

Abstract
C-mannosylation of Trp-7 in human ribonuclease 2 (RNase 2) is a novel kind of protein glycosylation that differs fundamentally from N- and O-glycosylation in the protein-sugar linkage. Previously, we established that the specificity determinant of the acceptor substrate (RNase 2) consists of the sequence W-x-x-W, where the first Trp becomes C-mannosylated. Here we investigated the reaction with respect to the mannosyl donor and the involvement of a glycosyltransferase. C-mannosylation of Trp-7 was reduced 10-fold in CHO (Chinese hamster ovary) Lec15 cells, which are deficient in dolichyl-phosphate-mannose (Dol-P-Man) synthase activity, compared with wild-type cells. This was not a result of a decrease in C-mannosyltransferase activity. Rat liver microsomes were used to C-mannosylate the N-terminal dodecapeptide from RNase 2 in vitro, with Dol-P-Man as the donor. This microsomal transferase activity was destroyed by heat and protease treatment, and displayed the same acceptor substrate specificity as the in vivo reaction studied previously. The C-C linkage between the indole and the mannosyl moiety was demonstrated by tandem electrospray mass spectrometry analysis of the product. GDP-Man, in the presence of Dol-P, functioned as a precursor in vitro with membranes from wild-type but not CHO Lec15 cells. In contrast, with Dol-P-Man both membrane preparations were equally active. It is concluded that a microsomal transferase catalyses C-mannosylation of Trp-7, and that the minimal biosynthetic pathway can be defined as: Man –> –> GDP-Man –> Dol-P-Man –> (C2-Man-)Trp.
INTRODUCTION
Recently, a novel form of protein glycosylation was reported, the C-mannosylation of Trp-7 in human RNase 2. In this case the C1′ of an α-mannopyranosyl residue is directly linked to C2 of the indole moiety [(C2-Man-)Trp] (Hofsteenge et al., 1994; de Beer et al., 1995; Löffler et al., 1996). This modification takes place in cells from a variety of mammals, but it was not observed in two different cell types from insects, i.e. Sf9 cells from Spodoptera frugiperda and Schneider 2 cells from Drosophila melanogaster, in protoplasts from Orychophragmus violaceous, nor in Escherichia coli (Krieg et al., 1997). In contrast to the processes of protein N- and O-glycosylation (Hanover et al., 1982), nothing is known about the biosynthetic aspects of C-mannosylation. In the accompanying article we have examined the structural requirements of the protein acceptor, and found that the sequence -W-x-x-W/F-, in which the first Trp residue becomes mannosylated, forms the specificity determinant. Replacement of the second Trp by a Phe residue reduced the efficiency of C-mannosylation 3.5-fold (Krieg et al., 1998).
Here we address two other major questions: 1) which sugar precursor is involved in C-mannosylation of Trp-7 in RNase 2, and 2) does a transferase exist that carries out this reaction, or is C-mannosylation the result of self-glycosylation? These issues have been approached in different ways. First, the hybrid RNase 2.4–consisting of residues 1–13 of human RNase 2 and residues 11–119 of porcine RNase 4 (Krieg et al., 1998)–was purified from transfected NIH 3T3 cells that had been labeled with [2-3H]Man, and analyzed by peptide mapping and Edman degradation. Second, RNase 2.4 that was expressed in mutant CHO1 Lec151 cells, which lack Dol-P-Man synthase activity (Stoll et al., 1982; Beck et al., 1990), was compared with the enzyme obtained from wild-type cells by Western analysis with modification-specific antibodies, and by peptide mapping. Third, model peptides derived from human RNase 2 were C-mannosylated in vitro with Dol-P-[2-3H]-Man as the sugar donor and rat liver microsomes as the source of the transferase. The peptides were characterized by LC-ESIMS and Edman degradation. Fourth, membranes from wild-type and CHO Lec15 cells were assayed for C-mannosyl transferase activity, using either GDP-[2-3H]Man or Dol-P-[2-3H]Man as the sugar donor. The results established the biosynthetic pathway leading to (C2-Man-)Trp in RNase 2.
MATERIALS AND METHODS
D-[2-3H]Mannose (18.6 Ci/mmol) was from Amersham, England; GDP-[2-3H]Man (15.9 Ci/mmol) was from Dupont NEN, MA; Dol-P was obtained from Sigma, St. Louis, MO. DEAE-cellulose (DE 52) was from Whatmann, Maidstone, England. C18 Sep-Pak cartridges were from Waters, Milford, CT and protein A Sepharose beads were from Pharmacia, Uppsala, Sweden. Silica TLC plates (60F254) were from Merck, Darmstadt, Germany; Triton X-100 was from Fluka, Buchs, Switzerland; thermolysin and chymotrypsin were from Boehringer, Mannheim, Germany; and elastase was from Worthington, Freehold, NJ. Peptides were synthesized at the FMI with an automated solid-phase synthesizer with Fmoc chemistry. They were purified by reverse phase HPLC and characterized by mass spectrometry and amino acid analysis. Cell culture media, FCS and lipofectamine reagent (2 mg/ml) were from Life Technologies, Gaithersburg, MD. ECL Western analysis reagents were from Amersham Corp., Arlington Heights, IL. All other chemicals were from Fluka, Buchs, Switzerland.
Recombinant RNases 2 and 4 were purified from E. coli, and RNase-2/urine from a crude preparation of human chorionic gonadotropin as previously described (Hofsteenge et al., 1994; Vicentini et al., 1994). Antibodies α-RNase 2, α-RNase 4 and α-(5–10) were prepared as described by Krieg et al. (1997) and Löffler et al. (1996).
Cell Culture, Transfection and Metabolic Labeling with D-[2-3H]Mannose
NIH 3T3 (NIH swiss mouse) cells were obtained from the American Type Culture Collection, Rockville, MD (ATCC CRL 1658) and were maintained at 37°C and 5% CO2 in low-glucose Dulbecco’s modified Eagle medium (DMEM), supplemented with 10% newborn calf serum. They were transfected with the plasmid encoding RNase 2.4 (Krieg et al., 1998) with the cationic lipid lipofectamine as described by the supplier.
The CHO Lec15 cell line and the corresponding wild-type line were a kind gift from Dr. S. Krag, John Hopkins University, Baltimore, MD. These cells were propagated at 37°C and 5% CO2 in MEM α medium supplemented with 10% FCS. CHO wild-type and Lec15 cells were plated, at 3.6 × 106 and 1.6 × 106 cells per 10 cm plate, respectively, 20 h before transfection. Optimal transfection conditions were: 32 and 28.8 μl of lipofectamine, 8.0 and 7.1 μg of RNase 2.4 plasmid for CHO wild-type and Lec15 cells, respectively. After 5 h the cells were supplemented with 10% dialysed newborn calf serum, and incubated with 36 μCi/ml of D-[2-3H]mannose for 48 h. Conditioned medium was collected for immunoprecipitation or RNase purification.
Immunoprecipitation and Immunoblotting
SDS-PAGE and Western blotting were performed as described previously by using modification-specific [α(5–10)] antibodies (Krieg et al., 1997). Immunoprecipitation was performed as described (Harlow and Lane, 1988) with protein A Sepharose beads coupled to rabbit anti-RNase 4 antibodies. Immunoprecipitated, radiolabeled RNases were loaded onto a 12.5% SDS polyacrylamide gel and submitted to fluorography for 30 min in 1 M sodium salicylate pH 6.0. The dried gel was exposed to a Kodak XAR Omat preflashed film for 4 days at −70°C with an intensifying screen.
Purification of RNase 2.4 and Protein Chemical Analysis
Radioactively labeled RNase 2.4 that was secreted by transfected 3T3 cells, was purified from the conditioned medium with a three-step purification procedure (Krieg et al., 1998), after addition of 10 μg of purified RNase 4 as a carrier. RNase 2.4 was monitored by counting radioactivity in 2 ml of Ready Save scintillation cocktail (Beckman) with a Packard scintillation counter. The protein was digested with thermolysin (Löffler et al., 1996), in the presence of 1.2 μg each of RNase 2/urine and RNase 2/E. coli, which provided C-mannosylated and unmodified marker peptides. The radioactive thermolytic peptide was purified by C18 reversed phase HPLC and sequenced by solid-phase Edman degradation (Pisano et al., 1993). The position of the tritiated amino acid residue in the peptide was established by counting the anilinothiazolinone-amino acid liberated in each cycle.
RNase 2.4 from wild-type and CHO Lec15 cells was purified by the same procedure but without added carrier protein. The degree of C-mannosylation was determined after digestion with thermolysin as described (Krieg et al., 1997).
Preparation of Rat Liver Microsomes and Membranes from CHO Cells
Female adult rats were starved overnight and killed by decapitation. The liver was removed immediately, cleaned from connective tissue, and homogenized in 10 mM Tris-HCl, pH 7.4 containing 1 mM MgCl2 and 0.25 M saccharose with a Potter-Elvehjem homogenizer. Microsomes were prepared by sucrose gradient centrifugation according to the method of Graham (1992). The microsomal pellet was resuspended in 20 mM HEPES-NaOH pH 7.2, containing 110 mM potassium acetate and 2 mM magnesium acetate (KMH buffer) and stored in liquid nitrogen. The protein concentration was determined by the method of Bradford (1976).
Total membrane fraction from wild-type and CHO Lec15 cells was prepared as described (Chaney et al., 1989). The membrane pellet was resuspended in KMH buffer and stored at −70°C.
Synthesis and Purification of Dolichyl-Phosphate-Mannose
Dol-P-[2-3H]Man (5.61 Ci/mmol) was synthesized from GDP-[3H]Man and Dol-P with rat liver microsomes as a source of GDP-Man:Dolichol phosphorylmannosyltransferase (Cacan and Verbert, 1995). The Dol-P-[2-3H]Man was extracted with chloroform/methanol (3:2, vol/vol) and purified on a DE 52 column according to Cacan and Verbert (1995). The final product was judged to be pure from analysis by silica TLC with chloroform/methanol/water (60:25:4 vol/vol/vol) or (86:14:1 vol/vol/vol) as the eluent (Oliver et al., 1975).
In vitro C-Glycosylation Mannosylation of Model Peptides and Product Characterization
The reaction mixture contained in a final volume of 24 μl: 45 pmol of Dol-P-[3H]Man (5.61 Ci/mmol), 0.9 mM of peptide N-AC-KPPQFAWAQWFE-NH2, 145 μg of rat liver microsomal protein, 20 mM HEPES-NaOH pH 7.2, 110 mM potassium acetate, 2 mM magnesium acetate, proteases inhibitors (2 μg/ml Benzamidine, 5 μg/ml Pepstatin A, 5 μg/ml Leupeptin, 2 mM EDTA), 0.2% Triton X-100. The Triton X-100/protein ratio (wt/wt) was kept constant at 0.34. The mixture was incubated at 37°C for various lengths of time and the reaction was stopped by adding 2 ml of chloroform/methanol 3:2 (vol/vol) and 0.48 ml of water. After centrifugation, the upper, aqueous phase contained the peptide and the lower, organic phase the Dol-P-[3H]Man. The radioactivity in 0.2 ml of the upper phase was determined by scintillation counting.
When GDP-[3H]Man (5.61 Ci/mmol) was used as the mannosyl donor, the assay was performed as described above, except that 2 mM MgCl2, 2 mM MnCl2, 1 mM ATP and 0.4 mM Dol-P were added. Following extraction, the aqueous phase was lyophylised, resuspended in 0.1% trifluoroacetic acid (TFA) and loaded onto a C18 column by gravity to separate the radiolabeled peptide from the GDP-[3H]Man. The column was equilibrated in 0.1% TFA, washed with 20% of buffer B (0.085% TFA, 70% CH3CN) and eluted with buffer B. The eluate was dried, dissolved in water and radioactivity was measured by scintillation counting.
To test the effect of protease treatment, microsomes were digested with chymotrypsin (200 μg/ml, added in 5 aliquots at 20 min intervals) for 90 min at 26°C under the assay conditions except that the peptide was omitted. The chymotrypsin was blocked with soybean trypsin-inhibitor (1.3 mg/ml) for 30 min at 26°C. The peptide was added at usual concentration and the mixture was incubated for 2 h at 26°C. The complete inhibition of the chymotrypsin was demonstrated by the lack of cleavage of the peptide.
To characterize the radioactively labeled peptide, the aqueous phase of eight experiments was dried. The peptide was purified by chromatography by using a C18 Sep-Pak cartridge, and two HPLC steps by using a C18 and a C8 column with 0.1 or 0.05% TFA as the eluent. The last step was performed on a liquid chromatograph interfaced with a Perkin Elmer-Cetus API 300 mass spectrometer with a 9:1 flow split. The purified peptide was dried, digested with elastase (Löffler et al., 1996) and fractionated by C8 reversed phase LC-ESIMS. In a separate experiment, the radioactive peptide was mixed with 168 pmol of peptide 1–12 obtained from RNase 2/urine, digested with thermolysin (Krieg et al., 1997a), and purified by C18 reversed phase HPLC. The radioactive thermolytic peptide was subjected to solid-phase Edman degradation. Nano-ES was performed as described (Wilm and Mann, 1996).
RESULTS
Mannose Is the Earliest Precursor in C-Mannosylation
To identify the earliest sugar precursor in the biosynthesis of (C2-Man-)Trp in RNase 2, 3T3 cells were transfected with the gene encoding the hybrid RNase 2.4 and labeled with D-[2-3H]mannose. RNase 2.4, rather than RNase 2 itself, was used, because it lacks N-glycosylation sites, which simplified the protein chemical characterization (Krieg et al., 1998). Analysis of the secreted, immuno-precipitated RNase 2.4 by SDS-PAGE revealed a single radioactive protein, that comigrated with RNase 4 (Figure 1A, lane 1). In contrast, no radioactive band was observed at that position in a control experiment with nontransfected cells (Figure 1A, lane 2). This result strongly suggested that d-mannose is a precursor in the biosynthesis of (C2-MaN-)Trp.
Figure 1.

Characterization of RNase 2.4 labeled with D-[2-3H]Mannose. (A) NIH 3T3 cells were transfected with the plasmid encoding RNase 2.4 and labeled with D-[2-3H]mannose. RNase 2.4 was immunoprecipitated with polyclonal α-RNase 4 antibodies from 6.5 ml of conditioned medium, and fractionated on a 12.5% SDS-PAA gel (lane 1). The immunoprecipitate of conditioned medium from nontransfected control cells was loaded in lane 2. The gel was submitted to fluorography and exposed for 4 d at −70°C. The position of RNase 4, which was visualized by Coomasssie brilliant blue staining, has been indicated by an arrow. (B) Portion of the thermolytic peptide map of radiolabeled RNase 2.4. Tritiated RNase 2.4 was purified from conditioned medium from transfected 3T3 cells and mixed with 1.2 μg of r-RNase 2/E-coli and 1.2 μg of RNase 2/urine. The mixture was digested with thermolysin at 75°C and the peptides were fractionated by C18 reversed phase HPLC. Fractions were collected and examined for radioactivity. The width of the bar indicates the fraction size and the height represents the radioactivity in 10 μl. The peaks containing unmodified peptides 6–10 (TWAQW) and 5–10 (FTWAQW) have been labeled ‘a’ and ‘c’, respectively. C-mannosylated peptide 5–10 has been labeled with ‘b’. (C) Solid-phase Edman degradation of peptide 5–10. Radioactive peptide from ‘B’ was covalently coupled to a sequelon-AA membrane and sequenced. The radioactivity of the anilinothiazolinone-amino acid released at each cycle was determined.
The radiolabeled protein was further characterized by peptide mapping. RNase 2.4 was purified from the conditioned medium, and digested with thermolysin. Equal amounts of RNase 2/urine and RNase 2/E. coli were added before digestion to provide fully C-mannosylated and unmodified marker peptides, respectively. Fractionation of the thermolytic digest by C18 reversed phase HPLC resulted in a single radioactive peptide (Figure 1B, lower panel) eluting exactly at the position of the C-mannosylated marker peptide ‘b’. This indicated that the [3H]mannosylated residue of RNase 2.4 was located between residue 5 and 10. The peptide was subjected to Edman degradation to determine the position of the mannosylated residue. A burst of radioactivity appeared in cycle 3 (Figure 1C), coinciding with (C2-Man-)Trp in the known sequence of the thermolytic peptide 5–10, FT (C2-MaN-)WAQW (Hofsteenge et al., 1994).
Dol-P-Man Synthase Deficient Cells Poorly C-Mannosylate RNase
The biosynthesis of N-linked glycans employs both cytoplasmic GDP-Man, as well as Dol-P-Man in the lumen of the endoplasmic reticulum as mannosyl donors (Hanover et al., 1982; Abeijon and Hirschberg, 1992). Since RNase 2 contains a signal sequence for membrane translocation and is secreted from the cell, it must travel through the endoplasmic reticulum. Therefore, we hypothesized that Dol-P-Man is a precursor in the biosynthesis of (C2-Man-)Trp. To investigate this, RNase 2.4 was expressed in CHO Lec15 cells and compared with the enzyme obtained from wild-type cells. This mutant cell line has been reported to be deficient in Dol-P-Man synthase activity and to contain decreased levels of Dol-P-Man (Stoll et al., 1985; Beck et al., 1990; Rosenwald et al., 1990; Stoll et al., 1992). RNase 2.4 was purified from the conditioned medium of these cells and analyzed by Western blot with modification-specific antibodies [α(5–10)] (Krieg et al., 1997). There was a nearly complete absence of C-mannosylation in the CHO Lec15 cells compared with the wild type (Figure 2A). The protein from both cell lines was further characterized by digestion and peptide mapping by HPLC and ESIMS. This showed that in the Lec15 cells peak ‘b’, which was shown to contain the C-mannosylated peptide 5–10 ([M+H]+ = 1000.5), was strongly reduced. A concomitant increase in the unmodified peptides 6–10 and 5–10 (Figure 2B, peak ‘a’: [M+H]+ = 691.5; peak ‘c’: [M+H]+ = 835.8) was observed. From the peak areas, the degree of C-mannosylation at Trp-7 was determined to be 46.0% and 4.7% in the wild-type and CHO Lec15 cells, respectively. These results indicated that either Dol-P-Man is a precursor in the biosynthesis of (C2-Man-)Trp, or that the putative C-mannosyl transferase requires correct N-glycosylation for folding and/or activity. To distinguish between these possibilities, an in vitro C-mannosylation system was established.
Figure 2.

Characterization of RNase 2.4 expressed by wild-type and CHO Lec15 cells. (A) Western analysis of purified RNase 2.4 from wild-type and CHO Lec15 cells with modification specific antibodies. HPLC-purified RNase 2.4 was fractionated on a 12.5% SDS-PAA gel and detected by immunoblotting with modification specific antibodies [α(5–10)]. 400 μg of purified RNase 2.4 from the CHO cells, and 200 μg of human RNase 2/urine were loaded. (B) Part of the thermolytic peptide map of RNase 2.4 purified from CHO wild-type (upper panel) and CHO Lec15 cell (lower panel). RNase 2.4 (1.8 μg) was digested with thermolysin at 75°C and the peptides were fractionated by reversed phase C18 HPLC. For details, see the legend to Figure 1B. The portion of the chromatogram containing peptides ‘a’, ‘b’ and ‘c’ is shown. The shift of 1.5 min between the chromatograms was a result of aging of the C18 column. In both experiments the identity of peptides a-c was verified by coelution with peptides from RNase 2/urine and -/E. coli, and by ESIMS.
Model peptides of RNase 2 are mannosylated in vitro
An in vitro C-mannosylation system was established on the basis of the in vivo observation that the first 12 residues of RNase 2 are sufficient for modification (Krieg et al., 1998). The peptide N-AC-KPPQFAWAQWFE-NH2, with Ala replacing Thr-6 to prevent possible O-glycosylation, was selected as the mannosyl acceptor substrate. Incubation of the peptide with Dol-P-[3H]Man in the presence of rat liver microsomes, followed by extraction with chloroform/methanol (3:2 vol/vol), resulted in the appearance of tritiated material in the aqueous phase (Figure 3A). Control experiments carried out without microsomes resulted in 91% less radioactivity in the aqueous phase (Figure 3A). Furthermore, replacement of Trp-7 by Ala reduced the amount of radioactivity in the aqueous phase to essentially background levels (Figure 3B). These results are consistent with transfer of [3H]mannose from Dol-P-[3H]Man to Trp-7 of the peptide.
Figure 3.

In vitro C-mannosylation reaction. (A) The peptide N-AC-KPPQFAWAQWFE-NH2 (900 μM) was incubated with Dol-P-[2-3H]Man (0.25 μCi) and rat liver microsomes (145.8 μg of protein) for 30 min at 37°C or 2h at 26°C. After extraction with chloroform/methanol (3/2 vol/vol), the total amount of radioactivity in the aqueous phase, which contains the peptide, was determined. (B) Various polypeptides were incubated and analyzed as under ‘A’, except that the concentration of r-RNase 2 was 300 μM. The values were corrected for background by subtracting the value obtained in the absence of peptide. The data in A and B represent the average of two independent experiments.
At 37°C the appearance of radioactivity in the peptide increased linearly over 60 min (Figure 4A), at which time 8.5% of the radioactivity in Dol-P-[2-3H]Man had been transferred. A much longer linear phase was observed, however, at 26°C (Figure 4A, inset). Under these conditions, 52% of the radioactivity in Dol-P-[3H]Man had been transferred to the peptide after 22 h. The rate of reaction was proportional to the amount of microsomal protein added (Figure 4B). In all experiments, the reaction depended on the addition of peptide acceptor (Figure 3A and 4). To examine whether a protein transferase was involved in the process, microsomes were heated at 95°C or treated with chymotrypsin. In both cases the amount of radioactivity transferred to the peptide decreased to essentially background level (Figure 3A).
Figure 4.

Dependence of the in vitro C-mannosylation reaction on time and amount of microsomes. The peptide N-AC-KPPQTAWAQWFE-NH2 was incubated with Dol-P-[2-3H]Man (0.25 μCi) and rat liver microsomes. (A) After various length of time of incubation at 37°C, the reaction was stopped by extraction with chloroform/methanol (3/2 vol/vol). The amount of radioactivity in the aqueous phase, which contains the peptide, was determined (•). Control experiments were performed without acceptor peptide (▪). The inset shows the time course measured at 26°C. (B) The amount of radioactivity in the aqueous phase after 30 min incubation with various amount of microsomes were determined (•). The data in A and B represent the average of two independent experiments.
The In Vitro Reaction Results in C-Mannosylation of Trp-7
To characterize the radioactive peptide material, the aqueous phase was fractionated by reversed phase C18 HPLC. A single radioactive peak was observed (recovery: 43%), which eluted 1.5 min earlier than the remaining unmodified acceptor peptide (Figure 5A). These results are consistent with the presence of a mannosyl residue in the tritiated peptide (Hofsteenge et al., 1996). The radioactive peptide obtained from the C18 HPLC column was purified to apparent homogeneity by reversed phase C8 LC-ESIMS (68% recovery). Its molecular mass was in excellent agreement with that expected for the mono-C-mannosylated acceptor peptide ([M+H]+ = 1739). The exact position of attachment of the mannosyl residue was established by analysis of subpeptides. Fractionation of the elastase digest by LC-ESIMS yielded two peptides with a molecular mass of 1148 and 607.5 Da (Figure 5B). Nano-ES-MSMS demonstrated these to correspond to mannosylated peptide 1–8 and unmodified peptide 9–12, respectively. In agreement with this, the radioactivity coeluted with residues 1–8, showing that Trp-10 had not been modified (Figure 5B, lower panel). The modification of Trp-7 was confirmed by solid-phase Edman degradation of the peptide 5–10 obtained by thermolytic digestion. A burst of radioactivity in cycle 3 was observed, coinciding with (C2-Man-)Trp (Figure 6B). Evidence for a C-C link between the mannosyl residue and the tryptophan was obtained from a nano-ESI-MSMS experiment, which showed a 120-Da loss from the fragments containing Trp-7 (Figure 6A). This behaviour is characteristic for aromatic C-glycosides (Hofsteenge et al., 1996).
Figure 5.

Purification of the radioactive peptide. Purification of the radiolabeled peptide by reversed phase HPLC. The peptide N-AC-KPPQFAWAQWFE-NH2 was labeled as described in the legend to Figure 3A. (A) The peptide in the aqueous phase was lyophilized and purified by C18 reversed phase HPLC. Unmodified peptide 1–12 has been indicated with an asterisk and mannosylated tritiated peptide with an arrowhead (upper panel). The lower panel shows the radioactivity, the width of the bar indicates the fraction size. (B) The purified peptide 1–12 was digested with elastase and fractionated on a C8 reversed phase column. The peaks containing peptides 1–8 and 9–12 have been labeled (upper panel). Fractions were collected and examined for radioactivity (lower panel).
Figure 6.

Characterization of the radioactive peptides. (A) Nano-ESI-MSMS experiment of the radioactive peptide 1–8 obtained by elastase digestion as described in the legend to Figure 5B. The 120 Da loss typical for aromatic C-glycosides is shown. The portion of the spectrum that has been enlarged fourfold has been indicated with dashes. (B) Radiolabeled peptide obtained from an experiment as shown in Figure 5A was digested with thermolysin and purified by HPLC as described in the legend to Figure 1B. The radioactive thermolytic peptide 5–10 was subjected to solid-phase Edman degradation and the radioactivity of the anilinothiazolinone-amino acid released at each cycle was measured.
The C-Mannosylation Pathway
The C-mannosyltransferase activity in wild-type and CHO Lec15 cells was determined with the in vitro assay with the total membrane fraction as the enzyme source, and Dol-P-[3H]Man as the donor. The specific activity of the C-mannosyltransferase was essentially the same in both cell lines (Table 1).
Table 1.
C-Mannosyltransferase activity in membranes from CHO cells
| Source of membrane | cpm incorporated into peptide
|
||
|---|---|---|---|
| Dol-P-[3H]Man | GDP-[3H]Man − Dol-P | GDP-[3H]Man + Dol-P | |
| CHO wild type | 19480 | 664 | 12400 |
| CHO Lec15 | 22031 | 46 | 15 |
The C-mannosyltransferase activity in membranes (75.5 μg of total protein) from wild-type and CHO Lec15 cells was determined using the standard peptide as the acceptor (see Figure 3) and Dol-P-[2-3H]Man as the donor. Alternatively, GDP-[2-3H]Man was tested as a donor in the absence or presence of Dol-P (0.4 mM). The amount of radioactivity incorporated into the peptide was corrected for background. The data are the average of two independent experiments.
In the cell Dol-P-Man is synthesized from Dol-P and GDP-Man (Waechter and Lennarz, 1976; Stoll et al., 1982). It was, therefore, of interest to examine whether GDP-Man could serve as a precursor in C-mannosylation. In the absence of exogenously added Dol-P, a small but significant amount of incorporation of [3H]Man into the peptide was observed with membranes from wild-type CHO cells (Table 1). Essentially background values were obtained with membranes from CHO Lec15 cells (Table 1). Addition of exogenous Dol-P stimulated the reaction with wild-type membranes 19-fold, but not with Lec15 membranes. This result substantiates that Dol-P-Man is an obligate precursor of C-mannosylation.
The C-mannosylation reaction in vitro and in vivo has the same specificity
In the accompanying article (Krieg et al., 1998), it was found that a Trp (or Phe) is required at position +3 from the C-mannosylated tryptophan. In addition, several mutations in the N-terminal region caused an increase in the degree of C-mannosylation (Krieg et al., 1998). It was therefore of interest to examine the effect of mutations in the peptide substrate in the in vitro assay. Replacement of Trp-10 by Ala completely abolished C-mannosylation of Trp-7 (Figure 3B). The effect of this mutation reflected a difference in C-mannosylation and not in peptide recovery since the latter was the same (50% ± 5) for all peptides used. In vivo the T6A mutation caused an increase in the stoichiometry of C-mannosylation from 0.58 for wild-type RNase 2.4, to 0.88 for the mutant enzyme (Figure 4 in Krieg et al., 1998). The same mutation in vitro, however, did not show this effect (Figure 3B).
Interestingly, native recombinant RNase 2/E. coli (r-RNase 2/E.coli; 0.3 mM) did not serve as a substrate (Figure 3B), although the standard peptide at the same concentration was C-mannosylated (65% compared with standard assay conditions). Unfolded RNase 2/E. coli could not be tested, because of its insolubility.
DISCUSSION
The results presented here define the pathway for the synthesis of (C2-Man-)Trp in RNase 2, and demonstrate the requirement for a novel microsome-associated transferase activity. A mechanism in which RNase 2 undergoes some form of ‘self-glycosylation’, as has been demonstrated for glycogenin and amylogenin (Alonso et al., 1995; Singh et al., 1995), can be excluded for the following reasons: 1) a small portion of the RNase (in vitro, residues 1–12) is sufficient for C-mannosylation; 2) a protein associated with the microsomal membrane is required (Figure 3A); 3) native RNase 2 did not become C-mannosylated when incubated with Dol-P-[3H]Man in vitro (Figure 3B). Thorough characterization of the product of in vitro C-mannosylation and its subpeptides by LC-ESIMS and Edman degradation (Figure 5 and 6), unequivocally demonstrated that Trp-7, but not Trp-10 was the target of the transferase. Furthermore, replacement of Trp-10 by Ala abolished the capacity of the peptide to accept the mannosyl residue (Figure 3B). Exactly the same results were obtained in vivo by mutagenesis studies (Krieg et al., 1998), strongly suggesting that the C-mannosyltransferase activities are the same.
The pathway for the biosynthesis of (C2-Man-)Trp can be defined as
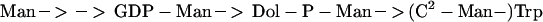 |
The obligate involvement of Dol-P-Man was concluded from a 10-fold reduction in the level of (C2-Man-)Trp in RNase 2.4 obtained from transfected CHO Lec15 cells (Figure 2), which are impaired in Dol-P-Man synthesis (Beck et al., 1990). It could be excluded that this decrease was a result of a reduction in the C-mannosyltransferase, since its specific activity in membranes from mutant and wild-type cells was the same (Table 1). Further evidence was obtained from in vitro experiments with GDP-[3H]Man as the donor. In the absence of added Dol-P a small amount of C-mannosylation took place in wild-type membranes, probably by using endogenous Dol-P (Chaudhary et al., 1982). Addition of Dol-P resulted in a strong increase in the level of C-mannosylation in the wild type, but not in the mutant cells (Table 1). Finally, Dol-P-Man by itself was sufficient for C-mannosylation (Figure 3). This is the third pathway in mammals in which Dol-P-Man is required; it is also involved in protein N-glycosylation (Behrens et al., 1973; Evans and Hemming, 1973; Waechter et al., 1973) and in the synthesis of glycosylphosphatidylinositol anchors (DeGasperi et al., 1990). The small amount of C-mannosylation found to occur in CHO Lec15 cells (Figure 2) deserves comment. Initially it was reported that CHO Lec15 cells were deficient in Dol-P-Man synthase activity (Beck et al., 1990). Subsequent studies showed, however, that they can synthesize functional glycosylphosphatidylinositol anchor, suggesting the presence of small amounts of Dol-P-Man (Rosenwald et al., 1990; Singh and Tartakoff, 1991).
It was not possible to detect intermediates between Dol-P-Man and the peptide substrate. Incubation of the peptide with Dol-P-[3H]Man and microsomes for 22 h at 26°C resulted in transfer of more than 50% of the radioactivity in Dol-P-Man to the peptide (Figure 4, inset). By omitting the peptide, a substantial amount of radioactivity would accumulate in a possible intermediate. Under these conditions very little radioactivity was transferred to the aqueous phase (Figure 4), which was not analyzed further. The organic phase contained 97% of the radioactive input. Using two different TLC systems, it was found that this radioactivity was entirely present in Dol-P-Man (our unpublished observations). These results are consistent with a direct transfer of Man from Dol-P-Man to the peptide. A less likely explanation that cannot be excluded is the existence of an intermediate that was either unstable under the conditions of analysis, or that is present at a very low level. Purification of the C-mannosyltransferase should answer this question.
The use of phosphodiester activated sugars in the biosynthesis of aromatic C-glycosides is not unprecedented. Franz and colleagues have provided evidence that the glucose in vitexin and isovitexin, two low-molecular-weight C-glucosides from plants, is donated by UDP- and ADP-glucose (Kerscher and Franz, 1987; Kerscher and Franz, 1988).
The specificity of the C-mannosylation reaction for Trp-7 in RNase 2 is determined by the presence of a Trp (or Phe) residue at position +3 (Krieg et al. (1998) and Figure 3). It seems that the primary, rather than the tertiary structure, forms the positive specificity signal for C-mannosylation. The observation that native r-RNase 2 does not function as a substrate in vitro but is effectively C-mannosylated in vivo (Krieg et al., 1998), indicates that folding actually has a negative effect. Inspection of the three-dimensional structure of r-EDN (= r-RNase 2) (Mosimann et al., 1996) revealed that the indole of Trp-7 is at the surface, and that a mannose residue can be accommodated without structural changes. In contrast, most of the indole ring of Trp-10 is buried between side chains from the N-terminal region and helix α2. The partial modification of Trp-7 in vivo would result from a competition between the C-mannosylation reaction and protein folding. This situation is akin to N-glycosylation, where the sequon Asn-X-Thr/Ser is required, but its usage can be modified by three-dimensional structural constraints (Goochee et al., 1992). This model would also explain the finding of mutants that improve C-mannosylation of RNase 2 and 2.4 in vivo, if it is assumed that they slow down the rate of folding (Krieg et al., 1998). This would predict that the same mutation in vitro, using an unfolded peptide, would not have an accelerating effect. Comparison of the data obtained for the T6A mutant in vitro confirms this prediction (Figure 3).
In conclusion, we have shown that 1) the biosynthesis of (C2-Man-)Trp in RNase 2 involves a transferase that most likely uses Dol-P-Man as the sugar donor; and 2) the recognition signal consists of the linear epitope W-x-x-W.
ACKNOWLEDGMENTS
We thank Dr. Sharon Krag for providing us with the CHO cells and for carefully reading the manuscript, Dr. Wolfgang Gläsner for expression and purification of RNase 2/E. coli, Anna-Maria Buxton-Vicentini for the RNase 2.4 plasmid construct, Renate Matthies for peptide sequencing, and Franz Fisher for peptide synthesis.
Footnotes
Abbreviations used: CHO, chinese hamster ovary; (C2-Man-)Trp, C2-α mannopyranosyltryptophan; nano-ES, nanoelectrospray; LC, liquid chromatography; MS, mass spectrometry; TLC, thin-layer chromatography.
REFERENCES
- Abeijon C, Hirschberg CB. Topography of glycosylation reactions in the endoplasmic reticulum. Trends Biochem Sci. 1992;17:32–36. doi: 10.1016/0968-0004(92)90424-8. [DOI] [PubMed] [Google Scholar]
- Alonso MD, Lomako J, Lomako WM, Whelan WJ. A new look at the biogenesis of glycogen. FASEB J. 1995;9:1126–1137. doi: 10.1096/fasebj.9.12.7672505. [DOI] [PubMed] [Google Scholar]
- Beck PJ, Orlean P, Albright C, Robbins PW, Gething MJ, Sambrook JF. The Saccharomyces cerevisiae DPM1 gene dolichol-phosphate-mannose synthase is able to comlement a glycosylation-defective mammalian cell line. Mol Cell Biol. 1990;10:4612–4622. doi: 10.1128/mcb.10.9.4612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrens NH, Carminatti H, Staneloni RJ, Leloir LF, Cantarella AI. Formation of lipid-bound oligosaccharides containing mannose: their role in glycoprotein synthesis. Proc Natl Acad Sci USA. 1973;70:3390–3394. doi: 10.1073/pnas.70.12.3390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Cacan R, Verbert A. Gycosyltransferases of the phosphodolichol pathways. In: Verbert A, editor. Methods on glycoconjugates. Chur, Switzerland: Harwood Academic Publishers; 1995. pp. 191–199. [Google Scholar]
- Chaney W, Sundaram S, Friedman N, Stanley P. The Lec4A CHO glycosylation mutant arises from miscompartmentalization of a Golgi glycosyltransferase. J Cell Biol. 1989;109:2089–2096. doi: 10.1083/jcb.109.5.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaudhary N, Freeman DJ, Rip JW, Carroll KK. Direct estimation of dolichyl phosphate in rat liver by high-pressure liquid chromatography. Lipids. 1982;17:558–560. doi: 10.1007/BF02535384. [DOI] [PubMed] [Google Scholar]
- de Beer T, Vliegenthart JFG, Löffler A, Hofsteenge J. The hexopyranosyl residue that is C-glycosidically linked to the side chain of tryptophan-7 in human RNase Us is α-mannopyranose. Biochemistry. 1995;34:11785–11789. doi: 10.1021/bi00037a016. [DOI] [PubMed] [Google Scholar]
- DeGasperi R, Thomas LJ, Sugiyama E, Chang HM, Beck PJ, Orlean P, Albright C, Waneck G, Sambrook JF, Warren CD, Yeh ETH. Correction of a defect in mammalian GPI anchor biosynthesis by a transfected yeast gene. Science. 1990;250:988–991. doi: 10.1126/science.1978413. [DOI] [PubMed] [Google Scholar]
- Evans PJ, Hemming FW. The unambiguous characterization of dolichol phosphate mannose as a product of mannosyl transferase in pig liver endoplasmic reticulum. FEBS Lett. 1973;31:335–338. doi: 10.1016/0014-5793(73)80135-8. [DOI] [PubMed] [Google Scholar]
- Goochee CF, Gramer MJ, Andersen DC, Bahr JB. The oligosaccharides of glycoproteins: factors affecting their synthesis and their influence on glycoprotein properties. In: Todd P, Sikdar SK, Milan B, editors. Frontiers in bioprocessing II. Washington, DC: American Chemical Society; 1992. pp. 199–240. [Google Scholar]
- Graham J. Isolation of subcellular organelles and mebranes. In: Rickwood D, editor. Centrifugation; a practical approach. Oxford: IRL Press; 1992. pp. 161–182. [Google Scholar]
- Hanover JA, Elting J, Mintz GR, Lennarz WJ. Temporal aspects of the N- and O-glycosylation of human chorionic gonadotropin. J Biol Chem. 1982;257:10172–10177. [PubMed] [Google Scholar]
- Harlow E, Lane D. Antibodies. A laboratory manual. Cold Spring Harbor Laboratory; 1988. [Google Scholar]
- Hofsteenge J, Müller DR, de Beer T, Löffler A, Richter WJ, Vliegenhart JFG. New type of linkage between a carbohydrate and a protein: C-glycosylation of a specific tryptophan residue in human RNase Us. Biochemistry. 1994;33:13524–13530. doi: 10.1021/bi00250a003. [DOI] [PubMed] [Google Scholar]
- Hofsteenge J, Löffler A, Müller DR, Richter WJ, de Beer T, Vliegenthart JFG. Techniques in protein chemistry VII. D.R. Marshak, New York: Academic Press; 1996. Protein C-glycosylation; pp. 163–171. [Google Scholar]
- Kerscher F, Franz G. Biosynthesis of vitexin and isovitexin: enzymatic synthesis of the C-glucosylflavones vitexin and isovittexin with an enzyme preparation from Fagopyrum esculentum m. seedlings. Z Naturforsch. 1987;42:519–524. [Google Scholar]
- Kerscher F, Franz G. Isolation and some properties of an UDP-glucose: 2-hydroxyflavanone-6 (or 8)-C-glucosyltransferase from Fagopyrum esculentum m. cotyledons. J Plant Physiol. 1988;132:110–115. [Google Scholar]
- Krieg J, Gläsner W, Vincentini A, Doucey M-A, Löffler A, Hess D, Hofsteenge J. Protein C-mannosylation is an intracellular process, performed by a variety of cultured cells. J Biol Chem. 1997a;272:26687–26692. doi: 10.1074/jbc.272.42.26687. [DOI] [PubMed] [Google Scholar]
- Krieg J, Hartmann S, Vicentini A, Gläsner W, Hess D, Hofsteenge J. Recognition signal for C-mannosylation of Trp-7 in RNase 2 consists of the sequence Trp-Xaa-Xaa-Trp. Mol Biol Cell. 1998;9:301–309. doi: 10.1091/mbc.9.2.301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Löffler A, Doucey M-A, Jansson AM, Müller DR, de Beer T, Hess D, Meldal M, Richter WJ, Vliegenthart JFG, Hofsteenge J. Spectroscopic and protein chemical analyses demonstrate the presence of C-mannosylated tryptophan in intact human RNase 2 and its isoforms. Biochemistry. 1996;35:12005–12014. doi: 10.1021/bi9610515. [DOI] [PubMed] [Google Scholar]
- Mosimann SC, Newton DL, Youle RJ, James MNG. X-ray crystallographic structure of recombinant eosinophil-derived neurotoxin at 1.83 Å resolution. J Mol Biol. 1996;260:540–552. doi: 10.1006/jmbi.1996.0420. [DOI] [PubMed] [Google Scholar]
- Oliver GJA, Harrison J, Hemming FW. The mannosylation of dolichol-diphosphate oligosaccharides in relation to the formation of oligosaccharides and glycoprteins in pig-liver endoplasmic reticulum. Eur J Biochem. 1975;58:223–229. doi: 10.1111/j.1432-1033.1975.tb02367.x. [DOI] [PubMed] [Google Scholar]
- Pisano A, Redmond JW, Williams KL, Gooley AA. Glycosylation sites identified by solid-phase edman degradation: O-linked glycosylation motifs on human glycophorin A. Glycobiology. 1993;3:429–435. doi: 10.1093/glycob/3.5.429. [DOI] [PubMed] [Google Scholar]
- Rosenwald AG, Stoll J, Krag SS. Regulation of glycosylation. J Biol Chem. 1990;265:14544–14553. [PubMed] [Google Scholar]
- Singh DG, Lomako J, Lomako WM, Whelan WJ, Meyer HE, Serwe M, Metzger JW. Beta-Glucosylarginine: a new glucose-protein bond in a self-glucosylating protein from sweet corn. FEBS Lett. 1995;376:61–64. doi: 10.1016/0014-5793(95)01247-6. [DOI] [PubMed] [Google Scholar]
- Singh N, Tartakoff AM. Two different mutants blocked in synthesis of dolichol-phosphoryl-mannose do not add glycolipid anchors to membrane proteins: quantitative correction of the phenotype of a CHO cell mutant with tunicamycin. Mol Cell Biol. 1991;11:391–400. doi: 10.1128/mcb.11.1.391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoll J, Robbins AR, Krag SS. Mutant of chinese hamster ovary cells with altered mannose 6-phosphate receptor activity is unable to synthesize mannosylphosphoryldolichol. Proc Natl Acad Sci USA. 1982;79:2296–2300. doi: 10.1073/pnas.79.7.2296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoll J, Rosenberg L, Carson DD, Lennarz WJ, Krag SS. A single enzyme catalyzes the synthesis of the mannosylphosphoryl derivative of dolichol and retinol in rat liver and chinese hamster ovary cells. J Biol Chem. 1985;260:232–236. [PubMed] [Google Scholar]
- Stoll J, Cacan R, Verbert A, Krag SS. Lec15 cells transfer glucosylated oligosaccharides to protein. Arch Biochem Biophys. 1992;299:225–231. doi: 10.1016/0003-9861(92)90268-2. [DOI] [PubMed] [Google Scholar]
- Vicentini A, Hemmings BA, Hofsteenge J. Residues 36–42 of liver RNase PL3 contribute to its uridine-preferring substrate specificity. Protein Sci. 1994;3:459–466. doi: 10.1002/pro.5560030311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waechter CJ, Lucas JJ, Lennarz WJ. Membrane glycoproteins. J Biol Chem. 1973;248:7570–7579. [PubMed] [Google Scholar]
- Waechter CJ, Lennarz WJ. The role of polyprenol-linked sugars in glycoprotein synthesis. Annu Rev Biochem. 1976;45:95–111. doi: 10.1146/annurev.bi.45.070176.000523. [DOI] [PubMed] [Google Scholar]
- Wilm M, Mann M. Analytical properties of the nanoelectrospray ion source. Anal Chem. 1996;68:1–8. doi: 10.1021/ac9509519. [DOI] [PubMed] [Google Scholar]