

Purine nucleoside phosphorylase (PNP) is involved in the catabolism and recycling of nucleosides and is a target for the development of novel therapeutic agents for T-cell apoptosis in leukemia and in autoimmune diseases.1-2 Our approach to the design of therapeutic inhibitors is transition state analysis based on experimentally measured kinetic isotope effects. Chemically stable analogues of a transition state complex are predicted to convert the energy of enzymatic rate acceleration (kcat/knon) into binding energy.3 Enzymatic transition state structures, however, cannot be directly observed but must be estimated by the measurement of kinetic isotope effects (KIE) and computational chemistry with appropriate constraints to provide agreement to the intrinsic KIE.3-6 Development of species-specific transition state analogues for use as therapeutic agents requires characterization of each specific transition state. Comparison of the transition state structures for human and bovine PNPs revealed surprising differences for proteins of 87% overall sequence identity.7,8 The goal of this study was to test the validity of exerimental transition state analysis by comparing the binding affinity of transition state analogues designed for specific transition states of the closely related bovine and human PNPs.
The transition state for the arsenolysis reaction catalyzed by bovine PNP (BtPNP) was solved by KIE measurements in 1993, and was characterized by a 0.38 Pauling bond order to the leaving group (1.8 Å separation) with only van der Waals contact to the attacking arsenate nucleophile (Figure 1).7 This structural guide for transition state analogue design led to the synthesis of Immucillin-H [1] and Immucillin-G [2], which approximate the transition state for the PNP catalyzed reactions with inosine and guanosine, respectively.9,10 These compounds resemble an early transition state with a distance between the ribose and the base of 1.5 Å. The transition state for the arsenolysis reaction catalyzed by human PNP (HsPNP) was solved in 2004 and the transition state was determined to resemble a fully dissociated ribooxacarbenium ion (Figure 1).8 The attacking arsenate nucleophile and the departing base are both in van der Waals contact with no significant bond character to the ribose. The 3.0 Å distance between ribose and the departing base was used as a design feature to give DADMe-Immucillin-H [3] and DADMe-Immucillin-G [4], which have a 2.5 Å distance between the hydroxyl-pyrrolidine and the 9-deaza-base.11 Chemical stability of 3 requires absence of the 2’-hydroxyl, a feature suitable for HsPNP as its primary physiological substrate is 2’-deoxyguanosine. A second design feature was to place the N-cationic center at the 1’-position to reflect the greater loss of electron density at this position in the transition state.8,11
Figure 1.
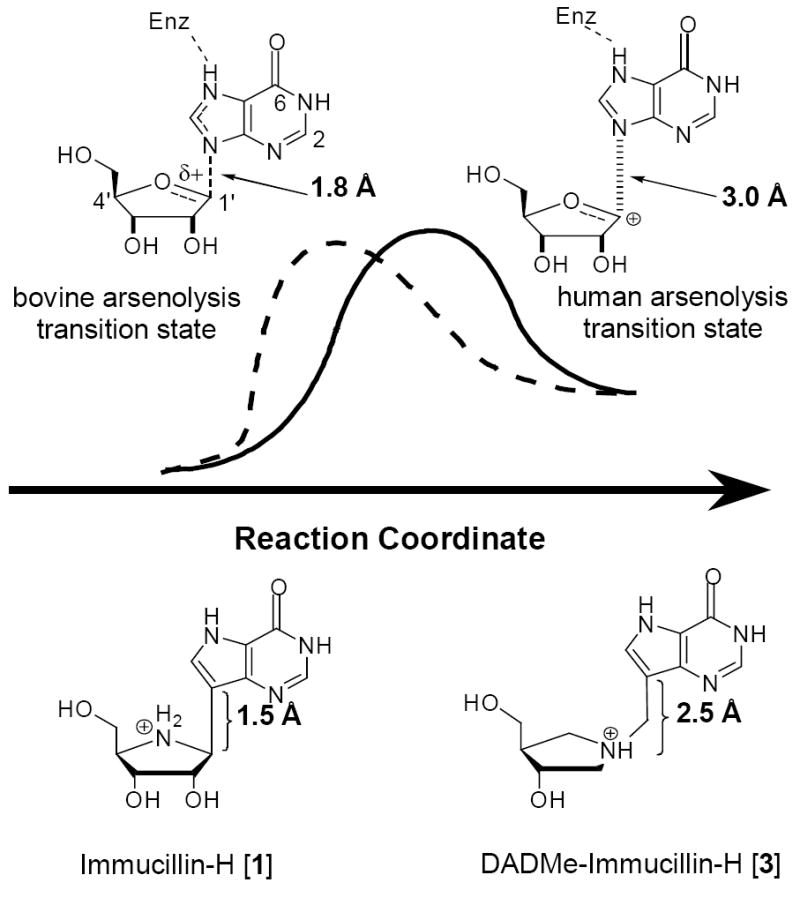
Transition state geometry for the arsenolysis of inosine by PNPs compared to analogs of the bovine [1] and human [3] transition states. 12
These transition state analogues have been found to be potent inhibitors with dissociation constants (Kd) in the picomolar range. In order to evaluate if the efficacy of these inhibitors is from their ability to differentially mimic the different transition states, these and other inhibitors were synthesized by methods described earlier and tested against both enzymes (Table 1).9,11
Table 1.
Dissociation Constants for Transition-State Analogue Inhibitors of Purine Nucleoside Phosphorylase.13
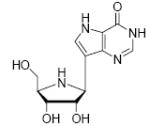
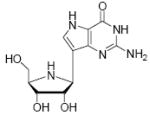
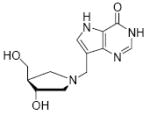
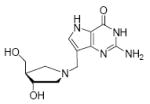
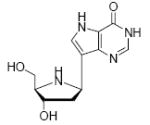
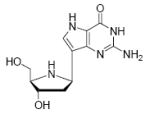
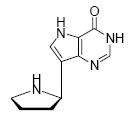
Binding of the transition state analogue inhibitors is consistent with inhibitor mimicry of the proposed transition states. Bovine PNP, which has an earlier transition state, binds more tightly to Immucillin-H than to DADMe-Immucillin-H, with dissociation constants of 23 pM and 110 pM, respectively. HsPNP has a later transition state and binds DADMe-Immucillin-H more tightly than Immucillin-H, with Kd values of 16 pM and 56 pM, respectively. This inhibition pattern holds true for Immucillin-G [2] and DADMe-Immucillin-G [4]. To examine if absence of the 2’-hydroxyl group caused the increased binding affinity between HsPNP and the DADMe compounds, 2’-deoxy-Immucillin-H [5] and 2’-deoxy-Immucillin-G [6] were examined with both enzymes. These inhibitors bound less tightly to both enzymes than their 2’-hydroxyl analogues and showed no discrimination between BtPNP and HsPNP.
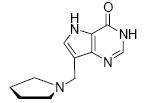
To further examine the contribution of pyrrolidine ring geometry and hydroxylation, 7-(pyrrolidin-2-yl)-3H-pyrrolo[3,2-d]pyrimidin-4(5H)-one [7] and 7-(pyrrolidin-1-yl-methyl)-3H-pyrrolo[3,2-d]pyrimidin-4(5H)-one [8] were tested as inhibitors of BtPNP and HsPNP. These inhibitors, though they have nanomolar dissociation constants, clearly demonstrate that the differences in inhibitor geometry between these two compounds is sufficient to result in binding affinity changes, where the Immucillin analogue [7] has a greater binding affinity for BtPNP and the DADMe-Immucillin analogue [8] binds more tightly to HsPNP. The differential binding of these inhibitors on the basis of their different transition state structures allows compounds 1 - 8 to be used as tools to derive insight into the relative transition state position of uncharacterized ribosyl transferases.
Although 1 - 4 are powerful inhibitors, it is not possible to generate chemically stable analogues that perfectly mimic unstable transition states. Human and bovine PNPs provide a rate enhancement of approximately 1012-fold over the uncatalyzed reaction. Therefore, since the Kd for inosine is approximately 10-5 M, the predicted binding affinity for a transition state analogue with perfect mimicry would be 10-17 M.3 The best inhibitor of Table 1 is 4 with HsPNP to give a Kd of 7 × 10-12 M, which corresponds to 7 × 105-fold weaker binding than a perfect transition state mimic. Compound 4 binds 5 × 106-fold tighter than substrate, thereby utilizing over half of the potential binding energy afforded by the enzymatic rate acceleration (kcat/knon) and capture of transition state features. Physiological experiments in mice with compound 3 suggest that its residence time on PNP in erythrocytes is in excess of tissue lifetime,16 thus capture of additional transition state binding energy is unnecessary for inhibitor design purposes.
The simplified inhibitor compound 8, though having higher Kd values than the other compounds in Table 1, has less than 350-fold higher Kd value than 3. DADMe-Immucillin-H [3] is currently being evaluated for T-cell immunosuppression and is in phase I clinical trials (http://www.biocryst.com/pipeline.htm). Compound 8 is especially novel in that it has no stereogenic centers, but its geometric similarity to the HsPNP transition state permits preferential binding to HsPNP.
The geometric differences between the Immucillins and the DADMe-Immucillins is sufficient to distinguish between the transition states of the bovine and human enzymes, even with the removal of all hydroxyl and hydroxylmethyl groups from the hydroxyl-pyrrolidine. These compounds can distinguish between two enzymes which have 87% sequence identity and have totally conserved active site residues, both in identity and position in the catalytic site.17,18 The ability to distinguish between enzymes with such great homology highlights the power of transition state determination and the subsequent synthesis and use of transition state analogues.
Supplementary Material
Synthetic methods, spectroscopic and analytical data for compounds 7 and 8. Assay conditions for Kd determination for Hs and BtPNPs. This material is available free of charge via the Internet at http://pubs.acs.org.
Acknowledgments
This work was supported by the NIH Research Grants No GM41916 and AI49512.
References
- 1.Stoeckler JD. Purine Nucleoside Phosphorylase: A Target for Chemotherapy. In: Glazer RJ, editor. Developments in Cancer Chemotherapy. CRC Press; Boca Raton: 1984. pp. 35–60. [Google Scholar]
- 2.Bantia S, Kilpatrick JM. Curr Opin Drug Discov Devel. 2004;7:243–247. [PubMed] [Google Scholar]
- 3.Schramm VL. Method Enzymol. 1999;308:301–355. doi: 10.1016/s0076-6879(99)08015-5. [DOI] [PubMed] [Google Scholar]; Schramm VL. Acc Chem Res. 2003;36:588–596. doi: 10.1021/ar0200495. [DOI] [PubMed] [Google Scholar]; Wolfenden R. Ann Rev Biophys Bioeng. 1976;5:271–306. doi: 10.1146/annurev.bb.05.060176.001415. [DOI] [PubMed] [Google Scholar]
- 4.Rodgers J, Femec DA, Schowen RL. J Am Chem Soc. 1982;104:3263–3268. [Google Scholar]
- 5.Cleland WW. Method Enzymol. 1982;87:625–641. doi: 10.1016/s0076-6879(82)87033-x. [DOI] [PubMed] [Google Scholar]
- 6.Berti PJ. Method Enzymol. 1999;308:355–397. doi: 10.1016/s0076-6879(99)08016-7. [DOI] [PubMed] [Google Scholar]
- 7.Kline PC, Schramm VL. Biochemistry. 1993;32:13212–13219. doi: 10.1021/bi00211a033. [DOI] [PubMed] [Google Scholar]
- 8.Lewandowicz A, Schramm VL. Biochemistry. 2004;43:1458–1468. doi: 10.1021/bi0359123. [DOI] [PubMed] [Google Scholar]
- 9.Furneaux RH, Tyler PC. J Org Chem. 1999;64:8411–8412. doi: 10.1021/jo990903e. [DOI] [PubMed] [Google Scholar]
- 10.Miles RW, Tyler PC, Furneaux RH, Bagdassarian CK, Schramm VL. Biochemistry. 1998;37:8615–8621. doi: 10.1021/bi980658d. [DOI] [PubMed] [Google Scholar]
- 11.Evans GB, Furneaux RH, Lewandowicz A, Schramm VL, Tyler PC. J Med Chem. 2003;46:5271–5276. doi: 10.1021/jm030305z. [DOI] [PubMed] [Google Scholar]
- 12.The arsenate, which is only found in van der Waals conctact with the ribose in both transition states, was not displayed for simplicity of visualizing the ribose-base geometry changes between the two enzymes.
- 13.Inhibition constants are reported from the indicated literature value with the least error. If no reference is given for a value it is previously unreported. Dissociation constants represent values corresponding to slow-onset, tight-binding inhibition, except for compound 7 for HsPNP and compounds 7 and 8 for BtPNP. Values shown here are final equilibrium dissociation constants.
- 14.Evans GB, Furneaux RH, Lewandowicz A, Schramm VL, Tyler PC. J Med Chem. 2003;46:3412–3423. doi: 10.1021/jm030145r. [DOI] [PubMed] [Google Scholar]
- 15.Lewandowicz A, Taylor Ringia EA, Ting L-M, Kim K, Tyler PC, Evans GB, Zubkova OV, Mee S, Painter GP, Lenz DH, Furneaux RH, Schramm VL. J Biol Chem. 2005;280:30320–30328. doi: 10.1074/jbc.M505033200. [DOI] [PubMed] [Google Scholar]
- 16.Lewandowicz A, Tyler PC, Evans GB, Schramm VL. J Biol Chem. 2003;278:31465–31468. doi: 10.1074/jbc.C300259200. [DOI] [PubMed] [Google Scholar]
- 17.Fedorov A, Shi W, Kicska G, Fedorov E, Tyler PC, Furneaux RH, Hanson JC, Gainsford GJ, Larese JZ, Schramm VL, Almo SC. Biochemistry. 2001;40:853–860. doi: 10.1021/bi002499f. [DOI] [PubMed] [Google Scholar]
- 18.Shi W, Lewandowicz A, Tyler PC, Furneaux RH, Almo SC, Schramm VL. unpublished data. Pdb file name 1RT9. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Synthetic methods, spectroscopic and analytical data for compounds 7 and 8. Assay conditions for Kd determination for Hs and BtPNPs. This material is available free of charge via the Internet at http://pubs.acs.org.