

Despite intensive efforts towards education, cocaine abuse continues to be prevalent and its escalation into a major medical and social problem has degraded the fabric of society in many countries the world over. Recent surveys for the United States indicated that more than 34 million people have tried cocaine, including an estimated 2.1 million considered regular users.1 A myriad of medical problems, including death, often accompany cocaine use and the association of the drug with the spread of AIDS is of concern.2 Furthermore, detrimental effects may be especially tragic for pregnant women where “crack” has been reported as the most abused illicit drug.3 There is no doubt that cocaine abuse has insidious and far-reaching consequences on health, the legal system, and the family unit.
Protein-based therapies for cocaine addiction have received considerable attention in recent years.4 Of particular interest are catalytic antibodies and enzymes specific for the hydrolysis of the benzoyl ester of cocaine 1, returning the nonpsychoactive products benzoic acid and methyl ecgonine 2 (Scheme 1). Several catalytic antibodies have been developed for this reaction,5 including GNL3A66 and GNL7A17 from our laboratory, as well as enzymes such as butyrylcholinesterase (BuChE)8 and the phage-displayed cocaine esterases cocE-pHI and -pIX.9 However, the efficacy of any treatment for cocaine addiction is dependent on limiting potential deleterious side reactions. Antibodies have long been touted as possessing exquisite specificity for their cognate antigens. Yet, while heteroclitic interactions are well known for binding antibodies, 11 no catalytic antibody has been reported to display activity for two distinct substrates of clinical relevance. With antibody therapies rapidly entering human testing, combined with the aftermath of the recent TGN1412 clinical trial disaster,12 potential therapies must be carefully scrutinized for adverse clinical consequences.
Scheme 1.
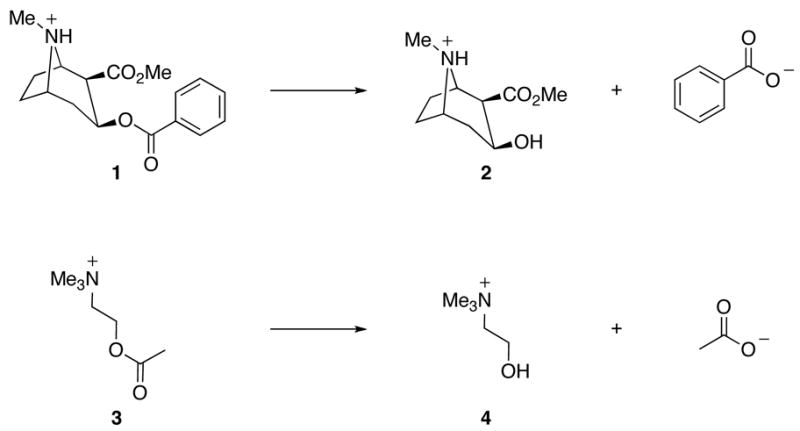
The products of enzymatic hydrolysis of cocaine 1 and acetylcholine 3.
Given that the common neurotransmitter acetylcholine 3 (ACh) and cocaine both contain a cationic nitrogen and reactive ester functionality, we became intrigued in the possibility that antibodies and enzymes displaying cocaine esterase activity might also hydrolyze ACh. Support for this hypothesis can be attained by an examination of the low energy conformations of these two molecules; indeed, in both molecules the nitrogen atom and catalytically cleaved ester moieties are separated by ~5.1 Å. Furthermore, crystallographic studies have revealed that cation-π interactions between aryl residues and the cationic tropane amine of cocaine are key elements of substrate binding for both GNL7A17 and cocE,10 suggesting that the quaternary nitrogen of ACh may similarly bind to these enzymes in a productive orientation.
To test this hypothesis, in silico binding studies were performed with GNL7A1 and cocE by determining the potential binding sites, followed by ligand docking in MPSim-Dock.13 Crystallographic studies suggest that the catalytic activity of GNL7A1 results from a conformational change, thereby stabilizing the generated tetrahedral intermediate.7 Thus, the computational model of GNL7A1 was first validated by comparing the docked cocaine transition state analog with the crystal structure (Figure 1A & 1C); gratifyingly, these two molecules displayed a high degree of overlap (RMDS = 0.384 Å). To examine if the antibody would similarly stabilize the transition state for ACh hydrolysis, a tetrahedral approximation of the ACh transition state was docked into the crystal structure of 7A1 Fab′ bound to a cocaine transition state analog (accession code 2AJX). Using this model, ACh was found to bind at the same site on the catalytic antibody and in a very similar conformation as cocaine (Figure 1A–D). In the lowest-energy structure, the quaternary nitrogen of ACh makes cation-π contacts to the same residues as cocaine, and docks only 1.3 Å away from the tropane nitrogen. Other residues such as TyrL94 and TyrH50 interact with the tetrahedral intermediate of both ACh and cocaine.
Figure 1.
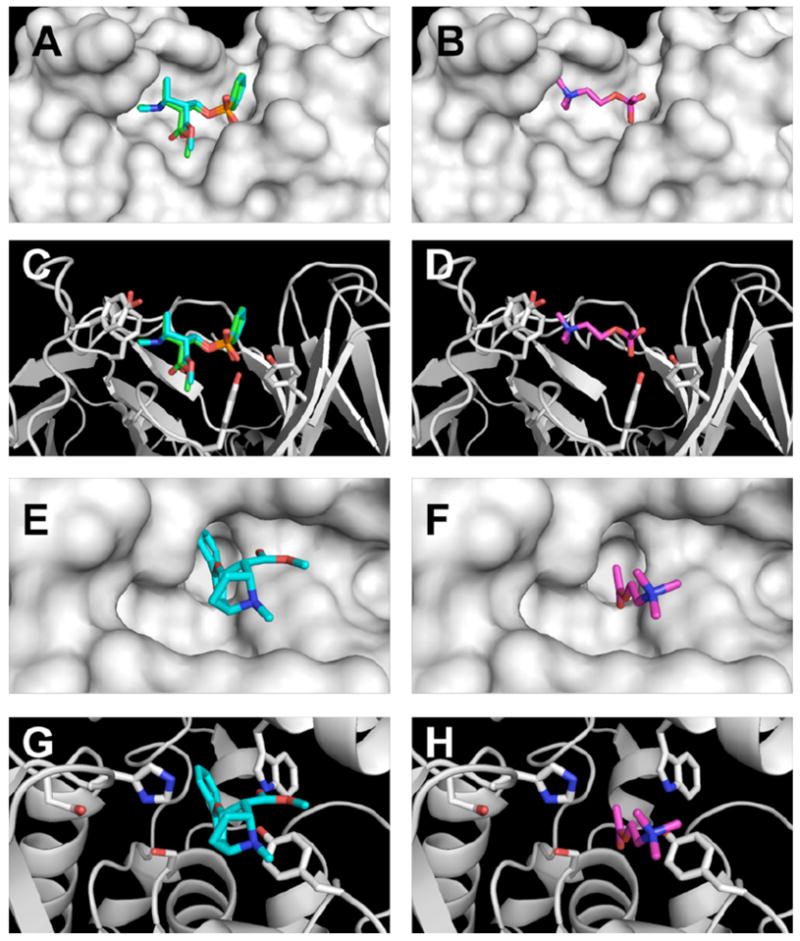
The docked (cyan) and crystal structure (green) of a cocaine transition state analog in 7A1 Fab′ shown in surface (A) and ribbon (C) view, and the structure of an ACh transition state analog in 7A1 Fab′ (B, D). Cocaine (E, G) and ACh (F, H) dock into the same site of cocE.
As with GNL7A1, cocE also has been crystallized in multiple conformations.10 For our studies, we chose the crystal structure of cocE bound to its product, benzoic acid (accession code 1JU4), as no structural data for cocaine-bound cocE has been reported. As was previously observed in the catalytic antibody dockings, the lowest-energy structure of cocaine binds to the active site of cocE (Figure 1E & 1G). The methyl and benzoyl esters of cocaine make hydrogen bond contacts with Trpl66 and His287 respectively, and the tropane nitrogen makes a cation-π interaction with Tyr44. The catalytic Ser117 also is in proximity with the reactive benzoyl carbon. The validity of the docked structure is supported by the high degree of overlap between the benzoyl moiety of cocaine and benzoic acid of the crystal structure.10 Replacement of cocaine with acetylcholine revealed that these two molecules bind cocE in similar conformations, and make similar contacts to the enzyme (Figure 1E–H). Specifically, the quaternary nitrogen of acetylcholine docks only 0.78 Å away from the tropane nitrogen of cocaine and the respective carbonyl carbons are separated by 1.02 Å.
Encouraged by the results of these in silica studies, we tested three cocaine esterase catalytic antibodies, GNL7A1, GNL3A6 and GNL4D3, as well as cocE for acetylcholinesterase activity using Ellman’s colorimetric assay.14 As expected from our docking simulations, all proteins displayed acetylcholine hydrolysis activity consistent with standard Michaelis-Menten kinetics (Table 1). Perhaps more surprising, all antibodies hydrolyzed acetylthiocholine (ATCh) with comparable rate enhancement to that of cocaine. This phenomenon was not due to non-specific IgG effects, as neither the cocaine-binding monoclonal antibody GNC92H215 nor catalytic antibody GNL23A66 hydrolyzed ATCh at enhanced rates relative to background reactions. In all cases, the Km for ATCh was much higher than the Km for cocaine, likely due to the loss of van der Waals contacts with the smaller substrate. An increase in Km was most pronounced for cocE, which has the highest specificity for cocaine; however, the endogenous enzyme butyrylcholinesterase (BuChE) has a comparable Km for acetylthiocholine (Km = 0.5 mM).16 Interestingly, it is as a result of this relatively promiscuous substrate binding that BuChE has been implicated in a wide range of physiological roles including organophosphate detoxification, Alzheimer’s disease, and co-regulation of cholinergic neurotransmission.17
Table 1.
Kinetic Parameters of ATCh and Cocaine Hydrolysis.
| Acetylthiocholine | Cocainea | |||||||
|---|---|---|---|---|---|---|---|---|
| catalyst | kcat (min−1) | Km (μM) | kcat/Km (M−1 s−1 | kcat/kuncat | kcat (min−1) | Km (μM) | kcat/Km (M−1 s−1) | kcat/kuncat |
| GNL3A6 | 0.016 ± 0.002 | 910 ± 220 | 0.29 ± 0.08 | 3.0×l02 ± 0.3×l02 | 0.03 | 55 | 9.2 | 3.7×l03 |
| GNL7A1 | 0.010 ± 0.001 | 1250 ± 320 | 0.13 ± 0.04 | 1.9×l02 ± 0.2×l02 | 0.025 ± 0.001 | 750 ± 37 | 0.56 ± 0.04 | 2.7×l03 ± 0.l×l03 |
| GNL4D3 | 0.014 ± 0.001 | 960 ± 110 | 0.23 ± 0.03 | 2.5×l02 ± 0.2×l02 | 0.01 | 70 | 2.3 | 1.2×l03 |
| cocE | 1207 ± 24 | 8430 ±270 | 2.4×l03 ± 0.1×l03 | 2.2×l07 ± 0.1×107 | 468 ± 6 | 0.64 ± 0.02 | 1.2×l07 ± 0.1×107 | 5.0×107 ± 0.1×107 |
We suggest these results argue that the ability to hydrolyze ACh may be inherent for all cocaine esterases. Additionally, we note that the catalytic activity of a cocaine esterase for ATCh hydrolysis closely tracks with the kinetics of cocaine cleavage; the most proficient cocaine esterase (i.e., cocE) displays the most rapid ATCh hydrolysis. While this finding does not preclude the use of cocaine esterases in the treatment of drug dependence, it does argue for future studies directly examining the effects of these proteins in vivo on the cholinergic nervous system. Furthermore, given the favorable kinetic parameters of cocE relative to BuChE for ACh hydrolysis, this enzyme may also prove useful as a scavenger in the treatment of exposure to agents of chemical warfare.18 With a large number of monoclonal antibodies currently in clinical trials, our study highlights the vital importance of thoroughly examining specificity and reactivity prior to human administration.
Supplementary Material
Acknowledgments
We gratefully acknowledge Dr. Neil C. Bruce of the University of Cambridge for the generous gift of the cocaine esterase vector pCocE, Peter M. Clark and Dr. William A. Goddard, III for assistance with docking studies, and Jenny Mee for assistance with cocE preparation. This work was supported by the National Institutes of Health (DA 08590 and DA 17228) and the Skaggs Institute for Chemical Biology.
Footnotes
Supporting Information Available: Experimental procedures and additional figures for docking and kinetics studies. This material is available free of charge via the Internet at http://pubs.acs.org/.
References
- 1.Office of Applied Studies, Substance Abuse and Mental Health Services Administration, The NSDUH Report, August 12, 2005.
- 2.(a) Benowitz NL. Pharmacol Toxicol. 1993;72:3–12. doi: 10.1111/j.1600-0773.1993.tb01331.x. [DOI] [PubMed] [Google Scholar]; (b) Cregler LL, Herbert M. New Engl J Med. 1986;315:1495–1500. doi: 10.1056/NEJM198612043152327. [DOI] [PubMed] [Google Scholar]
- 3.Frank DA, Zuckerman B, Amaro H, Aboagye K, Bauchner H, Cabral H, Freid L, Hingson R, Kayne H, Levenson SM, Parker S, Reece H, Vinci R. Pediatrics. 1988;82:888–895. [PubMed] [Google Scholar]
- 4.Cashman JR. NIDA Res Monogr. 1997;173:225–258. [PubMed] [Google Scholar]
- 5.Landry DW, Zhao K, Yang GXP, Glickman M, Georgiadis TM. Science. 1993;259:1899–1901. doi: 10.1126/science.8456315. [DOI] [PubMed] [Google Scholar]
- 6.Matsushita M, Hoffman TZ, Ashley JA, Zhou B, Wirsching P, Janda KD. Bioorg Med Chem Lett. 2001;11:87–90. doi: 10.1016/s0960-894x(00)00659-4. [DOI] [PubMed] [Google Scholar]
- 7.Zhu X, Dickerson TJ, Rogers CJ, Kaufmann GF, Mee JM, McKenzie KM, Janda KD, Wilson IA. Structure. 2006;14:205–216. doi: 10.1016/j.str.2005.10.014. [DOI] [PubMed] [Google Scholar]
- 8.Gorelick DA, Gardner EL, Xi ZX. Drugs. 2004;64:1547–1573. doi: 10.2165/00003495-200464140-00004. [DOI] [PubMed] [Google Scholar]
- 9.Rogers CJ, Mee JM, Kaufmann GF, Dickerson TJ, Janda KD. J Am Chem Soc. 2005;127:10016–10017. doi: 10.1021/ja053086a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Larsen NA, Turner JM, Stevens J, Rosser SJ, Basran A, Lerner RA, Bruce NC, Wilson IA. Nat Struct Biol. 2002;9:17–21. doi: 10.1038/nsb742. [DOI] [PubMed] [Google Scholar]
- 11.Mäkelä O. J Immunol. 1965;95:378–386. [PubMed] [Google Scholar]
- 12.Self CH, Thompson S. Lancet. 2006;367:1038–1039. doi: 10.1016/S0140-6736(06)68396-7. [DOI] [PubMed] [Google Scholar]
- 13.Cho AE, Wendel JA, Vaidehi N, Kekenes-Huskey PM, Floriano WB, Maiti PK, Goddard WA., III J Comput Chem. 2005;26:48–71. doi: 10.1002/jcc.20118. [DOI] [PubMed] [Google Scholar]
- 14.Ellman GL, Courtney KD, Andres V, Jr, Featherstone RM. Biochem Pharmacol. 1961;7:88–95. doi: 10.1016/0006-2952(61)90145-9. [DOI] [PubMed] [Google Scholar]
- 15.Carrera MRA, Ashley JA, Zhou B, Wirsching P, Koob GF, Janda KD. Proc Natl Acad Sci USA. 2000;97:6202–6206. doi: 10.1073/pnas.97.11.6202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Masson P, Adkins S, Gouet P, Lockridge O. J Biol Chem. 1993;268:14329–14341. [PubMed] [Google Scholar]
- 17.Darvesh S, Hopkins DA, Geula C. Nat Rev Neurosci. 2003;4:131–138. doi: 10.1038/nrn1035. [DOI] [PubMed] [Google Scholar]
- 18.Doctor BP, Saxena A. Chem Biol Interact. 2005;157–158:167–171. doi: 10.1016/j.cbi.2005.10.024. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.