

Abstract
Background and Aims
Successful cryopreservation of bryophytes is linked to intrinsic desiccation tolerance and survival can be enhanced by pre-treatment with abscisic acid (ABA) and sucrose. The pioneer moss Ditrichum plumbicola is naturally subjected to desiccation in the field but showed unexpectedly low survival of cryopreservation, as well as a poor response to pre-treatment. The effects of the cryopreservation protocol on protonemata of D. plumbicola were investigated in order to explore possible relationships between the production in vitro of cryopreservation-tolerant asexual propagules and the reproductive biology of D. plumbicola in nature.
Methods
Protonemata were prepared for cryopreservation using a four-step protocol involving encapsulation in sodium alginate, pre-treatment for 2 weeks with ABA and sucrose, desiccation for 6 h and rapid freezing in liquid nitrogen. After each stage, protonemata were prepared for light and electron microscopy and growth on standard medium was monitored. Further samples were prepared for light and electron microscopy at intervals over a 24-h period following removal from liquid nitrogen and re-hydration.
Key Results
Pre-treatment with ABA and sucrose caused dramatic changes to the protonemata. Growth was arrested and propagules induced with pronounced morphological and cytological changes. Most cells died, but those that survived were characterized by thick, deeply pigmented walls, numerous small vacuoles and lipid droplets in their cytoplasm. Desiccation and cryopreservation elicited no dramatic cytological changes. Cells returned to their pre-dehydration and cryopreservation state within 2 h of re-hydration and/or removal from liquid nitrogen. Regeneration was normal once the ABA/sucrose stimulus was removed.
Conclusions
The ABA/sucrose pre-treatment induced the formation of highly desiccation- and cryopreservation-tolerant propagules from the protonemata of D. plumbicola. This parallels behaviour in the wild, where highly desiccation-tolerant rhizoids function as perennating organs allowing the moss to endure extreme environmental conditions. An involvement of endogenous ABA in the desiccation tolerance of D. plumbicola is suggested.
Key words: ABA, asexual propagules, cell-biology, cryopreservation, desiccation tolerance, Ditrichum plumbicola, extremophiles, protonemata, sucrose
INTRODUCTION
The production of asexual propagules in bryophytes, in particular mosses, is well documented (Whitehouse, 1973, 1980; Side and Whitehouse, 1987; Duckett and Ligrone, 1992; Goode et al., 1993a, b, 1994). Propagules include gemmae, tubers or brood cells and are continually being described from both tissue cultures and natural populations. Their suggested function is as perennating organs, produced as survival mechanisms in unfavourable circumstances (Duckett and Pressel, 2003; Preston, 2004; Pressel et al., 2005). Previous studies, particularly on the formation of brood cells, indicate that abscissic acid (ABA) and/or desiccation are stimuli for formation (Goode et al., 1993a).
Specialized propagules with the ability to survive prolonged periods without water add an extra dimension to the knowledge of desiccation tolerance in mosses (for an overview, see Oliver et al., 2000a, b; Proctor, 2001). In common with other plant tissues, some mosses when suitably dried, are able to tolerate extreme temperature stress (Bequerel, 1951; Bewley, 1973; Glime and Carr, 1974; Norr, 1974; Hearnshaw and Proctor, 1982). This is exploited in cryopreservation protocols for the storage of plant tissues, where material is often first dried before being immersed in liquid nitrogen (LN) at − 196 °C. Indeed there is evidence that post-cryopreservation survival in mosses depends on intrinsic desiccation tolerance (Burch, 2003) and that this can be enhanced by pre-treatment with ABA and sucrose (Pence, 1998; Burch and Wilkinson, 2002).
The use of cryopreservation techniques is advocated for the long-term storage of plant material when conventional methods, such as seed banking, are inappropriate (Benson, 1999). Such tools are increasingly being used for the ex situ conservation of threatened species (Guerrant et al., 2004). Between 2000 and 2006, the Royal Botanic Gardens, Kew supported a project for the ex situ conservation of threatened bryophytes in the UK. One aim of the project was to provide a cryopreserved collection of gametophytic material, to store taxa from which spores were not necessarily available (Ramsay and Burch, 2001), and methods were developed accordingly (Burch and Wilkinson, 2002; Burch, 2003).
One of the target species of the project was Ditrichum plumbicola, a moss classified as near threatened on the British red list (Church et al., 2001), the distribution of which in the British Isles is restricted to lead mine spoil (Porley and Hodgetts, 2005). Gametangia and sporophytes are unknown for this species, but rhizoidal tubers have been described (Arts, 1994). Ditrichum plumbicola grows exclusively on bare soil subjected to desiccation in the summer and surface cryoturbation after winter frosts. It was therefore reasonable to assume that the moss was both desiccation and freezing tolerant. Preliminary cryopreservation trials on the protonemata, however, showed that this moss responded to the protocol differently from any other desiccation-tolerant species tested (Burch, 2003; J. K. Rowntree, unpubl. res.). In particular, exposure to ABA and sucrose elicited profound changes in protonemal morphology, producing cells which initially failed to regenerate following cryopreservation.
Thus, the aims of this study were firstly to determine if ABA or sucrose, or a combination of the two, induced the morphological changes observed in D. plubicola; and whether any other stages of the protocol played a part in this process. In a wider context, the objectives were (a) to characterize the cytology of these cells and compare them with brood cells; (b) to report on the cytological changes elicited in these cells by desiccation and cryopreservation; (c) to explore possible relationships between the production in vitro of asexual propagules that survive cryopreservation and the reproductive biology of D. plumbicola in nature.
MATERIALS AND METHODS
Plant material
Gametophytic material of Ditrichum plumbicola Crundw. was collected from a single lead mine spoil site in County Durham for initiation into tissue culture. Wild protonemata were collected for comparison from the same site and also from two further lead mine spoil sites in North and mid-Wales. The latter were kept in herbarium packets and observed over a 1-year period from the time of collection. As the species is designated under the UK Biodiversity Action Plan initiative, collection was only undertaken with permission of the lead partner (see http://www.ukbap.org.uk/ for more information) and specific site details are undisclosed.
Culture conditions
Protonematal cultures were prepared from surface-sterilized gametophore fragments. Stock cultures were maintained on sucrose-free quarter strength Murashige and Skoog medium (1/4 MS) with micro- and macro-elements including vitamins (Duchefa Biochemie B.V., Haarlem, The Netherlands) at pH 5·8, solidified with 4·0 g L−1 Gelrite™ (standard medium) in Petri dishes sealed with Micropore™ tape to allow for gas exchange. They were maintained in a growth room with a 16 h light/8 h dark regime, under a 1 : 1 mixture of NARVA 58W/077 and GE58W/29 fluorescent tubes (15–50 µmol m−2 s−1 PAR) at 20·5 °C (±3·5).
Cryopreservation experiment
Protonematal material was prepared for cryopreservation in LN according to the methods developed by Burch and Wilkinson (2002) and Burch (2003). The protocol was a four-stage process: (1) encapsulation in 3 % sodium alginate, solidified with 100 mm CaCl solution, (2) pre-treatment for 2 weeks with 10 µm ABA and 50 g L−1 sucrose, (3) dried for 6 h followed by (4) rapid immersion and storage in LN.
Ten protonematal plugs (approx. 1 mm diameter) were removed from the stock cultures, transferred onto standard medium and their growth monitored weekly for a total of 5 weeks by measuring plug diameter along two permanent transects marked on the base of the dishes with vernier callipers according to Rowntree et al. (2005) (control treatment). Forty protonematal plugs were each encapsulated in an alginate strip (for methods, see Burch and Wilkinson, 2002). The strips were then placed singly in Petri dishes containing standard medium and maintained in the growth room for 1 week to allow for recovery from possible damage incurred during their preparation. Protonematal growth from ten strips was monitored weekly for a total of 5 weeks (encapsulation treatment). The remaining 30 strips were transferred to Petri dishes containing standard medium supplemented with 50 g L−1 sucrose and 10 µm ABA. Sucrose was added to the medium prior to autoclaving and ABA was added post-autoclaving from a stock solution (50 mm in 96 % ethanol) to cooled, but still liquid, medium. After 2 weeks, ten strips were transferred back onto standard medium and their growth monitored for 5 weeks (ABA/sucrose treatment). The remaining 20 strips were placed in clean, open Petri dishes in a laminar flow hood, under a sterile air flow (0·45 m s−1) for 6 h to dry. Final water content was approx. 0·1 g H2O g−1 d. wt. Ten of these strips were transferred back onto standard medium and growth was monitored for 5 weeks (dehydration treatment), while the remaining ten strips were placed into 2·0-mL cryovials (Nalgene, Rochester, NY, USA) and sealed. The vials were frozen by rapid immersion in LN and stored at −196 °C for 1 week. Vials were then removed from the LN, immersed in a water bath at 40 °C for 2 min and dried. Strips were removed from the vials in a laminar flow hood, placed onto standard medium and growth was monitored for 5 weeks (cryopreservation treatment).
After 5 weeks, five replicates were randomly selected from each of the treatments. Protonemal material was removed from inside the alginate strips, transferred onto plates of standard medium and growth was monitored. Additional strips were prepared from each treatment for examination via light and electron microscopy. Cryopreserved material remained frozen in LN until processing and was transported in a dry shipper (IC-4VS, Wessington Cryogenics Ltd, Houghton-le-Spring, Tyne and Wear, UK) between facilities.
ABA/sucrose experiment
Standard medium was prepared without additional sucrose as above or with 50 g L−1 sucrose added. ABA (10 µm) was then added to half of the medium, giving a total of four treatments. Plugs of D. plumbicola protonemata (approx. 2 mm diameter) were removed from the stock cultures and transferred onto 5-cm-diameter, shallow Petri dishes (15 mL) containing one of the four media. Fifteen replicates were used per treatment, and there were a total of 60 plates for the whole experiment. Mean diameter growth was measured for 5 weeks as above and any structural and colour changes in the protonemata were monitored. After 5 weeks, five replicates were again randomly selected from each treatment and small protonemal plugs were transferred onto plates of standard medium. Growth was monitored for a further 5 weeks as above. Protonemata representative of each treatment after 2 weeks growth were prepared for light microscopy.
Light microscopy
Control, encapsulated and ABA/sucrose-treated specimens were mounted in water and photographed with a digital camera under a Leica DM RXAZ microscope using interference contrast optics. Dehydrated and cryopreserved specimens were mounted in immersion oil to prevent re-hydration, and photographed as above.
Electron microscopy
Protonemata were fixed and embedded for electron microscopy following the methods of Duckett and Ligrone (1992). Cryopreserved protonemata were either placed immediately in fixative upon removal from LN or allowed to regenerate for 0·5, 1, 2, 4, 12 and 24 h on standard medium. Protonemata were fixed in a mixture of 3 % (v/v) glutaraldehyde, 1 % (v/v) formaldehyde (freshly prepared from paraformaldehyde) and 0·5 % tannic acid (w/v) in 0·05 m Na-cacodylate buffer at pH 6·9 for 4 h at room temperature. Material was rinsed in 0·1 m Na-cacodylate buffer and post-fixed with 1 % (w/v) osmium tetroxide in 0·1 m Na-cacodylate buffer overnight at 4 °C. It was then dehydrated in ethanol and embedded in Spurr's resin via propylene oxide. Thin sections, cut with a diamond knife and sequentially stained with 5 % (v/v) methanolic uranyl acetate for 15 min and lead citrate for 10 min, were examined under a Jeol 1200 EX2 electron microscope. Specimens for scanning electron microscopy were dehydrated in ethanol, critical point dried and observed in a Hitachi S570 scanning electron microscope operating at 20 kV.
Sections (0·5 µm thick) stained with 1 % toluidine blue were photographed with a Leica DM RXA2 microscope equipped with differential interference contrast optics.
Data analysis
Data from week 5 of the cryopreservation experiment were analysed with a fixed effects (Model I) one-way ANOVA and Bonferroni post hoc tests, with the five treatments as independent categorical variables. Data from week 5 and then week 2 of the ABA/sucrose experiment were analysed with separate fixed effects (Model I) two-way ANOVAs, with sucrose and ABA as independent categorical variables. A Bonferroni correction factor for multiple tests was used where αadj = (α/k) and k was the number of tests (Sokal and Rohlf, 1995, 239–240). One replicate from the control treatment did not survive the original transfer; no growth was observed for the duration of the experiment and protonemata were colourless. This was removed from the statistical analysis giving a total n = 59. Normality was tested using a method for small sample sizes according to Sokal and Rolf (1995). Heteroskedasticity was not directly tested for, as Zar (1999, 185) and Scheffe (1959) indicate the robustness of ANOVAs from deviations in homoskedasticity if sample sizes are equal or nearly equal. Statistical analyses were performed using Systat 10 software (Systat Software, 2004)
RESULTS
Controls
Growth of D. plumbicola in culture was observed mainly as a steady expansion of the protonematal plug in two dimensions across the surface of the medium, although three-dimensional growth was also observed (Figs 1 and 2A). The protonemal system produced was typical (Fig. 2A), with well-differentiated chloronema (Fig. 3A) and caulonema filaments with longitudinally aligned, starch-filled chloroplasts (Fig. 3B). Rhizoidal filaments with deeply pigmented walls and minute plastids were also produced, but only after a prolonged period (>5 weeks) of culture (Fig. 3C). An aerial system of tightly packed chloronema filaments arising vertically from the prostrate system gave a characteristic ‘bushy’ appearance to the cultures (Fig. 2A).
Fig. 1.

Growth data (mean diameter in mm) at week 5 for control (1), encapsulation (2), ABA/sucrose (3), desiccation (4) and cryopreservation (5) treatments of Ditrichum plumbicola. Error bars = s.e., n = 50. Encapsulation treatment shows significantly reduced growth compared with the control (P < 0·001). ABA/sucrose, desiccation and cryopreservation treatments show significantly reduced growth compared with both the control and encapsulation treatments (P < 0·001), but not each other.
Fig. 2.

Ditrichum plumbicola protonema after 5 weeks of treatment: (A) control, (B) encapsulation, (C) ABA/sucrose, (D) desiccation and (E) cryopreservation treatments.
Fig. 3.

Light micrographs of cultured protonemata of Ditrichum plumbicola – controls and encapsulated material. (A–D) Controls: (A) typical chloronema cell with ovoid peripheral chloroplasts; (B) typical caulonema cell with elongated, longitudinally aligned plastids; (C) typical rhizoid with thick brown-pigmented walls, minute elongated plastids and spindle-shaped nucleus (n); (D) dedifferentiation of aerial chloronemal cells into thin-walled brood cells after 2–3 weeks in culture. (E–H) Encapsulated material: (E, F) typical chloronema (E) and caulonema (F) cells indistinguishable from the controls; (G) thin-walled brood cells on chloronema filaments after 2–3 weeks in culture; (H) 1-μm section of chloronema cell – note the large central vacuole and the peripherally located plastids (p) and nucleus. Scale bars: D, E, G = 50 µm; A–C, F = 20 µm; H = 10 µm.
The control protonemata exhibited a marked tendency towards broodiness. In young cultures (2–3 weeks old), numerous chloronema cells, especially those of the aerial system, readily dedifferentiated into spherical, thin-walled brood cells (Fig. 3D).
Encapsulation
Post-encapsulation, initial growth was within the alginate strips but, after a few weeks, protonemata grew through the alginate matrix onto the standard medium (Fig. 2B). There was a significant reduction in growth compared with the controls (week 5: P < 0·001; Fig. 1). Encapsulation did not affect the morphology (Figs 2B and 3E, F) and cytology (Fig. 3H) of protonemata and the cells showed the same tendency towards broodiness as the controls (Fig. 3G).
Combined ABA/sucrose treatment
Pre-treatment with a combination of sucrose and ABA elicited pronounced changes in the protonemata. Growth was significantly reduced compared with both the controls and encapsulated treatments (week 5: P < 0·001; Fig. 1) and appeared to have been arrested within the alginate strips (Fig. 2C). The majority of the protonemal system died, including almost all the main caulonemal axes (Fig. 4A, B). The parts that survived consisted of uniseriate rows of three to eight cells with thick, deeply pigmented walls and an apical cell ± subapical cell with less dense cytoplasm and thinner, non-pigmented walls (Fig. 4C). Incomplete cross walls were often present towards the tip of the apical cells (Fig. 4D). Some chloronemal side branches and adjacent cells along the filaments also survived. These were characterized by a dense cytoplasm containing numerous lipid droplets and by thick, but not always pigmented, cell walls (Fig. 4C, E).
Fig. 4.

Light micrographs of sucrose/ABA-treated protonemata: (A) the majority of the cells are dead; those that survive are characterized by heavily pigmented walls; (B) detail of dead cells showing complete disruption of cell contents; (C) heavily pigmented filament with dense cytoplasm and with non-pigmented apical cell (arrowed); (D) detail of non-pigmented apical cell – the arrow indicates incomplete cross wall; (E) lipid-laden chloronemal cells with non-pigmented thick walls; (F–H) encapsulated, sucrose/ABA-treated cells after 2 weeks from transfer to standard growth medium. The protonemal system remains virtually unchanged except for increased brood cell development (F) and detachment of the same (arrowed) from the parent filament (H). Scale bars: A, F = 200 µm; B, C = 100 µm; E, H = 50 µm; D, G = 20 µm.
The cells remained largely unchanged when the alginate strips containing them were transferred intact to standard growth medium (Fig. 4F–H), except for a pronounced dedifferentiation of chloronema side branches into brood cells, which often became detached from the parent filament (Fig. 4H).
Transmission electron microscopy confirmed observations from light microscopy. The majority of the cells that remained viable had a highly heterogeneous, thick (>3 µm) cell wall, comprising a densely fibrillar central layer with multiple strata of varying density on each side (Fig. 5A). Prominent projections of the loosely fibrillar, innermost layer containing a denser central core (Fig. 5B) were a frequent feature of most cells along the filaments except for the apical cells. Their walls were relatively thin (>1 µm) and homogeneous and never developed inner projections (Fig. 5C). The presence of incomplete cross walls in the apical cells was confirmed (Fig. 5C). The cytoplasm was packed with numerous small vacuoles (ranging from 200 nm to a few micrometres in diameter) often containing small deposits of electron-opaque material (Fig. 5D, E). The apical cells usually contained fewer vacuoles (Fig. 5C). The plastids were ovoid to discoidal with numerous, large starch grains (Fig. 5A, D). The mitochondria were rounded to elongate, with a dense stroma and saccate cristae (Fig. 5E). Nuclei were rounded with a prominent nucleolus (Fig. 5D).
Fig. 5.

Transmission electron micrographs of sucrose/ABA-treated protonemal cells before and after dehydration. (A–E) Fully hydrated cells: (A) cell with starch-filled plastid and with thick, multi-layered cell wall – note the numerous projections of the cell wall innermost layer; (B) detail of cell wall projection consisting of loosely fibrillar material; (C) incomplete cross wall (arrowed) in apical cell – note the thin, homogeneous cell wall (cw); (D) cell packed with small vacuoles – also note the centrally located round nucleus (n) and the starch-filled plastids; (E) cytoplasm containing mitochondria (m) with dense matrix and saccate cristae and vacuoles (v) with electron-opaque content. (F–H) Dehydrated cells: (F) ovoid plastids with small starch grains (arrowed) and with an intact, undulating thylakoid system; (G) nucleus (n) with conspicuous blocks of condensed chromatin surrounded by small vacuoles; (H) heterogeneous cell wall (cw) with irregular internal projections. Scale bars: D = 5 µm; A–C, E–G = 2 µm; H = 1 µm.
Separate ABA/sucrose treatments
ABA and sucrose applied singly caused changes similar to, but less pronounced than, the combined treatment, with the majority of the cells remaining alive. Both treatments caused a significant reduction in growth compared with the controls (week 5: P < 0·001; Fig. 6) and a significant interaction between ABA and sucrose was detected (week 5: P < 0·001; Fig. 6). Analysis of data at week 2 did not change the effects or significance values determined but R-values were slightly reduced.
Fig. 6.

Mean diameter (mm) of Ditrichum plumbicola protonema grown for 5 weeks on control (open circles, continuous line), 5 % sucrose (open triangles, continuous line), 10 µM ABA (closed circles, dashed line) and 5 % sucrose/10 µM ABA (closed triangle, dashed line) media. Error bars are s.e., n = 59. Significance values from two-way ANOVA at week 5 are shown.
After 2 weeks on medium supplemented solely with 10 µm ABA, most of the filaments had become thick-walled and deeply brown-pigmented (Fig. 7A). The majority of chloronemal cells had dedifferentiated into spherical brood cells (Fig. 7B, C), while caulonemal cells were shorter, with denser contents than the controls and bore only very short side branches (Fig. 7D).
Fig. 7.
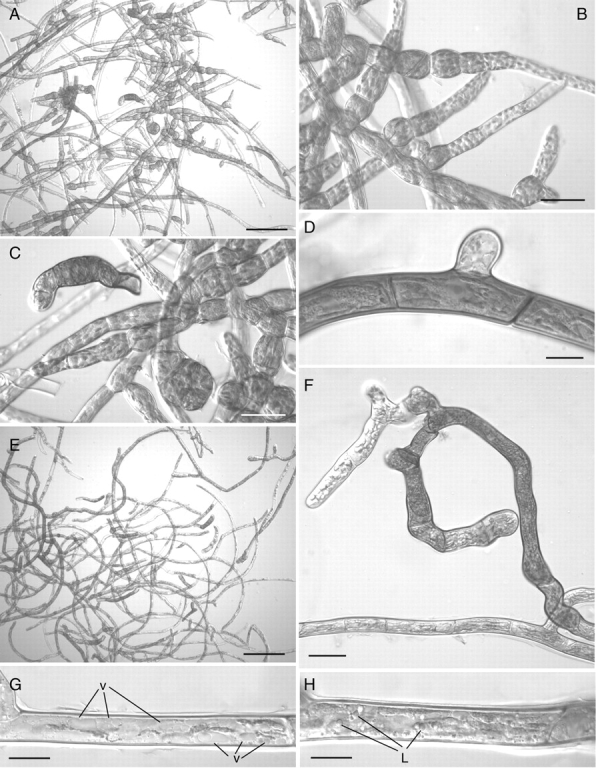
Light micrographs of ABA- and sucrose-treated protonemal cells. (A–D) Treatment for 2 weeks with 10 µM ABA: (A) most filaments are thick-walled and deeply pigmented; (B) dedifferentiation of chloronema filaments into spherical brood cells; (C) caulonema filament with thick pigmented walls – note the short cells with dense cytoplasm and the very short side branch. (E–H) Treatment for 2 weeks with 5 % sucrose: (E) general aspect showing thick-walled pigmented filaments as in (A) but no brood cells; (F) detail of thick-walled pigmented filament – note the thin-walled non-pigmented apical cells in the side branch; (G, H) caulonema cells with numerous small vacuoles (v) (G) and abundant lipid droplets (L) (H) in their cytoplasm. Scale bars: A, E = 200 µm; B, C, F = 50 µm; D, G, H = 20 µm.
The addition of 50 g L−1 sucrose only to the medium elicited similar changes to the appearance of protonemal cultures especially with regards to an increase in cell wall thickness and pigmentation (Fig. 7E). In contrast, the sucrose treatment affected mainly the prostrate system, with filaments of the aerial system appearing largely unaffected (data not shown). The prostrate chloronemal filaments did not dedifferentiate into brood cells. With the exception of apical and a few sub-apical cells, which remained thin-walled (Fig. 7F), the majority of these filaments became thick-walled and deeply brown-pigmented (Fig. 7E, F). Numerous lipid droplets were present in the cytoplasm of these cells (Fig. 7G) together with abundant small vacuoles (Fig. 7H).
Dehydration
Dehydration caused few additional cytological changes and although growth was significantly reduced compared with the controls and encapsulated treatment (week 5: P <0·001), there were no statistical differences with the ABA/sucrose treatment (Fig. 1). Numerous small vacuoles were present in the cytoplasm of dehydrated cells (Fig. 5G) and the cell wall remained highly heterogeneous and with conspicuous extensions in their innermost layers (Fig. 5H). The starch grains in the plastids, however, were less numerous and much smaller than in the hydrated cells (Fig. 5F), and within the nucleus, conspicuous blocks of condensed chromatin surrounded the highly compacted nucleolus (Fig. 5G). Mitochondria were generally rounded and with straight tubular cristae (Fig. 5F, H).
Cryopreservation and subsequent re-hydration
Light microscopy showed the frozen cells to be flattened with rounded or discoidal plastids distributed throughout the cell lumen (Fig. 8A). One hour after removal from LN, the cytoplasm had expanded but remained packed with numerous small vacuoles and large lipid droplets (Fig. 8B, C). The cytoplasm in the apical cells was less dense (Fig. 8D) and incomplete cross walls, resembling those in the sucrose/ABA-treated hydrated cells, were often present (Fig 8E). The overall appearance of the cells remained unchanged even after several weeks of recovery (Fig. 8F). Growth at 5 weeks was significantly reduced compared with the control and encapsulated treatments (P < 0·001), but no statistical differences were detected between the three post-ABA/sucrose treatments (Fig. 1).
Fig. 8.

Light micrographs of frozen and thawed protonemal cells. (A) Frozen cells mounted in immersion oil, the cells are flattened and the discoidal plastids are distributed throughout the cell lumen. (B–E) Frozen cells 1 h after thawing: (B, C) elongate plastids (p) in cytoplasm packed with lipid droplets (L) and small vacuoles – note the ovoid nucleus (n) with prominent nucleolus and no evidence of condensed chromatin in (C); (D) apical cells with round nuclei and less dense cytoplasm; (E) apical cell with incomplete cross wall (arrowed). (F) Cells kept in the original alginate strip 2 weeks after thawing – the cytology remains unchanged; (G, H) Production of normal filaments 2 d (G) and 4 d (H) after removing the thawed protonema from the alginate strips. Scale bars: G, H = 200 µm; A–F = 20 µm.
The cytology of the cryopreserved cells resembled that of the dehydrated cells (Fig. 9A–E): cells were packed with small vacuoles and numerous lipid droplets; nuclei were rounded with conspicuous blocks of condensed chromatin (Fig. 9A) and some starch grains remained in the plastids (Fig. 9B), especially those of the apical cells (Fig. 9D, E). In the latter the cytoplasm was less dense, the vacuoles less numerous than in the other cells (Fig. 9D and E), and short segments of tubular endoplasmic reticulum (ER) were often present in the proximity of the cell wall (Fig. 9E). As in the dried cells, mitochondria were ovoid with tubular, parallel cristae (Fig. 9C).
Fig. 9.

Transmission electron micrographs of frozen and thawed protonemal cells. (A–E) Frozen cells: (A) elongated plastids and nucleus with condensed chromatin – note the numerous lipid droplets and the small vacuoles in the cell lumen; (B) plastids with intact thylakoids and small starch grains; (C) rounded mitochondria with thin, parallel-sided cristae; (D) ovoid plastids with starch grains in an apical cell. (E) Short segments of ER (arrowed) remaining in the lumen of an apical cell (cw, cell wall). (F–H) Frozen cell 1–4 h after thawing: (F) the cytoplasm is packed with small vacuoles and numerous lipid droplets; (G) profiles of tubular ER (arrowed) are scattered in the cytoplasm amongst the mitochondria and the plastids, which now contain numerous starch grains, especially those of the apical cells (H). cw, Cell wall. Scale bars: A, F = 5 µm; B, D, E, G, H = 2 µm; C = 1 µm.
After 1–4 h following removal from LN, the cytoplasm had increased in volume but remained packed with numerous small vacuoles and large lipid droplets (Fig. 9F). Small pockets of cytoplasm between the nuclei, plastids and mitochondria contained dictyosomes and profiles of tubular ER (Fig. 9G). The plastids, especially those of the apical cells (Fig. 9H) contained numerous starch grains and were indistinguishable from the controls. Thereafter, the cytology of the protonemata remained unchanged and 24 h after removal from LN, the appearance of the cells was the same as that after 4 h (data not shown).
When re-hydrated cells were removed from the alginate strips and then placed onto standard growth medium they promptly regenerated, producing a normal protonemal system within 2–3 d (Fig. 8G, H). The same was also true for sucrose/ABA-treated and dehydrated cells (data not shown).
Ditrichum plumbicola in the wild
Gemmiferous protonemata around the periphery of gametophore colonies were not detected in the three wild populations of D. plumbicola examined in the present study. Older colonies of D. plumbicola, particularly where the soil showed no signs of recent disturbance, were often overgrown by the perennial protonemata of Pogonatum aloides (Fig. 10A). Analysis of the rhizoids revealed a similar absence of asexual propagules. Both freshly collected rhizoids (Fig. 10C) and those kept as herbarium specimens (Fig. 10B) had thick brown-pigmented walls and were packed with food reserves mainly in the form of large lipid droplets interspersed with occasional starch deposits (Fig. 10C). Some had minute longitudinally aligned plastids like those seen in culture (Fig. 10D). When both freshly collected and stored rhizoids were placed on culture medium they produced typical protonemata indistinguishable from the cultured controls within 2–3 d (Fig. 10E).
Fig. 10.
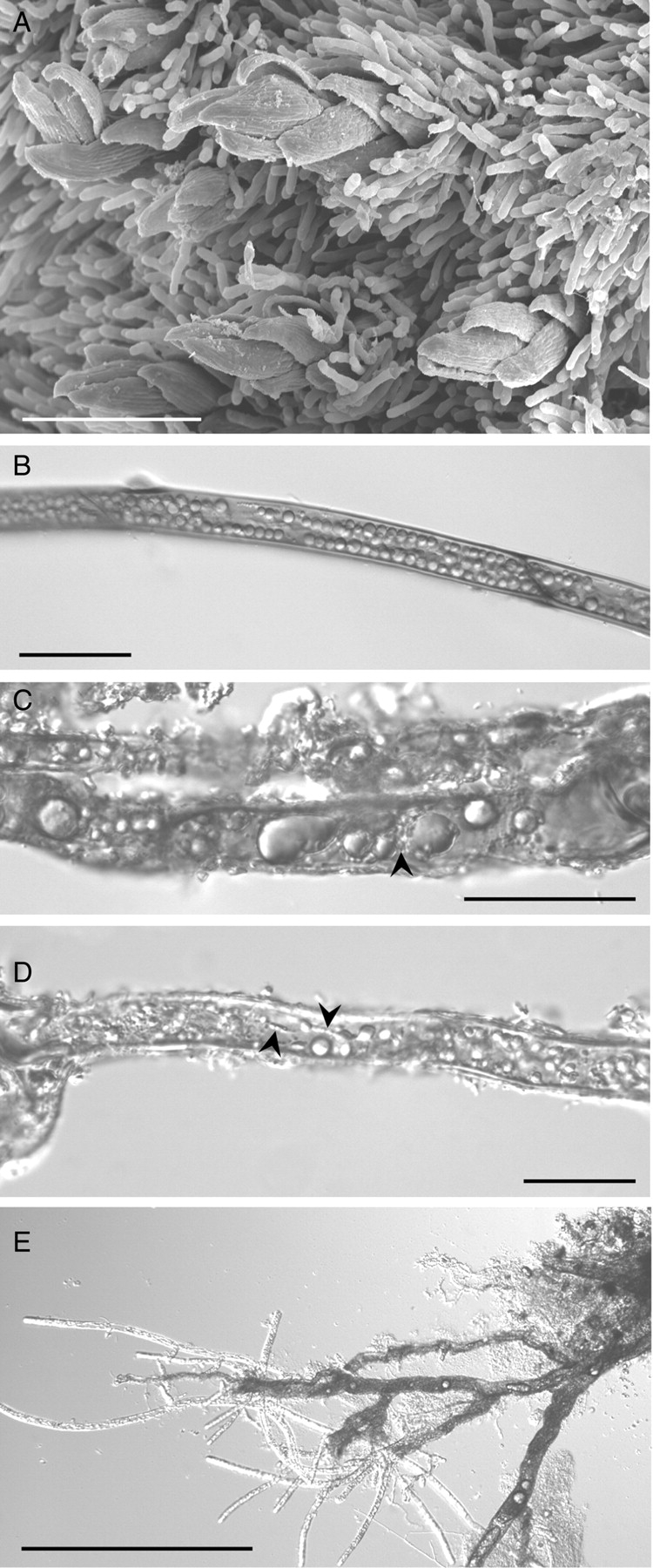
Ditrichum plumbicola from the wild: (A) scanning electron micrograph of mature colony showing D. plumbicola gametophores overgrown by the perennial protonemata of Pogonatum aloides; (B–D) light micrographs of herbarium (B) and freshly collected (C and D) material showing thick-walled rhizoids packed with lipid droplets of various sizes and only occasional starch grains (C) – in (D) note the minute plastids (arrowed) aligned along cytoplasmic strands like those seen in culture (see Fig. 4C); (E) normal protonemal system regenerating from > 1-year-old herbarium material. Scale bars: E = 100 µm; B–D = 50 µm; A = 20 µm.
DISCUSSION
The results clearly showed that exposure to a combination of sucrose and ABA inhibited growth and caused marked morphological and cytological changes to the protonemal cells of Ditrichum plumbicola. The majority of cells died but those that remained viable were characterized by thick walls, abundant lipid droplets, and by numerous small vacuoles in their cytoplasm. These cells had the ability to survive desiccation and cryopreservation in LN largely unperturbed. Once the ABA/sucrose stimulus was removed, i.e. following re-hydration and removal from the original alginate-matrix, the cells quickly regenerated into a normal protonemal system. Encapsulation in an alginate-matrix alone had little or no effect, apart from a reduction in protonemal growth rate. ABA and sucrose, applied singly, elicited changes similar to, but less pronounced than, those of the combined treatment, with the majority of the cells remaining alive. The effects of ABA were more pronounced and extended to all the filaments, while sucrose affected exclusively the prostrate system. Cells with morphology typical of brood cells were produced only in response to ABA but not sucrose.
Cytological considerations
Duckett and Ligrone (1992) and Goode et al. (1993a) described the formation of typical brood cells, as spherical or ovate cells ‘derived by re-differentiation from initially cylindrical chloronemal cells by swelling and breakdown of the middle lamellae of the cross walls’. The cells formed from D. plumbicola after pre-treatment with sucrose/ABA were similar to, even if not entirely congruent with, this description and will therefore be referred to as ‘brood bodies’.
The cytology of the brood bodies was remarkably similar to that described by Goode et al. (1994) for Aloina aloides and by Goode (1992) for a number of other species from ageing cultures or after exposure to ABA alone. Ultrastructural observations also revealed the D. plumbicola brood bodies to have characteristics typical of drought-tolerant propagules, e.g. thick cell walls and dense cytoplasm containing numerous small vacuoles and lipid droplets (Goode et al., 1994; Schnepf and Reinhard, 1997).
The presence of abundant food reserves is a common feature of bryophyte spores (Brown and Lemmon, 1987, 1988), and leaf cells of desiccation-tolerant mosses often contain small vacuoles (Tucker et al., 1975; Proctor and Tuba, 2002). Vacuolar fragmentation in response to drying has been extensively reported in desiccation-tolerant vascular (resurrection) plants (Gaff et al., 1976; Thomson and Platt, 1997; Vander Willigen et al., 2003) and, recently, in both the leaf cells (Proctor et al., 2007) and food-conducting cells (Pressel et al., 2006) of the desiccation-tolerant moss Polytrichum formosum. Nagao et al. (2005) showed that exposure to ABA induced vacuolar fragmentation in protonemal cells of Physcomitrella patens.
The dehydration treatment caused relatively few cytological changes to the brood bodies of D. plumbicola, and the integrity of the membranes and organelles was retained throughout the drying process. This supports the assertion that the brood bodies themselves are highly desiccation tolerant. Chromatin condensation was most likely a direct consequence of the withdrawal of water from the cells (Pressel et al., 2006) and is a typical feature of seeds during the dehydration phase of maturation (Klein and Pollock, 1968). Changes in the internal structure of mitochondria have been described previously in dried vegetative tissues in numerous plant systems, and are indicative of metabolic inactivity (Dalla Vecchia et al., 1998; Navari-Izzo et al., 2000; Proctor et al., 2007).
The disappearance of starch from the plastids in the dry state has been previously reported, e.g. in the mosses Polytrichum formosum (Pressel et al., 2006; Proctor et al., 2007) and Sphagnum (Gerdol et al., 1996), and the resurrection angiosperms Sporobolus stapfianus (Quartacci et al., 1997; Dalla Vecchia et al., 1998), Craterostigma plantagineum (Bianchi et al., 1991; Norwood et al., 2000) and Craterostigma wilmsii (Vicre et al., 2004). Perhaps the most likely explanation for the persistence of a few small starch grains in the plastids of D. plumbicola after drying, is that these cells already contained high levels of soluble carbohydrates following the sucrose pre-treatment. It is also possible that the rate of drying in the laminar flow hood did not allow sufficient time for all the starch to be hydrolysed. Further, death of the caulonemal cells may have prevented the translocation of soluble carbohydrates.
Once dried, the brood bodies of D. plumbicola survived cryopreservation without incurring further cytological changes. Their prompt return to the pre-drying and pre-cryopreservation state upon removal from LN and re-hydration was shown by an increase in the ER network, the reappearance of numerous starch grains in the plastids, and changes in the internal structure of the plastids and mitochondria. This confirms previous reports that dried plant tissues can survive extreme cold (Bequerel, 1951; Pence, 2000) and further emphasizes the link between desiccation tolerance and freezing survival (Burch, 2003).
ABA, sugars and desiccation biology
Sugars and ABA have been linked to improved desiccation and freezing tolerance in a wide range of plant tissues (Pence, 1998; Sreedhar et al., 2002; e.g. Werner et al., 1991; Zhu et al., 2006) with potentially interacting protective effects (Suzuki et al., 2006). They are often incorporated singly or in combination into cryopreservation protocols (e.g. Burch and Wilkinson, 2002; Fang et al., 2004).
Sugars and specific proteins play important roles during the loss of cellular water, by protecting membranes and macromolecules via direct interaction and by immobilizing the cytoplasm into a highly viscous, glassy state (for a review, see also Buitink and Leprince, 2004; Alpert, 2006). Sugars are also osmotic agents, reducing cell water content (Benson, 1999; Zhu et al., 2006) and there is evidence that they play a further protective role by scavenging damaging free radicals (Linders et al., 1997).
Many of the responses elicited by ABA in bryophytes are the same, or very similar to, those described in higher plants (Beckett, 2001). In bryophytes specifically, besides increasing desiccation and freezing tolerance (Hartung et al., 1987; Nagao et al., 2005; Pence et al., 2005), exogenous ABA has been shown to induce the production of specific proteins (Bopp and Werner, 1993; Werner et al., 1991), inhibit cytokinin-stimulated bud induction (Valadon and Mummery, 1971) and reduce protonemal growth rates (Burch and Wilkinson, 2002). Following ABA treatment, the moss Funaria hygrometrica (Werner et al., 1991) and the liverworts Riccia fluitans and Pallavicinia lyellii (Pence et al., 2005) were able to withstand faster rates of desiccation, while tolerance of the moss Atrichum androgynum to ion leakage during rehydration was enhanced (Beckett, 2001; Guschina et al., 2002). ABA has also been shown to be an important stimulus in the transition of the liverwort Riccia fluitans from water to land form (Hellwege et al., 1992).
The failure of protonemal cells of D. plumbicola to regenerate within the alginate strips following removal from LN and re-hydration is attributed to an accumulation and retention of ABA in the alginate strip following pre-treatment. That these cells promptly regenerated once removed from the alginate strips and placed onto fresh culturing medium, supports this suggestion. These results also agree with studies on brood cell formation (Goode, 1992; Goode et al., 1993a), which showed that once produced, brood cells did not germinate in situ but only after they were transferred to new growth medium. The same has also been demonstrated for protonemal gemmae (Duckett and Ligrone, 1991).
Extremeophile propagules
The desiccation- and cryopreservation-tolerant brood bodies of D. plumbicola resemble such ‘extremophile’ propagules as those produced by microbes in the high arctic and the encysted embryos of the crustacean Artemia franciscana (Clegg, 2005; Mueller et al., 2005). Mueller et al. (2005) suggested that pigmentation of the microbial propagules provided protection from a range of environmental stresses, whilst Clegg (2005) indicated that the tough shell of Artemia cysts ensured survival of the embryos within. The Ditrichum brood bodies with their thick, pigmented walls clearly join the category of ‘extremophile’ propagules.
Ecological considerations
Although wild asexual propagules of Ditrichum plumbicola were not detected in the present study, they have been observed previously. Arts (1994) described the occasional occurrence of rhizoidal tubers comprising uniseriate rows of five to eight swollen thick-walled cells, 50–90 µm in diameter, terminated distally by smaller (20–30 µm) lenticular apical cells. The tubers also regenerated into green chloronemata when placed on fresh medium. This description bears a striking resemblance to the brood bodies reported in the present study and it is likely that they are the same structures.
Analysis of both freshly collected rhizoids (Fig. 10C) and rhizoids kept as herbarium specimens (Fig. 10B) revealed that these were also packed with lipid and starch reserves similar to those in the ABA/sucrose-induced brood bodies. Again, they regenerated rapidly when transferred to fresh culture medium (Fig. 10E). The rapid formation of protonemata from structures such as rhizoids, tubers and brood bodies would likely give D. plumbicola a competitive advantage after cryoturbatic disturbance of the soil over species, such as Pogonatum aloides, the protonemal/rhizoidal systems of which lack asexual propagules (Duckett et al., 2004). When the ground remains undisturbed, Ditrichum does indeed become overrun by the perennial protonemata of Pogonatum (Fig. 10A) and other slower growing, but ultimately more competitive species. This is further demonstrated by another pioneer moss, Discelium nudum, which also gains a temporal advantage over ultimately more successful competitors, following winter exfoliation of the clay banks on which it grows (Duckett and Pressel, 2003). These data support a growing body of evidence that asexual propagules are potentially very important in the continued survival and dispersal of mosses, including threatened species (Preston, 2004; Mallon et al., 2006).
The role of natural endogenous ABA in the desiccation and freezing tolerance of D. plumbicola merits further consideration. Certainly the marked tendency to dedifferentiate into brood cells exhibited by protonemal cultures as young as 2–3 weeks, suggests that high levels of endogenous ABA may be present. The production of brood cells in vitro is usually associated with ageing (presumably in response to dehydration and/or accumulation of ABA in the medium) or with exposure to exogenous ABA (Goode, 1992; Goode et al., 1993a). While it is well known that exogenous ABA increases the desiccation and freezing tolerance of bryophytes (e.g. Beckett et al., 2000; Nagao et al., 2005; Pence et al., 2005), only a handful of studies have reported the presence of endogenous ABA (Hartung et al., 1987; Werner et al., 1991; Hellwege et al., 1994), with only two demonstrating an increase in endogenous ABA levels during drying (Werner et al., 1991; Hellwege et al., 1994). Clearly, and as discussed previously by Proctor and Tuba (2002), there is now the need for a systematic search for endogenous ABA in bryophytes and of a comprehensive study to determine whether endogenous ABA levels increase during drying.
CONCLUSIONS
Exposure to a combination of exogenous sucrose and ABA causes the protonema of the moss D. plumbicola to transform into specialized resting propagules or brood bodies. The cytology of these is consistent with that of moss brood cells previously described. The brood bodies are able to withstand extreme desiccation and cryopreservation stress, confirming that they are resting cells pre-adapted to withstand desiccative stress. The formation of such cells confers on D. plumbicola an additional survival strategy, and this has been exploited for the conservation of the species via cryopreservation.
ACKNOWLEDGEMENTS
We thank English Nature (Natural England), Scottish Natural Heritage and Countryside Council for Wales for funding the ex situ bryophyte project, and the Natural History Museum, lead partner on the Biodiversity Action Plan for Ditrichum plumbicola, for permission to collect material for the project and this study. Dr R. Preziosi, University of Manchester, gave statistical advice. S.P. held a Natural Environment Research Council (UK)/CASE studentship with the Royal Botanic Gardens, Kew. Thanks also to two anonymous reviewers for comments on an earlier draft of this manuscript.
LITERATURE CITED
- Alpert P. Constraints of tolerance: why are desiccation-tolerant organisms so small or rare? Journal of Experimental Biology. 2006;209:1575–1584. doi: 10.1242/jeb.02179. [DOI] [PubMed] [Google Scholar]
- Arts T. Rhizoidal tubers and protonemal gemmae in European Ditrichum species. Journal of Bryology. 1994;18:43–61. [Google Scholar]
- Beckett RP. ABA-induced tolerance to ion leakage during rehydration following desiccation in the moss Atrichum androgynum. Plant Growth Regulation. 2001;35:131–135. [Google Scholar]
- Beckett RP, Csintalan Z, Tuba Z. ABA treatment increases both the desiccation tolerance of photosynthesis, and nonphotochemical quenching in the moss Atrichum undulatum. Plant Ecology. 2000;151:65–71. [Google Scholar]
- Benson EE. Cryopreservation. In: Benson EE, editor. Plant conservation biotechnology. London: Taylor & Francis; 1999. pp. 83–95. [Google Scholar]
- Bequerel P. La suspension de la vie des algues, lichens, mousses au zero absolu et role de la synérèse reversible pour l'existence de la flore polaire et des hautes altitudes. Compte Rendu Hébomadaire des Séances de l'Académie des Sciences. 1951;232:22. [Google Scholar]
- Bewley JD. The effect of liquid nitrogen temperatures on proteins and RNA synthesis in the moss Tortula ruralis. Plant Science Letters. 1973;1:303–308. [Google Scholar]
- Bianchi G, Gamba A, Murelli C, Salamini F, Bartels D. Novel carbohydrate metabolism in the resurrection plant Craterostigma plantagineum. The Plant Journal. 1991;1:355–359. doi: 10.1046/j.1365-313X.1991.t01-11-00999.x. [DOI] [PubMed] [Google Scholar]
- Bopp M, Werner O. Abscisic acid and desiccation tolerance in mosses. Botanica Acta. 1993;106:103–106. [Google Scholar]
- Brown RC, Lemmon BE. Division polarity, development and configuration of microtubule arrays in bryophyte meiosis. 2. Anaphase-I to the tetrad. Protoplasma. 1987;138:1–10. [Google Scholar]
- Brown RC, Lemmon BE. Sporogenesis in bryophytes. In: Miller NG, editor. Bryophyte ultrastructure. Stuttgart: Gebrüder Borntraeger Verlagsbuchhandlung; 1988. pp. 159–223. [Google Scholar]
- Buitink J, Leprince O. Glass formation in plant anhydrobiotes: survival in the dry state. Cryobiology. 2004;48:215–228. doi: 10.1016/j.cryobiol.2004.02.011. [DOI] [PubMed] [Google Scholar]
- Burch J. Some mosses survive cryopreservation without prior pretreatment. Bryologist. 2003;106:270–277. [Google Scholar]
- Burch J, Wilkinson T. Cryopreservation of protonemata of Ditrichum cornubicum (Paton) comparing the effectiveness of four cryoprotectant pretreatments. Cryoletters. 2002;23:197–208. [PubMed] [Google Scholar]
- Church JM, Hodgetts NG, Preston CD, Stewart NF. British red data books – mosses and liverworts. Peterborough: JNCC; 2001. [Google Scholar]
- Clegg JS. Desiccation tolerance in encysted embryos of the animal extremophile, Artemia. Integrative and Comparative Biology. 2005;45:715–724. doi: 10.1093/icb/45.5.715. [DOI] [PubMed] [Google Scholar]
- Dalla Vecchia F, El Asmar T, Calamassi R, Rascio N, Vazzana C. Morphological and ultrastructural aspects of dehydration and rehydration in leaves of Sporobolus stapfianus. Plant Growth Regulation. 1998;24:219–228. [Google Scholar]
- Duckett JG, Ligrone R. Gemma germination and morphogenesis of a highly differentiated gemmiferous protonema in the tropical moss Calymperes (Calymperaceae, Musci) Cryptogamic Botany. 1991;2/3:219–228. [Google Scholar]
- Duckett JG, Ligrone R. A survey of diaspore liberation mechanisms and germination patterns in mosses. Journal of Bryology. 1992;17:335–354. [Google Scholar]
- Duckett JG, Pressel S. Studies of protonemal morphogenesis in mosses. IX. Discelium nudum: exquisite pioneer of unstable clay banks. Journal of Bryology. 2003;25:241–245. [Google Scholar]
- Duckett JG, Burch J, Fletcher PW, Matcham HW, Read DJ, Russell AJ, Pressel S. In vitro cultivation of bryophytes: a review of practicalities, problems, progress and promise. Journal of Bryology. 2004;26:3–20. [Google Scholar]
- Fang JY, Wetten A, Hadley P. Cryopreservation of cocoa (Theobroma cacao L.) somatic embryos for long-term germplasm storage. Plant Science. 2004;166:669–675. [Google Scholar]
- Gaff DF, Zee SY, Obrien TP. Fine-structure of dehydrated and reviving leaves of Borya nitida Labill., a desiccation-tolerant plant. Australian Journal of Botany. 1976;24:225–236. [Google Scholar]
- Gerdol R, Bonora A, Gualandri R, Pancaldi S. CO2 exchange, photosynthetic pigment composition, and cell ultrastructure of Sphagnum mosses during dehydration and subsequent rehydration. Canadian Journal of Botany – Revue Canadienne de Botanique. 1996;74:726–734. [Google Scholar]
- Glime JM, Carr RE. Temperature survival of Fontinalis novae-angliae Sull. The Bryologist. 1974;77:17–22. [Google Scholar]
- Goode JA. The moss protonema: morphogenesis and the cytoskeleton. University of London; 1992. PhD Thesis. [Google Scholar]
- Goode JA, Stead AD, Duckett JG. Redifferentiation of moss protonemata – an experimental and immunofluorescence study of brood cell formation. Canadian Journal of Botany – Revue Canadienne de Botanique. 1993a;71:1510–1519. [Google Scholar]
- Goode JA, Stead AD, Duckett JG. Studies of protonemal morphogenesis in mosses. 2. Orthotrichum obtusifolium Brid. Journal of Bryology. 1993b;17:409–419. [Google Scholar]
- Goode JA, Stead AD, Ligrone R, Duckett JG. Studies of protonemal morphogenesis in mosses. 4. Aloina (Pottiales) Journal of Bryology. 1994;18:27–41. [Google Scholar]
- Guerrant EO, Jr, Havens K, Maunder M. Ex situ plant conservation – supporting species survival in the wild. Washington, DC: Island Press; 2004. [Google Scholar]
- Guschina IA, Harwood JL, Smith M, Beckett RP. Abscisic acid modifies the changes in lipids brought about by water stress in the moss Atrichum androgynum. New Phytologist. 2002;156:255–264. doi: 10.1046/j.1469-8137.2002.00517.x. [DOI] [PubMed] [Google Scholar]
- Hartung W, Weiler EW, Volk OH. Immunochemical evidence that abscisic acid is produced by several species of Anthocerotae and Marchantiales. Bryologist. 1987;90:393–400. [Google Scholar]
- Hearnshaw GF, Proctor MCF. The effect of temperature on the survival of dry bryophytes. New Phytologist. 1982;90:221–228. [Google Scholar]
- Hellwege EM, Dietz KJ, Volk OH, Hartung W. Abscisic acid and the induction of desiccation tolerance in the extremely xerophilic liverwort Exormotheca holstii. Planta. 1994;194:525–531. [Google Scholar]
- Hellwege EM, Volk OH, Hartung W. A physiological role of abscisic acid in the liverwort Riccia fluitans L. Journal of Plant Physiology. 1992;140:553–556. [Google Scholar]
- Klein S, Pollock BM. Cell fine structure of developing lima bean seeds related to seed desiccation. American Journal of Botany. 1968;55:658–672. [Google Scholar]
- Linders LJM, Wolkers WF, Hoekstra FA, vantRiet K. Effect of added carbohydrates on membrane phase behavior and survival of dried Lactobacillus plantarum. Cryobiology. 1997;35:31–40. doi: 10.1006/cryo.1997.2021. [DOI] [PubMed] [Google Scholar]
- Mallon R, Reinoso J, Rodriguez-Oubina J, Gonzalez ML. In vitro development of vegetative propagules in Splachnum ampullaceum: brood cells and chloronematal bulbils. Bryologist. 2006;109:215–223. [Google Scholar]
- Mueller DR, Vincent WF, Bonilla S, Laurion I. Extremotrophs, extremophiles and broadband pigmentation strategies in a high arctic ice shelf ecosystem. FEMS Microbiology Ecology. 2005;53:73–87. doi: 10.1016/j.femsec.2004.11.001. [DOI] [PubMed] [Google Scholar]
- Nagao M, Minami A, Arakawa K, Fujikawa S, Takezawa D. Rapid degradation of starch in chloroplasts and concomitant accumulation of soluble sugars associated with ABA-induced freezing tolerance in the moss Physcomitrela patens. Journal of Plant Physiology. 2005;162:169–180. doi: 10.1016/j.jplph.2004.06.012. [DOI] [PubMed] [Google Scholar]
- Navari-Izzo F, Quartacci MF, Pinzino C, Rascio N, Vazzana C, Sgherri CLM. Protein dynamics in thylakoids of the desiccation-tolerant plant Boea hygroscopica during dehydration and rehydration. Plant Physiology. 2000;124:1427–1436. doi: 10.1104/pp.124.3.1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norr M. Heat resistance of mosses. Flora. 1974;163:388–397. [Google Scholar]
- Norwood M, Truesdale MR, Richter A, Scott P. Photosynthetic carbohydrate metabolism in the resurrection plant Craterostigma plantagineum. Journal of Experimental Botany. 2000;51:159–165. doi: 10.1093/jexbot/51.343.159. [DOI] [PubMed] [Google Scholar]
- Oliver MJ, Tuba Z, Mishler BD. The evolution of vegetative desiccation tolerance in land plants. Plant Ecology. 2000a;151:85–100. [Google Scholar]
- Oliver MJ, Velten J, Wood AJ. Bryophytes as experimental models for the study of environmental stress tolerance: Tortula ruralis and desiccation-tolerance in mosses. Plant Ecology. 2000b;151:73–84. [Google Scholar]
- Pence VC. Cryopreservation of bryophytes: the effects of abscisic acid and encapsulation dehydration. Bryologist. 1998;101:278–281. [Google Scholar]
- Pence VC. Cryopreservation of in vitro grown fern gametophytes. American Fern Journal. 2000;90:16–23. [Google Scholar]
- Pence VC, Dunford SS, Redella S. Differential effects of abscisic acid on desiccation tolerance and carbohydrates in three species of liverworts. Journal of Plant Physiology. 2005;162:1331–1337. doi: 10.1016/j.jplph.2005.02.002. [DOI] [PubMed] [Google Scholar]
- Porley RD, Hodgetts N. Mosses and liverworts. London: Harper Collins; 2005. [Google Scholar]
- Pressel S, Matcham HW, Duckett JG. Studies of protonemal morphogenesis in mosses. X. Ephemeraceae revisited; new dimensions underground. Journal of Bryology. 2005;27:311–318. [Google Scholar]
- Pressel S, Ligrone R, Duckett JG. Effects of de- and rehydration on food-conducting cells in the moss Polytrichum formosum: a cytological study. Annals of Botany. 2006;98:67–76. doi: 10.1093/aob/mcl092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preston CD. An updated list of British and Irish bryophytes from which tubers have been reported. Field Bryology. 2004;83:2–13. [Google Scholar]
- Proctor M. Patterns of desiccation tolerance and recovery in bryophytes. Plant Growth Regulation. 2001;35:147–156. [Google Scholar]
- Proctor MCF, Tuba Z. Poikilohydry and homoihydry: antithesis or spectrum of possibilities? New Phytologist. 2002;156:327–349. doi: 10.1046/j.1469-8137.2002.00526.x. [DOI] [PubMed] [Google Scholar]
- Proctor MCF, Ligrone R, Duckett JG. Desiccation tolerance in the moss Polytrichum formosum: physiological and fine-structural changes during desiccation and recovery. Annals of Botany. 2007;99:75–93. doi: 10.1093/aob/mcl246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quartacci MF, Forli M, Rascio N, DallaVecchia F, Bochicchio A, NavariIzzo F. Desiccation-tolerant Sporobolus stapfianus: lipid composition and cellular ultrastructure during dehydration and rehydration. Journal of Experimental Botany. 1997;48:1269–1279. [Google Scholar]
- Ramsay MM, Burch J. Ex situ techniques in support of UK bryophyte conservation. Novitates Botanicae ex Universitatis Carolinae Pragensis. 2001;15:27–33. [Google Scholar]
- Rowntree JK, Sheffield E, Burch J. Growth and development of mosses are inhibited by the common herbicide asulam. Bryologist. 2005;108:287–294. [Google Scholar]
- Scheffe H. The analysis of variance. New York, NY: John Wiley and Sons; 1959. [Google Scholar]
- Schnepf E, Reinhard C. Brachycytes in Funaria protonemata: induction by abscisic acid and fine structure. Journal of Plant Physiology. 1997;151:166–175. [Google Scholar]
- Side AG, Whitehouse HLK. Colorless tubers in Discelium nudum Brid. Journal of Bryology. 1987;14:741–743. [Google Scholar]
- Sokal RR, Rohlf FJ. Biometry. 3rd edn. New York, NY: W.H. Freeman & Co; 1995. [Google Scholar]
- Sreedhar L, Wolkers WF, Hoekstra FA, Bewley JD. In vivo characterization of the effects of abscisic acid and drying protocols associated with the acquisition of desiccation tolerance in alfalfa (Medicago sativa L.) somatic embryos. Annals of Botany. 2002;89:391–400. doi: 10.1093/aob/mcf057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki M, Ishikawa M, Okuda H, Noda K, Kishimoto T, Nakamura T, et al. Physiological changes in gentian axillary buds during two-step preculturing with sucrose that conferred high levels of tolerance to desiccation and cryopreservation. Annals of Botany. 2006;97:1073–1081. doi: 10.1093/aob/mcl054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Systat Software. Richmond, CA: 2004. Systat statistics I. Systat Software Inc. [Google Scholar]
- Thomson WW, Platt KA. Conservation of cell order in desiccated mesophyll of Selaginella lepidophylla ([Hook and Grev] Spring) Annals of Botany. 1997;79:439–447. [Google Scholar]
- Tucker EB, Costerton JW, Bewley JD. The ultrastructure of the moss Tortula ruralis on recovery from desiccation. Canadian Journal of Botany – Revue Canadienne de Botanique. 1975;53:94–101. [Google Scholar]
- Valadon LRG, Mummery RS. Quantitative relationship between various growth substances and bud production in Funaria hygrometrica – bioassay for abscisic acid. Physiologia Plantarum. 1971;24:232–234. [Google Scholar]
- Vander Willigen C, Pammenter NW, Jaffer MA, Mundree SG, Farrant JM. An ultrastructural study using anhydrous fixation of Eragrostis nindensis, a resurrection grass with both desiccation-tolerant and -sensitive tissues. Functional Plant Biology. 2003;30:281–290. doi: 10.1071/FP02221. [DOI] [PubMed] [Google Scholar]
- Vicre M, Lerouxel O, Farrant J, Lerouge P, Driouich A. Composition and desiccation-induced alterations of the cell wall in the resurrection plant Craterostigma wilmsii. Physiologia Plantarum. 2004;120:229–239. doi: 10.1111/j.0031-9317.2004.0234.x. [DOI] [PubMed] [Google Scholar]
- Werner O, Espin RMR, Bopp M, Atzorn R. Abscisic acid induced drought tolerance in Funaria hygrometrica Hedw. Planta. 1991;186:99–103. doi: 10.1007/BF00201503. [DOI] [PubMed] [Google Scholar]
- Whitehouse HLK. Occurrence of tubers in Pohlia pulchella (Hedw) Lindb. and Pohlia lutescens (Limpr) Lindb Fil. Journal of Bryology. 1973;7:533–540. [Google Scholar]
- Whitehouse HLK. The production of protonemal gemmae by mosses growing in deep shade. Journal of Bryology. 1980;11:133–138. [Google Scholar]
- Zar JH. Biostatistical analysis. 4th edn. Upper Saddle River, NJ: Prentice-Hall; 1999. [Google Scholar]
- Zhu G-Y, Geuns JMC, Dussert S, Swennen R, Panis B. Change in sugar, sterol and fatty acid composition in banana meristems caused by sucrose-induced acclimation and its effects on cryopreservation. Physiologia Plantarum. 2006;128:80–94. [Google Scholar]