inflammation by regulatory T cells. Immunol Rev. 2001;182:190–200. doi: 10.1034/j.1600-065x.2001.1820115.x. [DOI] [PubMed] [Google Scholar]
- 12.Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature. 2002;420:502–507. doi: 10.1038/nature01152. [DOI] [PubMed] [Google Scholar]
- 13.Suvas S, Kumaraguru U, Pack CD, Lee S, Rouse BT. CD4+CD25+ T Cells Regulate Virus-specific Primary and Memory CD8+ T Cell Responses. J Exp Med. 2003;198:889–901. doi: 10.1084/jem.20030171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Golgher D, Jones E, Powrie F, Elliott T, Gallimore A. Depletion of CD25+ regulatory cells uncovers immune responses to shared murine tumor rejection antigens. Eur J Immunol. 2002;32:3267–3275. doi: 10.1002/1521-4141(200211)32:11<3267::AID-IMMU3267>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- 15.Shimizu J, Yamazaki S, Sakaguchi S. Induction of tumor immunity by removing CD25+CD4+ T cells: A common basis between tumor immunity and autoimmunity. J Immunol. 1999;163:5211–5218. [PubMed] [Google Scholar]
- 16.Sakaguchi S. Regulatory T cells: key controllers of immunologic self-tolerance. Cell. 2000;101:455–458. doi: 10.1016/s0092-8674(00)80856-9. [DOI] [PubMed] [Google Scholar]
- 17.Shevach EM. Regulatory T cells in autoimmmunity. Annu Rev Immunol. 2000;18:423–449. doi: 10.1146/annurev.immunol.18.1.423. [DOI] [PubMed] [Google Scholar]
- 18.Bach JF. Regulatory T cells under scrutiny. Nat Rev Immunol. 2003;3:189–198. doi: 10.1038/nri1026. [DOI] [PubMed] [Google Scholar]
- 19.Taylor PA, Lees CJ, Blazar BR. The infusion of ex vivo activated and expanded CD4(+)CD25(+) immune regulatory cells inhibits graft-versus-host disease lethality. Blood. 2002;99:3493–3499. doi: 10.1182/blood.v99.10.3493. [DOI] [PubMed] [Google Scholar]
- 20.Hoffmann P, Ermann J, Edinger M, Fathman CG, Strober S. Donor-type CD4(+)CD25(+) Regulatory T Cells Suppress Lethal Acute Graft-Versus-Host Disease after Allogeneic Bone Marrow Transplantation. J Exp Med. 2002;196:389–399. doi: 10.1084/jem.20020399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cohen JL, Trenado A, Vasey D, Klatzmann D, Salomon BL. CD4(+)CD25(+) Immunoregulatory T Cells: New Therapeutics for Graft-Versus-Host Disease. J Exp Med. 2002;196:401–406. doi: 10.1084/jem.20020090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Trenado A, Charlotte F, Fisson S, et al. Recipient-type specific CD4+CD25+ regulatory T cells favor immune reconstitution and control graft-versus-host disease while maintaining graft-versus-leukemia. J Clin Invest. 2003;112:1688–1696. doi: 10.1172/JCI17702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Graca L, Thompson S, Lin CY, Adams E, Cobbold SP, Waldmann H. Both CD4(+)CD25(+) and CD4(+)CD25(−) regulatory cells mediate dominant transplantation tolerance. J Immunol. 2002;168:5558–5565. doi: 10.4049/jimmunol.168.11.5558. [DOI] [PubMed] [Google Scholar]
- 24.Trani J, Moore DJ, Jarrett BP, et al. CD25+ Immunoregulatory CD4 T Cells Mediate Acquired Central Transplantation Tolerance. J Immunol. 2003;170:279–286. doi: 10.4049/jimmunol.170.1.279. [DOI] [PubMed] [Google Scholar]
- 25.Hara M, Kingsley CI, Niimi M, et al. IL-10 Is Required for Regulatory T Cells to Mediate Tolerance to Alloantigens In Vivo. J Immunol. 2001;166:3789–3796. doi: 10.4049/jimmunol.166.6.3789. [DOI] [PubMed] [Google Scholar]
- 26.Taylor PA, Noelle RJ, Blazar BR. CD4+CD25+ Immune Regulatory Cells Are Required for Induction of Tolerance to Alloantigen via Costimulatory Blockade. J Exp Med. 2001;193:1311–1318. doi: 10.1084/jem.193.11.1311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kingsley CI, Karim M, Bushell AR, Wood KJ. CD25+CD4+ Regulatory T Cells Prevent Graft Rejection: CTLA-4- and IL-10-Dependent Immunoregulation of Alloresponses. J Immunol. 2002;168:1080–1086. doi: 10.4049/jimmunol.168.3.1080. [DOI] [PubMed] [Google Scholar]
- 28.Sanchez-Fueyo A, Weber M, Domenig C, Strom TB, Zheng XX. Tracking the Immunoregulatory Mechanisms Active During Allograft Tolerance. J Immunol. 2002;168:2274–2281. doi: 10.4049/jimmunol.168.5.2274. [DOI] [PubMed] [Google Scholar]
- 29.van Maurik A, Herber M, Wood KJ, Jones ND. Cutting edge: CD4+CD25+ alloantigen-specific immunoregulatory cells that can prevent CD8+ T cell-mediated graft-rejection: Implications for anti-CD 154 immunotherapy. J Immunol. 2002;169:5401–5404. doi: 10.4049/jimmunol.169.10.5401. [DOI] [PubMed] [Google Scholar]
- 30.Gregori S, Casorati M, Amuchastegui S, Smiroldo S, Davalli AM, Adorini L. Regulatory T Cells Induced by 1 {alpha},25-Dihydroxyvitamin D3 and Mycophenolate Mofetil Treatment Mediate Transplantation Tolerance. J Immunol. 2001;167:1945–1953. doi: 10.4049/jimmunol.167.4.1945. [DOI] [PubMed] [Google Scholar]
- 31.Walker MR, Kasprowicz DJ, Gersuk VH, et al. Induction of FoxP3 and acquisition of T regulatory activity by stimulated human CD4+CD25− T cells. J Clin Invest. 2003;112:1437–1443. doi: 10.1172/JCI19441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chen W, Jin W, Hardegen N, et al. Conversion of Peripheral CD4+CD25− Naive T Cells to CD4+CD25+ Regulatory T Cells by TGF-{beta} Induction of Transcription Factor Foxp3. J Exp Med. 2003;198:1875–1886. doi: 10.1084/jem.20030152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Fisson S, Darrasse-Jeze G, Litvinova E, et al. Continuous Activation of Autoreactive CD4+ CD25+ Regulatory T Cells in the Steady State. J Exp Med. 2003;198:737–746. doi: 10.1084/jem.20030686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Walker LSK, Chodos A, Eggena M, Dooms H, Abbas AK. Antigen-dependent Proliferation of CD4+ CD25+ Regulatory T Cells In Vivo. J Exp Med. 2003;198:249–258. doi: 10.1084/jem.20030315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Yamazaki S, Iyoda T, Tarbell K, et al. Direct Expansion of Functional CD25+ CD4+ Regulatory T Cells by Antigen-processing Dendritic Cells. J Exp Med. 2003;198:235–247. doi: 10.1084/jem.20030422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hori S, Haury M, Coutinho A, Demengeot J. Specificity requirements for selection and effector functions of CD4+CD25+ regulatory T cells in anti-myelin basic protein T cell receptor transgenic mice. Proc Natl Acad Sci USA. 2002;99:8213–8218. doi: 10.1073/pnas.122224799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Thornton AM, Shevach EM. Suppressor effector function of CD4+CD25+ immunoregulatory T cells is antigen nonspecific. J Immunol. 2000;164:183–190. doi: 10.4049/jimmunol.164.1.183. [DOI] [PubMed] [Google Scholar]
- 38.Edinger M, Hoffmann P, Ermann J, et al. CD4(+)CD25(+) regulatory T cells preserve graft-versus-tumor activity while inhibiting graft-versus-host disease after bone marrow transplantation. Nat Med. 2003;17:17. doi: 10.1038/nm915. [DOI] [PubMed] [Google Scholar]
- 39.Annacker O, Pimenta-Araujo R, Burlen-Defranoux O, Barbosa TC, Cumano A, Bandeira A. CD25+ CD4+ T cells regulate the expansion of peripheral CD4 T cells through the production of IL-10. J Immunol. 2001;166:3008–3018. doi: 10.4049/jimmunol.166.5.3008. [DOI] [PubMed] [Google Scholar]
- 40.Gavin MA, Clarke SR, Negrou E, Gallegos A, Rudensky A. Homeostasis and anergy of CD4+CD25+ suppressor T cells in vivo. Nat Immunol. 2001;3:33–41. doi: 10.1038/ni743. [DOI] [PubMed] [Google Scholar]
- 41.Barthlott T, Kassiotis G, Stockinger B. T Cell Regulation as a Side Effect of Homeostasis and Competition. J Exp Med. 2003;197:451–460. doi: 10.1084/jem.20021387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Luther SA, Acha-Orbea H. Mouse mammary tumor virus: immunological interplays between virus and host. Adv Immunol. 1997;65:139–243. [PubMed] [Google Scholar]
- 43.Sullivan KM, Parkman R, Walters MC. Bone Marrow Transplantation for Non-Malignant Disease. Hematology. 2000:319–338. doi: 10.1182/asheducation-2000.1.319. [DOI] [PubMed] [Google Scholar]