

Abstract
We studied whether temporal sequences can be learned implicitly using a process dissociation procedure (PDP). Participants performed repeated serial recalls of sequential stimuli with a random ordinal structure and fixed temporal structure. Explicit knowledge was evaluated through verbal questions and PDP analysis of two generation tasks (inclusion and exclusion). Participants were divided into two groups: in the Ordinal group, stimulus presentation was visual and the participants were instructed to repeat the ordinal structure; in the Temporal+Ordinal group, stimulus presentation was audio-visual and the participants were instructed to repeat temporal and ordinal structure. We expected predominantly implicit learning in the Ordinal group and explicit learning in the Temporal+Ordinal group. This was supported by two findings. First, a significant difference between inclusion and exclusion performance was seen only in the Temporal+Ordinal group. Second, in both groups, a negative relation was found between the degree of improvement during serial recall and a measure of explicit knowledge in the generation tasks. This relation was independent of the final level of performance during serial recall. These findings suggest that distinct implicit and explicit systems may exist for learning of temporal sequences: implicit learning is gradual and gives rise to knowledge that is inaccessible to conscious control while the explicit system is fast and results in representations that can be used to control performance in inclusion and exclusion tasks.
INTRODUCTION
Implicit learning is classically defined as learning occurring without conscious awareness of what is being learned (Reber 1967; Stadler and Frensch 1997). This phenomenon has been studied extensively for different forms of sequential skill learning. In the simplest type of sequence, the deterministic sequence, the elements have a fixed ordinal structure (serial order). Most studies of implicit learning of such sequences have used the serial reaction time task (SRTT). The SRTT consists of series of choice reaction time trials. The serial order of the stimuli can be manipulated, unbeknownst to the participant, and learning is observed as decreases in reaction time (see Nissen and Bullemer 1987; Willingham et al. 1993). Deterministic sequence learning in the SRTT presumably involves forming element–element associations between stimuli and between responses, as indicated by transfer studies (Mayr 1996; Seger 1997; Willingham et al. 1989) and designs where the stimulus–response mapping is randomized from trial to trial (Dennis et al. 2006; Goschke et al. 2001). Both the length and the structure of the sequence influence learning: implicit learning is more difficult for longer sequences as well as for sequences that do not contain any unique associations between two consecutive elements (e.g.: 43, but not 14, is a unique association in the sequence 1431343; Cohen et al. 1990).
Further insights into the nature of the learned sequence representations have come from studies of probabilistic sequence learning. Probabilistic sequences are generated by processes that include stochastic elements and thus lack a fixed serial order. Implicit learning has been reported for probabilistic sequences of different complexity, ranging from relatively simple types where the conditional probability that a particular element follows a preceding run of two or more elements is controlled (Hunt and Aslin 2001; Stadler 1992), to complex sequences generated by “noisy” finite state machines (Cleeremans and McClelland 1991). Participants seem able to use different statistics to improve performance, including absolute element frequencies and transitional probabilities between elements (Hunt and Aslin 2001; Lungu et al. 2004).
A large literature thus suggests that humans can extract and use information about the ordinal structure of sequential stimuli, with little or no awareness of the obtained knowledge. Dissociating implicit and explicit learning faces notorious methodological difficulties, however (see Stadler and Frensch 1997 and Shanks and St. John 1994 with commentaries). In general, the operational definition of explicit learning will obviously depend on which awareness test is used. A multitude of such tests have been used in the literature. Two serious concerns have been raised in this context. First, the awareness test may be too insensitive. Verbal questions may, for instance, fail to detect partial explicit knowledge. Second, the test may not be process pure; i.e., a test intended to measure only explicit knowledge—e.g., a generation task—may be influenced by implicit knowledge to an unknown degree.
The process dissociation procedure (PDP) is an approach to measuring explicit knowledge, which avoids assumptions about process purity. The basic idea (Jacoby 1991) is to design two tasks: one in which explicit and implicit knowledge both facilitate performance and one in which explicit knowledge facilitates performance while implicit knowledge acts as a source of interference. The difference in level of performance in the two tasks can be used as an estimate of obtained explicit knowledge. The PDP has been adapted to study implicit and explicit learning of deterministic ordinal sequences in the SRTT by Destrebecqz and Cleeremans (2001). Participants were divided into two groups with response-to-stimulus intervals (RSIs) of 0 or 250 ms to inhibit or facilitate explicit learning. After training, the participants were given two generation tasks. In the inclusion task, participants were asked to generate as much of the trained sequence as they remembered, whereas in the exclusion task, they were instructed to generate sequences that were different from the trained one. Both groups performed above chance on the inclusion task. However, the 0-ms RSI group was unable to exclude the trained sequence in the exclusion task. The likelihood of reproducing elements of the trained sequence in this group was in fact the same for both inclusion and exclusion, suggesting that the participants had gained sequence knowledge that was inaccessible to intentional control, i.e., implicit. Since then, explicit ordinal sequence learning with 0-ms RSI has been found in a number of experiments (Wilkinson and Shanks 2004: experiments 1–3; Destrebecqz et al. 2005; Norman et al. 2006; Shanks et al. 2005: experiment 3), which indicates that insufficient power may have been an issue in the pioneering study by Destrebecqz and Cleeremans (2001). Neverthelss, the PDP remains a useful design to evaluate explicit and implicit knowledge in the SRTT.
The large majority of studies on implicit sequence learning have focused on the learning of ordinal structures. Only a few published studies have directly addressed whether the other dimension of a sequence, its temporal structure—i.e., the pattern of temporal intervals between the elements—can be learned implicitly. Salidis (2001) used a modification of the auditory SRTT where the RSIs in a series of simple reaction time trials followed a temporal sequence. A decrease of mean reaction time was seen, with no signs of explicit rhythm knowledge on awareness tests, which included verbal questions and a generation test where the participants represented the temporal sequence as a series of numbers. Shin and Ivry (2002) studied both ordinal and temporal learning in the SRTT. Temporal learning was only found when the temporal sequence was coupled to an ordinal sequence. Verbal questions indicated that the acquired knowledge on temporal regularities was implicit. Ullén and Bengtsson (2003) found learning of temporal sequences in a repeated immediate serial recall paradigm where the ordinal structure of the stimuli was random. Learning was seen also in subjects who verbally reported having detected no regularities in the stimuli. Although these studies indicate that implicit temporal sequence learning is possible, they can be criticized on the same, abovementioned grounds as many studies of implicit ordinal learning: the used awareness measures may not have been process pure and/or sensitive enough.
The main purpose of this study was to investigate whether stronger evidence for implicit temporal sequence learning can be obtained using a modified version of the PDP used by Destrebecqz and Cleeremans (2001). As we have argued previously (see Ullén and Bengtsson 2003), the SRTT may be less suitable for temporal sequence learning. The training was therefore designed as 50 repeated immediate serial recall trials, where the participants were instructed to reproduce spatiotemporal sequences that had a random ordinal structure and a fixed temporal structure. The participants were divided into two groups that were given different instructions to promote explicit temporal learning in one group but not the other. The group Ordinal was thus asked to reproduce the ordinal structure of the stimuli, whereas the group Temporal+Ordinal was instructed to reproduce the ordinal as well as temporal structure of the stimuli. The sequences were presented visually. To further facilitate explicit learning in the Temporal+Ordinal group, a brief drum sound occurred at the onset of each stimulus in this group. Psychophysical studies show that subjects more easily extract the temporal structure of auditory than of visual stimuli (Repp and Penel 2002). None of the groups were informed about stimulus regularities before training. Procedural learning was estimated from the difference in performance (mean relative error of the reproduced rhythms compared with the stimulus rhythm) in the first and last five trials of the training. After training, explicit knowledge was estimated by verbal questions, as well as by comparing performance in two generation tasks (PDP). In one of these tasks, participants were instructed to freely generate temporal patterns that were as similar as possible to the patterns of training stimuli. In the other task, participants were asked to generate rhythms that were as different as possible to the training stimuli. Explicit learning scores were estimated from the difference in performance between these two tasks. To avoid task order confounds, the order of the two generation tasks was randomized.
We predicted significant procedural learning in both groups, small or absent explicit knowledge in the Ordinal group, and high explicit learning scores in the Temporal+Ordinal group. To further characterize implicit and explicit learning systems, we performed correlational analyses between the degree of improvement during serial recall and explicit learning scores in each group. Finally, the relation between explicit learning scores and the responses to the verbal questions on explicit knowledge was explored through regression analyses.
METHODS
Participants
Forty-three normal subjects (mean age, 28.0 ± 10.1 yr), 15 males and 28 females, gave informed consent to participate in the study. All participants were recruited through advertisement and received payment for participation. The study was approved by the Ethical Committee of Karolinska Institutet (KI Forskningsetikkommitté Nord; Dnr 2007/83–/32). Three participants were excluded: two because of self-reported problems with following the experimental procedure and one because of extreme outliers (generated intervals > 1 min) in the exclusion task. Forty participants where thus included in the analyses.
Materials
The experiment was performed on a PC. Stimulus presentation and data collection were controlled by a script written in the E-Prime software package (Psychological Software Tools). Visual stimuli were presented on the computer monitor, and auditory stimuli were presented through the computer loudspeakers. All visual stimuli were designed using CorelDraw 11 (Corel). The auditory stimulus was a brief (62 ms) recording of a single bongo drum beat downloaded from a database of licensed sounds (http://freesound.iua.upf.edu). Responses were collected from the numerical keypad of the PC keyboard. Processing of behavioral data were performed in MatLab 7 (The MathWorks), and statistical analyses were made in Statistica 7.1 (StatSoft).
Experimental procedures
During the experiment, the participant was seated in front of the computer, at a distance of ∼60 cm from the screen. The experiment consisted of a serial recall task followed by two tests of explicit rhythm knowledge, i.e., verbal questions on stimulus regularities and generation tasks. Participants were randomly assigned to either the Ordinal group and Ordinal/Temporal group, with 20 individuals in each. The task instruction and the modality of the sequence stimuli differed for the ordinal only group and the ordinal/temporal group participants in the serial recall task. In all other respects, the two groups received identical treatment.
SERIAL RECALL TASK.
This task consisted of a number of serial recall trials, where the participants reproduced rhythmic sequences of stimuli. Before the experimental session, participants were given a verbal task instruction. This instruction differed in the two participant groups to facilitate explicit learning in the Temporal+Ordinal group. The Ordinal group was instructed to pay attention to the order of the sequential stimuli and reproduce them, using the right index finger on the four arrow keys of the numerical keypad. The Temporal+Ordinal group was instructed to pay attention to both the order and the temporal pattern of the stimuli and reproduce them. No further instructions were provided. The task started with three practice trials to familiarize the participants with the procedure. After this, 50 trials were performed without interruption. Data from these 50 trials were analyzed.
In each trial, a sequence with an ordinal structure of eight elements and a temporal structure of seven intervals between the elements was presented. The ordinal structure of the sequences was always random. Four different elements were used, corresponding to the four arrow keys of the numerical keypad. These four elements had equal probability, with the constraint that the same element never appeared twice or more in succession. The temporal structure was always the same: 375-375-750-375-750-375-750 (all in ms). An example of a sequence is shown in Fig. 1A. During the presentation of a stimulus sequence, the keys of the numerical keypad were shown schematically on the computer screen as four squares (Fig. 1B). At the onset of each element in the sequence, the background of the corresponding square changed color from white to red for 50 ms. For participants of the Temporal+Ordinal group, the stimulus onset was in addition marked with a brief drum sound to further promote explicit learning of the rhythmic pattern.
FIG. 1.

Experimental task. A: an example of a stimulus sequence in the serial recall task. The sequences had a random ordinal structure of 8 elements and a temporal structure of 7 intervals that were always as shown. B: schematic representation of the arrow keys of the numerical keyboard that was used for sequence production.
After the presentation of a sequence the participant attempted to reproduce it, using the right index finger. At each keypress, the background of the corresponding square on the screen changed to red, as during the presentation. After each trial of eight key presses, feedback was provided on the number of correctly reproduced ordinal elements. No feedback was given on the temporal structure of the response.
VERBAL QUESTIONS.
After completion of the serial recall task, participants were asked to answer two verbal questions that were presented consecutively on the computer screen. In one of the questions, the participant had to rate whether she or he experienced the ordinal pattern of the sequences as predictable or random, on a scale from 1 to 4, where 1 was “always random”, 2 was “most of the time random”, 3 was “most of the time predictable”, and 4 was “always predictable.” In the other question, the same scale was used to rate the experienced regularity of the rhythmic pattern of the sequence. The order of the two questions was randomized between participants.
GENERATION TASKS.
Following the verbal questions, the participant was informed that the temporal sequence had always been the same, whereas the ordinal structure had been random. The participant had to perform two free generation tasks. In the inclusion task, the instruction was to generate rhythmical sequences that were similar to the temporal pattern presented in the serial recall task. In the exclusion task, the participant was asked to generate different rhythmical sequences but to try not to include the temporal pattern of the serial recall task. The participant was asked to produce the rhythms on a single arrow key. Each generation task was interrupted after 10 sequences of eight elements had been generated. The order of the two generation tasks was randomized between participants.
Data analysis
In both the serial recall task and the generation tasks, participants produced temporal sequences of seven intervals. As a basic measure of the accuracy of a produced sequence we used the mean absolute relative error of the trial, RE, that is
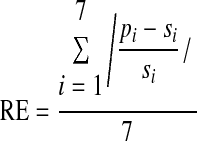 |
(1) |
where pi is the duration of produced interval i and si is the duration of interval i in the stimulus sequence.
Performance at the beginning and end of training was quantified as the mean RE of the first five trials and last five trials, respectively. Improvement during serial recall was defined as the within-subject difference of these two scores. RE was also used to quantify performance in the two generation tasks. A participant's Inclusion score was calculated as the mean RE of all 10 generated sequences in the inclusion task, whereas the Exclusion score was the mean RE of the 10 exclusion task sequences. An assumption of the study (see Introduction) is that both implicit and explicit knowledge decrease Inclusion, whereas Exclusion is increased by explicit knowledge and decreased by implicit knowledge acting as interference. A participant's Explicit learning score was therefore calculated as the difference between these two scores, i.e., Exclusion−Inclusion.
Differences in performance measures in the the serial recall and in the generation task were tested using repeated-measures ANOVA, complemented by post hoc analyses of differences in means with the Tukey HSD test. Group differences in Explicit scores were investigated using Student's t-test. Correlations between measures were tested using Pearson product-moment correlations. Since scores on the verbal questions were on an ordinal scale, within-subject and between-group differences in these variables were studied using the nonparametric Friedman ANOVA by ranks test and Mann-Whitney U test, respectively. Finally, multiple regression was used to study the relation between scores on the verbal questions and performance measures.
RESULTS
Procedural learning in the serial recall task
The performance of the Ordinal and Temporal+Ordinal group in the serial recall task is shown in Fig. 2A. The learning curves show the mean RE of each trial (i.e., reproduction) across all subjects, as a function of trial number, separately for the two groups. Error bars represent SE. A gradual improvement during the first 10–15 trials is seen for the Ordinal group, whereas the Temporal+Ordinal group performed at a high level throughout the task.
FIG. 2.

Performance in the serial recall task. A: learning curves, i.e., performance in the serial recall task, for the Ordinal and the Temporal + Ordinal groups. Mean relative error of each reproduction across all participants is shown as a function of trial number. Error bars represent SE. B: mean relative error in the first (mean of trials 1–5) and last (mean of trials 46–50) blocks of the serial recall task for the Ordinal and the Temporal + Ordinal group. Bars show 95% CIs. The main effects of Group and Block and the Group × Block interaction were all significant.
Three different measures were used to quantify performance. Performance at the beginning of serial recall was quantified as the mean RE of the first five trials of training (First). Performance at the end was measured as the mean RE of the last five trials (Last). First−Last, the within-subject difference between First and Last, was used as a measure of the degree of improvement in the task. We performed a repeated-measures ANOVA (Fig. 2B) with mean RE as the dependent variable, Block (i.e., First or Last) as the within-subject variable, and Group (i.e., Ordinal or Temporal+Ordinal) as the between-subject variable. Significant main effects were seen for Block [F(1,38) = 19.68; P < 0.0001] and Group [F(1,38) = 14.13; P = 0.0005]. There was also a significant Group × Block interaction [F(1,38) = 7.14; P = 0.01]. Post hoc analyses using the Tukey HSD test showed a significant difference between First and Last within the Ordinal group (P = 0.0002) but not within the Temporal+Ordinal group (P = 0.60). A gradual improvement of behavior across trials was thus only seen in the Ordinal group. Post hoc testing of between-group differences in means showed that First was significantly lower in the Temporal+Ordinal group than in the Ordinal group (P = 0.003) but that there was no group difference in Last (P = 0.37). The main effect for Group was thus caused by group differences in performance in the beginning but not the end of serial recall.
Performance on the generation tasks
Performance on the free generation tasks was quantified by three measures. Inclusion scores reflected how different the rhythms of the inclusion task were from the trained rhythm and were calculated as the mean RE of the rhythms in that task. Exclusion scores were calculated as mean RE of the sequences generated in the exclusion task, and Explicit scores as the within-subject difference in Exclusion and Inclusion scores. Inspection of the sequences produced in the exclusion task showed that no participant used simple strategies to avoid reproducing the trained sequence, such as the production of isochronous sequences or extremely long or short intervals. Figure 3 shows an example of a typical performance in the inclusion (Fig. 3A) and exclusion (Fig. 3B) task. All 10 generated rhythms are shown in each task. In the exclusion task, the participant generated a number of patterns, all different from the trained rhythm.
FIG. 3.

Example of a typical performance in the two generation tasks. Each diagram represents the generated rhythm in one trial, with interval number on the x-axis and duration on the y-axis. The scale is the same in all diagrams. A: the 10 trials generated in the inclusion task. B: the 10 trials generated in the exclusion task.
We analyzed the performance in the generation tasks by applying a repeated-measures ANOVA with mean RE as the dependent variable, Task (inclusion or exclusion) as a within-participant variable, and Group (Ordinal or Temporal+Ordinal) as a between-participant variable (Fig. 4A). A significant main effect was seen for Task [F(1,38) = 48.08, P < 0.0001] but not for Group [F(1,38) = 0.01, P = 0.91]. The Group × Task interaction was significant [F(1,38) = 12,22, P = 0.001]. Post hoc analyses showed a significant difference between Inclusion and Exclusion in the Temporal+Ordinal group (Tukey HSD test; P = 0.0001) but only a trend for a difference in the Ordinal group (P = 0.08). A Student's t-test was used to test for group differences in Explicit scores. Explicit scores were significantly higher in the Temporal+Ordinal group [t(38) = 3.5; P = 0.001].
FIG. 4.

Performance in the generation tasks. A: mean relative error in the generation tasks for the Ordinal and Temporal + Ordinal groups. The main effect of Task (inclusion or exclusion) and the Group × Task interaction were significant but not the main effect for group. Bars show 95% CIs. B: correlation between improvement of performance during serial recall and generation task performance for the Ordinal group. C: the same correlation for the Temporal + Ordinal group. The raw correlation had a trend for significance at P = 0.11 but was significant when controlling for performance at the end of serial recall (Last).
Correlations between the serial recall task and the generation tasks
Additionally, the relation between improvement of performance in the serial recall task (First−Last) and performance in the generation tasks (explicit scores) was studied for both groups. For the Ordinal group, a negative correlation (r = −0.52, P = 0.02) was found (Fig. 4B), and for the Temporal+Ordinal group, a similar negative trend was observed (r = −0.36, P = 0.11; Fig. 4C). Performance differences at the beginning of training (First) were more important than final performance (Last) for these correlations. The partial correlation between First−Last and Explicit scores was thus significant when controlling for Last in both the Ordinal (r = −0.52, P = 0.02) and the Temporal+Ordinal group (r = −0.53, P = 0.02). When controlling for First, only trends for negative correlations were found in the Ordinal (r = −0.39, P = 0.1) and the Temporal+Ordinal group (r = −0.30 P = 0.2). Furthermore, negative relations were found between First and Explicit in both groups (Ordinal: trend at r = −0.38, P = 0.1; Temporal+Ordinal: r = −0.48, P = 0.03), whereas Last and Explicit were unrelated (ordinal: r = 0.12, P = 0.62; Temporal+Ordinal: r = −0.16, P = 0.49).
Relation between verbal questions and scores on the generation tasks
Both groups had higher scores on the verbal question about temporal regularities than on the question about regularities in the random ordinal pattern. The scores on the temporal question in the Ordinal group (M = 2, i.e., “most of the time random”) were significantly higher (Friedman ANOVA; χ2 = 6.4; P = 0.01) than the scores on the ordinal question (M = 1, i.e., “always random”). A significant difference (χ2 = 9.0; P = 0.003) with the same median scores on both questions was found in the Temporal+Ordinal group. No between-group differences were seen on either the temporal (Mann-Whitney U = 172; P = 0.45) or the ordinal question (U = 193; P = 0.82).
In the whole dataset, scores on the temporal question were significantly correlated with Explicit scores (r = 0.50, P = 0.001), scores on the ordinal question (r = 0.35, P = 0.03), and initial performance on the serial recall task (First; r = −0.34, P = 0.03). In a multiple regression of temporal question scores on these three variables, the multiple R was 0.58 [F(3,36) = 6.11, P < 0.002]. The regression coefficients were significantly different from zero for Explicit scores [t(36) = 2.81; P = 0.008] and scores on the ordinal question [t(36) = 2.17; P = 0.04] but not for First [t(36) = 0.07; P = 0.95].
DISCUSSION
Independent learning of temporal and ordinal sequences
We studied temporal sequence learning in a repeated serial recall paradigm, where the ordinal stimulus structure was random while the temporal structure was fixed. Although the Ordinal group was not told to reproduce the rhythm, their performance at the end of the serial recall task was not significantly different from the performance of the Temporal+Ordinal group. Temporal sequence learning was found in both groups, thus replicating a main finding of Ullén and Bengtsson (2003) that temporal sequence learning does not require a fixed ordinal structure or explicit instruction to repeat the temporal pattern. In the SRTT, in contrast, temporal learning has been reported to be absent when the ordinal stimulus structure is variable (Shin and Ivry 2002). This may reflect differences between the SRTT and serial recall learning, which make temporal learning more difficult in the former task (Ullén and Bengtsson 2003). In this task, the whole stimulus sequence was presented without interruption before reproduction. In the SRTT, in constrast, the encoding of interstimulus durations coincides with response preparation and generation. This may interfere with temporal encoding. In general, there will probably also be interference between the regular pattern of interstimulus intervals and the interlocked and noisy, i.e., irregular, patterns of stimulus-response, and interresponse intervals. However, another serious concern is that temporal learning with a random ordinal structure in the SRTT may remain undetected: the measure of learning in the SRTT is reaction time, and with a random ordinal structure, decreases in reaction time will be minimal since the participant cannot anticipate the next response even with perfect knowledge of the temporal structure.
Implicit and explicit temporal sequence learning
One prediction in this study was that learning in the Ordinal group would be predominantly implicit, whereas learning in the Temporal+Ordinal group would be explicit. Support for this hypothesis is that only the Temporal+Ordinal group could significantly differentiate their performance in the inclusion and exclusion tasks. Also the Explicit scores that were calculated as the within-subject difference in Exclusion and Inclusion scores were significantly higher in the Temporal+Ordinal group. A trend for a difference between Inclusion and Exclusion was also found in the Ordinal group.
However, we found a negative correlation between improvement in the serial recall task and Explicit scores in both groups, when controlling for final performance level during serial recall. Participants with a large, gradual improvement in the serial recall task thus showed limited ability to differentiate their performance in the inclusion and exclusion tasks. Participants with a constantly accurate performance during serial recall in contrast showed high explicit knowledge.
We argue that these two findings taken together are difficult to explain as solely because of differences in degree of knowledge acquired via a single explicit system. Rather, they suggest the existence of two learning systems giving different kinds of rhythmic knowledge: an implicit system in which learning is gradual and gives rise to rhythm representations that are unavailable to conscious control and an explicit system that is fast and gives rise to consciously accessible rhythm information. An additional possibility is that participants with a fast initial learning reproduced the rhythm correctly a larger number of times, which in turn, promoted explicit learning. In applications of the PDP to ordinal sequence learning in the SRTT, generation task performance is often compared with a baseline (Destrebecqz and Cleeremans 2001; Norman et al. 2006; Wilkinson and Shanks 2004). The similarity of the generation task sequences to the trained sequence is contrasted with the similarity of the generation sequences to a baseline sequence. The baseline sequence has a different ordinal structure than the trained sequence but is identical to the latter in terms of features such as the frequency of elements, first-order transitions, reversals, and repetitions. Performance above baseline allows the inference that participants have learned the second-order transitional structure of the trained sequence. A limitation of this study is that we did not include a baseline. The main reason for this is that the understanding of how temporal sequences are represented is still limited, and it is therefore very difficult to determine what would constitute an adequate temporal baseline sequence. In particular, the representation of temporal sequences is very sensitive to the metrical structure of the sequence (i.e., where stimulus onsets occur in relation to an isochronous beat) (Essens and Povel 1985; Shmulevich and Povel 2000) and a procedure analogous to the one used to construct ordinal baseline sequences is therefore not appropriate. If appropriate temporal baseline tasks can be developed, future studies may give more information regarding which type of patterns within a temporal sequence can be learned implicitly. Another interesting question for further experiments is how implicit and explicit temporal sequence learning depend on the metrical structure of the trained rhythm.
It should also be mentioned that the Ordinal and Temporal+Ordinal groups differed both in task instruction and stimulus modality, which was both auditory and visual in the latter group. It thus cannot be excluded that some group differences may reflect differences in auditory and visual processing of rhythmic stimuli that are unrelated to differences in explicit knowledge between the groups. However, the fact that the within-group correlations Explicit × First and Explicit × Last mimicked between-group differences in First and Last suggests that the observed group differences in these variables mainly are caused by differences in explicit knowledge.
Verbal questions and generation tasks
The verbal question on temporal regularities in the stimuli was a relatively poor predictor of performance in the generation tasks. Notably, the median score on this question was low (2, i.e., “most of the time random”), although participants performed well on the generation tasks. This supports the idea that verbal reports may be relatively insensitive measures of explicit knowledge (Shanks and St. John 1994 and commentaries). Individual differences in explicit learning have earlier been shown to be positively related to general intelligence (Feldman et al. 1995; Waber et al. 2003) and the personality trait openness to feelings, a subscale of the big-five trait openness to experience (Norman et al. 2006). Interestingly, responses to regularities in temporal stimuli in this study were positively related to a tendency to report the random ordinal stimuli as regular. This suggests that individual difference variables may influence responses to verbal questions on regularities, regardless of whether the stimuli were regular or not. This is of course a further reason to be cautious with interpreting such responses as evidence for explicit knowledge.
GRANTS
This work was supported by the Swedish Research Council and the Söderberg Foundation. A. Karabanov was supported by a grant from the National Institutes of Health/Karolinska Institutet Graduate Partnership Program.
Acknowledgments
We thank M. Hallett for discussions and comments on an earlier version of this manuscript.
The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
REFERENCES
- Cleeremans and McClelland 1991.Cleeremans A, McClelland JL. Learning the structure of event sequences. J Exp Psychol Gen 120: 235–253, 1991. [DOI] [PubMed] [Google Scholar]
- Cohen et al. 1990.Cohen A, Ivry RI, Keele SW. Attention and structure in sequence learning. J Exp Psychol Learn Mem Cogn 16: 17–30, 1990. [Google Scholar]
- Dennis et al. 2006.Dennis NA, Howard JH, Howard DV. Implicit sequence learning without motor sequencing in young and old adults. Exp Brain Res 175: 153–164, 2006. [DOI] [PubMed] [Google Scholar]
- Destrebecqz and Cleeremans 2001.Destrebecqz A, Cleeremans A. Can sequence learning be implicit? New evidence with the process dissociation procedure. Psychon Bull Rev 8: 343–350, 2001. [DOI] [PubMed] [Google Scholar]
- Destrebecqz et al. 2005.Destrebecqz A, Peigneux P, Laureys S, Degueldre C, Del Fiore G, Aerts J, Luxen A, Van Der Linden M, Cleeremans A, Maquet P. The neural correlates of implicit and explicit sequence learning: interacting networks revealed by the process dissociation procedure. Learn Mem 12: 480–490, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Essens and Povel 1985.Essens PJ, Povel D-J. Metrical and nonmetrical representations of temporal patterns. Percept Psychophysiol 37: 1–7, 1985. [DOI] [PubMed] [Google Scholar]
- Feldman et al. 1995.Feldman J, Kerr B, Streissguth AP. Correlational analyses of procedural and declarative learning performance. Intelligence 20: 87–114, 1995. [Google Scholar]
- Goschke et al. 2001.Goschke T, Friederici AD, Kotz SA, van Kampen A. Procedural learning in Broca's aphasia: dissociation between the implicit acquisition of spatio-motor and phoneme sequences. J Cogn Neurosci 13: 370–388, 2001. [DOI] [PubMed] [Google Scholar]
- Hunt and Aslin 2001.Hunt RH, Aslin RN. Statistical learning in a serial reaction time task: access to separable statistical cues by individual learners. J Exp Psychol Gen 130: 658–680, 2001. [DOI] [PubMed] [Google Scholar]
- Jacoby 1991.Jacoby LL A process dissociation framework: separating automatic from intentional uses of memory. J Mem Lang 30: 513–541, 1991. [Google Scholar]
- Jenkins et al. 1994.Jenkins IH, Brooks DJ, Nixon PD, Frackowiak RS, Passingham RE. Motor sequence learning: a study with positron emission tomography. J Neurosci 14: 3775–3790, 1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karni et al. 1995.Karni A, Meyer G, Jezzard P, Adams MM, Turner R, Ungerleider LG. Functional MRI evidence for adult motor cortex plasticity during motor skill learning. Nature 377: 155–158, 1995. [DOI] [PubMed] [Google Scholar]
- Lungu et al. 2004.Lungu OV, Wachter T, Liu T, Willingham DT, Ashe J. Probability detection mechanisms and motor learning. Exp Brain Res 159: 135–150, 2004. [DOI] [PubMed] [Google Scholar]
- Mayr 1996.Mayr U Spatial attention and implicit sequence learning: evidence for independent learning of spatial and nonspatial sequences. J Exp Psychol Learn Mem Cogn 22: 350–364, 1996. [DOI] [PubMed] [Google Scholar]
- Nissen and Bullemer 1987.Nissen MJ, Bullemer P. Attentional requirements of learning: evidence from performance measures. Cogn Psychol 19: 1–32, 1987. [Google Scholar]
- Nitsche et al. 2003.Nitsche MA, Schauenburg A, Lang N, Liebetanz D, Exner C, Paulus W, Tergau F. Facilitation of implicit motor learning by weak transcranial direct current stimulation of the primary motor cortex in the human. J Cogn Neurosci 15: 619–626, 2003. [DOI] [PubMed] [Google Scholar]
- Norman et al. 2006.Norman E, Price MC, Duff SC. Fringe consciousness in sequence learning: the influence of individual differences. Conscious Cogn 15: 723–760, 2006. [DOI] [PubMed] [Google Scholar]
- Reber 1967.Reber AS Implicit learning of artificial grammars. J Verbal Learn Verbal Behav 6: 855–863, 1967. [Google Scholar]
- Repp and Penel 2002.Repp BH, Penel A. Auditory dominance in temporal processing: new evidence from synchronization with simultaneous visual and auditory sequences. J Exp Psychol Hum Percept Perform 28: 1085–1099, 2002. [PubMed] [Google Scholar]
- Salidis 2001.Salidis J Nonconscious temporal cognition: learning rhythms implicitly. Mem Cogn 29: 1111–1119, 2001. [DOI] [PubMed] [Google Scholar]
- Seger 1997.Seger CA Two forms of sequential implicit learning. Conscious Cogn 6: 108–131, 1997. [DOI] [PubMed] [Google Scholar]
- Shanks et al. 2005.Shanks DR, Rowland LA, Ranger MS. Attentional load and implicit sequence learning. Psychol Res 69: 369–382, 2005. [DOI] [PubMed] [Google Scholar]
- Shanks and St. John 1994.Shanks DR, St. John MF. Characteristics of dissociable human learning systems. Behav Brain Sci 17: 367–395, 1994. [Google Scholar]
- Shin and Ivry 2002.Shin JC, Ivry RB. Concurrent learning of temporal and spatial sequences. J Exp Psychol Learn Mem Cogn 28: 445–457, 2002. [DOI] [PubMed] [Google Scholar]
- Shmulevich and Povel 2000.Shmulevich I, Povel DJ. Measures of temporal pattern complexity. J New Music Res 29: 61–69, 2000. [Google Scholar]
- Stadler 1992.Stadler MA Statistical structure and implicit serial learning. J Exp Psychol Learn Mem Cogn 18: 318–327, 1992. [Google Scholar]
- Stadler and Frensch 1997.Stadler MA, Frensch PA (Editors). Handbook of Implicit Learning. New York: Sage, 1997.
- Ullén and Bengtsson 2003.Ullén F, Bengtsson S. Independent processing of the temporal and ordinal structure of movement sequences. J Neurophysiol 90: 3725–3735, 2003. [DOI] [PubMed] [Google Scholar]
- Waber et al. 2003.Waber DP, Marcus DJ, Forbes PW, Bellinger DC, Weiler MD, Sorensen LG, Curran T. Motor sequence learning and reading ability: is poor reading associated with sequencing deficits? J Exp Child Psychol 84: 338–354, 2003. [DOI] [PubMed] [Google Scholar]
- Wilkinson and Shanks 2004.Wilkinson L, Shanks DR. Intentional control and implicit sequence learning. J Exp Psychol Learn Mem Cogn 30: 354–369, 2004. [DOI] [PubMed] [Google Scholar]
- Willingham et al. 1993.Willingham DB, Greeley T, Bardone AM. Dissociation in a serial respone time task using a recognition measure: comment on Perruchet and Amorim (1992). J Exp Psychol Learn Mem Cogn 19: 1424–1430, 1993. [Google Scholar]
- Willingham et al. 1989.Willingham DB, Nissen MJ, Bullemer P. On the development of procedural knowledge. J Exp Psychol Learn Mem Cogn 15: 1047–1060, 1989. [DOI] [PubMed] [Google Scholar]