

Abstract
Cluster of differentiation antigen 4 (CD4), the T lymphocyte antigen receptor component and human immunodeficiency virus coreceptor, is down-modulated when cells are activated by antigen or phorbol esters. During down-modulation CD4 dissociates from p56lck, undergoes endocytosis through clathrin-coated pits, and is then sorted in early endosomes to late endocytic organelles where it is degraded. Previous studies have suggested that phosphorylation and a dileucine sequence are required for down-modulation. Using transfected HeLa cells, in which CD4 endocytosis can be studied in the absence of p56lck, we show that the dileucine sequence in the cytoplasmic domain is essential for clathrin-mediated CD4 endocytosis. However, this sequence is only functional as an endocytosis signal when neighboring serine residues are phosphorylated. Phosphoserine is required for rapid endocytosis because CD4 molecules in which the cytoplasmic domain serine residues are substituted with glutamic acid residues are not internalized efficiently. Using surface plasmon resonance, we show that CD4 peptides containing the dileucine sequence bind weakly to clathrin adaptor protein complexes 2 and 1. The affinity of this interaction is increased 350- to 700-fold when the peptides also contain phosphoserine residues.
INTRODUCTION
Efficient endocytosis of cell surface glycoproteins through clathrin-coated vesicles requires the presence of endocytosis signals (Pelchen-Matthews et al., 1991; Kirchhausen et al., 1997). These signals interact with adaptor proteins that couple the glycoproteins to clathrin lattices. Two types of endocytic adaptor have been identified to date; 1) adaptor protein complex 2 (AP-2)1 complexes, which have been implicated in the endocytosis of cell surface proteins carrying either YXXØ (where X is any amino acid, and Ø is a large hydrophobic amino acid) or dileucine-containing endocytosis signals (Trowbridge et al., 1993; Kirchhausen et al., 1997); and 2) the nonvisual arrestins, which facilitate the internalization of certain seven-transmembrane-domain G-protein–coupled receptors (Ferguson et al., 1996a,b; Goodman et al., 1996). The mechanisms through which these proteins recognize and bind sorting signals are not well understood. Here we describe our studies of the dileucine-containing endocytosis signal in cluster of differentiation antigen 4 (CD4) and its interaction with the AP-2 and AP-1 adaptor complexes.
CD4 is a type I integral membrane glycoprotein expressed primarily on the surfaces of major histocompatibility complex (MHC) class II restricted T lymphocytes, thymocytes, and cells of the macrophage–monocyte lineage. This protein undergoes endocytosis in clathrin-coated vesicles; however, its internalization is tightly regulated (reviewed in Marsh and Pelchen-Matthews, 1996). In T cells, association of CD4 with the Src family protein tyrosine kinase p56lck prevents CD4 association with coated pits and endocytosis (Pelchen-Matthews et al., 1992). After T cell activation by antigen, antibody-mediated cross-linking of cell surface proteins, or treatment of cells with phorbol esters (PEs), CD4 dissociates from p56lck and undergoes endocytosis (Pelchen-Matthews et al., 1992, 1993). In CD4+ve cells, which lack p56lck (e.g., monocytes and macrophages), CD4 is constitutively internalized, although the rates of internalization are increased when the cells are treated with PEs (Pelchen-Matthews et al., 1991). Significantly, the endocytic properties of CD4 observed on tissue culture cells are also seen with primary cells ex vivo (Pelchen-Matthews et al., 1998).
The endocytic properties of CD4 in the absence of p56lck have been studied in most detail in HeLa and NIH 3T3 cells stably transfected with human CD4 (Pelchen-Matthews et al., 1989). In these cells CD4 undergoes constitutive endocytosis at rates of ∼4% of the cell surface pool per minute. This endocytosis is dependent on the presence of the cytoplasmic domain of the molecule and is signal mediated. Removal of the cytoplasmic domain reduces the uptake rate to 1%/min. The truncated molecule is neither included in nor excluded from coated pits and is internalized through bulk flow endocytosis (Pelchen-Matthews et al., 1991), which we subsequently referred to as basal endocytosis. After PE treatment, CD4 undergoes rapid internalization at rates up to 10–12%/min. This increased internalization is due to a three- to fourfold increase in CD4 association with coated pits (Pelchen-Matthews et al., 1991). The cytoplasmic domain of CD4 has no tyrosines, but it does contain a pair of leucine residues (L413 and L414 in human CD4) that are required for PE-induced down-modulation (Shin et al., 1990). In addition, the phosphorylation of adjacent serine residues (S408 and S415) has been implicated in down-modulation (Shin et al., 1991). These serines are phosphorylated in PE-treated cells, and substitution of the serines reduces PE-induced down-modulation (Shin et al., 1990, 1991). Together these data suggest that phosphorylation may be required for rapid endocytosis of CD4 and may regulate the interaction of CD4 with clathrin adaptors.
CD4 down-modulation is a multistep process that involves endocytosis, endosomal sorting, and degradation (Pelchen-Matthews et al., 1993). Here we have used a series of CD4 mutants to investigate whether phosphorylation of the serine residues adjacent to the dileucine sequence is required for clathrin adaptor complex binding and endocytosis. The results indicate that the dileucine sequence is essential for endocytosis, but that the endocytosis signal is only active when S408 and/or S415 are phosphorylated. The motifs for constitutive and PE-induced endocytosis differ, with phosphorylation of S408 being essential for constitutive endocytosis, whereas phosphorylation of S408 and/or S415 allows PE-induced internalization. Surface plasmon resonance (SPR) analysis demonstrated that peptides encoding a portion of the CD4 cytoplasmic domain including L413, L414, S408, and S415 show weak binding to purified AP-2 adaptors. However, peptides in which S408 and/or S415 are phosphorylated show up to 700-fold increase in binding affinity. Significantly, constitutive endocytosis and adaptor binding are not observed when the serine residues are substituted with glutamic acid, indicating that negative charges cannot substitute for phosphoserine in this signal. Together the data indicate that 1) the endocytosis signal in CD4 involves a dileucine sequence that is only active when adjacent serine residues are phosphorylated, and 2) this phosphoserine- and dileucine-containing signal is recognized by clathrin adaptor complexes. We also observed that some CD4 constructs lacking endocytosis signals were internalized relatively efficiently in PE-treated cells. This suggested that PE treatment may also modify adaptor complexes, or other components of the endocytic machinery, to enhance CD4 recruitment into coated pits.
MATERIALS AND METHODS
Reagents and Cells
All tissue culture reagents were purchased from Life Technologies (Paisley, United Kingdom). Other chemicals were from Sigma (Poole, United Kingdom) unless indicated otherwise, and radioactive reagents were from Amersham (Little Chalfont, United Kingdom). The murine anti-human CD4 mAbs Q4120, 4, and 19 were all immunoglobulin G1. Q4120 (Healey et al., 1990) was obtained from Q. Sattentau through the Medical Research Council AIDS Reagents Program (National Institute for Biological Standards and Control, South Mimms, Potters Bar, United Kingdom); antibodies 4 and 19 (Endres et al., 1996) were provided by J. Hoxie (University of Pennsylvania, Philadelphia, PA). Q4120 was radiolabeled using 125I-3-(p-hydroxyphenyl)-propionic acid N-hydroxy-succinimide ester (Bolton and Hunter reagent) as described (Pelchen-Matthews et al., 1998). Specific activities were typically 300–600 Ci/mmol.
HeLa cells were maintained in Dulbecco’s modified Eagle’s medium containing 4% FCS, 2 mM glutamine, 100 U/ml penicillin, and 0.1 mg/ml streptomycin.
Generation and Expression of CD4 Mutants
CD4 mutants made for this study are detailed in Figure 1. The truncation mutants, S to E and L to A substitutions, were made by oligonucleotide-directed, site-specific mutagenesis using the dut/ung phagemid system (Bio-Rad, Hemel Hempstead, United Kingdom). Other mutants were made using the QuikChange site-directed mutagenesis kit (Stratagene, Cambridge, United Kingdom). Mutagenesis reactions were performed using the oligonucleotides and templates described in Table 1.
Figure 1.

CD4 and CD4 cytoplasmic domain mutants used in this study. The full-length cytoplasmic domain is illustrated for wt CD4 commencing at residue R396. Single amino acid substitutions at S408, L413, L414, and S415 are marked in bold.
Table 1.
Oligonucleotides used for site-directed mutagenesis
| Construct | Oligonucleotide |
|---|---|
| K418 | 5′-AGGGCACTGGCATCACTTCTTCTCACT |
| R412 | 5′-CTTCTCACTGAGTCATCTCTTGATCTG |
| H399 | 5′-CTTGGCGCCTTCAGTGCCGGCAC |
| K418 S408A | 5′-GAGCGGATGGCACAGATCAAG |
| and 5′-CTTGATCTGTGCCATCCGCTC | |
| K418 S415A | 5′-AGACTCCTCGCGAGAAGAAG |
| and 5′-CTTCTTCTCTGCGAGGAGTCT | |
| K418 S408A/ S415A | As for K418 S415A |
| K418 S408E | 5′-TCTCTTGATCTGCTCCATCCGCTCTGC |
| K418 S415E | 5′-CTTCTTCTCCTCGAGGAGTCTCTT |
| K418 S408E/ S415E | As for K418 S415E |
| K418 L413A/ L414A | 5′-CTTCTTCTCACTGGCGGCTCTCTTGAT |
| K418 L413V | 5′-CAGATCAAGCGAGTACTCAGTGAGAAGAAG |
| and 5′-CTTCTTCTCACTGAGTACTCGCTTGATCTG | |
| K418 L413V/ | 5′-CAGATCAAGAGAGTGGTCAGTGAGAAGAAG |
| L414V | and 5′-CTTCTTCTCACTGACCACTCTCTTGATCTG |
All CD4 constructs were generated and expressed in the eukaryotic expression vector pSG5 (Stratagene). Sequences of constructs were checked using either Sequenase version 2.0 T7 DNA polymerase (Amersham) or the Applied Biosystems Prism dye terminator cycle sequencing ready reaction kit (Perkin Elmer-Cetus, Warrington, United Kingdom). Two oligonucleotides were used for sequencing, one directed to the 3′ untranslated sequence of CD4 (5′-GTCCGCAGGCAGCAAACGCGG-3′) and the other to the extracellular domain of the protein (5′-AACCCTGAGGCGGGGATGTGG-3′).
HeLa cells were transfected by electroporation. Plasmids encoding the CD4 constructs were transfected together with pSV2-Neo at a plasmid ratio of pSG5-CD4:pSV2-Neo of 10:1. Resistant colonies were selected in medium containing 1 mg/ml Geneticin sulfate (Life Technologies) and screened by immunofluorescence using Q4120 as described (Signoret et al., 1997). DNA was prepared from the stable lines as described (Strauss, 1987), and the oligonucleotides 5′-CCACATCCCCGCCTCAGGGTT-3′ and 5′-GTCCGCAGGCAGCAAA-CGCGG-3′ were used to reisolate the transfected CD4 cDNAs by PCR. The PCR products were sequenced using the Applied Biosystems Prism system.
Measurement of CD4 Endocytosis
Endocytosis of CD4 was measured using 125I-Q4120 essentially as described (Pelchen-Matthews et al., 1989). Cells were seeded in 16-mm wells, in 24- or 4-well plates, and grown to confluence over 2 d. The cells were washed and labeled with 0.3 nM 125I-Q4120 in BM (RPMI 1640 medium containing 0.2% BSA and buffered to pH 7.4 with 10 mM HEPES) for 2 h at 4°C. After labeling, unbound antibody was washed away, and the cells were either held on ice or warmed to 37°C for various times in the presence or absence of 100 ng/ml phorbol 12-myristate 13-acetate (PMA). Cells were warmed rapidly by immersion of plates in a large volume of prewarmed medium. After incubation at 37°C, the cells were cooled by immersion in 4°C BM and either harvested directly using 0.2 M NaOH (to give total cell-associated 125I-Q4120 activity), or after treatment with pH 2.0 medium to remove the cell surface 125I-Q4120 (to give the intracellular 125I-Q4120 activity). Endocytosis was calculated as the proportion of acid-resistant to total cell-associated radioactivity for each time point. A background value for acid-resistant counts on cells maintained at 4°C (∼200 cpm) was subtracted from each time point. Endocytosis rates were calculated by linear regression over the first 4 min after warmup or, for PMA-treated cells, after the 1-min lag phase (see below).
Fluid Phase Endocytosis
Fluid phase endocytosis was measured using HRP (type II) as described (Griffiths et al., 1989). Cells, plated on 6-cm plates 2 d before the assay, were washed twice with 37°C BM and incubated in BM containing 5 mg/ml HRP at either 4 or 37°C. At the indicated times the cells were placed on ice and washed extensively with 4°C PBS. Cells were recovered in 2.5 mg/ml trypsin/EDTA, washed twice in 10 ml of 4°C PBS containing 0.5 mg/ml soya bean trypsin inhibitor, and lysed in ice-cold PBS containing 0.5% Triton X-100. HRP was quantitated using o-dianisidine in a microtiter plate assay (Pelchen-Matthews et al., 1991). Cell protein was measured using bicinchoninic acid (Pierce and Warriner, Chester, United Kingdom) as described (Smith et al., 1985).
Phosphate Labeling and CD4 Immunoprecipitation
Cells were cultured on 6-cm plates for 2 d before labeling. The cells were washed twice with phosphate-free medium and equilibrated with 0.5 mCi of [32P]orthophosphoric acid for 4 h. The cells were incubated for 3 min in 37°C BM with or without 100 ng/ml PMA and then washed three times with 4°C PBS. Cell lysates were prepared by incubating the cells for 10 min in 4°C lysis buffer (20 mM Tris-HCl, pH 8.0, containing 0.5% NP40, 150 mM NaCl, 2 mM EDTA, protease inhibitors (1 mM PMSF and 10 μg/ml chymostatin, leupeptin, antipain, and pepstatin), and phosphatase inhibitors (10 mM sodium pyrophosphate, 50 mM sodium fluoride, and 1 mM orthovanadate). The lysates were recovered by scraping, and the detergent-insoluble material was removed by centrifugation in a benchtop microfuge (13,000 rpm for 20 min at 4°C).
Before immunoprecipitation, the lysates were “precleared” by incubation for 30 min with 5 μl of a nonspecific rabbit serum and 20 μl of protein A-Sepharose CL-4B (Pharmacia, St. Albans, United Kingdom). The beads were removed by centrifugation (13,000 rpm for 1 min at 4°C), and the lysate was precleared twice more in the presence of 2.5 μl of nonspecific rabbit serum or beads alone. The final “preclears” were analyzed together with the immunoprecipitates. To immunoprecipitate CD4, lysates were mixed for 2 h, or overnight at 4°C, with protein A-Sepharose CL-4B covalently cross-linked to antibodies 4 and 19. The beads were collected by centrifugation and washed four times with lysis buffer and once in mixed micelle buffer (10 mM Tris-HCl, 0.3 M NaCl, 0.1% SDS, 0.05% Triton X-100, and 0.02% NaN3). The precipitates were eluted with nonreducing sample buffer, separated on 10% SDS-polyacrylamide gels, and transferred to nitrocellulose. The blots were exposed to Kodak (Rochester, NY) X-OMAT AR film for autoradiography. Levels of 32P incorporation were determined using a Bio-Rad GS250 Molecular Imager and/or by excision of the labeled bands and Cerenkov counting. To quantify the amount of CD4 precipitated in each lane, the transfers were immunoblotted with Q4120. Blots were blocked for 2 h at room temperature using 10% dried skimmed milk powder in PBS containing 0.1% Tween 20 and then incubated in Q4120 diluted 1:2000 in PBS and 0.1% Tween 20 for 1 h. After washing, the blots were probed with peroxidase-conjugated goat anti-mouse antibody (Pierce and Warriner), developed using ECL (Amersham) and quantified using the Bio-Rad Molecular Imager.
Preparation of Clathrin Adaptors
AP-1 and AP-2 clathrin adaptor complexes were purified from pig brain as described (Höning et al., 1997). Briefly, adaptors were released from cell membranes by treatment with 0.5 M Tris, pH 7.8, and purified by gel filtration on Superose-6 by fast-performance liquid chromatography (Pharmacia). AP-1 was separated from AP-2 by hydroxyapatite chromatography. The purity of AP-1 and AP-2 was confirmed by Coomassie Blue staining fractions after SDS-PAGE and by Western blotting using AP-1–specific (100/3) and AP-2–specific (100/2) mAbs (Sigma). Before SPR analysis AP-1 and AP-2 preparations were centrifuged for 15 min at 275,000 × g to remove any aggregated material. The supernatants were analyzed by gel filtration using a SMART system (Pharmacia) both before and after SPR analysis. No adaptor aggregation was detected under the experimental conditions used.
Peptide Synthesis
Peptides encoding the cytoplasmic domain of human CD4 from A398 to K418 (see Table 3) were synthesized by Chiron Technologies (Meltek Scientific, Feltham, United Kingdom). The peptides were synthesized with biotin at the N terminus to facilitate coupling for SPR analysis. Phosphorylated peptides were synthesized using phosphoserine instead of serine. The phosphorylation of these peptides was stable for >7 d in the buffers and conditions used for SPR analysis, as determined by mass spectroscopy.
Table 3.
SPR analysis of CD4 peptide–adaptor complex binding
| CD4 peptide | Sequence |
KD (μM)
|
|
|---|---|---|---|
| AP-1 | AP-2 | ||
| wt | Biotin-RHRRRQAERMSQIKRLLSEK | ND | 92 |
| L413A/L414A | Biotin-RHRRRQAERMSQIKRAASEK | ND | ND |
| S408 | Biotin-RHRRRQAERMSQIKRLLSEK | 0.60 | 0.12 |
| S415 | Biotin-RHRRRQAERMSQIKRLLSEK | 0.76 | 0.35 |
| S408/S415 | Biotin-RHRRRQAERMSQIKRLLSEK | 0.52 | 0.08 |
| S408/L413A/L414A | Biotin-RHRRRQAERMSQIKRAASEK | ND | 82 |
| S408/S415/L413A/L414A | Biotin-RHRRRQAERMSQIKRAASEK | 46 | 12 |
| S408E/L413A/L414A | Biotin-RHRRRQAERMEQIKRAASEK | 86 | ND |
CD4 cytoplasmic domain peptides were immobilized on streptavidin-coated sensor chips at similar densities (variation, <8%). Purified AP-1 and AP-2 complexes were used at concentrations ranging from 50 to 500 nM. The equilibrium constant (KD = the association rate [ka]/dissociation rate [kd]) was calculated from the sensorgrams (see Fig. 7) as described in MATERIALS AND METHODS. Phosphoserine residues are indicated in underlined type; ND, reactions in which the interactions were too weak to determine the binding constants.
SPR Analysis
The interaction of CD4 peptides with AP-1 and AP-2 adaptor complexes was analyzed by SPR (Jonsson et al., 1991) using a BIAcore-2000 biosensor (BIAcore, Uppsala, Sweden). The peptides were coupled via their N-terminal biotin moiety to a streptavidin-coated SA5 sensor chip according to the manufacturer’s instructions.
All interaction experiments were performed with buffer A (20 mM HEPES-NaOH, pH 7.0, 150 mM NaCl, 10 mM KCl, 2 mM MgCl2, and 0.2 mM DTT) at a flow rate of 20 μl/min unless otherwise stated. Purified adaptors were used at 50–500 nM in buffer A. Association for 2 min was followed by dissociation for 2 min, during which buffer A alone was perfused. A short pulse injection (15 s) of 20 mM NaOH and 0.5% SDS was used to regenerate the sensor chip surface after each experimental cycle. The peptide-derivitized sensor chip remained stable and retained its specific binding capacity for >100 experimental cycles of association–dissociation and regeneration.
Determination of Kinetic Rate Constants
The rate constants (ka for association and kd for dissociation) and the equilibrium constant (KD = kd/ka) for the interaction of the CD4 peptides and purified adaptors were calculated using the evaluation software of the BIAcore-2000 (version 3.0) assuming first-order kinetics A + B = AB. The mathematical models used are described in detail elsewhere (Schuck and Minton, 1996; Schuck, 1997). Association was determined 15–20 s after switching to the adaptor solution to avoid distortions caused by injection and mixing. The dissociation rate constants were determined 5–10 s after switching to buffer flow. After a dissociation phase of ∼30 s, further dissociation of adaptors was slow. The model calculates the ka and the steady-state response level Req by fitting data to the equation:
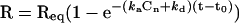 |
where t is time in seconds; Req is the steady-state response level; and C is the molar concentration of adaptors in the injection solution. The steric interference factor N, which describes the valency of the interaction between the adaptors and the CD4 peptides, was set to 1. The kd was determined by fitting data to the equation:
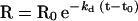 |
where R0 is the response level at the beginning of the dissociation phase. This model, which has been applied to other adaptor–cytoplasmic domain interactions (Heilker et al., 1996), is described in more detail elsewhere (Karlsson et al., 1991; O’Shannessy et al., 1993). It should be noted that these models allow the determination of rate constants without reaching equilibrium during the experimental cycle.
RESULTS
The CD4 Endocytosis Signal Is Located between Residues 399 and 418
To examine the relationship between serine phosphorylation and CD4 recruitment into endocytic clathrin-coated vesicles, we expressed CD4 and CD4 cytoplasmic domain mutants in HeLa cells. HeLa cells do not express p56lck and allow the endocytic properties of CD4 to be examined unfettered. To identify the location of the endocytosis signal(s) in CD4 we first used site-directed mutagenesis to replace amino acids R400, L413, and T419 with stop codons and to produce the truncation mutants H399, R412, and K418 (Figure 1). The constructs were transfected into HeLa cells by electroporation, and stable CD4-positive clones were selected. CD4 expression was monitored by immunofluorescence and SDS-PAGE. All three truncation mutants were expressed on the cell surface and, as expected, demonstrated increased mobility in SDS-PAGE compared with wild-type (wt) CD4 (Figure 2B).
Figure 2.

Endocytosis and phosphorylation of CD4 truncations mutants. (A) HeLa cells stably expressing CD4 or CD4 truncation mutants (Figure 1) were labeled with 0.3 nM 125I-Q4120 at 4°C and then warmed to 37°C in binding medium (open symbols) or medium containing 100 ng/ml PMA (solid symbols) for the indicated times. The total and acid-resistant intracellular radioactivities were determined for each time point as described in MATERIALS AND METHODS. Data from one representative experiment are illustrated, and each point represents the mean ± SD for duplicate samples. □ and ▪, wt CD4; ○ and ●, K418; ▵ and ▴, R412; ⋄ and ♦, H399. (B) Cells were labeled with [32P]orthophosphate (as described in MATERIALS AND METHODS) and incubated in medium with (+) or without (−) 100 ng/ml PMA for 3 min. The cells were rapidly cooled to 4°C and lysed, and the CD4 molecules were immunoprecipitated, separated by SDS-PAGE, and transferred to nitrocellulose paper. CD4 protein was visualized and quantitated by Western blot, and the 32P-activity was determined on the same blots using a phosphorimager. The positions of the molecular weight standards are shown; p, preclear lane (see MATERIALS AND METHODS). Note that the K418 32P lanes illustrated here have been exposed for shorter times than the other panels.
An assay based on the internalization of prebound 125I-Q4120 was used to measure CD4 uptake. In this assay, the amount of antibody internalized in the first 10 min after warming cells to 37°C gives a direct measure of the rate of CD4 endocytosis (Pelchen-Matthews et al., 1989). To ensure that all cells were treated identically, plates of cells were warmed by immersion in 37°C medium. This allowed a more rapid rate of temperature change than previous procedures, and, as a consequence, we observed slightly faster CD4 uptake (cf. Pelchen-Matthews et al., 1989). The rates of internalization were calculated for the first 4 min after warmup. Under these conditions full-length CD4 was internalized at ∼5% of the cell surface pool per minute (Figure 2A and Table 2). K418 was internalized at a similar rate. By contrast, R412 and H399 were internalized at rates of 1–2%/min. The internalization of H399 is considered to occur through basal endocytosis (Pelchen-Matthews et al., 1991). The faster uptake of wt CD4 and K418 reflects constitutive signal-dependent endocytosis. R412 consistently showed slightly faster internalization rates than H399. Whether this indicates the presence of a weak endocytosis signal in R412 or reflects some nonspecific interaction of the truncated cytoplasmic domain with the endocytosis machinery is unclear.
Table 2.
Effects of phorbol ester on CD4 endocytosis and phosphate incorporation
| Construct | Endocytosis ratea
|
Relative phosphate incorporationb
|
||||
|---|---|---|---|---|---|---|
| −PMA (%/min) | +PMA (%/min) | +PMA:−PMA | −PMA | +PMA | +PMA:−PMA | |
| wtCD4 | 5 ± 0.9 | 19.2 ± 3.3 | 3.8 | 1 | 6.1 | 6.1 |
| n = 11 | n = 8 | n = 7 | n = 7 | |||
| K418 | 4.2 ± 0.9 | 18.2 ± 3.4 | 4.3 | 1.2 | 6.3 | 5.2 |
| n = 3 | n = 3 | n = 7 | n = 7 | |||
| R412 | 2.4 ± 0.2 | 2.3 ± 0.4 | 1 | 0.9 | 4.4 | 4.9 |
| n = 3 | n = 3 | n = 1 | n = 1 | |||
| H399 | 1.2 ± 0.4 | 1.0 ± 0.2 | 1 | NDc | ND | |
| n = 10 | n = 8 | |||||
| Mutations in K418 background | ||||||
| K418 S408A/S415A | 1.7 ± 1.0 | 4.2 ± 0.2 | 2.4 | ND | ND | |
| n = 3 | n = 4 | |||||
| K418 S408E/S415E | 1.9 ± 0.8 | 4.1 ± 0.1 | 2.2 | ND | ND | |
| n = 3 | n = 3 | |||||
| K418 S408A | 1.7 ± 1.0 | 7.1 ± 1.7 | 4.1 | 0.7 | 4.0 | 5.7 |
| n = 3 | n = 3 | n = 4 | n = 4 | |||
| K418 S408E | 2.7 ± 0.5 | 13.7 ± 4.5 | 5 | 2.0 | 8.3 | 4.1 |
| n = 7 | n = 7 | n = 3 | n = 3 | |||
| K418 S415A | 2.1 ± 0.4 | 6.6 ± 1.2 | 3.1 | 0.1 | 2.4 | 24 |
| n = 3 | n = 3 | n = 1 | n = 1 | |||
| K418 S415E | 2.0 ± 0.3 | 5.0 ± 0.6 | 2.5 | 0.1 | 2.5 | 25 |
| n = 3 | n = 2 | n = 2 | n = 2 | |||
| K418 L413A/L414A | 2.2 ± 0.7 | 2.5 ± 0.4 | 1 | 1.1 | 7.1 | 6.5 |
| n = 4 | n = 3 | n = 3 | n = 3 | |||
| K418 L413V | 1.6 ± 0.4 | 1.9 ± 0.2 | 1 | 2.7 | 5.9 | 0.5 |
| n = 2 | n = 3 | n = 2 | n = 1 | |||
| K418 L413V/L414V | 1.9 ± 0.5 | 2.3 ± 0.4 | 1 | 1.7 | 7.3 | 4.4 |
| n = 2 | n = 3 | n = 1 | n = 1 | |||
Endocytosis rates were normally calculated over the first 4 min after warmup except in PE-treated cells. Because the effects of PE were observed after a lag of ∼1 min (see Fig. 2A), the rates for these cells were calculated 1–4 min after warmup. The effects of PE on CD4 endocytosis are given as the ratio of the rates +PMA:−PMA.
Phosphate incorporation for each construct (− and +PMA) is compared with wt CD4 immunoprecipitated from untreated cells (see MATERIALS AND METHODS). Wild-type CD4 is set to 1. n, number of experimental data sets used in these calculations.
ND, not detected.
As previously described (Pelchen-Matthews et al., 1989, 1991), treatment of wt CD4 expressing cells with PMA rapidly induced a fourfold increase in the rate of CD4 endocytosis (Figure 2A and Table 2). This increase occurred after a lag of ∼1 min (Figure 2A), which we attribute to the time required for the activation of PKC and phosphorylation of CD4. K418 showed a similar increase in endocytosis rate. However, PMA had no effect on the endocytosis of either H399 or R412 (Figure 2A). Together these data indicate that constitutive and PE-induced CD4 endocytosis require cytoplasmic domain elements residing between the transmembrane domain and K418, and that sequences downstream of K418 are not required for CD4 endocytosis.
The observation that H399 endocytosis was not modulated by PE treatment suggested that PMA had no effect on bulk flow endocytosis in these cells. To determine this directly, we measured the effect of PMA on fluid phase endocytosis using HRP. Similar levels of HRP were accumulated over 10 min for all cell lines in the presence and absence of PMA (Figure 3). A slight increase in HRP accumulation was observed after 30 min of endocytosis in the presence of PMA, as previously reported (Pelchen-Matthews et al., 1993), and may reflect PE effects on later steps in the endocytic pathway.
Figure 3.

Effect of PMA on fluid phase endocytosis. Cells expressing CD4, K418, or H399 were incubated in BM containing HRP in the presence (filled symbols) or absence (open symbols) of 100 ng/ml PMA. At the end of the incubation period the cells were rapidly cooled to 4°C and washed extensively, and the cell-associated HRP was determined. □ and ▪, wt CD4; ○ and ●, K418; ▵ and ▴, H399.
CD4 Phosphorylation
It has been demonstrated that PEs induce rapid phosphorylation of the three serine residues in the CD4 cytoplasmic domain (S408, S415, and S431) and that this phosphorylation is necessary, but not sufficient, for CD4 down-regulation (Shin et al., 1991). Phosphorylation occurs mainly on S408, and substitution of this amino acid with alanine inhibits down-modulation (Shin et al., 1990). To study the role of phosphorylation in CD4 endocytosis specifically, cells expressing wt CD4 and truncation mutants were labeled with [32P]orthophosphate for 4 h and then incubated in BM with or without 100 ng/ml PMA for 3 min at 37°C. A 3-min time point was chosen, because we previously found that maximal clustering of CD4 in endocytic, clathrin-coated pits occurs 2–3 min after PMA addition (Pelchen-Matthews et al., 1993). Subsequently, the cells were lysed and CD4 immunoprecipitated, and the immunoprecipitates were separated by SDS-PAGE and transferred to nitrocellulose. CD4 was quantitated by Western blotting, and CD4 32P incorporation was determined on the same blots. To compare phosphate incorporation for each construct, 32P activity associated with each CD4 band was normalized for the amount of CD4 protein. The phosphate:protein ratios obtained were then compared with the phosphate:protein ratio for CD4 precipitated from cells that had not been treated with PE.
Wild-type CD4 was constitutively phosphorylated (Figure 2B). After 3 min of PE treatment, phosphate incorporation on this molecule was increased approximately sixfold. K418 and R412 were also constitutively phosphorylated and showed approximately fivefold increases in incorporation after PE treatment. As expected, H399 was not phosphorylated either constitutively or after PE treatment. Together, these data suggest that S415 phosphorylation does not require sequences downstream of K418, and S408 phosphorylation does not require sequences down stream of R412. The data also suggest that S431 is not significantly phosphorylated after PE treatment. In addition, because R412 was phosphorylated to a similar level as wt CD4 and K418 but did not undergo efficient endocytosis, the data also indicated that phosphorylation of the truncated R412 molecule is not sufficient to induce its endocytosis. Thus elements between R412 and K418 are required for CD4 internalization.
Because the 23 membrane proximal amino acids in the CD4 cytoplasmic domain are sufficient for phosphorylation and endocytosis, and the C-terminal 15 amino acids make little contribution to the total level of phosphorylation or the endocytic activity of the molecule, the K418 construct was used for site-directed mutagenesis.
Two Leucine Residues Are Essential for CD4 Endocytosis
To investigate the role of leucines 413 and 414, we generated a mutant CD4 molecule in which these residues were substituted with alanine (K418 L413A/L414A; Figure 1). This molecule showed no constitutive or PMA-induced endocytosis (Figure 4). Strikingly, the molecule was constitutively phosphorylated to the same extent as K418 (Figure 4C), and the level of phosphorylation was increased approximately sevenfold after PE treatment. In addition, we made two mutants in which either one or both leucine residues were mutated to valine (K418 L413V and K418 L413V/L414V; Figure 1). Both mutants were internalized at basal endocytosis rates (Figure 4), despite the fact that both were phosphorylated with similar or higher efficiency than K418 (Figure 4C). Thus the loss of a single methyl group from the side chain of residue 413 is sufficient to disrupt the endocytosis signal.
Figure 4.

Effects of leucine substitutions on K418 CD4 endocytosis and phosphorylation. (A and B) Endocytosis of K418 CD4 L substitutions (Figure 1) was measured as described in Figure 2A. Open symbols indicate the constitutive endocytosis of constructs in the absence of PMA, and solid symbols indicate endocytosis in the presence of 100 ng/ml PMA. (A) □ and ▪, K418; ○ and ●, K418 L413A/L414A; ⋄ and ♦, H399. (B) ▵ and ▴, K418 L413V/L414V; ○ and ●, K418 L413V. (C) Cells expressing K418 CD4 L413/L414 substitutions were labeled with [32P]orthophosphate, incubated in medium with (+) or without (−) 100 ng/ml PMA for 3 min, and CD4 immunoprecipitated as described in Figure 2B. The left six lanes of the top panel were exposed to film for twice as long as the right six lanes.
The CD4 Endocytosis Signal Requires Serine Phosphorylation
Phosphorylation of S408 in R412, and of the leucine substitution constructs discussed above (Figure 4), was not sufficient to generate an active endocytosis signal. However, the basal phosphorylation of wt CD4 and K418, together with the increased phosphorylation that accompanied the PE-induced CD4 endocytosis, suggested that phosphorylation might be necessary for efficient CD4 endocytosis. We examined the role of phosphorylation by substituting S408 and S415 with alanine (K418 S408A/S415A; Figure 1). As expected, no basal or PE-induced phosphorylation of this double mutant was observed (Figure 5). Moreover, this construct was internalized at rates similar to the basal endocytosis of H399 (1.7%/min; Figure 5 and Table 2). Thus the constitutive internalization of wt CD4 and K418 may be due in part to the constitutive phosphorylation of these molecules.
Figure 5.

Effects of serine substitutions on K418 CD4 endocytosis and phosphorylation. (A and B) Endocytosis of K418 CD4 S to A substitutions (Figure 1) was measured as described in Figure 2A. (A) Constitutive endocytosis of the constructs in the absence of PMA. (B) Endocytosis in the presence of 100 ng/ml PMA. □ and ▪, K418; ○ and ●, K418 S408A; ▿ and ▾, K418 S415A; ▵ and ▴, K418 S408A/S415A; ⋄ and ♦, H399. (C) Cells expressing K418 CD4 S to A or E (see Figure 5) substitutions were labeled with [32P]orthophosphate, incubated in medium with (+) or without (−) PMA, and CD4 immunoprecipitated as described in Figure 2B.
After PE treatment, the rate of K418 S408A/S415A endocytosis increased from 1.7 to 4.2%/min (Figure 5 and Table 2). The uptake of the R412 and H399 constructs, which do not contain leucines 413 and 414, was not affected by PE (Figure 2 and Table 2). This suggests that PE treatment may induce some modification of the endocytic machinery so that it interacts more effectively with the nascent LL-containing endocytosis signal in CD4.
Both S408 and S415 Contribute to the CD4 Endocytosis Signal
The individual contributions of S408 and S415 to CD4 endocytosis were analyzed with constructs containing single alanine substitutions (K418 S408A and K418 S415A; Figure 1). K418 S408A was internalized at 1.7%/min (Figure 5), a rate comparable to H399 and R412 (Figure 2 and Table 2). On PMA treatment the rate of K418 S408A internalization increased ∼4.5-fold to 7.1%/min. K418 S408A was constitutively phosphorylated (Figure 5), indicating that S415 could be phosphorylated in the absence of S408, although the level of constitutive phosphorylation was less than that seen for K418 (Table 2). PE treatment induced an approximately sixfold increase in phosphate incorporation.
The intermediate values for the rates of endocytosis seen with these constructs suggest that the interplay between phosphorylation and endocytosis is complex. The basal endocytosis of the S408A mutant indicates that the constitutive phosphorylation of S415 does not generate an efficient endocytosis signal. After PE treatment, however, the endocytosis and phosphorylation of S408A were increased by four- and sixfold, respectively. We suggested above that PE treatment may modify the interaction of adaptors with the nascent signal in CD4; some of the PE-induced increase in S408A uptake may be attributed to this effect. Nevertheless, in PE-treated cells the S408A construct was internalized faster than the double serine mutant (S408A/S415A), indicating that phosphorylation of S415 may activate the CD4 signal, although this activation is not as effective as phosphorylation on S408. The notion that phosphorylation of S415 may generate a weak signal was supported by in vitro SPR studies (see below).
K418 S415A was also internalized at a basal rate (2.1%/min; Figure 5). Very little constitutive phosphorylation was seen with this construct (Figure 5). PE treatment induced a threefold increase in the rate of internalization of this construct (Figure 5) and a significant increase in phosphorylation. The level of this increase was exaggerated by the low basal labeling, and the level observed was comparable to that of wt CD4 in unstimulated cells. Because the R412 construct was phosphorylated both constitutively and after PE (Figure 2), these results suggest that residues 413–418, and S415 in particular, may influence phosphorylation on S408.
Glutamic Acid Does Not Substitute for Serine Phosphorylation in CD4 Endocytosis
A consequence of phosphorylation is the addition of negative charge to a protein. In some cases phosphoserine can be mimicked by glutamic or aspartic acid (for example, see Hicke et al., 1998). To determine whether the introduction of negative charges into CD4 might generate an active endocytosis signal, S408E, S415E, and double S408E/S415E mutants were made in K418 (Figure 1). When these mutants were stably expressed in HeLa cells, cell surface expression equivalent to that of wt CD4 was observed by immunofluorescence and 125I-Q4120 binding, indicating that the substitutions did not impede delivery of the protein to the plasma membrane. K418 S408E/S415E behaved identically to the K418 S408A/S415A mutant. Basal internalization was seen (Figure 6), and PE induced an approximately twofold increase in the endocytosis rate, similar to the K418 S408A/S415A construct discussed above (Table 2).
Figure 6.

Effects of serine to glutamic acid substitutions on K418 CD4 endocytosis. Endocytosis of K418 CD4 S to E substitutions (Figure 1) was measured as described in Figure 2A. (A) Constitutive endocytosis of the constructs in the absence of PMA. (B) Endocytosis in the presence of 100 ng/ml PMA. □ and ▪, wt CD4; ▵ and ▴, K418 S408E; ○ and ●, K418 S415E; ▿ and ▾, K418 S408E/S415E; ⋄ and ♦, = H399.
Single glutamic acid substitutions exhibited some intermediate properties. S408E showed some constitutive endocytosis activity (2.7%/min; Table 2). This construct also showed relatively high constitutive phosphorylation (Figure 5). PE treatment increased the rate of S408E endocytosis approximately fivefold, and phosphate incorporation increased the rate fourfold (Figure 5). Together these data are consistent with the notion that phosphorylation on S415 can generate a weak endocytosis signal. Why this construct is phosphorylated more efficiently than S408A is unclear; possibly the presence of a negative charge at S408 may increase the efficiency of phosphorylation on S415.
S415E showed internalization kinetics similar to S415A (Table 2). In addition, as with S415A, there was little constitutive phosphorylation of S415E. S415E was phosphorylated and internalized after PE treatment (Figure 5), although the rates were slow compared with K418. Thus a negative charge at position 415 does not generate an active endocytosis signal, but, as suggested above, the residues in this domain of CD4 may influence S408 phosphorylation. Together the data indicate that glutamic acid residues do not substitute for phosphoserine in the CD4 tail and that negative charges alone are not sufficient to generate an active endocytosis or sorting signal.
SPR Analysis of CD4 Peptide–Adaptor Interactions
Previous reports have suggested that dileucine-containing endocytosis signals interact with clathrin adaptor complexes (Dietrich et al., 1997; Bremnes et al., 1998; Rapoport et al., 1998), although the site of interaction may be different from that recognized by tyrosine-containing signals (Marks et al., 1996; Rapoport et al., 1998). Attempts to coprecipitate CD4 with adaptor protein complexes either under steady-state conditions or after PE treatment were not successful. We therefore investigated the interaction of purified adaptors with CD4 cytoplasmic domain peptides using SPR. N-terminally biotinylated peptides equivalent to the cytoplasmic domain of K418 CD4 were synthesized. Peptides containing specific amino acid substitutions are indicated in Table 3. The N-terminal biotin modification facilitated coupling to a streptavidin-coated SPR chip such that the peptide was oriented with the C terminus free, as may occur on the internal face of the plasma membrane. Interactions with purified pig brain AP-2 and AP-1 adaptors were determined as described (Höning et al., 1997).
Little interaction with AP-2 was seen using a wt K418 peptide in which neither serine was phosphorylated (Figure 7) or with a peptide containing L413A/L414A substitutions (Table 3). The observed signal was comparable to that seen when adaptor complexes were passed over a nonderivatised surface. However, a significant increase in AP-2 binding was seen when a wt K418 peptide with phosphoserine at position 408 was used (Figure 7). Analysis of the association and dissociation phases of the interaction suggested a KD of 92 μM for the wt peptide (Table 3), and that this value was increased >700-fold (KD ≈ 120 nM) for the phosphorylated peptide. AP-2 binding was also increased when a peptide with phosphoserine at 415 was used, supporting the notion that phosphorylation of S415 may generate a weak endocytosis signal. This later binding was subsequently reduced to background levels when the peptide was dephosphorylated using calf alkaline phosphatase (our unpublished results). Significantly, highest affinity binding was seen with a peptide in which both serine residues were phosphorylated. Binding was dependent on the presence of leucines 413 and 414, because peptides containing alanine at these positions failed to interact with the adaptors regardless of whether phosphoserine was present at either 408 or 415 or both. Moreover, substitution of S408 with glutamic acid also failed to elicit significant binding. We also observed binding of these peptides to the AP-1 complex. Binding was on the order of three- to fourfold lower affinity than that seen for AP-2 but was also dependent on phosphorylation and the presence of the dileucine sequence. Together these data indicate that the endocytosis signal in CD4 binds the clathrin adaptor complexes AP-1 and AP-2 and that the sequence requirements for binding show a very close correlation to those required for endocytosis (see above).
Figure 7.

SPR analysis of CD4 peptide–adaptor complex binding. Biotinylated CD4 tail peptides were immobilized on an SA5 sensor surface and analyzed for binding of purified AP-2 and AP-1 adaptor complexes. The wt peptide bound both adaptor complexes with very low affinity (see Table 3); however, high-affinity adaptor binding was observed when peptides containing phosphorylated S408 were used.
DISCUSSION
Cell surface receptors, and other proteins, that traffic between the plasma membrane and endocytic compartments require signals that allow their efficient recruitment into endocytic vesicles. To date four classes of endocytosis signal have been identified that target surface proteins to clathrin-coated vesicles. First, tyrosine-based signals within FXNPXY or YXXØ motifs were initially identified in the low-density lipoprotein and transferrin receptors, respectively (Trowbridge et al., 1993). The low-density lipoprotein receptor FXNPXY motif is reported to adopt a reverse turn configuration and to bind directly to clathrin heavy chains (Kibbey et al., 1998), whereas at least some YXXØ motifs interact with the μ chains of adaptor complexes (reviewed in Kirchhausen et al., 1997; Owen and Evans, 1998). In many cases these signal are constitutively active; i.e., receptors containing the motifs undergo continuous rounds of endocytosis and recycling whether ligands are bound.
However, endocytosis signals can be regulated. For CTLA-4 at least, phosphorylation of a tyrosine can inactivate μ chain binding (Shiratori et al. 1997). By contrast, three other types of endocytosis signal can be activated by phosphorylation. The first type is signals involving pairs of hydrophobic amino acids, frequently leucines (Letourneur and Klausner, 1992), although isoleucine, methionine, or valine can substitute for one of the leucines in some circumstances (Bremnes et al., 1994; Sandoval and Bakke, 1994; Pond et al., 1995). These so-called “dileucine” signals also bind adaptor complexes through a site(s) different from that involved in YXXØ motif recognition (Marks et al., 1996; Dietrich et al., 1997; Bremnes et al., 1998; Rapoport et al., 1998). Dileucine motifs have been implicated in the endocytic activities of a number of proteins and receptors, and for some at least, the endocytic activities are regulated by phosphorylation (Shin et al., 1991; Corvera et al., 1994; Dietrich et al., 1994). A second type of regulated signal is contained in members of the family of seven-transmembrane-domain G protein–coupled receptors. For several of these receptors, in particular the β-adrenergic receptor, ligand-induced phosphorylation of serine residues in the C-terminal domain of the molecule leads to recruitment of nonvisual arrestins that both uncouple associated heterotrimeric G proteins and act as adaptors to recruit the receptor into clathrin-coated pits (reviewed in Ferguson et al., 1996a). Finally, in Saccharomyces cerevisiae ligand-induced phosphorylation of serines in the α factor receptor (Ste2p) C-terminal domain leads to ubiquitination of Ste2p and its subsequent internalization (Hicke et al., 1998). A number of mammalian cell surface receptors are also ubiquitinated after ligand binding (Strous et al., 1997), although a role in endocytosis is still to be clearly established. Nevertheless, it is apparent that in many instances endocytic signals are regulated, and this regulation occurs primarily through phosphorylation of serine residues.
Here we show that a dileucine-containing signal in CD4 is activated by phosphorylation and that this reaction allows the CD4 cytoplasmic domain sorting signal to bind clathrin adaptor complexes. Our study refines earlier work (Shin et al., 1990, 1991) in which the complex phenomenon of down-modulation was analyzed. Moreover, we have combined direct endocytosis assays with determinations of serine phosphorylation and adaptor binding assays. The results indicate a mechanism through which CD4 endocytosis is controlled by a phosphorylation-regulated interaction with clathrin adaptors. We have shown that the endocytosis signal in CD4 requires at least two components: a pair of leucine residues that are essential for endocytosis and a neighboring phosphorylated serine residue.
A pair of leucine residues is essential for CD4 endocytosis. These amino acids are absolutely conserved in all CD4 sequences published to date. Modification of either one or both of the leucines to alanine completely disrupts CD4 endocytosis regardless of whether the serines at 408 and 415 are phosphorylated. Moreover, substitution of just one of the leucines (L413) with valine, a change that removes a single methyl group from the side chain, is sufficient to disrupt the signal. The two leucine residue were also essential for in vitro adaptor binding: peptides containing leucine to alanine substitutions showed weak, or undetectable, binding to both AP-1 and AP-2 adaptors even when phosphorylated. We have not tested other hydrophobic substitutions; however, our studies with the CXCR4 chemokine receptor suggest that hydrophobic substitutions may be tolerated in some dileucine-containing signals (Signoret et al., 1998). At present we know little about the architecture of the CD4 cytoplasmic domain that is recognized by the endocytic machinery. (A nuclear magnetic resonance structure for a CD4 cytoplasmic domain peptide has been published; however this peptide was not phosphorylated, and it is unclear whether the structure represents that of the native protein [Willbold and Roesch, 1996].) The fact that the endocytosis signal in CD4 is regulated may impose a tighter sequence requirement for two leucines than constitutively active dileucine-containing signals in which other hydrophobic amino acids can be tolerated.
The second component of the signal is a phosphorylated serine residue. The CD4 cytoplasmic domain contains three serine residues that are potential targets for PKC-mediated phosphorylation. Of these, S431 is phosphorylated least efficiently (Shin et al., 1991) and has no apparent involvement in the endocytic properties of the molecule. However, both S408 and S415 influence endocytosis, and both residues undergo phosphorylation on treatment of cells with PEs. S408 appears to be phosphorylated more efficiently (Shin et al., 1991), and the increase in the level of phosphate incorporation is similar to increases in the measured rates of CD4 endocytosis. Substitution of both S408 and S415 with either alanine or glutamic acid and single S408 substitutions completely abolish constitutive CD4 endocytosis, even though the dileucine sequence is intact. In addition, PE-induced endocytosis of these constructs is reduced (see below). Although serine phosphorylation is required for CD4 internalization, phosphorylation alone is not sufficient for endocytosis, because constructs with leucine mutations and the R412 protein fail to internalize, even though they are phosphorylated efficiently. In keeping with these results, in vitro binding of adaptor complexes was seen with peptides phosphorylated on either S408 or S415, although higher-affinity binding was seen when S408 was phosphorylated. Together, the data indicate that both constitutive and PE-induced CD4 endocytosis is dependent on serine phosphorylation, the normal CD4 molecule being phosphorylated to some extent even in unstimulated cells. Only basal CD4 internalization, which occurs by bulk flow endocytosis, is independent of both phosphorylation and the dileucine sequence. The precise relationship between S408 and S415 is complex. Both residues appear to facilitate endocytosis when phosphorylated, because the single substitutions show PE-induced internalization. For this function, which involves interaction with the adaptor proteins, phosphoserine cannot be substituted with glutamic acid. However, the phosphorylation of S415 in particular may be influenced by the presence of a negative charge at position 408. In addition, residues 413–418 may modulate the phosphorylation of S408. Thus the efficacy of CD4 endocytosis is a balance between the interaction of kinases (and possibly phosphatases) that phosphorylate the cytoplasmic domain of the molecule and the subsequent recognition of the active endocytosis signal by clathrin adaptor complexes.
Why serine phosphorylation is required to activate the CD4 endocytosis signal is unclear. One notion is that phosphorylation of the CD4 cytoplasmic domain may increase the affinity of the motif for binding sites on AP-2. However, the substitution of the serine residues with glutamic acid did not generate a constitutively active endocytosis signal, indicating that the role of phosphorylation is not simply the introduction of a negative charge. This contrasts the situation in, for example, Ste2p, in which negatively charged amino acids can partially replace phosphoserine (Hicke et al., 1998). Another possibility is that the incorporation of phosphate would induce a structural constraint or conformational change in CD4 that allows the display of the dileucine sequence in a form that can interact with the adaptor complexes. Such a conformational change may indeed occur as the interaction of CD4 with p56lck is also regulated by serine phosphorylation. Under normal circumstances the cytoplasmic domain of CD4 interacts with the N-terminal domain of p56lck. This interaction involves cysteines 421 and 423 in CD4, cysteines 21 and 23 in Lck, and the coordination of a zinc ion (Huse et al., 1998). PE-induced phosphorylation of S408 triggers dissociation of this complex (Pelchen-Matthews et al., 1993), whereas a complex formed between Lck and CD4 with an S408A mutation fails to dissociate in PE-treated cells. More detailed structural studies will be required to fully understand the nature of the CD4 endocytosis signal and the role of phosphoserine and the dileucine sequence in regulating endocytosis.
The clathrin adaptor complex associated with endocytic coated vesicles is AP-2 (Robinson, 1994). Peptides encoding a portion of the cytoplasmic domain of CD4 can bind purified AP-2 complexes in vitro. The features of the peptides required for this association closely parallel the endocytic properties of CD4. Thus we believe that the interactions observed by SPR are likely to reflect events that occur within the cell. Binding is dependent on the presence of the two leucine residues and requires phosphorylation of the serine residues. The binding affinity measured by SPR is highest when S408 is phosphorylated. However, significant binding also occurs when S415 is phosphorylated, supporting the endocytosis data suggesting that a weak endocytosis signal may be generated after phosphorylation of S415. The highest affinity CD4 peptide–AP-2 interaction measured using SPR (kD = 100 nM) was similar to that recorded for a synthetic dileucine-containing sequence (Heilker et al., 1996) but is somewhat less than values determined for Y-based signals (Höning et al., 1997). At present it is unclear how closely the affinities we have measured by SPR reflect signal–adaptor interactions in the cell. Nevertheless, it is clear that the properties we observe for binding in vitro correlate well with the endocytic properties of CD4.
Significantly, we also observed the CD4 peptides binding to AP-1 complexes. This interaction appeared to be weaker than that seen for AP-2 but required similar features in the peptides, i.e., the pair of leucines and phosphoserine. The relevance of this binding is at present unclear. AP-1 is believed to function in association with clathrin-coated vesicles derived from the trans-Golgi network (Robinson, 1994). At present we have no indication that CD4 is phosphorylated during transit through the biosynthetic pathway. Indeed our studies of p56lck biosynthesis suggest that CD4 and p56lck form a complex early in the exocytic pathway and then move together to the cell surface (Bijlmakers and Marsh, unpublished results). Because phosphorylation of the CD4 cytoplasmic domain induces dissociation of p56lck, it seems unlikely that CD4 will be phosphorylated during transit through the Golgi apparatus. However, during down-modulation, internalized CD4 is delivered to early endosomes and from this site is sorted to late endosomes and lysosomes, where it is believed to be degraded (Pelchen-Matthews et al., 1993). At present we know little of the machinery responsible for this endosomal sorting step. It is possible that AP-1 plays some role. Alternatively, the recently characterized AP-3 complex, which has been implicated in sorting to late endocytic organelles (Ooi et al., 1997; Simpson et al., 1997), might function in this step. We have yet to determine whether CD4 peptides are able to bind AP-3.
A number of dileucine-containing endocytosis and sorting sequences contain one or more negatively charged amino acids three or four positions upstream of the leucine pair. The role of these residues is unclear. An aspartic acid residue appears to be important for the binding of a dileucine motif from the T cell receptor–CD3 γ subunit to AP-1 and AP-2 (Dietrich et al., 1997), although the endocytic activity of this signal is regulated by phosphorylation (Dietrich et al., 1994). By contrast, in the MHC II invariant chain, polar residues upstream of both dileucine-like signals can influence endocytosis but appear to be required for intracellular sorting events (Pond et al., 1995). Significantly, upstream negatively charged amino acids have been shown to be important for the interaction of tyrosinase and the lysosomal integral membrane protein II, two proteins that are sorted to lysosomal-type organelles, with AP-3 complexes. Moreover, the dileucine signals in these proteins do not appear to interact with AP-1 and AP-2 (Höning et al., 1998). A similar signal has been identified in the yeast t-SNARE Vam3p that mediates sorting of Vam3p to the vacuole via the AP-3–dependent pathway (Darsow et al., 1998). Whether other amino acids adjacent to the leucines play critical roles in adaptor binding is also unclear. M407 and I410 in CD4 have been implicated in PE-induced CD4 down-modulation (Shin et al., 1991), but it remains to be determined whether these residues modulate endocytosis or adaptor binding specifically. How dileucine motifs within specific sequence contexts interact with different adaptor complexes will require more detailed systematic analyses.
The binding experiments described here used purified adaptor complexes in which the two heavy chains (α/β1 and γ/β2), the medium chains (μ1 and μ2), and the small chains (ς1 and ς2) were present. The binding site(s) for dileucine-containing signals on adaptor complexes are unclear. Competition studies with overexpressed receptors indicate that dileucine-containing signals are not competed by YXXØ based signals (Marks et al., 1996). Experiments using phage display suggest that dileucine-type signals in the MHC II invariant chain interact with adaptor μ subunits (Bremnes et al., 1998), whereas labeling experiments with dileucine-containing peptides suggest an interaction with β subunits (Rapoport et al., 1998). Significantly, a phosphopeptide was used for these latter experiments, although the phosphoserine was not required for the binding activity (Rapoport et al., 1998). Further experiments will be required to establish exactly how the endocytosis signal sequence in CD4 interacts with clathrin adaptor complexes.
The observation that PEs could increase the endocytosis of some CD4 constructs containing substitutions in both phosphorylation sites has interesting implications. The in vitro adaptor binding assays indicate that adaptors only bind to the phosphorylated CD4 peptides (Table 3). Moreover, PE treatment did not increase bulk flow or fluid phase endocytosis over the course of these experiments. Thus one explanation for the PE-induced uptake of some nonphosphorylated constructs is that PE treatment leads to modification of the adaptors, or other clathrin-associated proteins, to enhance recognition of the nascent endocytosis signal in CD4. Components of the adaptor complex are known to be phosphorylated (Pauloin and Thurieau, 1994; Wilde and Brodsky, 1996; Bar-Zvi et al., 1998). To date this phosphorylation has been linked to adaptor association with clathrin (Wilde and Brodsky, 1996). However, our preliminary results suggest that phosphorylation of AP-2 can also modulate the interaction of this adaptor with the cytoplasmic domains of some membrane proteins (Fingerhut and Höning, unpublished results). We are currently investigating the possibility that adaptors are phosphorylated in PE-treated cells and that this phosphorylation may modify the association of adaptor complexes with endocytic sorting signals.
ACKNOWLEDGMENTS
We thank our colleagues for constant criticism during this work and Annegret Pelchen-Matthews, Nathalie Signoret, and Marie-José Bijlmakers for critically reading the manuscript. C.P., K.B., and M.M. were supported by grants from the United Kingdom Medical Research Council; M.M. and S.H. were supported by European Union Training and Mobility of Researchers program grant FMRX-CT96-0058.
Abbreviations used:
- AP
adaptor protein complex
- CD4
cluster of differentiation antigen 4
- MHC
major histocompatibility complex
- PE
phorbol ester
- PMA
phorbol 12-myristate 13-acetate
- SPR
surface plasmon resonance
- wt
wild-type
REFERENCES
- Bar-Zvi D, Mosley ST, Branton D. In vivo phosphorylation of clathrin-coated vesicle proteins from rat reticulocytes. J Biol Chem. 1998;263:4408–4415. [PubMed] [Google Scholar]
- Bremnes T, Lauvrak V, Lindqvist B, Bakke O. A region from the medium chain adaptor subunit (μ) recognizes leucine- and tyrosine-based sorting signals. J Biol Chem. 1998;273:8638–8645. doi: 10.1074/jbc.273.15.8638. [DOI] [PubMed] [Google Scholar]
- Bremnes B, Madsen T, Geddedahl M, Bakke O. An LI and ML motif in the cytoplasmic tail of the MHC- associated invariant chain mediate rapid internalization. J Cell Sci. 1994;107:2021–2032. doi: 10.1242/jcs.107.7.2021. [DOI] [PubMed] [Google Scholar]
- Corvera S, Chawla A, Chakrabarti R, Joly M, Buxton J, Czech MP. A double leucine within the GLUT4 glucose transporter COOH-terminal domain functions as an endocytosis signal. J Cell Biol. 1994;126:979–989. doi: 10.1083/jcb.126.4.979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darsow T, Burd CG, Emr SD. Acidic di-leucine motif essential for AP-3-dependent sorting and restriction of the functional specificity of the Vam3p vacuolar t-SNARE. J Cell Biol. 1998;142:913–922. doi: 10.1083/jcb.142.4.913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dietrich J, Hou X, Wegener A-M, Geisler C. CD3γ contains a phosphoserine-dependent di-leucine motif involved in down-regulation of the T cell receptor. EMBO J. 1994;13:2156–2166. doi: 10.1002/j.1460-2075.1994.tb06492.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dietrich J, Kastrup J, Nielsen BL, Odum N, Geisler C. Regulation and function of the CD3γ DxxxLL motif: a binding site for adaptor protein-1 and adaptor protein-2 in vitro. J Cell Biol. 1997;138:271–281. doi: 10.1083/jcb.138.2.271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endres MJ, et al. CD4-independent infection by HIV-2 is mediated by fusin. Cell. 1996;87:745–756. doi: 10.1016/s0092-8674(00)81393-8. [DOI] [PubMed] [Google Scholar]
- Ferguson SS, Barak LS, Zhang J, Caron MG. G-protein-coupled receptor regulation: role of G-protein-coupled receptor kinases and arrestins. Can J Physiol Pharmacol. 1996a;74:1095–1110. doi: 10.1139/cjpp-74-10-1095. [DOI] [PubMed] [Google Scholar]
- Ferguson SS, Downey WE, III, Colapietro AM, Barak LS, Menard L, Caron MG. Role of β-arrestin in mediating agonist-promoted G protein-coupled receptor internalization. Science. 1996b;271:363–366. doi: 10.1126/science.271.5247.363. [DOI] [PubMed] [Google Scholar]
- Goodman OBJ, Krupnick JG, Santinin F, Gurevich VV, Penn RB, Gagnon AW, Keen JH, Benovic JL. β-Arrestin acts as a clathrin adaptor in endocytosis of the β2-adrenergic receptor. Nature. 1996;383:447–450. doi: 10.1038/383447a0. [DOI] [PubMed] [Google Scholar]
- Griffiths G, Back R, Marsh M. A quantitative analysis of the endocytic pathway in baby hamster kidney cells. J Cell Biol. 1989;109:2703–2720. doi: 10.1083/jcb.109.6.2703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Healey D, et al. Novel antiCD4 monoclonal antibodies separate human immunodeficiency virus infection and fusion of CD4+ cells from virus binding. J Exp Med. 1990;172:1233–1242. doi: 10.1084/jem.172.4.1233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heilker R, Manning-Krieg U, Zuber JF, Spiess M. In vitro binding of clathrin adaptors to sorting signals correlates with endocytosis and basolateral sorting. EMBO J. 1996;15:2893–2899. [PMC free article] [PubMed] [Google Scholar]
- Hicke L, Zanolari B, Riezman H. Cytoplasmic tail phosphorylation of the α-factor receptor is required for its ubiquitination and internalization. J Cell Biol. 1998;141:349–358. doi: 10.1083/jcb.141.2.349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Höning S, Sandoval IV, von Figura K. A di-leucine-based motif in the cytoplasmic tail of LIMP-II and tyrosinase mediates selective binding of AP-3. EMBO J. 1998;17:1304–1314. doi: 10.1093/emboj/17.5.1304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Höning S, Sosa M, Hille-Rehfeld A, von Figura K. The 46-kDa mannose 6-phosphate receptor contains multiple binding sites for clathrin adaptors. J Biol Chem. 1997;272:19884–14890. doi: 10.1074/jbc.272.32.19884. [DOI] [PubMed] [Google Scholar]
- Huse M, Eck MJ, Harrison SC. A Zn2+ ion links the cytoplasmic tail of CD4 and the N-terminal region of Lck. J Biol Chem. 1998;273:18729–18733. doi: 10.1074/jbc.273.30.18729. [DOI] [PubMed] [Google Scholar]
- Jonsson U, et al. Real-time biospecific interaction analysis using surface plasmon resonance and a sensor chip technology. Biotechniques. 1991;11:620–627. [PubMed] [Google Scholar]
- Karlsson R, Michaelsson A, Mattsson L. Kinetic analysis of monoclonal antibody-antigen interactions with a new biosensor based analytical system. J Immunol Methods. 1991;145:229–240. doi: 10.1016/0022-1759(91)90331-9. [DOI] [PubMed] [Google Scholar]
- Kibbey RG, Rizo J, Gierasch LM, Anderson RGW. The LDL receptor clustering motif interacts with the clathrin terminal domain in a reverse turn conformation. J Cell Biol. 1998;142:59–67. doi: 10.1083/jcb.142.1.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirchhausen T, Bonifacino JS, Riezman H. Linking cargo to vesicle formation: receptor tail interactions with coat proteins. Curr Opin Cell Biol. 1997;9:488–495. doi: 10.1016/s0955-0674(97)80024-5. [DOI] [PubMed] [Google Scholar]
- Letourneur F, Klausner RD. A novel di-leucine motif and a tyrosine-based motif independently mediate lysosomal targeting and endocytosis of CD3 chains. Cell. 1992;69:1143–1157. doi: 10.1016/0092-8674(92)90636-q. [DOI] [PubMed] [Google Scholar]
- Marks MS, Woodruff L, Ohno H, Bonifacino JS. Protein targeting by tyrosine- and di-leucine-based signals: evidence for distinct saturable components. J Cell Biol. 1996;135:341–354. doi: 10.1083/jcb.135.2.341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsh M, Pelchen-Matthews A. Endocytic and exocytic regulation of CD4 expression and function. Curr Top Microbiol Immunol. 1996;205:107–135. doi: 10.1007/978-3-642-79798-9_6. [DOI] [PubMed] [Google Scholar]
- Ooi CE, Moreira JE, Dell’Angelica EC, Poy G, Wassarman DA, Bonifacino JS. Altered expression of a novel adaptin leads to defective pigment granule biogenesis in the Drosophila eye color mutant garnet. EMBO J. 1997;16:4508–4518. doi: 10.1093/emboj/16.15.4508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Shannessy D, Brigham-Burke M, Soneson K, Hensley P, Brooks I. Determination of rate and equilibrium constants for macromolecular interactions using surface plasmon resonance: use of non linear least squares analysis methods. Anal Biochem. 1993;212:457–468. doi: 10.1006/abio.1993.1355. [DOI] [PubMed] [Google Scholar]
- Owen DJ, Evans PR. A structural explanation for the recognition of tyrosine-based endocytosis signals. Science. 1998;282:1327–1332. doi: 10.1126/science.282.5392.1327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauloin A, Thurieau C. The 50 kDa protein subunit of assembly polypeptide (AP) AP-2 adapter from clathrin-coated vesicles is phosphorylated on threonine-156 by AP-1 and a soluble AP50 kinase which copurifies with the assembly polypeptides. Biochem J. 1994;296:409–415. doi: 10.1042/bj2960409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelchen-Matthews A, Armes JE, Griffiths G, Marsh M. Differential endocytosis of CD4 in lymphocytic and nonlymphocytic cells. J Exp Med. 1991;173:575–587. doi: 10.1084/jem.173.3.575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelchen-Matthews A, Armes JE, Marsh M. Internalization and recycling of CD4 transfected into HeLa and NIH-3T3 cells. EMBO J. 1989;8:3641–3649. doi: 10.1002/j.1460-2075.1989.tb08538.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelchen-Matthews A, Boulet I, Littman D, Fagard R, Marsh M. The protein tyrosine kinase p56lck inhibits CD4 endocytosis by preventing entry of CD4 into coated pits. J Cell Biol. 1992;117:279–290. doi: 10.1083/jcb.117.2.279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelchen-Matthews A, DaSilva R, Bijlmakers M-J, Signoret N, Gordon S, Marsh M. Lack of p56lck expression correlates with CD4 endocytosis in primary lymphoid and myeloid cells. Eur J Immunol. 1998;28:3639–3647. doi: 10.1002/(SICI)1521-4141(199811)28:11<3639::AID-IMMU3639>3.0.CO;2-Q. [DOI] [PubMed] [Google Scholar]
- Pelchen-Matthews A, Parsons IJ, Marsh M. Phorbol ester-induced down-regulation of CD4 is a multi-step process involving dissociation from p56lck, increased association with clathrin-coated pits and altered endosomal sorting. J Exp Med. 1993;178:1209–1222. doi: 10.1084/jem.178.4.1209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pond L, Kuhn LA, Teyton L, Schutze MP, Tainer JA, Jackson MR, Peterson PA. A role for acidic residues in di-leucine motif-based targeting to the endocytic pathway. J Biol Chem. 1995;270:19989–19997. doi: 10.1074/jbc.270.34.19989. [DOI] [PubMed] [Google Scholar]
- Rapoport I, Chen YC, Cupers P, Shoelson SE, Kirchhausen T. Dileucine-based sorting signals bind to the β-chain of AP-1 at a site distinct and regulated differently from the tyrosine-based motif-binding site. EMBO J. 1998;17:2148–2155. doi: 10.1093/emboj/17.8.2148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson MS. The role of clathrin, adaptors and dynamin in endocytosis. Curr Opin Cell Biol. 1994;6:538–544. doi: 10.1016/0955-0674(94)90074-4. [DOI] [PubMed] [Google Scholar]
- Sandoval I, Bakke O. Targeting of membrane proteins to endosomes and lysosomes. Trends Cell Biol. 1994;4:292–297. doi: 10.1016/0962-8924(94)90220-8. [DOI] [PubMed] [Google Scholar]
- Schuck P. Use of surface plasmon resonance to probe the equilibrium and dynamic aspects of interactions between biological macromolecules. Annu Rev Biophys Biomol Struct. 1997;26:541–566. doi: 10.1146/annurev.biophys.26.1.541. [DOI] [PubMed] [Google Scholar]
- Schuck P, Minton AP. Analysis of mass transport-limited binding kinetics in evanescent wave biosensors. Anal Biochem. 1996;240:262–272. doi: 10.1006/abio.1996.0356. [DOI] [PubMed] [Google Scholar]
- Shin J, Doyle C, Yang Z, Kappes D, Strominger JL. Structural features of the cytoplasmic region of CD4 required for internalization. EMBO J. 1990;9:425–434. doi: 10.1002/j.1460-2075.1990.tb08127.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin J, Dunbrack RL, Lee S, Strominger JL. Phosphorylation-dependent down-modulation of CD4 requires a specific structure within the cytoplasmic domain of CD4. J Biol Chem. 1991;266:10658–10665. [PubMed] [Google Scholar]
- Shiratori T, Miyatake S, Ohno H, Nakaseko C, Isono K, Bonifacino JS, Saito T. Tyrosine phosphorylation controls internalization of CTLA-4 by regulating its interaction with clathrin-associated adaptor complex AP-2. Immunity. 1997;6:583–589. doi: 10.1016/s1074-7613(00)80346-5. [DOI] [PubMed] [Google Scholar]
- Signoret N, et al. Phorbol esters and SDF-1 induce rapid endocytosis and down-modulation of the chemokine receptor CXCR4. J Cell Biol. 1997;139:651–664. doi: 10.1083/jcb.139.3.651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Signoret N, Rosenkilde MM, Klasse PJ, Schwartz TW, Malim MH, Hoxie JA, Marsh M. Differential regulation of CXCR4 and CCR5 endocytosis. J Cell Sci. 1998;111:2819–2830. doi: 10.1242/jcs.111.18.2819. [DOI] [PubMed] [Google Scholar]
- Simpson F, Peden AA, Christopoulou L, Robinson MS. Characterization of the adaptor-related protein complex, AP-3. J Cell Biol. 1997;137:835–845. doi: 10.1083/jcb.137.4.835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith PK, et al. Measurement of protein using bicinchoninic acid. Anal Biochem. 1985;150:76–85. doi: 10.1016/0003-2697(85)90442-7. [DOI] [PubMed] [Google Scholar]
- Strauss WM. Preparation of genomic DNA from mammalian tissue. In: Ausubel FM, et al., editors. Current Protocols in Molecular Biology. 1987. , Current Protocols, 2.2.1–2.2.3. [DOI] [PubMed] [Google Scholar]
- Strous GJ, van Kerkhof P, Govers R, Rotwein P, Schwartz AL. Growth hormone-induced signal transduction depends on an intact ubiquitin system. J Biol Chem. 1997;272:40–43. doi: 10.1074/jbc.272.1.40. [DOI] [PubMed] [Google Scholar]
- Trowbridge IS, Collawn JF, Hopkins CR. Signal-dependent membrane protein trafficking in the endocytic pathway. Annu Rev Cell Biol. 1993;9:129–161. doi: 10.1146/annurev.cb.09.110193.001021. [DOI] [PubMed] [Google Scholar]
- Wilde A, Brodsky FM. In vivo phosphorylation of adaptors regulates their interaction with clathrin. J Cell Biol. 1996;135:635–645. doi: 10.1083/jcb.135.3.635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willbold D, Roesch P. Solution structure of the human CD4 (403–419) receptor peptide. J Biochem Sci. 1996;3:435. doi: 10.1007/BF02258047. [DOI] [PubMed] [Google Scholar]