

INTRODUCTION
Intermediate filaments (IFs) are the least understood of the three major cytoskeletal elements. One of the difficulties of studying the functions of IFs in vitro is that the IF proteins are, unlike microtubules (MTs) and microfilaments (MFs), quite insoluble in nondenaturing buffers and the assembly and disassembly of IFs can therefore not be studied under physiological conditions. Despite this limitation, in vitro studies have revealed important aspects of IF assembly (see Herrmann and Aebi, 1998), but they have not been able to address specific questions about IF dynamics in vivo. IFs are known to undergo dynamic changes in distribution and organization during cell growth, polarization, and differentiation. These changes in distribution occur without complete disassembly of IFs into individual subunits. Thus, unlike MTs or MFs, whose dynamics are driven by the concentration of the soluble subunits, IFs are dynamic despite the fact that there is very little IF protein found in the soluble fraction. By injection of rhodamine-labeled vimentin or expression of tagged vimentin, it has been shown that IF turnover occurs along the length of the filaments, unlike the turnover of MTs or MFs, which only add or subtract subunits from the filament ends. One question that arises from these results is whether the turnover of the IF array is the result of an insoluble intermediate and whether one can visualize these intermediates in vivo.
MTs and perhaps other cellular structures are required to form the extended array of IFs in cells. Drugs that induce breakdown of MTs cause collapse of IFs to the perinuclear region (Goldman, 1971; Hynes and Destree, 1978; Wang and Choppin, 1981; Masurovsky et al., 1982; Gurland and Gundersen, 1995). Two separate studies have demonstrated that microinjection of tubulin antibodies promotes collapse of IFs to the perinuclear region (Blose and Feramisco 1984; Gurland and Gundersen, 1995). This apparent association of IFs with MTs has also been shown to require kinesin as a linker molecule, because microinjection of kinesin antibodies (Gyoeva and Gelfand, 1991) or tubulin fragments that interfere with kinesin binding to MTs (Kreitzer et al., 1999) promote collapse of the IF array. Furthermore, a kinesin protein has been shown to bind directly to vimentin filaments in vitro (Liao and Gundersen, 1988). These results suggest that IFs may be extended in cells by MT-based motor proteins.
To study IF assembly and dynamics in vivo, we have used stable cell lines expressing green fluorescent protein (GFP)-vimentin (Ho et al., 1998). We were particularly interested in understanding how IFs are extended in cells and whether intermediates in the turnover and assembly of IFs can be detected. In this assay, we summarize some of our previously published results and include examples that support the idea that IFs move along MTs and that small fragments of IFs may be intermediates in the turnover of the IF array. Since our initial report (Ho et al., 1998), similar findings have been reported from the Goldman laboratory (Prahlad et al. 1998; Yoon et al., 1998).
VIDEO MOVIES
Movie 1: The IF Array Is Dynamic
To analyze the behavior of vimentin IFs in living cells, a chimeric protein was constructed. The enhanced form of GFP was fused to the amino-terminal end of vimentin and expressed in NIH3T3 cells. GFP-vimentin constituted 20% of the cellular vimentin in the stably transfected NIH3T3 cells (for more detail, see Ho et al., 1998). Recordings of >4 h were possible with little photobleaching of GFP fluorescence, making GFP-vimentin a good reporter for observing IF behavior in living cells. For experiments presented in this essay, we studied cells at the edge of a wounded monolayer, because these cells become highly polarized when they migrate into the wound, and we suspect that IFs undergo corresponding rearrangements (Gurland and Gundersen, 1995). Specific conditions have been described in detail by Ho et al. (1998).
Initial low-magnification recordings of GFP-vimentin–expressing cells revealed the dynamic behavior of the IF array. IFs were observed to change curvature and orientation over the course of several minutes. In longer recordings it was possible to visualize a wave-like motion for IFs. Presented in Movie 1 (Ho et al., 1998) is a recording where many of the IFs were seen to shift and exhibit this wavy motion. This recording was 90 min in length (frame interval, 2.5 min). The still figure presented (Figure 1) is the first image from Movie 1, and the arrow indicates an IF that bends in a wave-like motion. It is important to note that larger bundles of IFs did not appear to bend as much as the smaller IF bundles. In general, individual IFs were observed to be extending toward the edge of the cell, while concurrently the entire array appeared to be pulled inward toward the nucleus. The movement toward the nucleus is similar to what has been described as centripetal transport of other cytoskeletal elements (Theriot and Mitchison, 1992; Mikhailov and Gundersen, 1995). The nucleus in this cell moved dramatically at the beginning of the movie and distorted the IFs. Some IFs also appeared to move with the nucleus, suggesting that they may be tethered to the nuclear envelope. This movie illustrates the usefulness of using GFP-vimentin, because these types of movements would not have been observed using other methods.
Figure 1.

Movie 1: The IF array is dynamic. Shown is the behavior of the IF array in an NIH3T3 cell expressing GFP-vimentin. The movements of the IF array in this movie are typical of other arrays we have recorded. Bar, 15 μm.
Movie 2: IFs in a Mitotic Cell Form Cage-like Structures around Dividing Nuclei
The ability to detect IFs in living cells with GFP fusion proteins offered us the opportunity to examine the IF network of mitotic cells in three dimensions using confocal microscopy. To obtain optimal three-dimensional resolution, optical sections of 0.5 μm were scanned eight times. Additionally, to avoid flattening, the cells were imaged in coverslip dishes in which a hole had been punched in the bottom and a coverslip had been placed on the bottom of the dish. Confocal microscopy was performed with a Zeiss LSM 410 scanning laser confocal attachment mounted on a Zeiss Axiovert 100 TV inverted fluorescence microscope (Carl Zeiss, Jena, Germany). The stack of images for all the scanned z-planes was processed using Zeiss LSM software to produce the final three-dimensional image.
The fixed cell shown in Movie 2 was in telophase (Figure 2); from Ho et al., 1998, cover photo). Through the 360° rotation, we clearly see cables of filaments that appear to run through the midbody connecting the two daughter cells. These data are compatible with the idea that the vimentin network does not completely disassemble during mitosis but rather remains in cage-like structures around the dividing nuclear material. The GFP-vimentin stable cell lines should be useful to explore further the dynamics and rearrangements of IFs during mitosis.
Figure 2.

Movie 2: IFs in a mitotic cell form cage-like structures around dividing nuclei. The organization of GFP-vimentin in a dividing NIH3T3 cell is shown. The IF array forms cage-like structures around the daughter nuclei, and it is connected by a bridge.
Movie 3: Individual Vimentin IFs Extend, Retract, and Translocate
The dynamic behavior of the IF array must be dependent on movements of individual IF bundles. To analyze movements of individual filament bundles, we focused on the leading edge of migrating cells (Movie 3, from Ho et al., 1998; Figure 3). During the recording process, the focus was maintained on the edge of the cell, sometimes at the expense of focus on the entire array. The intensity of the image had to be scaled so that the filaments (or bundles of filaments) at the edge of the cell were detectable. Images were processed every 2.5 min over the course of ∼2.5 h. Several types of movements were observed for individual filament bundles in the cells we observed. Extension and retraction of several IFs at the edge of the cell were detected. The average rate of filament extension measured for vimentin IFs was 0.61 μm/min. Translocation, defined as a lateral shift in a filament without obvious change in length, could also be observed for many filaments at the edges of the cells. In addition to the movements of intact IFs, we observed the presence of vimentin fragments at the edge of the cell. Further analysis of vimentin fragments revealed that they also exhibit dynamic behavior. These movements are better described in Movies 4 and 6.
Figure 3.

Movie 3: Vimentin IFs extend, retract, and translocate. IFs at the edge of the cell undergo continuous changes in position. The filaments extend, retract, and shift. Shown is the first frame of the movie, which demonstrates the movements of IFs at the edge of the cell. Bar, 10 μm.
Movie 4: Fountains—the Movements of IF Fragments
In some cells, we observed an abundance of fragments in the cell periphery, particularly at the leading edge of the cell (Movie 4). The movie presented is from the region enclosed in the box in Figure 4D. Movie 4 is almost 50 min, with images acquired every 2 min. This movie shows fragments that moved in a linear manner toward the edge of the cell and then turned and moved back toward the cell body. In this movie, individual fragments were continuously generated during the entire sequence. Sequentially formed fragments followed the same general paths on their way out toward the leading edge of the cell. The overall path followed by these fragments resembled a fountain, and so we have dubbed them “vimentin fountains.” The fragments within these fountains moved at an average rate of 15 μm/min. This is ∼25 times faster than that seen for extension of individual IF bundles. We have seen numerous vimentin fragments in >50% of the cells at the wound edge, suggesting that this is a common feature of IF dynamics in these cells. We do not know whether fragments only form at the edge of the cell or, rather, can only be detected there. On occasion we have observed them along the lateral edges of the cell.
Figure 4.

Movie 4: Fountains—the movements of IF fragments. IF fragments are present in GFP-vimentin–expressing cells and in the parental NIH3T3 cells. (A and B) GFP-vimentin–expressing NIH3T3 cells. (A) GFP fluorescence. (B) Indirect immunofluorescent pattern of the same cells fixed and stained as described by Gurland and Gundersen (1995) using a polyclonal antibody (1274) (Wang et al., 1984). Arrows point to two of the many fragments present. (C) NIH3T3 cells fixed and stained as in B. Arrows point to IF fragments present in the parental cells. (D) First frame of a movie in which the IF fragments display dynamic movements. The movie is from the region enclosed in the box and is played four times. There is a pause only after the first time through the movie. Bars: (A, B, and D) 10 μm; (C) 5 μm.
One concern with the fragments is that they are artifacts produced either by the GFP-vimentin chimera or by photo damage during fluorescent imaging. To address this question, we examined parental NIH3T3 cells that were fixed in −20°C methanol and immunofluorescently stained for endogenous vimentin. The parental cell line exhibited immunofluorescently detectable fragments near the leading edge (Figure 4C) that were similar in number and size to those detected in the GFP-vimentin cell line by either direct GFP fluorescence (Figure 4A) or immunofluorescence (Figure 4B). This demonstrates that GFP fragments were not a product of GFP-vimentin expression or of the imaging conditions.
Movie 5: GFP-Vimentin Fragments—Intermediates of IF Turnover
One possible explanation for the presence of the GFP-vimentin fragments is that they are an intermediate in the turnover of vimentin filaments. In agreement with this possibility, we observed the apparent joining of two fragments onto the end of a single filament bundle (Movie 5A). Figure 5A shows selected frames from a recording in which the fragments have been outlined. Over the course of the 13-min movie, the two fragments were seen to align with a longer IF that appeared to come from the IF array. These aligned fragments then extended together toward the edge of the cell. Although we cannot be certain that the fragments actually annealed into a single IF bundle, the unified extension of these coaligned fragments is consistent with this possibility. It is interesting to speculate that IFs may increase in length through en bloc incorporation of fragments at the ends of filaments and that the fragments we have observed are an intermediate in IF formation.
Figure 5.

Movie 5: GFP-vimentin fragments—possible intermediates in IF turnover. (A) The coalignment of two GFP-vimentin fragments with an IF is observed. Every other frame from the first 10 min of the movie is shown, with the IFs of interest outlined. In the first frame the fragments of interest are outlined as they are in the movie. Bar, 5 μm. (B) We observed the formation of two fragments from a longer fragment. The six frames of the movie are shown. As in A, the fragment(s) of interest are outlined. Bar, 5 μm.
We have also detected the formation of two smaller fragments from a larger IF segment (Movie 5B). At this time we are not able to rule out the possibility that the apparent formation of fragments in this recording may be explained by an initial coalignment of two separate fragments that subsequently separate from each because of movements at different speeds. Yet, it is reasonable to suggest that vimentin fragment intermediates may form through this mechanism. We speculate that if one end of the fragment is immobilized and kinesin pulls on the other, or if there is an uneven distribution of kinesin, small fragments of IFs may be formed. The observations presented in these two movies, although not conclusive, suggest that IF fragments are an important component of IF dynamics, possibly an intermediate, and may be directly involved in IF turnover and rearrangement. Prahlad et al. (1998) observed similar IF fragments, termed squiggles, before the formation of a filamentous IF array in spreading cells.
Movie 6: Movement of GFP-Vimentin Fragments Depends on Intact MTs
There is considerable evidence that MTs play an important role in the distribution of IFs. We wanted to know whether MTs played a direct role in the observed movements of GFP-vimentin filaments or GFP-vimentin fragments that we observed in our recordings. To address this question, we recorded the movements of IFs and fragments before and after perfusion of 20 μm nocodazole. In Movie 6, the effect of nocodazole on both filament extension and fragment movement is presented (Figure 6). The movements of IFs were recorded for ∼20 min during perfusion with nocodazole. Before nocodazole addition, the filaments can be seen extending and moving as in the other movies presented above. Additionally, both outward and inward movements of vimentin fragments were observed. During nocodazole perfusion (4 ml at 1 ml/min), imaging was difficult, and there is a break of ∼4 min between the last frame before perfusion and the first frame after perfusion. After perfusion the recording was continued for another 35 min with images acquired every minute. Almost immediately after perfusion, the dynamic behavior of both the IF bundles and fragments decreased. The filaments no longer extended, and the fragments stopped moving forward toward the edge of the cell. Movement in the direction of the nucleus continued as the IF array began to collapse toward the perinuclear region. This experiment and an earlier one reported by Ho et al. (1998) show that both the slower-moving intact IFs and the faster-moving IF fragments depend on MTs for their outward movements.
Figure 6.
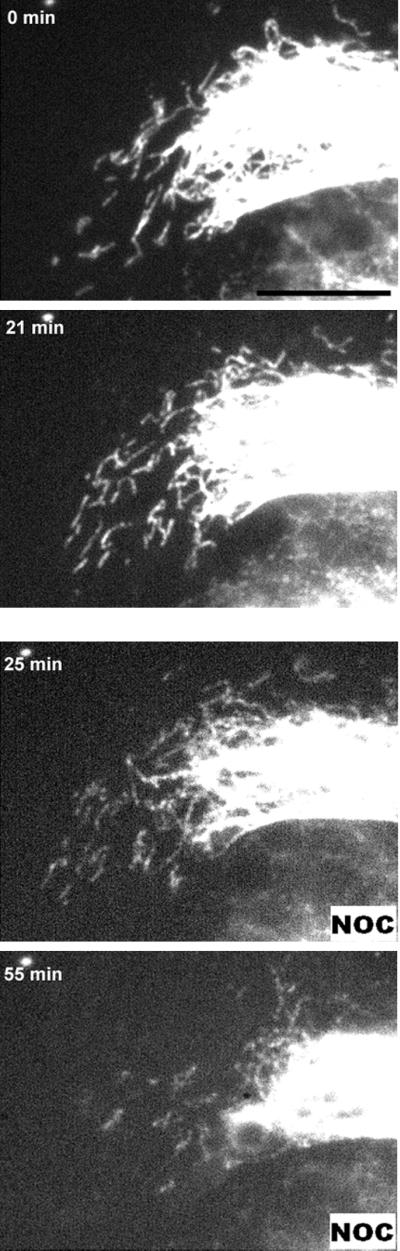
Movie 6: Movement of GFP-vimentin fragments depends on intact MTs. Vimentin fragments stop moving in the presence of nocodazole. Shown are a number of fragments at the edge of the cell. The fragments move during the first part of the recording (before perfusion), as shown by the change in fragment location from 0 to 21 min. After perfusion of 20 μM nocodazole (NOC, bottom right), the fragments stop moving toward the edge of the cell and are only seen moving away from the edge, as is shown by the different fragment distributions at 25 and 55 min. Bar, 10 μm.
Movie 7: Collapse of the IF Array Occurs through Redistribution of Intact Filaments
There are a number of cellular treatments (microinjection of antibodies to IFs, tubulin, or kinesin or treatment with MT antagonists) that promote collapse of the IF array to a perinuclear location (Goldman, 1971; Hynes and Destree, 1978; Wang and Choppin, 1981; Klymkowsky, 1982; Masurovsky et al., 1982; Blose and Feramisco, 1984; Gyoeva and Gelfand, 1991; Gurland and Gundersen, 1995). From these earlier studies, it was unclear whether the rearrangement represented a redistribution of intact filaments or some dynamic disassembly–reassembly process as has been observed for MTs and MFs. In Movie 7, we examined the collapse of the IF array, in real time, by microinjecting anti-IFA antibody, which had previously been shown to collapse IFs (Klymkowsky, 1982) (Figure 7). Cells at a wound edge were injected with anti-IFA and imaged as previously described (see Ho et al., 1998). The arrow in the still image presented as the start of the movie points to an IF that is observed to move in bulk to an area on the other side of the nucleus. The GFP-vimentin IF array did not disassemble but instead appeared to be moved back toward the center of the cell. Control experiments (see Ho et al., 1998) demonstrated that the cell remained spread and extended and that the MT array was not disrupted. Although anti-IFA can induce disassembly of IFs in vitro, the use of time-lapse recording in this experiment demonstrates that the collapse of IFs to a perinuclear area in vivo involves bulk movement of the IFs rather than a disassembly–reassembly process.
Figure 7.

Movie 7: Collapse of the IF array occurs through redistribution of intact filaments. Anti-IFA antibody was microinjected into cells expressing GFP-vimentin, and the resultant redistribution of the IF array was recorded. Bar, 10 μm.
SUMMARY
In the movies presented in this essay we have demonstrated that IFs are dynamic in nature. Movements of IFs include both extension and retraction and are dependent on the presence of intact MTs. Furthermore, we have characterized a possible intermediate of IF turnover, IF fragments. These fragments are also dynamic and can be visualized moving at the leading edge of cells migrating into a wound edge. Interestingly, the movement of these fragments is also dependent on intact MTs, suggesting that MTs may play a role in turnover of IFs.
Supplementary Material
ACKNOWLEDGMENTS
The confocal images were taken by Teresa Swayne. Her efforts are greatly appreciated. This work was supported by National Institutes of Health grants NS-15182 (to R.K.H.L.) and GM-42026 (to G.G.G.). J.L.M. is a postdoctoral trainee supported by National Research Service Award training grant GM-19695.
Footnotes
Online version of this essay contains video material for Figures 1–7. Online version available at www.molbiocell.org.
REFERENCES
- Blose SH, Meltzer DI, Feramisco JR. 10-nm filaments are induced to collapse in living cells microinjected with monoclonal and polyclonal antibodies against tubulin. J Cell Biol. 1984;98:847–858. doi: 10.1083/jcb.98.3.847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldman RD. The role of three cytoplasmic fibers in BHK-21 cell motility. I. Microtubules and the effects of colchicine. J Cell Biol. 1971;51:752–762. doi: 10.1083/jcb.51.3.752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gurland G, Gundersen GG. Stable, detyrosinated microtubules function to localize vimentin intermediate filaments in fibroblasts. J Cell Biol. 1995;131:1275–1290. doi: 10.1083/jcb.131.5.1275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gyoeva FK, Gelfand VI. Coalignment of vimentin intermediate filaments with microtubules depends on kinesin. Nature. 1991;353:445–448. doi: 10.1038/353445a0. [DOI] [PubMed] [Google Scholar]
- Herrmann H, Aebi U. Intermediate filament assembly: fibrillogenesis is driven by decisive dimer-dimer interactions. Curr Opin Struct Biol. 1998;8:177–185. doi: 10.1016/s0959-440x(98)80035-3. [DOI] [PubMed] [Google Scholar]
- Ho C-L, Martys JL, Mikhailov A, Gundersen GG, Liem RKH. Novel features of intermediate filament dynamics revealed by green fluorescent protein chimeras. J Cell Sci. 1998;111:1767–1778. doi: 10.1242/jcs.111.13.1767. [DOI] [PubMed] [Google Scholar]
- Hynes RO, Destree AT. 10 nm filaments in normal and transformed cells. Cell. 1978;13:151–163. doi: 10.1016/0092-8674(78)90146-0. [DOI] [PubMed] [Google Scholar]
- Klymkowsky MW. Vimentin and keratin intermediate filament systems in cultured PtK2 epithelial cells are interrelated. EMBO J. 1982;1:161–165. doi: 10.1002/j.1460-2075.1982.tb01141.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreitzer G, Liao G, Gundersen GG. Detyrosination of tubulin regulates the interaction of intermediate filaments with microtubules in vivo through a kinesin-dependent mechanism. Mol Biol Cell. 1999;10:1105–1118. doi: 10.1091/mbc.10.4.1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamb N, Fernandes A, Feramisco JR, Welch WJ. Modulation of vimentin containing intermediate filament distribution and phosphorylation in living fibroblasts by the cAMP-dependent protein kinase. J Cell Biol. 1989;108:2409–2422. doi: 10.1083/jcb.108.6.2409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao G, Gundersen GG. Kinesin is a candidate for cross-bridging microtubules and intermediate filaments. J Biol Chem. 1988;273:9797–9803. doi: 10.1074/jbc.273.16.9797. [DOI] [PubMed] [Google Scholar]
- Masurovsky EB, Peterson ER, Crain SM, Horowitz SB. Taxol-induced microtubule formations in fibroblasts of fetal mouse dorsal root ganglion-spinal cord cultures. Biol Cell. 1982;46:213–216. [Google Scholar]
- Mikhailov AL, Gundersen GG. The centripetal transport of microtubules in motile cells. Cell Motil Cytoskeleton. 1995;32:173–186. doi: 10.1002/cm.970320303. [DOI] [PubMed] [Google Scholar]
- Prahlad V, Yoon M, Moir RD, Vale RD, Goldman RD. Rapid movements of vimentin on microtubule tracks: kinesin-dependent assembly of intermediate filament networks. J Cell Biol. 1998;143:159–170. doi: 10.1083/jcb.143.1.159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theriot JA, Mitchison TS. Comparison of actin and cell surface dynamics in motile fibroblasts. J Cell Biol. 1992;119:367–377. doi: 10.1083/jcb.119.2.367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang E, Cairncross JG, Liem RK. Identification of glial filament protein and vimentin in the same intermediate filament system in human glioma cells. Proc Natl Acad Sci USA. 1984;81:2102–2106. doi: 10.1073/pnas.81.7.2102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang E, Choppin PW. Effect of vanadate on intracellular distribution and function of 10-nm filaments. Proc Natl Acad Sci USA. 1981;78:2363–2367. doi: 10.1073/pnas.78.4.2363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon M, Moir RD, Prahlad V, Goldman RD. Motile properties of vimentin intermediate filament networks in living cells. J Cell Biol. 1998;143:147–157. doi: 10.1083/jcb.143.1.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.