

Abstract
In this study of reading development, children (ages 7–10) and adults (ages 18–32) performed overt single-word reading and aural repetition tasks on high-frequency word stimuli during functional magnetic resonance imaging. Most regions showed similar activity across age groups. These widespread regions of similarity indicate that children and adults use largely overlapping mechanisms when processing high-frequency words. Significant task-related differences included greater activity in occipital cortex for the read task, and greater activity in temporal cortex for the repeat task; activity levels in these regions were similar for adults and children. However, age group differences were found in several posterior regions, including a set of regions implicated in adult reading: the left supramarginal gyrus, the left angular gyrus, and bilateral anterior extrastriate cortex. The angular and supramarginal gyrus regions, hypothesized to play a role in phonology, showed decreased activity in adults relative to children for high-frequency words. The extrastriate regions had significant activity for both the visual read task and auditory repeat task in children, but just for the read task in adults, showing significant task and age interactions. These results are consistent with decreasing reliance on phonological processing, and increasing tuning of visual mechanisms, with age.
Keywords: angular gyrus, child, extrastriate cortex, specialization, supramarginal gyrus
Introduction
Mastery of the skill of reading is an important predictor of an individual's success in literate societies. This reliance upon an ability that is uniquely human, and that requires instruction for expert acquisition, has generated considerable research interest in its neurobiological foundation and developmental trajectory. Reading-related brain research has been conducted using many different tools, including behavioral methods, event-related potentials (ERP), magnetoencephalography (MEG), and neuroimaging methods (e.g., functional magnetic resonance imaging [fMRI] and positron emission tomography [PET]) (for an overall review, see Schlaggar and McCandliss 2007). From this wide variety of investigations come several consistent regions of activation. In general, it is agreed that adults show greater left than right hemisphere increases in activity for reading in many brain regions, including parts of inferior occipital–temporal cortex, inferior frontal gyrus/frontal operculum, and superior temporal and inferior-parietal cortex (for an fMRI/PET review, see Fiez and Petersen 1998; Turkeltaub et al. 2002; Jobard et al. 2003; Palmer et al. 2004; Binder et al. 2005; Bolger et al. 2005; for an ERP review, see Barber and Kutas 2007). MEG has also demonstrated a similar set of locations for reading-related tasks (Simos et al. 2002). Of note, some studies have demonstrated a less lateralized pattern of activity developmentally (Gaillard et al. 2000; Turkeltaub et al. 2003). Literature reporting the effect of damage to these 3 left hemisphere locations in the adult has indicated that separable reading-related deficits can result (Dejerine 1892; Geschwind 1965; Damasio and Damasio 1983; Binder and Mohr 1992; Hillis et al. 1999; Fiez et al. 2006).
In 1 of these 3 locations, the left inferior occipital–temporal gyrus, several recent adult studies have provided evidence for specialization in processing visual words (a “visual word form area” [vWFA]) (e.g., Cohen et al. 2002; Dehaene et al. 2002; McCandliss et al. 2003). The characterization of the vWFA is not without controversy, however (see e.g., Price and Devlin 2003; Devlin et al. 2006). This region, located approximately at Talairach coordinates −43, −54, −12, shows greater activity for words than consonant strings and may be visually specific in adults. Its location appears to be well-conserved across individuals (McCandliss et al. 2003). In addition, there is a consistent observation in the ERP literature of a negativity at the left posterior inferior aspect of the scalp (near the proposed vWFA) during the N170 time frame (first 200 ms) for presentation of visual words (Maurer et al. 2005). This negativity has been shown to be absent in nonreaders, largest in early readers, and to decline in magnitude with age (Brem et al. 2006; Maurer et al. 2006).
Since 1861, when Broca reported the consequences of a lesion in the left inferior frontal cortex (Broca's Area), that region has been implicated in fluent language production (Broca 1861). Recent studies have indicated that reading pseudowords, as compared with real words, produces greater activity in a region in the left frontal operculum (Fiez et al. 1999; Hagoort et al. 1999; Mechelli et al. 2003). Damage to this region (located approximately at Talairach coordinates −45, +12, +13) results in difficulty reading pseudowords and inconsistent low frequency words (Fiez et al. 2006).
Studies of typical adult readers and those with developmental dyslexia indicate that phonological and/or semantic processing might also be occurring in temporo-parietal regions including the superior temporal gyrus, the supramarginal gyrus, and the angular gyrus (Fiez and Petersen 1998; Jobard et al. 2003; Palmer et al. 2004). Regions in these locations are consistently reported in imaging studies of reading, although interpretations differ for the processes they perform (e.g., Pugh et al. 2001; Joubert et al. 2004). For example, Binder et al. (2005) have proposed that the left angular gyrus is a site of semantic processing similar in function to the semantic component of a connectionist model of reading (Plaut et al. 1996; Seidenberg 2005). This idea is based on a correlation between word imageability and blood oxygenation level–dependent (BOLD) activity in this region. However, Booth et al. (2004) do not support a role for semantics in the angular gyrus, and instead propose that the angular gyrus extracts statistical regularities between orthography and phonology, more consistent with earlier lesion studies (Geschwind 1965).
Overall, the idea that damage within the left temporo-parietal region results in language and/or reading disorders has been supported for over a century (in the temporal lobe by Wernicke (1874), and in the inferior-parietal lobe by Dejerine (1891)). More recently, Hillis et al. (2005) studied 80 patients within 24 h of onset of acute left hemisphere ischemic stroke, and compared the location of the stroke with the patient's abilities in word naming, oral reading, and reading comprehension, as well as other tasks. In addition to other results, damage to the angular gyrus was associated with impaired naming and oral reading. Moreover, adult and child dyslexia research suggests that impairment in left superior temporal and inferior-parietal regions is fundamental to the disorder, resulting in a failure to develop successful reading skills (e.g., Eden and Zeffiro 1998; Horwitz et al. 1998; Shaywitz et al. 2002; Hoeft et al. 2006).
Review of Developmental Studies
Although the literature of developmental studies of brain mechanisms used while reading is less extensive than for adult studies, the literature has grown with the advent of noninvasive brain research techniques. For example, researchers using MEG have revealed developmental differences in the timing of activation in some key reading regions between adults and children for lexical tasks that varied difficulty by age (Simos et al. 2001). Using fMRI, Schlaggar et al. (2002) studied children and adults performing lexical processing tasks (i.e., rhyme, verb, and opposite generation). In this study, regions were found in frontal cortex that showed greater activity for adults than children, whereas extrastriate regions showed greater activity for children than adults (see also Brown et al. 2005). Similarly, Turkeltaub et al. (2003) found that children performing a covert reading task showed a greater reliance on posterior superior temporal cortical regions than adults, and that an increase in frontal activity correlated with an increase in reading skill. Booth et al. (2004) reported an increase in activity in regions in the angular and supramarginal gyri in adults compared with children for spelling and rhyming cross-modal tasks, and suggest that this increase is due to adults having more elaborated mapping between orthographic and phonologic representations. None of these studies directly addressed overt single-word reading, but the results imply that the reading networks in the brain are dynamic between the ages of 7 years and adulthood, and suggest that targeted investigations of single-word reading during development could help clarify apparently discrepant findings. To this end, we studied adults and children on simple single-word reading and aural single-word repetition tasks.
Materials and Methods
Participants
Subjects were chosen from a data set of 96 healthy right-handed, native English speakers, ages 7–35 years who had participated in a large cross-sectional study of lexical processing. Some portions of this data set, describing different tasks, have been published previously (Schlaggar et al. 2002; Brown et al. 2005; Fair et al. 2006a; Fair et al. 2006b). For this analysis, the data from 50 of these participants were selected from the data set on the basis of behavioral measures (see below).
Subjects under 18 years had parental informed consent and gave assent. They were recruited from community schools and through parents connected to the Washington University community. Children and teenagers were screened similarly to the adults, but also were examined by a pediatric neurologist (B.L.S.). To characterize the children and teenagers further, participants under 18 were tested with Wechsler Abbreviated Scale of Intelligence (Wechsler 1999). Participants under 18 were also acclimated to the imaging environment via a “mock scanner” several days prior to the experiment.
All adults gave informed consent. Adult participants were college, graduate, or medical students, or college graduates, and many were from the Washington University community. Adult subjects were screened for neurological, psychiatric, and/or psychopathological diagnoses by telephone interview and questionnaire. Unfortunately, IQ testing was not obtained for these subjects. However, IQ scores were estimated for the adults based on their years of education, which provides a good general estimate (Matarazzo 1972).
The Washington University Human Studies Committee approved the study and all participants were reimbursed for their participation.
Lexical Tasks
All subjects were scanned while performing 2 simple lexical tasks: 1) Read (aloud) a visually presented word, and 2) Repeat (aloud) an aurally presented word. All subjects performed 1 or 2 runs of each task. In both tasks, participants viewed a white fixation cross on a black background using a rear projector and mirrors. Words for both tasks were high frequency, and derived from available lists of US first-grade reading words. Word lists were counterbalanced for syllables, and words were presented in random order. For visually presented words, each letter subtended 0.5°. Visual stimulus duration was relatively long (0.95 s), which was intentionally chosen to aid reading in the youngest subjects. Auditory stimulus duration varied appropriately with word length spoken by an adult male raised in the Midwestern United States. The means and ranges of the length, frequency, and orthographic neighbors of the stimuli are summarized in Table 1. Although there is a numerical difference between the frequencies of the word items, the read and repeat task stimuli were both quite high frequency on average, and were not significantly different, as determined by t-test (P = 0.44).
Table 1.
Stimulus properties of the Read and Repeat tasks
| Read | Repeat | |
| Avg. length (letters) | 4.4 | 4.3 |
| Range | 2–9 | 2–8 |
| Avg. frequency (HAL) | 369, 361 | 447, 594 |
| Range | 2,682–3,095,437 | 1,614–6,474,135 |
| Avg. ortho. neighbors | 9.8 | 9.6 |
| Range | 0–29 | 0–28 |
In each run, 21 words were presented every second, third, or fourth MR frame (time repetition [TR] 3.08 s; average interstimulus interval 9.24 s) in pseudorandom fashion. This jittering allowed the event-related time course of the response to be extracted (Miezin et al. 2000). For each task, participants were asked to say aloud the correct response as quickly, clearly, and accurately as possible.
Behavioral Measures
Behavioral data were collected with digital voice recording software for later analysis as described by Nelles et al. (2003). Reaction time and accuracy (% correct) were computed for each individual.
In any study of development, researchers have to address a potential confound due to differing task performance between age groups (Johnson 2001; Schlaggar et al. 2002; Palmer et al. 2004). We used performance matching of behavioral measures (reaction time and accuracy) to select our sample from a larger data set. First, participants without at least one run of each task, with less than 70% accuracy on either task, or with excessive movement (greater than 1.5 mm root mean square [rms]), were removed from analysis. The remaining set of participants was subjected to individual removals to balance the average reaction time and accuracy for the set of adults and children aged 7–10 years. The final, matched set of data contained the endpoints of our developmental span of 7–35 years, and consisted of 25 adults (18–35 years, 14 female), and 25 children (7–10 years, 16 female). The characteristics of the larger child and adult data set and of the matched child and adult groups used for this analysis can be seen in Table 2.
Table 2.
Demographic and task performance characteristics of the child and adult groups
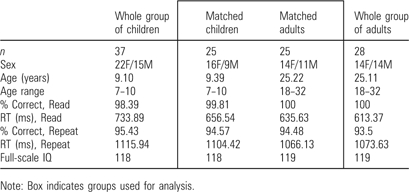 |
Note: Box indicates groups used for analysis.
MRI Data acquisition and Preprocessing
A Siemens 1.5 Tesla MAGNETOM Vision system (Erlangen, Germany) was used to collect the anatomical and functional MRI data. In brief, 3-dimensional high-resolution structural images were acquired using a sagittal magnetization-prepared rapid gradient echo (MP–RAGE) sequence (slice time echo = 4 ms, TR = 9.7 ms, inversion time = 300 ms, flip angle = 12°, 128 slices, 1.25 × 1 × 1 mm voxels). Functional data were collected parallel to the anterior commissure–posterior commissure plane using an asymmetric spin-echo echo-planar pulse sequence (TR = 2.18 s with a 904 ms delay [total TR = 3.08 s], T2* evolution time = 50 ms, flip angle = 90°). Complete brain coverage was achieved during each scan by obtaining 73 frames of 16 contiguous interleaved 8-mm axial slices (3.75 × 3.75 mm in-plane resolution). The brief delay after each TR was added to each frame to minimize scanner noise and allow subject task responses to be more easily monitored. Steady state was assumed starting with the fourth frame.
Preliminary image processing was carried out to reduce noise and artifacts (see Miezin et al. 2000, for detailed procedures). Preprocessing included removal of a single pixel spike caused by signal offset, whole-brain normalization of signal intensity across MR frames, correction for subject movement within and across runs, and slice-by-slice normalization to correct for changes in signal intensity due to acquisition of interleaved slices.
Data for all subjects were transformed into standard stereotactic space (based on Talairach and Tournoux 1988) using a single common atlas (Lancaster et al. 1995; Snyder 1996) and resampled isotropically at 2 mm × 2 mm × 2 mm. Stereotactic registration was accomplished by 12-parameter affine warping of each individual MP–RAGE image to an atlas-representative target using difference image variance minimization as the objective function. The atlas-representative target image was produced by mutual coregistration of independent groups of 12 healthy adults and 12 healthy children. This target-image strategy minimizes systematic age differences in the atlas location of cortical structures (see Burgund et al. 2002). The composite target was made to conform to the Talairach atlas using the spatial normalization method of Lancaster et al. (1995). The atlas-transformed image for each of our participants was checked against a reference average to ensure appropriate registration. This method allows direct voxel-wise statistical comparisons to be made between children and adults, and has been used previously (Burgund et al. 2002; Schlaggar et al. 2002; Kang et al. 2003; Monk et al. 2003; Blumenfeld et al. 2006; Ciesielski et al. 2006).
Participants were encouraged to minimize movements throughout the scan, including during their vocal responses. In order to enhance participants’ abilities to remain still, a thermoplastic mask that molds to an individual's face was applied. Participant motion was corrected and quantified using an analysis of head position based on rigid body translation and rotation. The data derived from adjustments needed to realign head movement on a frame-by-frame basis were calculated as rms values for translation and rotation in the x, y, and z planes in millimeters. A scan run was discarded if the movement was more than 1.5 mm rms. Movement rms values ranged from 0.114 to 1.298, with an average overall movement of 0.379 rms across both tasks for all subjects. As has been reported elsewhere, children moved significantly more than adults (Schlaggar et al. 2002; Brown et al. 2005).
Concern about artifacts from scanning during overt verbal responses in the MRI environment has been addressed by multiple researchers (Birn et al. 1999; Palmer et al. 2001). These papers demonstrate that acquisition of interpretable, relatively artifact-free images of tasks during overt verbal responses is possible, and other studies have also not observed large speaking-related artifacts (Schlaggar et al. 2002; Binder et al. 2005; Brown et al. 2005; Fair et al. 2006b).
fMRI Processing and Data Analysis
General Features
Statistical analyses of event-related fMRI data were based on the general linear model (GLM) as previously described (Miezin et al. 2000; Schlaggar et al. 2002; Brown et al. 2005), and conducted using in-house software programmed in the Interactive Data Language (Research Systems, Inc., Boulder, CO).
The GLM design for each subject included time as a 7-level factor, made up of the 7 MR frames (∼22 s, 3.08 s per frame) following presentation of the stimulus. No assumptions were made regarding the shape of the hemodynamic response function. All analyses included only activity corresponding to correct trials.
Voxel-Wise Analysis
The first analysis, a 2 × 2 × 7 voxel-wise, whole-brain analysis of variance (ANOVA), was constructed with 2 levels of task (read and repeat), 2 levels of age (child and adult), and the 7 points in time (7 MR frames) for all correct trials. Error trials were coded separately from correct trials and not analyzed for this study. For all voxel-wise analyses, a correction based on a Monte Carlo simulation was implemented to guard against false positives that may result from conducting a large number of statistical comparisons over many images (Forman et al. 1995; McAvoy et al. 2001). To achieve P < 0.05 corrected for multiple comparisons, a threshold of 24 contiguous voxels with a Z value > 3.5 was applied.
The voxel-wise analysis produced 4 images, each showing voxels with either a significant effect of time, time × age interaction, time × task interaction, or a time × task × age interaction. From these images, regions were created using an in-house algorithm (courtesy of Avi Snyder) that located peaks within the Monte Carlo corrected contiguous voxels in each image, and defined regions by first smoothing with a 4-mm kernel, then extracting peaks with a minimum distance of 10 mm from other peaks, a peak Z-value of at least 3.5, and a minimum size of 24 voxels. For all region-wise analyses, Box's sphericity correction was used, adjusting for temporal correlation and possible inhomogeneity of variance over the repeated measure (i.e., time) (Box 1954; McAvoy et al. 2001).
Region of Interest Analyses
Regions extracted from the 4 images were examined for overlap, and whenever a center of mass coordinate for a region was within 5 mm of another region, the smaller of the 2 was removed. The regions derived from the voxel-wise images were then interrogated for the combinations of effects that were found. No regions had significantly negative-going time courses. The regions derived from a particular interaction image (e.g., regions of interest [ROIs] derived from the task × time image) are already known to be significant for that interaction (e.g., the task × time effect), so a statistic for that effect in those ROIs cannot be considered a significance test, and is only useful as an index of relative reliability. On the other hand, a test for task × time interaction in an ROI defined in the main effect of time image provides an appropriate unbiased test.
In many regions, the only statistically significant effect matched the effect in the original voxel-wise image. However, for some regions interactions were found that were not reflected in the image of origin. For example, of the 63 regions derived from the main effect of time image, 22 regions showed at least one secondary interaction effect. These regions were then grouped with the other regions showing those effects for further analyses. The different classes of statistical measurement are denoted in each result table by using an asterisk to indicate regions derived from the particular interaction effect image and thus not by a secondary statistical test. Post hoc analyses determined which factor(s) drove each interaction. Main effect of time statistics were considered significant if they were greater than a sphericity-corrected Z-value of 4.0 (P < 0.00005). Interaction effects were considered significant if they had a sphericity-corrected Z-value greater than 2.5 (P < 0.01).
Time courses for all 50 subjects were extracted for each region. The individual time courses across all regions were screened for highly aberrant values, blind to age. Outlier time courses were defined as those showing any time point with greater than 2% signal change, because 2% signal change represented 3 standard deviations away from the mean when 96 subjects from the larger data set were examined for signal magnitudes (Brown et al. 2005). Regions containing 10% or more outlying time courses across subjects were removed and are not reported here. Regions showing fewer than 10% outlying time courses across subjects were retained, but data from those particular “outlier” subjects were removed from analyses of that region only.
A subsidiary ROI analysis was performed on 3 left hemisphere regions whose location was similar to regions described in prior studies of reading (Cohen et al. 2002; Palmer et al. 2004; Binder et al. 2005). These regions of interest were located in the left angular gyrus (−49, −62, +29), supramarginal gyrus (−52, −42, +24), and anterior extrastriate cortex (−38, −60 −12).
Results
Behavioral Results
The behavioral data for the performance-matched groups are depicted in Table 2. As designed, the 2 groups did not differ by reaction time or accuracy. Average full-scale IQ for the 25 children was 118 and estimated IQ for the 25 adults was 119.
Children and adults were slower on the repeat task (child average 1104 ms, adult average 1066 ms) than the read task (child average 657 ms, adult average 636 ms) (P < 0.05 for task differences, no group difference). Both groups were highly accurate on both tasks (>94%), indicating that both groups were successful at performing the 2 tasks.
Voxel-Wise Analysis
The 2 × 2 × 7 ANOVA resulted in a preponderance of regions derived from the main effect of time image: 69 regions qualified for further analysis. Six of these regions were removed due to overlap with other regions (see Methods). The remaining 63 regions were located bilaterally in a wide variety of locations in the brain. These regions were analyzed for secondary effects (see Methods).
Other images showing more complex interactions provided 16 regions (task × time interaction image), and 25 regions (age × time interaction image). Five of these regions were removed due to overlap. Age × time interaction image regions were located only in the posterior half of the brain, and included regions in the precuneus, inferior-parietal, superior temporal, and occipital lobes.
ROI Analysis
Regions showing only a main effect of time potentially reveal the shared functional neuroanatomy of reading and repeating for people ages 7–35 years (Fig. 1). These regions were in a wide variety of locations, including visual cortex, auditory cortex, and primary motor cortex, as well as other regions commonly associated with reading tasks (Fiez et al. 1999; Price 2000; Joseph et al. 2001; Turkeltaub et al. 2002; Palmer et al. 2004) (Table 3).
Figure 1.

Regions showing no significant differences between ages or tasks. An example region, location indicated by the arrow, is shown at right, with percent signal change on the y-axis, and time (in MR frames; each MR frame is 3.08 s) on the x-axis. Time courses are shown with standard error of the mean. Coordinates reported for this and all figures are in Talairach space. The orientation of all subsequent brain surfaces will follow the convention above. All surface-rendered images were created using CARET software and surface-based atlases (Van Essen 2002; Van Essen et al. 2001).
Table 3.
Forty-one brain regions showing a main effect of time in children and adults
| x | y | z | Size (cm3) | Location | Time (Z-score) | Approximate Brodmann area |
| Left | ||||||
| −2 | −9 | 50 | 2.45 | Sup. frontal gyrus | 9.05 | 6 |
| −3 | 11 | 37 | 4.18 | Sup. frontal gyrus | 10.23 | 32 |
| −47 | −16 | 34 | 4.16 | Precentral gyrus | 13.84 | 4 |
| −44 | 2 | 28 | 3.25 | Inf. frontal gyrus | 8.80 | 44 |
| −34 | 27 | 17 | 0.27 | Inf. frontal gyrus | 7.26 | 45 |
| −29 | 43 | 25 | 0.35 | Mid. frontal gyrus | 6.92 | 9 |
| −2 | −81 | 17 | 3.90 | Cuneus | 9.61 | 18 |
| −9 | −76 | 5 | 3.46 | Lingual gyrus | 11.11 | 18 |
| −14 | −67 | 1 | 3.67 | Lingual gyrus | 9.51 | 18 |
| −26 | −76 | 21 | 1.74 | Mid. occipital gyrus | 7.29 | 18 |
| −20 | −34 | 55 | 1.77 | Sup. parietal lobe | 9.37 | 7 |
| −27 | −49 | 48 | 2.14 | Sup. parietal lobe | 8.10 | 7 |
| −52 | −45 | 8 | 4.02 | Mid. temporal gyrus | 10.40 | 21 |
| −51 | 4 | 0 | 3.60 | Sup. temporal gyrus | 10.39 | 22 |
| −39 | 3 | 12 | 3.26 | Insula | 8.80 | — |
| −40 | 18 | 9 | 2.87 | Insula | 8.27 | — |
| −15 | −23 | 11 | 2.49 | Thalamus | 10.44 | — |
| −28 | −16 | 10 | 2.93 | Putamen | 9.41 | — |
| −28 | −24 | 3 | 2.46 | Thalamus | 10.20 | — |
| −11 | −47 | −2 | 2.54 | Cerebellum | 7.57 | — |
| −29 | −79 | −19 | 3.67 | Cerebellum | 9.46 | — |
| Right | ||||||
| 45 | −15 | 35 | 4.02 | Precentral gyrus | 13.96 | 4 |
| 43 | 5 | 30 | 1.18 | Inf. frontal gyrus | 8.03 | 44 |
| 8 | −73 | 3 | 3.65 | Lingual gyrus | 10.91 | 18 |
| 17 | −66 | 5 | 3.62 | Lingual gyrus | 10.19 | 18 |
| 25 | −77 | −10 | 3.66 | Mid. occipital gyrus | 9.40 | 19 |
| 19 | −34 | 55 | 2.64 | Sup. parietal lobe | 9.85 | 7 |
| 59 | −10 | 16 | 3.50 | Postcentral gyrus | 12.44 | 43 |
| 1 | −31 | 29 | 2.16 | Post. cingulate | 8.07 | 23 |
| 11 | −44 | 1 | 2.13 | Post. cingulate | 7.95 | 30 |
| 54 | −10 | −6 | 1.55 | Mid. temporal gyrus | 8.43 | 21 |
| 48 | 7 | 3 | 3.14 | Sup. temporal gyrus | 9.23 | 22 |
| 47 | −40 | 8 | 3.68 | Sup. temporal gyrus | 12.54 | 22 |
| 47 | −7 | 9 | 2.09 | Sup. temporal gyrus | 8.61 | 42 |
| 30 | −26 | 10 | 2.90 | Trans. temporal gyrus | 9.79 | 41 |
| 32 | −16 | 13 | 2.38 | Insula | 8.91 | — |
| 36 | −1 | 11 | 1.70 | Insula | 7.54 | — |
| 34 | 14 | 10 | 1.94 | Insula | 7.65 | — |
| 16 | −23 | 11 | 1.98 | Thalamus | 9.62 | — |
| 14 | −64 | −19 | 4.20 | Cerebellum | 10.53 | — |
| 1 | −52 | −22 | 1.14 | Cerebellum | 8.43 | — |
Note: These regions showed no significant differences between tasks or groups.
Regions with an interaction of task × time, as would be anticipated, were located in sensory cortex (Fig. 2A). The regions that had greater activation for the repeat task were located in temporal auditory-related cortex, whereas the regions that had greater activation for the read task were in extrastriate visual cortex (Table 4). For 6 visual regions, only the read task produced significant time courses, whereas the other visual and auditory regions had significant time courses for both tasks.
Figure 2.

Regions showing significant task-related (A) or age-related (B) effects. (A) Task-related regions: Pink indicates regions showing significant activity only for Read; Red indicates regions showing significantly more activity for Read than Repeat; Yellow indicates regions showing significantly more activity for Repeat than Read. An example region, location indicated by the arrow, is shown below the brain images with percent signal change on the y-axis, and time (in MR frames) on the x-axis. Time courses are shown with standard error of the mean. (B) Age-related regions: Light blue indicates regions showing significant activity only in children; Dark blue indicates regions showing significantly more activity in children than in adults. An example region, location indicated by the arrow, is shown below the brain images. All differences surpassed a Z-score of 2.5.
Table 4.
Fifteen brain regions showing an interaction of time × task effect in children and adults
| x | y | z | Size (cm3) | Location | Time × task (Z-score) | Approximate Brodmann area | Time × task (effect) |
| Left | |||||||
| −4 | −16 | 42 | 2.58 | Cingulate gyrus | 2.56 | 24 | Read > Repeat |
| −33 | −86 | −1 | 1.16 | Inf. occipital gyrus | 5.23 | 18 | Read only* |
| −37 | −79 | −7 | 0.46 | Mid. occipital gyrus | 3.46 | 19 | Read only* |
| −28 | −70 | 32 | 0.29 | Sup. occipital gyrus | 2.95 | 19 | Read only* |
| −38 | −43 | 40 | 0.7 | Inf. parietal lobe | 4.46 | 40 | Read only* |
| −43 | −65 | 4 | 0.5 | Mid. temporal gyrus | 4.66 | 37 | Read only* |
| −42 | −31 | 9 | 0.34 | Sup. temporal gyrus | 4.17 | 42 | Repeat > Read* |
| −56 | −30 | 8 | 1.61 | Sup. temporal gyrus | 5.19 | 42 | Repeat > Read* |
| −52 | −16 | 2 | 0.82 | Sup. temporal gyrus | 2.96 | 22 | Repeat > Read* |
| −53 | −25 | −7 | 0.66 | Mid. temporal gyrus | 4.46 | 21 | Repeat > Read* |
| Right | |||||||
| 21 | −89 | −5 | 3.11 | Inf. occipital gyrus | 3.07 | 18 | Read > Repeat |
| 38 | −79 | −2 | 0.98 | Inf. occipital gyrus | 4.79 | 18 | Read > Repeat* |
| 37 | −59 | −14 | 1.95 | Fusiform gyrus | 5.07 | 37 | Read only* |
| 45 | −29 | 11 | 0.46 | Trans. temporal gyrus | 4.37 | 41 | Repeat > Read* |
| 58 | −26 | 8 | 1.41 | Sup. temporal gyrus | 4.6 | 42 | Repeat > Read* |
Note: Asterisk indicates regions from this interaction image, whereas other regions were from the main effect of time image, and also showed a significant effect of time × task.
Regions with an interaction of age × time all showed greater BOLD activity for children than adults. Regions showing this interaction were located primarily in the left hemisphere, and included many posterior parietal and extrastriate locations previously reported in the literature, including the supramarginal gyrus and angular gyrus (Fig. 2B). In 9 of the 38 regions, significant time courses were evident for the children, but not for the adults (Table 5).
Table 5.
Thirty-eight brain regions showing an interaction of time × age effect in children and adults
| x | y | z | Size (cm3) | Location | Time × age (Z-score) | Approximate Brodmann area |
| Left | ||||||
| −10 | 39 | 32 | 0.41 | Sup. frontal gyrus | 3.47 | 9 |
| −21 | 16 | 32 | 1.18 | Cingulate gyrus | 3.13 | 32 |
| −16 | −75 | 29 | 2.82 | Cuneus | 2.54 | 19 |
| −9 | −57 | 29 | 2.29 | Precuneus | 4.15 | 18* |
| −22 | −61 | 24 | 1.09 | Precuneus | 4.45 | 18* |
| −6 | −98 | −1 | 0.77 | Calcarine sulcus | 4.01 | 17* |
| −52 | −42 | 24 | 3.46 | Supramarginal gyrus | 2.93 | 40 |
| −43 | −27 | 25 | 0.66 | Inf. parietal lobe | 4.42 | 40* |
| −33 | −50 | 30 | 0.33 | Inf. parietal lobe | 2.61 | 40 |
| −26 | −64 | 43 | 2.73 | Inf. parietal lobe | 3.14 | 7 |
| −12 | −54 | 40 | 1.45 | Precuneus | 5.17 | 7* |
| −10 | −69 | 43 | 1.38 | Precuneus | 3.62 | 7 |
| −7 | −54 | 49 | 3.22 | Sup. parietal lobe | 3.57 | 7 |
| −49 | −62 | 29 | 1.34 | Angular gyrus | 4.82 | 39* |
| −9 | −35 | 25 | 1.34 | Post. cing. | 5.27 | 23* |
| −16 | −62 | −12 | 0.37 | Cerebellum | 3.21 | —* |
| −15 | −67 | −20 | 4.07 | Cerebellum | 2.65 | — |
| −5 | −69 | −11 | 1.18 | Cerebellum | 3.16 | —* |
| Right | ||||||
| 9 | 44 | 23 | 0.34 | Sup. frontal gyrus | 4.61 | 9* |
| 15 | −78 | 33 | 2.76 | Cuneus | 2.96 | 19 |
| 25 | −80 | 23 | 2.92 | Sup. occipital gyrus | 2.60 | 19 |
| 23 | −80 | 9 | 0.36 | Med. occipital gyrus | 3.15 | 19* |
| 9 | −46 | 61 | 1.18 | Sup. parietal lobe | 3.35 | 7 |
| 7 | −49 | 48 | 3.03 | Sup. parietal lobe | 2.81 | 7 |
| 7 | −61 | 48 | 1.38 | Sup. parietal lobe | 3.43 | 7 |
| 16 | −50 | 44 | 2.00 | Precuneus | 4.84 | 7* |
| 21 | −68 | 45 | 1.90 | Precuneus | 3.10 | 7 |
| 29 | −58 | 46 | 2.42 | Inf. parietal lobe | 2.70 | 7 |
| 43 | −57 | 44 | 0.41 | Inf. parietal lobe | 4.40 | 40* |
| 37 | −39 | 24 | 0.40 | Inf. parietal lobe | 4.20 | 40* |
| 45 | −50 | 24 | 1.97 | Inf. parietal lobe | 4.81 | 39* |
| 36 | −67 | 22 | 1.97 | Mid. temporal gyrus | 3.26 | 39 |
| 40 | −47 | 11 | 1.52 | Sup. temporal gyrus | 4.59 | 22* |
| 49 | −58 | 8 | 2.94 | Mid. temporal gyrus | 2.56 | 37 |
| 32 | −56 | −5 | 1.22 | Inf. temporal gyrus | 4.19 | 37* |
| 9 | −43 | 26 | 1.66 | Post. cingulate | 4.53 | 23* |
| 8 | −56 | 26 | 1.65 | Post. cingulate | 3.56 | 31* |
| 6 | −72 | −10 | 1.43 | Cerebellum | 3.39 | —* |
Note: Asterisk indicates regions from this interaction image, whereas other regions were from the main effect of time image, and also showed a significant effect of time × age. Bold indicates significant activity in children only (no significant adult activity), whereas the other regions were significantly active in both groups, but significantly less so in adults.
Regions showing both an interaction of age × time and task × time, but not a 3-way interaction, appeared most often to be driven by a flat BOLD response for the repeat task in adults. These 5 regions were all in the posterior temporal and extrastriate occipital cortex, and included a region close to the hypothesized vWFA (Table 6) (Fig. 3). No regions showed a 3-way interaction effect of time × age × task.
Table 6.
Five brain regions showing both time × task and time × age effects
| x | y | z | Size (cm3) | Location | Time × task (Z-score) | Time × age (Z-score) | Approximate Brodmann area |
| Left | |||||||
| −38 | −60 | −12 | 0.76 | Fusiform gyrus | 2.75 | 4.23 | 19/37 |
| −41 | −68 | −12 | 0.47 | Fusiform gyrus | 4.39 | 2.52 | 19 |
| −19 | −97 | −8 | 0.38 | Lingual gyrus | 3.61 | 4.12 | 17 |
| Right | |||||||
| 40 | −69 | −7 | 3.35 | Mid. occipital gyrus | 3.28 | 3.34 | 19 |
| 43 | −69 | 6 | 2.50 | Mid. occipital gyrus | 3.62 | 3.3 | 19/39 |
Figure 3.

Five bilateral extrastriate cortex regions showing both significant task-related and significant age-related effects. An example from each hemisphere, locations indicated by the arrows, are shown below the image with percent signal change on the y-axis, and time (in MR frames) on the x-axis. Time courses are shown with standard error of the mean. The left hemisphere exemplar region is within 8 mm of the purported vWFA.
Subsidiary Analyses
Subsidiary analyses were performed on 3 left hemisphere regions similar to those described in prior studies of reading: a region in the anterior extrastriate cortex, a region in the supramarginal gyrus, and 1 in the angular gyrus.
The left anterior extrastriate (−38, −60, −12) region is a vector distance of only 7.8 mm away from the location (−43, −54, −12) described by Cohen et al. (2000); McCandliss et al. (2003) as the vWFA. This region showed an interaction of task × time with greater activation for the read task than for the repeat task (Z value = 2.75), and also showed a significant interaction of age and time with children having greater activation than adults (Z value = 4.23). In post hoc analyses, both the read task and repeat task BOLD responses were significantly greater in children than adults (P < 0.05 for comparison of adults and children on the repeat task, P < 0.005 on the read task, P < 0.005 comparing groups collapsed across task). Interestingly, there was no significant difference for children between the 2 tasks (P = 0.3), whereas there was a significant difference between the 2 tasks in adults (P < 0.005), because adults did not activate the region for the repeat task. This pattern was also true for 4 other extrastriate regions (Table 6). As will be discussed, this combination is suggestive of increasing visual specificity within the extrastriate cortex with age (Fig. 3).
The region in the left supramarginal gyrus (−52, −42, +24) showed a significant interaction of age × time (Z value = 2.93), and all 4 time courses showed a significant main effect of time (P < 0.01). This region was activated significantly more by children than adults for both tasks (P < 0.05 for post hoc comparisons of adults and children on the repeat task, P < 0.005 on the read task, P < 0.01 comparing groups collapsed across task), although both groups showed significant activations (each time course, adult read, adult repeat, child read, and child repeat, was significantly active compared with baseline, P < 0.0001, as tested post hoc) (A in Fig. 4).
Figure 4.

Two left hemisphere regions of interest. Region A, in the supramarginal gyrus, shows greater activity for children than adults. Region B, in the angular gyrus, shows significant activity only in children. Time courses are shown with standard error of the mean.
Finally, the region in the left angular gyrus (−49, −62, +29) also showed a significant interaction of age × time (Z value = 4.82). Unlike the supramarginal gyrus region, the angular gyrus region was significantly activated only by children for both tasks; this region was not activated in adults (child read and child repeat time courses were significantly active compared with baseline, P < 0.005, adult read and adult repeat time courses were not, P = 0.62, P = 0.36, respectively, as determined post hoc) (B in Fig. 4).
Discussion
Summary of Key Points
This study has several implications for understanding the development of reading-related processes in the brain. First, children and adults are using the majority of regions of the brain similarly for these tasks, including regions previously implicated in adult studies of reading such as the left inferior frontal gyrus/frontal operculum and the superior temporal gyrus. Second, examination of the age-related differences indicates that compared with adults, children are recruiting additional posterior brain regions to perform these tasks. Third, bilateral extrastriate cortex shows evidence of multimodal-to-unimodal tuning over age. Finally, probing the developmental differences in regions previously reported to be engaged in phonological and/or orthographical processing, for example, regions in the left supramarginal and angular gyri, offers insight into how reading-related networks might be recruited dynamically over time.
Many Regions Behave Similarly in Children and Adults
Age-Invariant and Task-Invariant Regions
Our results indicate that, when performance is matched, children and adults share a common overall pattern of brain activity during overt reading of high-frequency single words. This pattern resembles the distribution of regions seen in meta-analyses of functional neuroimaging studies of single-word reading (Fiez and Petersen 1998; Turkeltaub et al. 2002; Jobard et al. 2003; Bolger et al. 2005). The similarity of activity between ages in numerous regions, including cingulate, insula, motor and primary sensory regions, implies that these regions are functioning at adult-like levels by age 7, which is consistent with previous results (e.g. Gaillard et al. 2003a, 2003b; Turkeltaub et al. 2003; Brown et al. 2005). Because reaction time and accuracy between groups were equated in this study, and because both of the tasks in this study had an overt verbal output, motor regions would not be anticipated to be significantly different between age groups or tasks.
A key distinction between the read task and the repeat task is that the former requires the translation of visual (orthographic) input to verbal (phonological) output, whereas the latter task emphasizes the translation of auditory (phonological) input to verbal (phonological) output. The different modality input for each task highlights regions of integration where both tasks show similar levels of activity.
Age-Invariant and Task-Dependent Regions
The regions that differed by task included mainly auditory and visual processing regions. All auditory cortex regions were activated significantly more by the repeat task, but were significantly active for both the read and repeat tasks. This result was expected due to the self-generated auditory stimulation from the verbal output demanded in both tasks.
Visual regions, although showing greater activity during reading tasks, in some cases also showed positive activity for the repeat task. This result may have occurred due to participants’ visualization of the words they were hearing. These task-related differences are similar to those reported in adults (Price 2000). As in regions showing no age or task effects, the lack of differences between children and adults for these regions implies a common sensory processing network that is operating at adult levels by age 7.
Reading-Related Regions
Significantly different activity was lacking between groups and between tasks in left frontal and insular regions around the left frontal operculum. This region has been described in other studies as integral to an anterior circuit for reading that is highly sensitive to regularity, and shows greater activity for pseudowords than real words (Fiez et al. 1999; Pugh et al. 2001; Mechelli et al. 2003). Properties of the words employed in the stimulus set (i.e., high frequency, high regularity) may have prevented the ability to identify a difference between the tasks or groups in this region. Another possibility is that the contribution of this region to these tasks matures earlier than other reading-related regions, and reached adult activity levels by age 7 years.
Significantly different activity was also lacking between groups and between tasks in the left superior temporal gyrus (−52, −45, +8). This general region has been reported in other studies of reading as important in phonological and/or semantic processing (e.g., Bolger et al. 2005; Vigneau et al. 2006). The positive activity for both tasks in both groups suggests that this region, like the left frontal operculum region, supports a consistent processing role for these tasks over the ages studied.
Age-Dependent Effects
Task-Invariant, Age-Dependent Regions
Age-dependent effects were all due to child BOLD activity being greater than adult BOLD activity. These regions were nearly all in the posterior half of the brain, and included bilateral activation in the parietal lobe, midline occipital lobe, and posterior cingulate, right hemisphere activation at the temporal–occipital junction, and also included activation in the left angular and supramarginal gyri (discussed below). The general location of these regions is consistent with other studies from our group which report a “growing down” pattern during development for lexical tasks in posterior portions of the brain (e.g., Brown et al. 2005). For many regions showing age-related differences, the differences were in the magnitude of activation, as adults still engaged the regions. This finding suggests that these regions become more efficient, or more specialized for certain processes over age, and thus show a decrease in activity in adults. Some regions, however, apparently dropped out completely in adults, suggesting that children may recruit additional regions of the brain to perform the tasks at the same level of proficiency as adults.
Of interest, regions that show greater activity for adults than children were not detected. Several regions did show a pattern of greater activity in adults than children in a previous report by our group, which analyzed 3 controlled lexical tasks (rhyme, verb, and opposite generation) (Brown et al. 2005). Other developmental studies have also shown regions of greater activity in adults (Bunge et al. 2002; Turkeltaub et al. 2003). One possibility for this lack of effect is that because children generally have more variable data than adults, potential differences are masked. There are at least 2 potential sources of this variability. First, variability could come from the groups being more variable on IQ or another subject characteristic. Although we were not able to do direct comparison of IQs, this does not appear to be the case, as both of our groups are estimated to be a standard deviation above the mean for intelligence. Second, greater variability in children could arise from the age range of the children representing a greater range of developmental difference than the adult sample. This source of variability is likely influencing our data and would decrease the likelihood of finding any group differences. Inspection of time courses, as was done in the present study, is critical to reducing incorrect classification of regions that may contain false positives or false negatives. This inherent bias against finding differences could have reduced reporting of regions with greater adult activity, however, in addition to statistical measures, no region time courses were found where that pattern was the case. Two additional possibilities for the lack of regions showing greater activity in adults than children could be either the relative ease and familiarity of the tasks and stimuli for both groups, or that regions which become more involved over development have already reached their adult levels by age 7.
Why do children show greater activity than adults in some regions during single-word repeating? The regressive effect of high activity in children to less or no activity in adults for a given region suggests that reading development involves intraregional specialization with maturity. This concept has also been suggested by others (e.g., Durston et al. 2006). Importantly, an isolated increase or decrease in a single region may not completely reflect the broader story of functional specialization. This concept has been discussed in the framework of interactive specialization advanced by Johnson and colleagues (Johnson 2000, 2003; Johnson et al. 2002), who posit that the functional role of a given region is shaped by its activity-dependent connections with other regions. Early in development, performance of a specific task might be served by a number of regions whose functional contributions in the face of that task's demands are not well defined. As these regions compete with each other to perform the task, the connections between the regions are altered and the relative contributions of the regions change. The presence of regions showing child-only activity in the present study indicates that functional relationships may indeed be reflecting alterations between these regions and those that remain active in adults. Alternatively, reduction in activation may also reflect, in part, more general developmental changes such as synaptic pruning (e.g., Huttenlocher et al. 1982). However, a general mechanism such as synaptic pruning will not be able to account for task-dependent developmental effects.
Bilateral Extrastriate Shows Modality Tuning Over Age
Five bilateral extrastriate regions showed interaction effects both of time × task and of time × age. One region with both interactions was in the left extrastriate cortex near the vWFA (discussed below). All 5 regions with both time × task and time × age interactions showed a similar pattern of positive activity for both tasks in children, and adult activity for only the reading task, suggesting that the regions become more tuned for unimodal (i.e., visual) information with age. These results are consistent with those of Booth et al. (2003) who have previously reported that children, compared with adults, had less modality-specific activity in visual regions. The tuning of bilateral extrastriate cortex for visual information over development is an intriguing finding, implying that in addition to the idea that different parts of the extrastriate cortex become specialized for particular types of visual information (such as a vWFA, McCandliss et al. 2003 or a fusiform face area; Kanwisher et al. 1997;), regions of the extrastriate cortex may show progressive specialization from a more multimodal (e.g., auditory and visual) to unimodal (e.g., visual) processing over development, or become more segregated in purpose.
A left anterior extrastriate region, with Talairach coordinates at −38, −60, −12, showed both age and task effects in this study; it showed similar activity for both tasks in children, but significantly different activity between tasks in adults. Children also had greater activity in this region for both tasks compared with adults, suggesting greater efficiency and specialization in this region in adults. This region has a vector distance of about 8 mm from the putative vWFA by Cohen et al. (2000) (−43, −54, −12). The present findings do support a visually predominant processing role for this region in adults. However, the present findings suggest a lack of specificity for the visual modality in children, suggesting that the region becomes more modality-specific with maturity and/or experience, and/or more general developmental mechanisms such as synaptic consolidation (but see Brown et al. 2006). In explaining this result, it is possible to surmise that children are performing more visualization of words during the repeat task than the adults, despite equivalent reaction times. On the other hand, this lack of specificity could reflect feedback from phonological processing regions in the angular and supramarginal gyri that show decreasing activity with age (see below). The possibility of increasing modality specificity in the vWFA with development needs to be further explored. Developmental studies with a wider variety of visual word stimuli may be better able to tease apart these 2 possibilities.
Putative Phonological Regions in the Angular and Supramarginal Gyri
Regions in the angular gyrus and supramarginal gyrus, widely reported in reading research, showed age-dependent effects in this study. For the supramarginal gyrus, activity in adults was weaker in amplitude than the activity for children. This finding is consistent with the possibility that this region is involved in phonological processing, or in orthographic to phonological conversions (Price 1998; Pugh et al. 2001; Lee et al. 2007). The fact that we find this activity decrease for both the read and repeat tasks suggests that the supramarginal gyrus may be more involved in general phonological processing than in orthographic to phonological conversions per se.
A region only 2 cm away, in the angular gyrus, was not active in adults, whereas it was active in children. Some authors have suggested that the angular gyrus is involved in semantic processing (Price 2000; Binder et al. 2005; Sabsevitz et al. 2005; Lee et al. 2007). Although, in general, the coordinates reported for regions showing semantic effects are more dorsally located than this region, the lack of activity in adults in the angular gyrus region for the familiar, high frequency, concrete words decreases the likelihood of (but does not rule out) a role for the currently reported angular gyrus region in semantic processing. The similar profile (decreasing activation across both tasks with age) is most consistent with the possibility that, along with the supramarginal gyrus region, the angular gyrus region is part of a dorsal phonological system as suggested by Pugh et al. (2001). However, the lack of activation in adults, which differs from the supramarginal region, suggests a separable role for this region, at least in adults.
Lateralization
Although not examined directly, this study did not find evidence for an increasing left lateralization for single-word reading across development. Several fMRI studies of the development of language (but not overt, single-word reading/repetition) have reported increasing left lateralization with development from early school-age into young adulthood (Gaillard et al. 2000; Turkeltaub et al. 2003; Brown et al. 2005; Szaflarski et al. 2006). Studies of controlled lexical processing (Gaillard et al. 2000; Szaflarski et al. 2006), including our own (Brown et al. 2005), are generally consistent with increasing left lateralization with development but these studies differ substantially from the present in terms of the task (e.g., verbal fluency, lexical generation) and analysis methods. Eden and colleagues (Turkeltaub et al. 2003) employed a covert task of word reading and found that task-related activity in left hemisphere regions increased with reading skill, whereas such activity in right hemisphere regions decreased with reading skill. The present data suggest that at least for single-word reading and repetition, both developmentally stable and developmentally dynamic regions are found in both hemispheres.
Future Studies
It is interesting to consider whether the developmental changes observed are the result of decreasing reliance on network components in general, or whether these changes are task or stimulus dependent. There is the possibility that the lack of activity for adults in the angular gyrus, and the decreased activity for adults in the supramarginal gyrus may be due to their extreme familiarity with the high-frequency stimuli. The high-frequency first-grade reading-level words are more familiar for adults than for children, due to the numerous experiences with these words throughout their lifetimes as readers. This result informs research using computational modeling as well. If, as has been suggested by others, interactions between a dorsal phonological system and a ventral orthographic system serve to train the latter (Pugh et al. 2001; McCandliss and Noble 2003), what happens to phonological system activity levels after the ventral system is trained? Our results suggest that as the ventral system (at least in the location of the vWFA) becomes more selective for visual over auditory word input, activity in the dorsal system decreases. We are currently investigating whether there are ways to modulate the amount of activation in the angular and supramarginal gyri in adults with more challenging reading stimuli, or whether these regions become permanently lessened or inactive for reading tasks when reading proficiency is established.
Conclusions
This study of reading in children and adults shows that these groups have much in common when reading high-frequency words aloud. However, the differences observed between children and adults in a key set of regions implicated in adult reading indicate that children are not fully adult-like when performing a reading task. The use of a comparison task of similar difficulty, and with some overlapping processing demands, but in a different modality (repeating aloud aurally presented words) allowed the observation of task and group similarities and differences. There is evidence from examining these tasks that children have bilateral extrastriate regions that are less unimodally (visually) tuned than adults. In addition, hypothesized phonological regions decrease in activity in adults compared with children for high-frequency words. Overall, the developmental differences found in the present study suggest that compared with adults, children recruit additional posterior brain regions to perform single-word reading, and that with development the reading network becomes better tuned for the processing and output of auditory and visual words.
Funding
Mr. and Mrs. Spencer T. Olin Fellowship Program to J.A.C.; National Institutes of Health (NIH) Neurological Sciences Academic Development Award to B.L.S., NIH (NS053425) to B.L.S., NIH (NS32979) to S.E.P., NIH (NS41255) to S.E.P., NIH (NS46424) to S.E.P.; The McDonnell Center for Higher Brain Function to S.E.P. and B.L.S; Brad Schlaggar is a Scholar of the Child Health Resource Center of Excellence in Developmental Biology at Washington University School of Medicine (HD01487).
Acknowledgments
The authors wish to thank all of the participants in this study. We thank Francis M. Miezin for helpful discussions about the manuscript. Portions of this work were presented at the 10th annual meeting of the Organization for Human Brain Mapping, and the 36th annual meeting of the Society for Neuroscience. Conflict of Interest: None declared.
References
- Barber HA, Kutas M. Interplay between computational models and cognitive electrophysiology in visual word recognition. Brain Res Rev. 2007;53:98–123. doi: 10.1016/j.brainresrev.2006.07.002. [DOI] [PubMed] [Google Scholar]
- Binder JR, Medler DA, Desai R, Conant LL, Liebenthal E. Some neurophysiological constraints on models of word naming. Neuroimage. 2005;27:677–693. doi: 10.1016/j.neuroimage.2005.04.029. [DOI] [PubMed] [Google Scholar]
- Binder JR, Mohr JP. The topography of callosal reading pathways. A case-control analysis. Brain. 1992;115(Pt 6):1807–1826. doi: 10.1093/brain/115.6.1807. [DOI] [PubMed] [Google Scholar]
- Birn RM, Bandettini PA, Cox RW, Shaker R. Event-related fMRI of tasks involving brief motion. Hum Brain Mapp. 1999;7:106–114. doi: 10.1002/(SICI)1097-0193(1999)7:2<106::AID-HBM4>3.0.CO;2-O. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blumenfeld HK, Booth JR, Burman DD. Differential prefrontal-temporal neural correlates of semantic processing in children. Brain Lang. 2006;99:226–235. doi: 10.1016/j.bandl.2005.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolger DJ, Perfetti CA, Schneider W. Cross-cultural effect on the brain revisited: universal structures plus writing system variation. Hum Brain Mapp. 2005;25:92–104. doi: 10.1002/hbm.20124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TB, Mesulam MM. Development of brain mechanisms for processing orthographic and phonologic representations. J Cogn Neurosci. 2004;16:1234–1249. doi: 10.1162/0898929041920496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Lei Z, Choy J, Gitelman DR, Parrish TB, Mesulam MM. Modality-specific and -independent developmental differences in the neural substrate for lexical processing. J Neurolinguist. 2003;16:383–405. [Google Scholar]
- Box GEP. Some theorems on quadratic forms applied in the study of analysis of variance problems, II. Effects of inequality of variance and of correlation between errors in the two-way classification. Ann Math Stat. 1954;25:484–498. [Google Scholar]
- Brem S, Bucher K, Halder P, Summers P, Dietrich T, Martin E, Brandeis D. Evidence for developmental changes in the visual word processing network beyond adolescence. Neuroimage. 2006;29:822–837. doi: 10.1016/j.neuroimage.2005.09.023. [DOI] [PubMed] [Google Scholar]
- Broca P. Perte de la parole, remollissement chronique et destruction partielle du lobe anterieur gauche du cerveau. Bull Soc Anthropol. 1861;2:235–238. [Google Scholar]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Developmental changes in human cerebral functional organization for word generation. Cereb Cortex. 2005;15:275–290. doi: 10.1093/cercor/bhh129. [DOI] [PubMed] [Google Scholar]
- Brown TT, Petersen SE, Schlaggar BL. Does human functional brain organization shift from diffuse to focal with development? Dev Sci. 2006;9:9–12. doi: 10.1111/j.1467-7687.2005.00455.x. [DOI] [PubMed] [Google Scholar]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Immature frontal lobe contributions to cognitive control in children: evidence from fMRI. Neuron. 2002;33:301–311. doi: 10.1016/s0896-6273(01)00583-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgund ED, Kang H-SC, Kelly JE, Buckner RL, Snyder AZ, Petersen SE, Schlaggar BL. The feasibility of a common stereotactic space for children and adults in fMRI studies of development. Neuroimage. 2002;17:184–200. doi: 10.1006/nimg.2002.1174. [DOI] [PubMed] [Google Scholar]
- Ciesielski KT, Lesnik PG, Savoy RL, Grant EP, Ahlfors SP. Developmental neural networks in children performing a Categorical N-Back Task. Neuroimage. 2006;33:980–990. doi: 10.1016/j.neuroimage.2006.07.028. [DOI] [PubMed] [Google Scholar]
- Cohen L, Dehaene S, Naccache L, Lehericy S, Dehaene-Lambertz G, Henaff MA, Michel F. The visual word form area: spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain. 2000;123(Pt 2):291–307. doi: 10.1093/brain/123.2.291. [DOI] [PubMed] [Google Scholar]
- Cohen L, Lehericy S, Chochon F, Lemer C, Rivaud S, Dehaene S. Language-specific tuning of visual cortex? Functional properties of the visual word form area. Brain. 2002;125:1054–1069. doi: 10.1093/brain/awf094. [DOI] [PubMed] [Google Scholar]
- Damasio AR, Damasio H. The anatomic basis of pure alexia. Neurology. 1983;33:1573–1583. doi: 10.1212/wnl.33.12.1573. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Le Clec HG, Poline JB, Le Bihan D, Cohen L. The visual word form area: a prelexical representation of visual words in the fusiform gyrus. Neuroreport. 2002;13:321–325. doi: 10.1097/00001756-200203040-00015. [DOI] [PubMed] [Google Scholar]
- Dejerine J. Sur un cas de cécité verbale avec agraphie, suivi d'autopsie. C R Hebd Seances Mem Soc Biol. 1891;3:197–201. [Google Scholar]
- Dejerine J. Contribution a l'étude anatomoclinique et clinique des differentes varietes de cecite verbal. C R Hebd Seances Mem Soc Biol. 1892;4:61–90. [Google Scholar]
- Devlin JT, Jamison HL, Gonnerman LM, Matthews PM. The role of the posterior fusiform gyrus in reading. J Cogn Neurosci. 2006;18:911–922. doi: 10.1162/jocn.2006.18.6.911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella JA, Casey BJ. A shift from diffuse to focal cortical activity with development. Dev Sci. 2006;9:1–8. doi: 10.1111/j.1467-7687.2005.00454.x. [DOI] [PubMed] [Google Scholar]
- Eden GF, Zeffiro TA. Neural systems affected in developmental dyslexia revealed by functional neuroimaging. Neuron. 1998;21:279–282. doi: 10.1016/s0896-6273(00)80537-1. [DOI] [PubMed] [Google Scholar]
- Fair DA, Brown TT, Petersen SE, Schlaggar BL. A comparison of ANOVA and correlation methods for investigating cognitive development with fMRI. Dev Neuropsychol. 2006a;30:531–546. doi: 10.1207/s15326942dn3001_2. [DOI] [PubMed] [Google Scholar]
- Fair DA, Brown TT, Petersen SE, Schlaggar BL. fMRI reveals novel functional neuroanatomy in a child with perinatal stroke. Neurology. 2006b;67:2246–2249. doi: 10.1212/01.wnl.0000249348.84045.0e. [DOI] [PubMed] [Google Scholar]
- Fiez JA, Balota DA, Raichle ME, Petersen SE. Effects of lexicality, frequency, and spelling-to-sound consistency on the functional anatomy of reading. Neuron. 1999;24:205–218. doi: 10.1016/s0896-6273(00)80833-8. [DOI] [PubMed] [Google Scholar]
- Fiez JA, Petersen SE. Neuroimaging studies of word reading. Proc Natl Acad Sci USA. 1998;95:914–921. doi: 10.1073/pnas.95.3.914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiez JA, Tranel D, Seager-Frerichs D, Damasio H. Specific reading and phonological processing deficits are associated with damage to the left frontal operculum. Cortex. 2006;42:624–643. doi: 10.1016/s0010-9452(08)70399-x. [DOI] [PubMed] [Google Scholar]
- Forman SD, Cohen JD, Fitzgerald M, Eddy WF, Mintun MA, Noll DC. Improved assessment of significant activation in functional magnetic resonance imaging (fMRI): use of a cluster-size threshold. Magn Reson Med. 1995;33:636–647. doi: 10.1002/mrm.1910330508. [DOI] [PubMed] [Google Scholar]
- Gaillard WD, Balsamo LM, Ibrahim Z, Sachs BC, Xu B. fMRI identifies regional specialization of neural networks for reading in young children. Neurology. 2003a;60:94–100. doi: 10.1212/wnl.60.1.94. [DOI] [PubMed] [Google Scholar]
- Gaillard WD, Hertz-Pannier L, Mott SH, Barnett AS, LeBihan D, Theodore WH. Functional anatomy of cognitive development: fMRI of verbal fluency in children and adults. Neurology. 2000;54:180–185. doi: 10.1212/wnl.54.1.180. [DOI] [PubMed] [Google Scholar]
- Gaillard WD, Sachs BC, Whitnah JR, Ahmad Z, Balsamo LM, Petrella JR, Braniecki SH, McKinney CM, Hunter K, Xu B, et al. Developmental aspects of language processing: fMRI of verbal fluency in children and adults. Hum Brain Mapp. 2003b;18:176–185. doi: 10.1002/hbm.10091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geschwind N. Disconnexion syndromes in animals and man, Part 1. Brain. 1965;88:237–294. doi: 10.1093/brain/88.2.237. [DOI] [PubMed] [Google Scholar]
- Hagoort P, Indefrey P, Brown C, Herzog H, Steinmetz H, Seitz RJ. The neural circuitry involved in the reading of German words and pseudowords: a PET study. J Cogn Neurosci. 1999;11:383–398. doi: 10.1162/089892999563490. [DOI] [PubMed] [Google Scholar]
- Hillis AE, Newhart M, Heidler J, Barker P, Herskovits E, Degaonkar M. The roles of the “visual word form area” in reading. Neuroimage. 2005;24:548–559. doi: 10.1016/j.neuroimage.2004.08.026. [DOI] [PubMed] [Google Scholar]
- Hillis AE, Rapp BC, Caramazza A. When a rose is a rose in speech but a tulip in writing. Cortex. 1999;35:337–356. doi: 10.1016/s0010-9452(08)70804-9. [DOI] [PubMed] [Google Scholar]
- Hoeft F, Hernandez A, McMillon G, Taylor-Hill H, Martindale JL, Meyler A, Keller TA, Siok WT, Deutsch GK, Just MA, et al. Neural basis of dyslexia: a comparison between dyslexic and nondyslexic children equated for reading ability. J Neurosci. 2006;26:10700–10708. doi: 10.1523/JNEUROSCI.4931-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horwitz B, Rumsey JM, Donohue BC. Functional connectivity of the angular gyrus in normal reading and dyslexia. Proc Natl Acad Sci USA. 1998;95:8939–8944. doi: 10.1073/pnas.95.15.8939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huttenlocher PR, de Courten C, Garey LJ, Van der Loos H. Synaptogenesis in human visual cortex—evidence for synapse elimination during normal development. Neurosci Lett. 1982;33:247–252. doi: 10.1016/0304-3940(82)90379-2. [DOI] [PubMed] [Google Scholar]
- Jobard G, Crivello F, Tzourio-Mazoyer N. Evaluation of the dual route theory of reading: a metanalysis of 35 neuroimaging studies. Neuroimage. 2003;20:693–712. doi: 10.1016/S1053-8119(03)00343-4. [DOI] [PubMed] [Google Scholar]
- Johnson MH. Functional brain development in infants: elements of an interactive specialization framework. Child Dev. 2000;71:75–81. doi: 10.1111/1467-8624.00120. [DOI] [PubMed] [Google Scholar]
- Johnson MH. Functional brain development in humans. Nat Rev Neurosci. 2001;2:475–483. doi: 10.1038/35081509. [DOI] [PubMed] [Google Scholar]
- Johnson MH. Development of human brain functions. Biol Psychiatry. 2003;54:1312–1316. doi: 10.1016/s0006-3223(03)00426-8. [DOI] [PubMed] [Google Scholar]
- Johnson MH, Halit H, Grice SJ, Karmiloff-Smith A. Neuroimaging of typical and atypical development: a perspective from multiple levels of analysis. Dev Psychopathol. 2002;14:521–536. doi: 10.1017/s0954579402003073. [DOI] [PubMed] [Google Scholar]
- Joseph J, Noble K, Eden G. The neurobiological basis of reading. J Learn Disabil. 2001;34:566–579. doi: 10.1177/002221940103400609. [DOI] [PubMed] [Google Scholar]
- Joubert S, Beauregard M, Walter N, Bourgouin P, Beaudoin G, Leroux JM, Karama S, Lecours AR. Neural correlates of lexical and sublexical processes in reading. Brain Lang. 2004;89:9–20. doi: 10.1016/S0093-934X(03)00403-6. [DOI] [PubMed] [Google Scholar]
- Kang H-SC, Burgund ED, Lugar HM, Petersen SE, Schlaggar BL. Comparison of functional activation foci in children and adults using a common stereotactic space. Neuroimage. 2003;19:16–28. doi: 10.1016/s1053-8119(03)00038-7. [DOI] [PubMed] [Google Scholar]
- Kanwisher N, McDermott J, Chun MM. The fusiform face area: a module in human extrastriate cortex specialized for face perception. J Neurosci. 1997;17:4302–4311. doi: 10.1523/JNEUROSCI.17-11-04302.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lancaster JL, Glass TG, Lankipalli BR, Downs H, Mayberg H, Fox PT. A modality-independent approach to spatial normalization of tomographic images of the human brain. Hum Brain Mapp. 1995;3:209–223. [Google Scholar]
- Lee H, Devlin JT, Shakeshaft C, Stewart LH, Brennan A, Glensman J, Pitcher K, Crinion J, Mechelli A, Frackowiak RS, et al. Anatomical traces of vocabulary acquisition in the adolescent brain. J Neurosci. 2007;27:1184–1189. doi: 10.1523/JNEUROSCI.4442-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matarazzo JD. Wechsler's measurement and appraisal of adult intelligence. Baltimore: Williams and Wilkins; 1972. [Google Scholar]
- Maurer U, Brandeis D, McCandliss BD. Fast, visual specialization for reading in English revealed by the topography of the N170 ERP response. Behav Brain Funct. 2005;1:13. doi: 10.1186/1744-9081-1-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maurer U, Brem S, Kranz F, Bucher K, Benz R, Halder P, Steinhausen HC, Brandeis D. Coarse neural tuning for print peaks when children learn to read. Neuroimage. 2006;33:749–758. doi: 10.1016/j.neuroimage.2006.06.025. [DOI] [PubMed] [Google Scholar]
- McAvoy MP, Ollinger JM, Buckner RL. Cluster size thresholds for assessment of significant activation in fMRI. Neuroimage. 2001;13:S198. [Google Scholar]
- McCandliss BD, Cohen L, Dehaene S. The visual word form area: expertise for reading in the fusiform gyrus. Trends Cogn Sci. 2003;7:293–299. doi: 10.1016/s1364-6613(03)00134-7. [DOI] [PubMed] [Google Scholar]
- McCandliss BD, Noble KG. The development of reading impairment: a cognitive neuroscience model. Ment Retard Dev Disabil Res Rev. 2003;9:196–204. doi: 10.1002/mrdd.10080. [DOI] [PubMed] [Google Scholar]
- Mechelli A, Gorno-Tempini ML, Price CJ. Neuroimaging studies of word and pseudoword reading: consistencies, inconsistencies, and limitations. J Cogn Neurosci. 2003;15:260–271. doi: 10.1162/089892903321208196. [DOI] [PubMed] [Google Scholar]
- Miezin FM, Maccotta L, Ollinger JM, Petersen SE, Buckner RL. Characterizing the hemodynamic response: Effects of presentation rate, sampling procedure, and the possibility of ordering brain activity based on relative timing. Neuroimage. 2000;11:735–759. doi: 10.1006/nimg.2000.0568. [DOI] [PubMed] [Google Scholar]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E, Charney DS, Ernst M, Pine DS. Adolescent immaturity in attention-related brain engagement to emotional facial expressions. Neuroimage. 2003;20:420–428. doi: 10.1016/s1053-8119(03)00355-0. [DOI] [PubMed] [Google Scholar]
- Nelles JL, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. An automated method for extracting response latencies of subject vocalizations in event-related fMRI experiments. Neuroimage. 2003;20:1865–1871. doi: 10.1016/j.neuroimage.2003.07.020. [DOI] [PubMed] [Google Scholar]
- Palmer ED, Brown TT, Petersen SE, Schlaggar BL. Investigation of the functional neuroanatomy of single word reading and its development. Sci Stud Read. 2004;8:203–223. [Google Scholar]
- Palmer ED, Rosen HJ, Ojemann JG, Buckner RL, Kelley WM, Petersen SE. An event-related fMRI study of overt and covert word stem completion. Neuroimage. 2001;14:182–193. doi: 10.1006/nimg.2001.0779. [DOI] [PubMed] [Google Scholar]
- Plaut DC, McClelland JL, Seidenberg MS, Patterson K. Understanding normal and impaired word reading: Computational principles in quasi-regular domains. Psychol Rev. 1996;103:56–115. doi: 10.1037/0033-295x.103.1.56. [DOI] [PubMed] [Google Scholar]
- Price CJ. The functional anatomy of word comprehension and production. Trends Cogn Sci. 1998;2:281–288. doi: 10.1016/s1364-6613(98)01201-7. [DOI] [PubMed] [Google Scholar]
- Price CJ. The anatomy of language: contributions from functional neuroimaging. J Anat. 2000;197:335–359. doi: 10.1046/j.1469-7580.2000.19730335.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price CJ, Devlin JT. The myth of the visual word form area. Neuroimage. 2003;19:473–481. doi: 10.1016/s1053-8119(03)00084-3. [DOI] [PubMed] [Google Scholar]
- Pugh KR, Mencl WE, Jenner AR, Katz L, Frost SJ, Lee JR, Shaywitz SE, Shaywitz BA. Neurobiological studies of reading and reading disability. J Commun Disord. 2001;34:479–492. doi: 10.1016/s0021-9924(01)00060-0. [DOI] [PubMed] [Google Scholar]
- Sabsevitz DS, Medler DA, Seidenberg M, Binder JR. Modulation of the semantic system by word imageability. Neuroimage. 2005;27:188–200. doi: 10.1016/j.neuroimage.2005.04.012. [DOI] [PubMed] [Google Scholar]
- Schlaggar BL, Brown TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Functional neuroanatomical differences between adults and school-age children in the processing of single words. Science. 2002;296:1476–1479. doi: 10.1126/science.1069464. [DOI] [PubMed] [Google Scholar]
- Schlaggar BL, McCandliss BD. Development of neural systems for reading. Annu Rev Neurosci. 2007;30:475–503. doi: 10.1146/annurev.neuro.28.061604.135645. [DOI] [PubMed] [Google Scholar]
- Seidenberg MS. Connectionist models of word reading. Curr Dir Psychol Sci. 2005;14:238–242. [Google Scholar]
- Shaywitz BA, Shaywitz SE, Pugh KR, Mencl WE, Fulbright RK, Skudlarski P, Constable RT, Marchione KE, Fletcher JM, Lyon GR, et al. Disruption of posterior brain systems for reading in children with developmental dyslexia. Biol Psychiatry. 2002;52:101–110. doi: 10.1016/s0006-3223(02)01365-3. [DOI] [PubMed] [Google Scholar]
- Simos PG, Breier JI, Fletcher JM, Foorman BR, Castillo EM, Papanicolaou AC. Brain mechanisms for reading words and pseudowords: an integrated approach. Cereb Cortex. 2002;12:297–305. doi: 10.1093/cercor/12.3.297. [DOI] [PubMed] [Google Scholar]
- Simos PG, Breier JI, Fletcher JM, Foorman BR, Mouzaki A, Papanicolaou AC. Age-related changes in regional brain activation during phonological decoding and printed word recognition. Dev Neuropsychol. 2001;19:191–210. doi: 10.1207/S15326942DN1902_4. [DOI] [PubMed] [Google Scholar]
- Snyder AZ. Difference image vs. ratio image error function forms in PET-PET realignment. In: Myer R, Cunningham VJ, Bailey DL, Jones T, editors. Quantification of brain function using PET. San Diego (CA): Academic Press; 1996. pp. 131–137. [Google Scholar]
- Szaflarski JP, Holland SK, Schmithorst VJ, Byars AW. fMRI study of language lateralization in children and adults. Hum Brain Mapp. 2006;27:202–212. doi: 10.1002/hbm.20177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talairach J, Tournoux P. Co-planar stereotaxic atlas of the human brain. New York: Thieme Medical Publishers, Inc; 1988. [Google Scholar]
- Turkeltaub PE, Eden GF, Jones KM, Zeffiro TA. Meta-analysis of the functional neuroanatomy of single-word reading: method and validation. Neuroimage. 2002;16:765–780. doi: 10.1006/nimg.2002.1131. [DOI] [PubMed] [Google Scholar]
- Turkeltaub PE, Gareau L, Flowers DL, Zeffiro TA, Eden GF. Development of neural mechanisms for reading. Nat Neurosci. 2003;6:767–773. doi: 10.1038/nn1065. [DOI] [PubMed] [Google Scholar]
- Van Essen DC. Windows on the brain: the emerging role of atlases and databases in neuroscience. Curr Opin Neurobiol. 2002;12:574–579. doi: 10.1016/s0959-4388(02)00361-6. http://pulvinar.wustl.edu:8081/sums/search.do?filename=ATLAS&;filetype=tar.gz. [DOI] [PubMed] [Google Scholar]
- Van Essen DC, Dickson J, Harwell J, Hanlon D, Anderson CH, Drury HA. An integrated software suite for surface-based analyses of cerebral cortex. J Am Med Inform Assoc. 2001;41:1359–1378. doi: 10.1136/jamia.2001.0080443. http://brainmap.wustl.edu/caret. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vigneau M, Beaucousin V, Herve PY, Duffau H, Crivello F, Houde O, Mazoyer B, Tzourio-Mazoyer N. Meta-analyzing left hemisphere language areas: phonology, semantics, and sentence processing. Neuroimage. 2006;30:1414–1432. doi: 10.1016/j.neuroimage.2005.11.002. [DOI] [PubMed] [Google Scholar]
- Wechsler D. Wechsler abbreviated scale of intelligence. San Antonio: The Psychological Corporation; 1999. [Google Scholar]
- Wernicke C. Der aphasische Symptomencomplex. Breslau: Kohn and Weigert; 1874. [Google Scholar]