

Abstract
The intermediate filament protein vimentin is a major phosphoprotein in mammalian fibroblasts, and reversible phosphorylation plays a key role in its dynamic rearrangement. Selective inhibition of type 2A but not type 1 protein phosphatases led to hyperphosphorylation and concomitant disassembly of vimentin, characterized by a collapse into bundles around the nucleus. We have analyzed the potential role of one of the major protein phosphatase 2A (PP2A) regulatory subunits, B55, in vimentin dephosphorylation. In mammalian fibroblasts, B55 protein was distributed ubiquitously throughout the cytoplasm with a fraction associated to vimentin. Specific depletion of B55 in living cells by antisense B55 RNA was accompanied by disassembly and increased phosphorylation of vimentin, as when type 2A phosphatases were inhibited using okadaic acid. The presence of B55 was a prerequisite for PP2A to efficiently dephosphorylate vimentin in vitro or to induce filament reassembly in situ. Both biochemical fractionation and immunofluorescence analysis of detergent-extracted cells revealed that fractions of PP2Ac, PR65, and B55 were tightly associated with vimentin. Furthermore, vimentin-associated PP2A catalytic subunit was displaced in B55-depleted cells. Taken together these data show that, in mammalian fibroblasts, the intermediate filament protein vimentin is dephosphorylated by PP2A, an event targeted by B55.
INTRODUCTION
Protein phosphatase 2A (PP2A) is implicated in a significant array of cellular processes, including metabolism, DNA replication, transcription, translation, cell cycle progression, and membrane-to-nuclear signal transduction (for review, see Shenolikar, 1994; Wera and Hemmings, 1995). Regulatory flexibility is conferred by the association of a constant dimeric core of a 36-kDa catalytic (PP2Ac) and a 65-kDa (PR65 or A) subunit with a third, variable B subunit (Mayer-Jaekel and Hemmings, 1994). To date three families of B subunits have been identified, which we will refer to as B55, B56, and B72, according to the predicted molecular weight of their founding member (Mayer et al., 1991; Hendrix et al., 1993a; McCright and Virshup, 1995). At least 10 individual genes code for B-type regulators, with this number being further increased by the additional presence of alternatively spliced forms of certain messages (Hendrix et al., 1993a; Csortos et al., 1996; McCright et al., 1996; Tanabe et al., 1996; Tehrani et al., 1996; Zolnierowicz et al., 1996). By analogy to PP1, in which associated noncatalytic subunits target the catalytic subunit to different subcellular compartments and/or substrates, it was proposed that the B-type regulators act as targeting subunits for PP2A (Hubbard and Cohen, 1993). Indeed, B subunits affect the substrate specificity of PP2A in vitro and in vivo (Agostinis et al., 1987; Cegielska et al., 1994; Kamibayashi et al., 1994; Mayer-Jaekel et al., 1994; Zhao et al., 1997). Distinct subcellular localization has been reported for some B subunits (McCright et al., 1996; Tehrani et al., 1996). The 55-kDa regulatory subunit B55 (or PR55) was one of the first B-type regulators to be identified in skeletal muscle preparations of PP2A, and subsequent cDNA cloning revealed evolutionary conservation from yeast to human (Healy et al., 1991; Mayer et al., 1991; Pallas et al., 1992; Mayer-Jaekel et al., 1993). The lack of functional B55 in yeast and Drosophila causes severe aberrations in mitotic transit (Healy et al., 1991; Mayer-Jaekel et al., 1993; Wang and Burke, 1997) presumably because of the lack of dephosphorylation of certain p34cdc2 phosphorylated sites (Ferrigno et al., 1993; Mayer-Jaekel et al., 1994). It was recently also reported that B55 associates with the microtubule network (Sontag et al., 1995).
Intermediate filaments (IFs) are major components of the cytoskeleton and nuclear envelope. They are characterized by 10 nm diameter and marked insolubility (for review, see Fuchs and Weber, 1994). The IF superfamily comprises five classes of proteins, which appear to share common secondary structure. Cytoplasmic IF proteins include vimentin in fibroblasts, cytokeratins in epithelial cells, neurofilaments and/or glial fibrillary acidic protein in neuronal tissues and desmin in muscle cells. IF protein are among the most prominent phosphoproteins found in cells, and reversible phosphorylation has been shown to play a key role in their dynamic rearrangement (for review, see Inagaki et al., 1996). For example the type III IF vimentin has been shown to be phosphorylated in vitro and/or in vivo by various protein kinases, including PKA and PKC, and p34cdc2. Most of these protein kinases phosphorylate vimentin in its amino-terminal head domain, resulting in disassembly and bundle formation. Inhibition of the phosphoserine–threonine protein phosphatases using okadaic acid (OA) or related compounds also leads to hyperphosphorylation and disassembly of vimentin IF networks (Yatsunami et al., 1991; Eriksson et al., 1992; Lee et al., 1992).
We have previously shown that specific inhibition of type 2A but not type 1 protein phosphatases1 induces hyperphosphorylation of cytokeratin 8 and 18 in MCF7 epithelial cells (Favre et al., 1997). In this report we have studied IF dephosphorylation in mammalian Hs68 fibroblasts. By a number of criteria, we identified the heterotrimeric PP2A-containing B55 regulatory subunit as the major vimentin phosphatases in interphase Hs68 cells. Specific inhibition of type 2A protein phosphatases using OA led to dramatic hyperphosphorylation and disassembly of vimentin. Depletion of B55 in living cells led to a similar vimentin disassembly and increased phosphorylation. Recombinant B55 strongly increased PP2A activity toward vimentin in vitro and in situ. Moreover, a fraction of cytoplasmic PP2A, namely PP2Ac, PR65, and B55, was found tightly associated with vimentin and could be displaced from vimentin by functional knockout of B55. Taken together these results suggest that PP2A, targeted by the B55 regulatory subunit, dephosphorylates vimentin directly.
MATERIALS AND METHODS
Subunit-specific Antibodies
Anti-PP1 and anti-PP2A antibodies AbCrecomb, AbC299/309, AbC169/182, Ab65recomb, and Ab65177/196 were used as described previously (Fernandez et al., 1992; Turowski et al., 1995; Favre et al., 1994). The B55 peptide CTNNLYIFQDKVN (Synthem, Nimes, France), corresponding to the carboxyl-terminal region of B55 α, β, and γ (Mayer et al., 1991; Zolnierowicz et al., 1994) was coupled to thyroglobulin using sulfo-m-maleimidobenzoyl-N-hydroxysulfo-succinimide ester (Pierce, Rockford, IL). Rabbits were immunized using 0.3–1 mg of the peptide–protein conjugate. Sera containing Ab55437/448 were screened by immunoblotting against recombinant B55 (Michelson et al., 1996) and affinity purified using the immunogenic peptide (conjugated to BSA) immobilized on cyanogen bromide-Sepharose (Pharmacia, Orsay, France) (Hendrix et al., 1993b). Monospecific Ab55recomb (Michelson et al., 1996) were obtained by absorption to recombinant B55α transferred to a polyvinylidene difluoride membrane and subsequent elution using 50 mM glycine, pH 2.3, 500 mM NaCl, 0.5% Tween 20, and 0.1 mg/ml BSA as described (Mayer-Jaekel et al., 1994). Affinity-purified antibodies were concentrated in the presence of BSA by dialysis against sucrose and redialysis against PBS as detailed elsewhere (Turowski and Lamb, 1998).
Expression Plasmids and Transient Transfections
An XbaI site was introduced by PCR into full-length human B55α cDNA (Mayer et al., 1991) at position −18. Subsequently the 2-kb XbaI fragment (using the 3′ XbaI from the multiple cloning site) was subcloned into the mammalian expression vector pECE (Ellis et al., 1986) in antisense orientation. The resulting construct pECE-B55as was verified by restriction endonuclease digestion and DNA sequence analysis. Hemagglutinin (HA)-tagged B55 and B56β were cloned into pCMV5 similar to the procedure described previously (Andjelkovic et al., 1996). B56 cDNAs were subcloned in antisense orientation into the pBI-EGPF Tet vector (Clontech, Palo Alto, CA), which was coinjected with pTet-Off (Clontech). For microinjection and transfection the plasmids were prepared by cesium chloride centrifugation using standard procedures (Sambrook et al., 1989).
Hs68 fibroblasts were transfected with either pECE-B55as or pCMV5-HA55α by the calcium phosphate method (Chen and Okayama, 1988) with the modifications described by Alessi et al. (1996). Five and 10 μg of pECE-B55as and pCMV5-HA55α, respectively, were used per 60-mm dish of Hs68 cells. Incubation with the DNA was done overnight, followed by 36 h of expression before analysis.
Microinjection and Immunofluorescence Analysis
Human Hs68 fibroblasts (CRL-1365) were cultured and synchronized by serum deprivation as described elsewhere (Girard et al., 1992). Cells were subcultured 2–3 d before use onto 12-mm acid-washed glass coverslips. Cells were injected as described (Lamb and Fernandez, 1997). Before injection the plasmid, pECE-B55as, was diluted to 0.1 mg/ml with PBS containing 1 mg/ml mouse, rabbit, or guinea pig immunoglobulin G (to act as a marker for injected cells).
For immunofluorescence studies, cells were fixed in 3.7% formaldehyde in PBS and extracted with acetone (−20°C) or in methanol (−20°C) as described (Turowski and Lamb, 1998). Alternatively, to stain for cytoskeletal associated proteins, cells were extracted with 0.01% Triton X-100 in PBS before fixation. Subsequently, cells were stained (Turowski et al., 1995; Lamb and Fernandez, 1997) using primary antibodies as follows: affinity-purified Ab55s at 1:10–1:20 (corresponding to a serum dilution of 1:100); affinity-purified AbCs at 1:50 (idem); monoclonal anti-vimentin (clone V9; Boehringer Mannheim, Meylan, France; and Sigma, St. Quentin Fallavier, France) at 1:10; monoclonal anti-tubulin (clone DM1A; Blose et al., 1984) at 1:2000; and Bodipy-phalloidin (Molecular Probes, Eugene, OR) at 1 U/coverslip. In competition experiments diluted Ab55s were preincubated for 30 min with immobilized recombinant B55. Microinjected cells were identified by costaining with FITC-conjugated anti-mouse or anti-rabbit antibodies (Organon Tecknika, Durham, NC) diluted 1:30 or amino-methylcoumarin–conjugated anti-guinea pig antibody diluted 1:10 (Jackson ImmunoResearch, West Grove, PA). DNA was visualized using Hoechst dye (1 μg/ml bisbenzimide). Cells were mounted and photographed on an Eastman Kodak (Rochester, NY) DCS 420 camera mounted on a Leica (Wetzlar, Germany) DMRB fluorescent microscope. Alternatively cells were photographed on a Zeiss (Thornwood, NY) Axiophot microscope using conventional photography on slide film (Girard et al., 1992). Images were scanned or acquired in Photoshop (Adobe Systems, Mountain View, CA) and further treated and mounted on a Silicon Graphics (Mountain View, CA) Indigo 2 workstation. In cases of double exposures the color balance of the scanned images was not varied to not distort colocalization information of the image. Confocal microscopy and fluorescence measurements of AbC169/182 were performed as previously described (using ImgCalc fluorescence sensitivity software; Turowski et al., 1995).
Metabolic Pulse Labeling of Injected Cells or Cells Treated with Phosphatase Inhibitors and Two-dimensional Gel Electrophoresis
Hs68 fibroblasts were grown on small (∼1 mm2) glass coverslips. All cells on the coverslip were counted and injected with pECE-B55as or pECE alone as a control and counted. For metabolic labeling the cells were transferred to a humidified chamber and overlaid with 7 μl of phosphate-free Dulbecco’s modified Eagle’s medium (supplemented with 8% dialyzed FCS) containing 1 mCi of [32P]H3PO4 (PBS13; Amersham, Les Ulis, France) as previously described (Lamb et al., 1989). Cells were briefly rinsed in PBS and lysed by dropping the chip into 30 μl of boiling 50 mM Tris-Cl, pH 7.5, 5 mM dithiothreitol, 0.5% SDS, and 2% NP-40 containing ∼106–107 unlabeled carrier cells. Samples were boiled, lyophilized, resuspended in 20 μl of 1 mg/ml RNase A (protease-free; Worthington, Freehold, NJ), and incubated for 10 min at 37°C. In a similar manner cells treated with OA or tautomycin (TAU) were metabolically labeled; Hs68 fibroblasts were grown on 12 mm coverslips to equal density and overlaid with 30 μl of medium containing OA at 1 μM or TAU at 10 μM and 3 mCi of [32P]H3PO4 (PBS13; Amersham). Cells were treated with OA and TAU for 1 and 3 h, respectively, as previously described (Favre et al., 1997). Labeling times were 1 h. Lysis was performed as described for chip-injected cells in a volume of 100 μl.
Before two-dimensional gel electrophoresis 150 μl of isoelectric focusing sample buffer (9.5 M urea, 4% NP-40, 0.1 M dithiothreitol, and 2% ampholines, pH 6–8) were added to each sample. Isoelectric focusing was performed according to the method of O’Farrell (1975) using ampholines, pH 3–10 (60%) and pH 4–7 (40%). Second dimensions were run on 12.5% acrylamide gels according to the method of Blattler et al. (1972). Gels were stained using Coomassie brilliant blue, dried, and exposed to autoradiography at −80°C on film using two intensifying screens. Alternatively, gels were quantified using a PhosphorImager and ImageQuant software (Molecular Dynamics, Bondoufle, France). The position of vimentin on two-dimensional gels was determined by immunblotting using the V9 mAb (Sigma).
Cell Fractionation, Vimentin Purification, and Immunoblotting
Total cell lysates and cytoplasmic and nuclear fractions of Hs68 fibroblasts were prepared as previously described (Turowski et al., 1995). Protein concentration in the soluble fractions was determined using a Bradford detection reagent (Bio-Rad, Ivry sur Seine, France) and BSA as a standard.
Extracts enriched in vimentin were essentially prepared as described by Franke et al. (1978). Briefly, subconfluent Hs68 were scraped into PBS, pelleted, and lysed in buffer V containing 50 mM HEPES, pH 7.2, 140 mM NaCl, 1% Triton X-100, 5 mM MgCl2, 1 mM dithiothreitol, and a mixture of protease inhibitors. The insoluble material was collected by centrifugation and subjected to three extractions in buffer V containing either 0.6 or 1.5 M KCl. The resulting vimentin pellet was solubilized in SDS-PAGE sample buffer. Proteins solubilized during the extraction steps were precipitated in 10% trichloroacetic acid at 4°C, extracted with ether:ethanol (4:1), and resuspended in the same volume of 1× SDS-PAGE sample buffer as the vimentin pellet to enable direct comparison.
Protein samples were made 1× in SDS-PAGE sample buffer (62.5 mM Tris-Cl, pH 6.8, 2% SDS, 8% glycerol, 0.001% bromphenol blue, and 10 mM dithiothreitol), boiled, and subjected to SDS-PAGE on 10% gels (Laemmli, 1970). Rabbit skeletal muscle PP2Ac (Stone et al., 1987), recombinant B55α (Michelson et al., 1996), recombinant PR65 (Turowski et al., 1997), and PP1 catalytic subunit (a gift from M. Bollen, Katholieke Universiteit, Leuven, Belgium) were included as positive controls. Proteins were subsequently electrotransferred onto an Immobilon-P membrane (Millipore, St. Quentin Yvelines, France), and immunodecoration was performed as described (Turowski et al., 1995). AbCs, Ab65s, and Ab55s were diluted 1:100–1:500, respectively. Anti-vimentin V9 (Sigma) was used at 1:400; anti-HA ascites (clone 12CA5; a generous gift from S. Leibovitch, Institut Gustave Roussy, Villejuif, France) was used at 1:10′000. Competition experiments were performed as described above for immunofluorescence.
Recombinant B55
The insert encoding human B55α (Mayer et al., 1991) was subcloned as a BamHI fragment into the baculovirus transfer vector pVL1392. The resulting plasmid was transfected with wild-type baculoviral DNA (AcMNPV) to generate recombinant baculovirus (AcHPR55α) by homologous recombination in Spodoptera frugiperda (Sf9) cells (Summers and Smith, 1987). For large-scale infection, 1 l of Sf9 was cultured in TC-100 medium supplemented with 10% FCS to a density of 1.5 × 106/ml, at which point they were infected with AcHPR55α at a multiplicity of infection of 5. Cells were harvested 72 h after infection and lysed in 50 mM Tris-Cl, pH 7.5, 100 mM NaCl, 1 mM EDTA, 1 mM DTT, 10% glycerol, 0.2% Triton X-100, 1 mM benzamidine, 0.1 mM tosyl-l-phenylanalylchloromethane, 0.1 mM tosyl-l-lysylchloromethylketone, 3 μM pepstatin A, 2 μM leupeptin, and 0.5 mM PMSF using a Dounce homogenizer. Subsequently, B55 was purified by chromatography on Q-Sepharose, heparin-Sepharose and Mono Q (all from Pharmacia). A detailed purification scheme will be published elsewhere (Myles and Hemmings, manuscript in preparation).
Protein Phosphatase Assays
Type 1 and 2A protein phosphatase activities of Hs68 cells remaining after OA and TAU treatment were determined using phosphorylase a and phosphopeptide as previously described (Favre et al., 1997). The functionality of recombinant B55 preparations was assayed using heterodimeric PP2A2 (a gift from J. Goris, Katholieke Universiteit, Leuven, Belgium), recombinant B55, and phosphorylase a at final concentrations of 1 nM, 10 nM, and 10 μM, respectively (Turowski et al., 1997). Alternatively, histone H1, phosphorylated by p34cdc2 (a gift from J.-C. Labbé, Centre de Recherche en Biochimie Macromoléculaire, Montpellier, France) was used as substrate at a final concentration of 5 μM (Mayer-Jaekel et al., 1994).
Vimentin Dephosphorylation In Vitro.
Six 60-mm dishes of Hs68 were each metabolically labeled with 2.5 ml of 90% phosphate-free medium containing 1 mCi of [32P]H3PO4 (PBS13; Amersham) and 1 μM OA for 1.5 h. Cells were scraped and recovered by centrifugation, washed in 20 mM HEPES and 150 mM NaCl, pH 7.4, and snap frozen. Subsequently, vimentin was purified as described above. Hyperphosphorylated vimentin was initially found to be completely insoluble. Solubilization was performed by incubation in 8 M urea, 50 mM Tris-Cl, and 10 mM DTT, pH 7.5, at 37°C for 15 min and dilution with SDS-PAGE sample buffer to 2× final (125 mM Tris-Cl, pH 6.8, 4% SDS, 16% glycerol, 0.002% bromphenol blue, and 20 mM DTT). The mixture was incubated for 1 h on a rocking platform, boiled, and centrifuged, and equal volumes were separated on SDS-PAGE. Proteins were transferred to nitrocellulose. The position of phosphovimentin was identified by autoradiography of the membrane and by immunoblotting of a slice of the membrane using antibody V9 (Sigma). Radioactive vimentin bands were excised and quantitated (∼5 μg of protein as estimated by staining with Ponceau red and 30,000 cpm, with <10% of variation between the different strips). Vimentin was partially renatured by incubation in 6 M guanidinium chloride and 50 mM Tris-Cl, pH 7.5, and stepwise dilution using Tris-buffered saline. Subsequently, strips were incubated in 100 μl of phosphatase buffer (50 mM Tris-Cl, 50 mM NaCl, 0.1 mM EDTA, 0.1% β-mercaptoethanol, and 0.67 mg/ml BSA) containing mixtures of 1 nM PP2A2, 10 nM B55, 10 nM OA, or 1 nM PP1c. Incubation was done at 30°C for 10–40 min. Reactions were stopped by retraction of the phosphovimentin strips, and the phosphate released into the reaction mixture was quantitated by liquid scintillation counting.
Vimentin Dephosphorylation In Situ.
Hs68 fibroblasts were grown on coverslips and treated with 1 μM OA for 1 h. Cells were then permeabilized in buffer V for 2 min, and soluble cellular proteins were extracted by subsequent extraction in buffer V supplemented with 1.5 M KCl for 10 min (Franke et al., 1978). Coverslips coated with vimentin lattices were equilibrated in protein phosphatase assay buffer (50 mM Tris-Cl, pH 7.5, 50 mM NaCl, 0.1 mM EDTA, 0.67 mg/ml BSA, and 0.1% β-mercaptoethanol). For dephosphorylation assays, coverslips were overlaid with 30 μl of phosphatase mixture diluted in assay buffer and incubated at 37°C in a humidified chamber. PP2A2, B55, and OA were used at final concentrations of 1, 10, and 10 nM, respectively. At various times thereafter (between 0 and 45 min) reactions were stopped by fixation in formalin and further processed for immunofluorescence analysis of vimentin.
RESULTS
Inhibition of Type 2A Protein Phosphatases Leads to Hyperphosphorylation and Disassembly of Vimentin
Type 1 and type 2A protein serine-threonine phosphatases1 can be selectively inhibited by defined concentrations of TAU and OA, respectively (Favre et al., 1997). Significantly higher concentrations of each compound, compared with their reported Ki values, are required to effectively inhibit each class of protein phosphatases in cultured MCF7 human breast cancer cells. We have also shown that such inhibition of type 2A but not type 1 protein phosphatases leads to hyperphosphorylation of cytokeratin IFs in MCF7 cells (Favre et al., 1997). Here we have examined whether vimentin, the IF protein present in fibroblasts, became hyperphosphorylated by similar, selective inhibition of type 2A protein phosphatases. In Hs68 human fibroblasts TAU and OA inhibited type 1 and type 2A activities at doses and time courses comparable with those found for MCF7 cells. Specifically, treatment with 1 μM OA for 1 h resulted in an inhibition of type 2A activity of 99% (±5% [SE]; n = 4), whereas type 1 activity was nearly unaffected (13 ± 8% inhibition; n = 4). In contrast, treatment with 10 μM TAU for 3 h inhibited type 1 phosphatases by 81% (±5%; n = 4), whereas type 2A activity remained normal (107 ± 6%; n = 4). In subsequent experiments, equal numbers of Hs68 fibroblasts were treated for either 1 h with 1 μM OA or 3 h with 10 μM TAU. Cells were metabolically labeled with [32P]H3PO4, and cellular phosphoproteins were analyzed by two-dimensional gel electrophoresis and autoradiography (Figure 1A). Vimentin was heavily hyperphosphorylated in cells treated with OA when compared with that of control or TAU-treated cells (Figure 1A, upper panel). In contrast, TAU treatment led to hyperphosphorylation of myosin light chain (Figure 1A, lower panel), a bona fide substrate for PP1 in fibroblasts (Fernandez et al., 1990). OA treatment rather induced a diminution of myosin light chain phosphorylation, which may be attributable to a role of PP2A in regulating myosin light chain kinase. We identified at least six other phosphoproteins (including src and hsp 90), the phosphorylation state of which was unaffected by OA or TAU treatment. Thus neither of the two inhibitors exhibited generalized effects on protein phosphorylation (such as affecting the cellular ATP pool), in agreement with previous reports (Haystead et al., 1989). To confirm that OA and TAU acted differentially on the cytoskeleton, Hs68 fibroblasts, incubated with either inhibitor, were analyzed for their vimentin and F-actin distribution by indirect immunofluorescence (Figure 1B). In OA-treated cells the normally fine, well-extended vimentin network bundled, forming a cage-like structure around the nucleus, characteristic of its disassembly in vivo (Inagaki et al., 1996, and references therein). Although the cells rounded up, no significant reorganization of stress fibers took place. In contrast, TAU treatment led to increased staining for phalloidin, indicative of enhanced polymerization of G- to F-actin. Again this observation is in agreement with the reported role of PP1 in dephosphorylating myosin light chain (Fernandez et al., 1990). Vimentin from TAU-treated cells underwent limited reorganization, but in contrast to OA, no thick bundles were observed. Under the experimental conditions used only 30–40% of the cells showed modifications of the cytoskeleton described here. After longer incubation times all cells showed modified phenotypes. However, more strongly inhibited cells rounded completely and frequently detached, making high-resolution microscopy impossible. For this reason we used the shorter incubation times when some cells had not yet reacted phenotypically to the drugs. From these results it is clear that a type 2A protein phosphatase is implicated in the modulation of vimentin phosphorylation and organization in Hs68 fibroblasts.
Figure 1.

Analysis of phosphoproteins and the cytoskeleton of OA- and TAU-treated cells. (A) Equal numbers of Hs68 fibroblasts, grown on 12-mm glass coverslips, were treated with OA or TAU and metabolically labeled with [32P]H3PO4, and the totality of cellular proteins was separated by two-dimensional gel electrophoresis as described in MATERIALS AND METHODS. Shown are portions of the autoradiograms containing vimentin (upper panels) and myosin light chain (lower panels). Untreated, phosphoproteins of untreated cells labeled for 60 min; Okadaic Acid, phosphoproteins of cells treated with 1 μM OA during the 60-min labeling period; Tautomycin, phosphoproteins of cells treated with 10 μM TAU during 2 h and the subsequent 60-min labeling period. Autoradiographs were exposed at −80°C for 2 h for vimentin and for 6 h for MLC. (B) Hs68 fibroblasts were either left untreated or incubated with OA (1 μM for 45 min) or TAU (10 μM for 2 h). Subsequently they were fixed in formalin and costained for vimentin and F-actin using mAb V9 and Bodipy-phalloidin, respectively. Bar, 5 μm.
The B55 Subunit Is a Ubiquitous, Cytoplasmic Protein
Taking into account the abundance of vimentin in fibroblasts, our further studies concentrated on a possible role of PP2A in the modulation of vimentin phosphorylation. PP2A constitutes a class of heterotrimeric enzymes in which a variable protein, termed B, confers substrate specificity (Mayer-Jaekel and Hemmings, 1994). We chose to study the B subunit involved in vimentin phosphorylation, because this would also conclusively identify PP2A as the vimentin phosphatase. Our initial studies examined the subcellular distribution of a major PP2A regulatory subunit in Hs68 fibroblasts, namely B55. Two B55-specific antisera were generated: Ab55473/448 against a carboxyl-terminal peptide of all three B55 isoforms and Ab55recomb, for which full-length recombinant B55 was used as antigen (Michelson et al., 1996). After affinity purification both antibodies were monospecific in immunoblot analyses of human Hs68 fibroblasts (Figure 2, A and B). A single band at ∼52 kDa, comigrating with recombinant B55, was revealed in total extracts of Hs68 cells. Soluble and insoluble fractions from cytoplasm and nuclei of Hs68 fibroblasts were prepared and subjected to immunoblotting. As shown in Figure 2B, B55 was only detectable in the soluble cytoplasmic fraction. A more detailed subcellular analysis was performed using the anti-B55 antibodies in indirect immunofluorescence of mammalian fibroblasts. As illustrated in Figure 2, C and E, Ab55473/448 as well as Ab55recomb stained essentially the cytoplasmic compartment. Some of the cytoplasmic staining of Ab55recomb (Figure 2, E and G) was fine and filamentous, reminiscent of IF network staining and possibly attributable to a direct association of B55 with vimentin. For both antibodies, the nuclei were nearly devoid of staining (Figure 2, C and E), which was confirmed by confocal sections through the nucleus, which revealed that <5% of B55 was nuclear (Figure 2G). Confocal sectioning also showed the light filamentous cytoplasmic staining pattern. The staining pattern of both antibodies was specific, because it was essentially lost by preincubating the antibodies with recombinant B55 (Figure 2, D, F, and H). Similar staining patterns were also observed when methanol was used as a fixative or in REF-52 cells. Expression of HA-tagged B55 by microinjection or transfection in Hs68 or 293 cells, respectively, revealed a similar, ubiquitous distribution in the cytoplasm (our unpublished results). To further confirm the specificity of this subcellular localization and for subsequent functional studies, an antisense B55 RNA-expressing construct, pECE-B55as, was prepared. When transiently transfected in Hs68 fibroblasts, a significant decrease of the B55 signal was observed by immunoblot analysis of total extracts 36 h after transfection (Figure 3A). Given a transfection efficiency of ∼60%, the antisense construct pECE-B55as appeared an effective tool in depleting cells of B55. Similarly, the microinjection of pECE-B55as induced a marked loss of anti-B55 staining in a time-dependent manner (Figure 3, B and C). Within 15 h pECE-B55as induced a 71% reduction (±11%; n = 23) of B55 immunofluorescence. Taken together these data suggested that B55 is a cytoplasmic protein which is in part associated with cytoskeletal structures, most likely vimentin (also see below).
Figure 2.

Subcellular distribution of the B55 subunit in Hs68 fibroblasts. (A) Approximately 20 μg of total cell extracts from Hs68 fibroblasts were electrophoretically separated (lane T) and immunoblotted; ∼10 ng of recombinant B55α (lane C) were included as positive control. Immunoblots were decorated with affinity-purified Ab55473/448. (B) Hs68 fibroblasts were differentially extracted for soluble cytoplasm (lane 1), insoluble cytoplasm (lane 2), nucleoplasm (lane 3), and the nuclear pellet also containing microfilament and IFs (lane 4). Twenty micrograms of the soluble (lanes 1 and 3) and equal volumes of the insoluble (lanes 2 and 4) fractions were electrophoretically separated and immunoblotted using Ab55recomb. An identical subcellular distribution was obtained using affinity-purified Ab55473/448. Note that both antibodies recognize different proteolytic fragments of recombinant B55 (lane C), namely an amino terminus with Ab55recomb and a carboxyl terminus for Ab55473/448. (C–H) Hs68 fibroblasts were grown on glass coverslips and formalin fixed. Cells were stained for B55 using either Ab55473/448 (C and D) or Ab55recomb (E–H). In D, F, and H, antibodies were preincubated with recombinant B55 as described in MATERIALS AND METHODS. The images resulting from the same antibody staining were acquired and treated identically. (G and H) Confocal section of ∼180 nm through the middle of the nucleus. Bar, 5 μm.
Figure 3.

Depletion of cellular B55 by pECE-B55as. (A) Hs68 were transiently transfected with the empty pECE plasmid (mock) or with the B55 antisense construct pECE-B55as (B55as). Thirty-six hours after transfection 50 μg of total lysate were electrophoretically separated and immunoblotted using Ab55recomb. Immunodecoration using Ab55473/448 revealed a similar disappearance of B55 in extracts of pECE-B55as-transfected cells. In parallel experiments an efficiency of 60% transfection was determined using a green fluorescent protein expression construct. (B and C) Cells were microinjected with the B55-antisense RNA–expressing construct pECE-B55as, mixed with mouse immunoglobulin G, cultured for a further 15 h, fixed, and stained for B55 using Ab55473/448 (B). The arrowheads indicate the microinjected cell as identified by costaining for the microinjection marker antibody (C). Note that this cell has been exclusively microinjected in the nucleus. Bar, 5 μm.
Lack of B55 Leads to Reorganization and Increased Phosphorylation of the Vimentin IF
Phase-contrast microscopy of cells deprived of B55 by antisense microinjection revealed the formation of dense cytoskeletal structures around the nuclei. Our results using OA (Figure 1) and the immunolocalization of B55 using Ab55recomb prompted us to analyze vimentin distribution in Hs68 cells lacking B55. Because B55-containing PP2A holoenzymes were reported to be involved during mitotic progression (Mayer-Jaekel et al., 1993), when both cellular morphology and vimentin distribution are highly modified (Chou et al., 1990), we used only cells in interphase. For this Hs68 fibroblasts were made quiescent by serum deprivation and microinjected 2–3 h after being refed, allowing analysis for 22–24 h before cells entered mitosis (Girard et al., 1992). Hs68 cells, deprived of B55 by antisense injection, showed a dramatic reorganization of vimentin distribution. Specifically, 6–9 h after microinjection the normally fine, well-extended vimentin network retracted from the cell edges and collapsed (Figure 4, A and B). The extent of this reorganization was dependent on the time after antisense B55 injection. Cells fixed 12–15 h after injection exhibited a complete contraction of the vimentin into dense filament bundles around the nucleus (Figure 4, C and D). Longer times (>20 h) resulted in complete rounding and subsequent detachment of B55-depleted cells. Similar effects on vimentin were also obtained after microinjection of affinity purfied anti-B55 antibody AB55473/488 (our unpublished results). Antisense depletion of B subunits other than B55 in Hs68 cells did not have such effects on vimentin (Turowski, Fernandez, and Lamb, manuscript in preparation). A representative example is shown in Figure 4, E and F.
Figure 4.

Vimentin disassembly and hyperphosphorylation after the functional knockout of B55. (A–D) Hs68 fibroblasts were microinjected with pECE-B55as and cultured for 6 h (A and B) or 12 h (C and D). Cells were then stained for vimentin (A and C). Arrowheads indicatemicroinjected cells as identified by costaining against the marker antibody (B and D). (E and F) Similarly, Hs68 fibroblasts were microinjected with a B56 antisense construct (manuscript in preparation) and subsequently stained for vimentin (E) and injection marker (F). Note that antisense B56 did not induce any changes in vimentin integrity. Bars, 5 μm. (G) Hs68 fibroblasts were grown on 1-mm2 glass chips and microinjected with the control plasmid pECE or the antisense B55-expressing construct pECE-B55as. Cells were subcultured for 6 h and then metabolically labeled with [32P]H3PO4 for 3 h. Subsequently, phosphoproteins were analyzed by two-dimensional gel electrophoresis as described in MATERIALS AND METHODS. Shown are portions of the autoradiograms containing vimentin (black arrowheads) and surrounding proteins only (white arrowheads indicate hsp 28). Shown are phosphoproteins of 230 cells injected with the control plasmid pECE (left panel) and 216 cells injected with the antisense B55 construct pECE-B55as (right panel). Autoradiographs were exposed for 4 d at −80°C and quantitated using a PhosphorImager.
To examine whether the dramatic reorganization of vimentin was associated with hyperphosphorylation, the phosphorylation state of vimentin in cells lacking B55 was analyzed. For this, Hs68 fibroblasts were grown to equal densities on small (1-mm2) glass coverslips, and all cells were counted and microinjected with either the control plasmid pECE or the B55 antisense construct pECE-B55as. Six hours after injection, cells were metabolically labeled with [32P]H3PO4 for 3 h, and the totality of the phosphoproteins was analyzed by two-dimensional gel electrophoresis and autoradiography. As shown in Figure 4G, vimentin from exactly 216 cells, all injected with antisense B55 (right panel), was phosphorylated to a greater extent than that from exactly 230 control-injected cells (left panel). Because the exact number of cells used in each experiment is known, a direct comparison of the phosphoproteins can be made. This was further illustrated by the fact that the phosphorylation of no other major phosphoprotein (such as hsp 28) changed between experiments using similar cell numbers. Significantly, we found from two such metabolic labeling experiments that the amount of labeled vimentin increased threefold.
Therefore, depletion of B55 induced an increased phosphorylation of vimentin, which accompanied its disassembly. In addition, reorganization of cytoskeletal structures after B55 antisense injection was restricted to the vimentin IF: none of the injected cells exhibited alterations of the microtubule network (Figure 5, A and B) or the F-actin (Figure 5, C and D).
Figure 5.

Microtubule and microfilament networks are unaffected in B55-depleted cells. Hs68 fibroblasts were microinjected with pECE-B55as and cultured for 12 h as in Figure 4. (A and B) Cells were then stained for tubulin (A). Arrowheads indicate microinjected cell as identified by costaining against the marker antibody (B). (C and D) Costaining for F-actin (C) and vimentin (D), respectively. Here the corresponding guinea pig marker antibody (revealed with amino-methylcoumarin) is not shown but indicated by arrowheads. In each case ∼50 cells were microinjected, and no effect on microtubule or actin distribution was observed. Bars, 5 μm.
Vimentin Dephosphorylation is Enhanced by B55-Type Regulator
These results suggested that vimentin was directly modulated by a B55-containing PP2A heterotrimer. We next tested whether purified B55-containing heterotrimer dephosphorylated vimentin. Human B55 protein was expressed in Sf9 cells infected with a recombinant baculovirus carrying B55α cDNA and purified to >80% homogeneity (Figure 6A). Added to homogeneous, dimeric PP2A (PP2A2), it induced an approximately sixfold increase of the phosphatase activity toward p34cdc2-phosphorylated histone H1, whereas phosphorylase phosphatase activity remained unchanged (Figure 6B). This is consistent with analyses of cell free extracts of Drosophila mutants lacking functional B55 (Mayer-Jaekel et al., 1994) and suggests that our recombinant B55 preparation was fully functional. To study the effect of B55 on vimentin dephosphorylation, vimentin, potentially hyperphosphorylated on PP2A-specific sites, was purified from OA-treated Hs68 cells by Triton X-100 and KCl extraction (see MATERIALS AND METHODS and Figure 7). It was further electrophoretically separated from all contaminants, electrotransfered onto nitrocellulose, and renatured by Tris-buffered saline–guanidinium cycles. Subsequently, strips containing equal amounts of phosphovimentin were incubated with various protein phosphatase mixtures, and the released phosphate was quantitated (Figure 6C). In this assay, addition of recombinant B55 stimulated the basal activity of dimeric PP2A (PP2A2) toward vimentin by a factor of ∼5. PP1 or PP2A mixtures containing 10 nM OA did not induce significant dephosphorylation of vimentin. To further confirm that trimeric, B55-containing PP2A was also competent to dephosphorylate native vimentin, we developed an in situ assay. Cells were grown on coverslips, treated with OA, and then extracted with Triton X-100 and KCl to wash out most soluble proteins (also see Figure 7). The resulting cytoskeletal lattices, highly enriched in vimentin, were incubated with PP2A mixtures, and at various times thereafter vimentin distribution was determined by immunofluorescence. Incubation with buffer or PP2A2 alone had no effect on vimentin distribution: vimentin was disassembled and strongly bundled in ∼80% of the cells (Figure 6, D and E). In contrast, after incubation with PP2A2 and an excess of recombinant B55, a significant number of the vimentin lattices had reorganized, as characterized by a fine, filamentous staining pattern (Figure 6, F and G). Quantitative analysis (∼600 vimentin lattices were assessed for each time point and incubation condition) revealed that only the incubation with PP2A2 plus recombinant B55 resulted in efficient reassembly of vimentin into filaments (Figure 6H). Most importantly, this reorganization was attributable to dephosphorylation by PP2A, because it could be abolished by adding 10 nM OA to the assay. Moreover, immunodepleted B55 preparations did not enhance vimentin reorganization by PP2A2. Likewise, semipurified, active preparations of B56β (McCright et al., 1996; Zolnierowicz et al., 1996), a different PP2A regulator present in Hs68 fibroblasts (Turowski, Fernandez, and Lamb, manuscript in preparation), was ineffective (our unpublished results). The extent of vimentin reorganization induced by B55/PP2A increased linearly with time (Figure 6I). Thus B55 enhanced the specificity of PP2A for vimentin both in vitro and in situ.
Figure 6.

Enhanced dephosphorylation of vimentin in the presence of recombinant B55. (A) Twenty microliters of recombinant B55 prepared as described in MATERIALS AND METHODS were electrophoretically separated, and proteins were visualized using Coomassie brilliant blue. The molecular masses of marker proteins in kilodaltons (lane M) are indicated at the left. The major full-length and minor proteolytically degraded forms of B55, as identified by immunoblotting, are indicated with arrowheads. (B) The effect of B55 on the activity of purified dimeric PP2A2 toward phosphorylase and (p34cdc2 phosphorylated) histone H1 was assayed. Before the assay, PP2A2 (at 1 nM final concentration) was preincubated with (black bars) or without (hatched bars) ∼10 nM (final) recombinant B55 on ice. (C) Nitrocellulose strips, containing hyperphosphorylated [32P]vimentin were prepared as described in MATERIALS AND METHODS. Similarly to the phosphatase assays shown in B, strips containing equal amounts of [32P]vimentin (∼30,000 cpm and 5 μg of protein) were incubated with either 1 nM dimeric PP2A2 preincubated with (open squares) or without (closed squares) 10 nM recombinant B55 or with 1 nM catalytic subunit of PP1 (triangles). Phosphate release was determined and plotted against the assay time. (D–I) Hs68 fibroblasts grown on coverslips were treated with 1 μM OA for 1 h. Soluble proteins were extracted using Triton X-100 and 1.5 M KCl, and the remaining vimentin lattices were extensively washed in phosphatase assay buffer. Subsequently coverslips were overlaid with 30 μl of protein phosphatase assay buffer containing no phosphatase (D), 1 nM PP2A2 (E), or 1 nM PP2A2 and ∼10 nM recombinant B55 (F and G) and incubated at 37°C for 45 min. Reactions were stopped by formalin fixation and further stained for vimentin distribution. Bar, 5 μm. (H) Quantitative representation of the experiment shown in D–G; vimentin distribution resembling D or E was counted as bundled, whereas that resembling F or G or Figure 1B, upper left (unaffected vimentin), was counted as (re)assembled. On average, 600 stained vimentin lattices of each coverslip (i.e., assay point) were counted. In all assays ∼20% of the vimentin lattices had normal appearance and had not been affected by the initial OA treatment. (I) Time course of vimentin dephosphorylation by 1 nM PP2A2 and 10 nM B55. Vimentin reassembly was determined as in H, and for each time point the corresponding background (no PPase) was subtracted.
Figure 7.

Association of PP2A with vimentin in Hs68 fibroblasts. (A and B) Vimentin was isolated from Hs68 fibroblasts by Triton X-100 and 0.6 M KCl extraction as described in MATERIALS AND METHODS. Material corresponding to one-third of a 100-mm dish of Hs68 was electrophoretically analyzed; equal proportions of the soluble (lanes S; S/10, 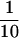 of S) and insoluble, vimentin-enriched (lanes VE) fractions were electrophoretically separated. Subsequently, proteins were stained with Coomassie brilliant blue (A). Arrows indicate the positions of vimentin (v) and actin (a). The migration of molecular mass markers is indicated at the left. Alternatively, gels were immunoblotted (B) for the presence of PP2Ac and PR65 using AbC302/309 (upper panel) and Ab65177/196 (middle panel). Approximately 5 ng of rabbit skeletal muscle PP2Ac and ∼10 ng of recombinant PR65 were included as a positive control (lanes C), and their positions are marked with arrowheads.Because of a high background staining in the region at ∼50 kDa, B55 could not be detected in an unequivocal way using our polyclonal Ab55. Vimentin was isolated from Hs68, transiently transfected to express HA-tagged B55. These preparations were immunoblotted using the monoclonal anti-HA (12CA5) (lower panel). nt, vimentin from nontransfected cells; t, from transfected cells. The arrowheads indicate the migration of HA-B55 as determined by anti-HA immunoprecipitates, run on the same gel and detected with Ab55recomb. (C) Hs68 fibroblasts were grown on glass coverslips and, before formalin fixation, extracted with 0.01% Triton X-100. Subsequently, cells were stained for PP2Ac, PR65, or B55 using AbCrecomb, Ab65177/196, or Ab55recomb, respectively, and costained for vimentin. The staining for PP2A subunits was revealed using FITC-conjugated reagents (upper panels), whereas vimentin was detected using Texas Red (middle panels). The lower panels show double exposures of the respective PP2A and vimentin staining (see MATERIALS AND METHODS). Bar, 5 μm.
of S) and insoluble, vimentin-enriched (lanes VE) fractions were electrophoretically separated. Subsequently, proteins were stained with Coomassie brilliant blue (A). Arrows indicate the positions of vimentin (v) and actin (a). The migration of molecular mass markers is indicated at the left. Alternatively, gels were immunoblotted (B) for the presence of PP2Ac and PR65 using AbC302/309 (upper panel) and Ab65177/196 (middle panel). Approximately 5 ng of rabbit skeletal muscle PP2Ac and ∼10 ng of recombinant PR65 were included as a positive control (lanes C), and their positions are marked with arrowheads.Because of a high background staining in the region at ∼50 kDa, B55 could not be detected in an unequivocal way using our polyclonal Ab55. Vimentin was isolated from Hs68, transiently transfected to express HA-tagged B55. These preparations were immunoblotted using the monoclonal anti-HA (12CA5) (lower panel). nt, vimentin from nontransfected cells; t, from transfected cells. The arrowheads indicate the migration of HA-B55 as determined by anti-HA immunoprecipitates, run on the same gel and detected with Ab55recomb. (C) Hs68 fibroblasts were grown on glass coverslips and, before formalin fixation, extracted with 0.01% Triton X-100. Subsequently, cells were stained for PP2Ac, PR65, or B55 using AbCrecomb, Ab65177/196, or Ab55recomb, respectively, and costained for vimentin. The staining for PP2A subunits was revealed using FITC-conjugated reagents (upper panels), whereas vimentin was detected using Texas Red (middle panels). The lower panels show double exposures of the respective PP2A and vimentin staining (see MATERIALS AND METHODS). Bar, 5 μm.
PP2A Association with Vimentin
At this point our results suggested that PP2A might dephosphorylate vimentin directly. PP2A is the major protein phosphatase acting on neurofilaments and found associated with this type of IF (Saito et al., 1995). To corroborate an association of PP2A with vimentin, fractions highly enriched in IFs were prepared from Hs68 fibroblasts (Figure 7A) and immunoblotted for the presence of PP2A subunits. Both PP2Ac and the PR65 regulatory subunit were detected in these preparations (Figure 7B). Immunoblotting of vimentin preparations with Ab55recomb or Ab55473/448 gave rise to several bands at ∼55 kDa, the nature of which could not be determined unequivocally. To circumvent this problem, vimentin was isolated from Hs68 transfected with pCMV-HA-B55α and expressing HA-tagged B55. Specifically, HA-tagged B55 was detected associated with vimentin from transfected cells (Figure 7B, lower panel). This association was considered specific, because all three PP2A subunits were still present at an ionic strength up to 1.5 M KCl. No PP1 was detected in these vimentin fractions by immunoblotting. Comparison of the levels of PP2A present on vimentin with that of the soluble pool (Figure 7B) showed that the majority of PP2A was soluble, and only a small subpopulation was associated with vimentin. An exact estimation of how much PP2A is associated with vimentin appeared difficult, because PP2Ac, PR65, and B55 were found in each of the high-salt washes of the vimentin pellet, implying that the actual amount detected might be inferior to that associated in living cells.
We have previously shown using a number of different antibodies that the majority of PP2Ac and PR65 is distributed throughout both the cytoplasmic and the nuclear compartments (Turowski et al., 1995). In turn B55 is essentially cytoplasmic (Figure 2). When any of the antibodies against PP2Ac, PR65, or B55 were used in immunfluorescence analyses of cells, mildly extracted with Triton X100 before fixation, most of the diffuse staining in the cytoplasm and the nucleus disappeared. A comparatively weak, filamentous staining pattern was left, which coincided with the corresponding vimentin staining (Figure 7C, upper and middle panels). This was especially striking in double exposures of the two staining patterns (Figure 7C, lower panels). This colocalization of a fraction of the three PP2A subunits, PP2Ac, PR65, and B55, with vimentin after detergent solubilization of the bulk of these proteins further reinforced that PP2A not only dephosphorylated vimentin in Hs68 fibroblasts but also associated with this substrate.
Targeting by B55
In this respect it was significant to find that an antibody, directed against an internal region of the catalytic subunit of PP2A (AbC169/182; Favre et al., 1994), also showed a colocalization of PP2Ac with vimentin. This antibody was specific for PP2Ac in immunoblots of Hs68 (Figure 8A) and in immunoprecipitates from whole-cell lysates (Favre et al., 1994). Immunofluorescence analysis of Hs68 cells stained with AbC169/182 showed a fine, filamentous staining (Figure 8B, left panel), which seemed to colocalize with vimentin IFs. Indeed, when superimposed with the corresponding vimentin staining, most of this PP2Ac staining coincided with vimentin (Figure 8B, right panel). Only a few, mostly peripheral areas showed no costaining of PP2Ac and vimentin (Figure 8B, residual, red staining, arrowheads; also see DISCUSSION). It is noteworthy that AbC169/182 gave rise to a signal colocalizing with vimentin without detergent pre-extraction (compared with the staining shown in Figure 7C), making it a valuable tool to follow vimentin-associated PP2Ac. With the use of this antibody we analyzed whether the PP2Ac–vimentin interaction was mediated by B55. For this, the distribution of vimentin-associated PP2Ac was determined in cells depleted of B55 by antisense injection. In these cells the AbC169/182 staining followed the bundling of vimentin, and this staining diminished at later time points of antisense injection (Figure 8C). Moreover, the disappearance of AbC169/182 staining was specific and restricted to vimentin-associated PP2Ac, because immunofluorescence analysis of the totality of PP2Ac (as revealed by AbCrecomb; Turowski et al., 1995) showed no discernible changes (Figure 8D). The mean, total fluorescence of B55 antisense-injected cells stained for vimentin-associated PP2Ac (using AbC169/182), total PP2Ac (using AbCrecomb), or vimentin was determined and compared with that of control-injected cells (Figure 8E). Whereas the amount of total PP2Ac remained unchanged, that of vimentin-associated PP2Ac decreased nearly fivefold in antisense B55 injected cells. This was not due to the disappearance or solubilization of vimentin, because its overall staining did not change to the same extent. Thus B55 depletion resulted in a decrease of vimentin-associated PP2Ac, indicating that B55 is required to target PP2A to vimentin.
Figure 8.

Targeting by B55. (A) Immunoblot analysis of PP2A catalytic subunit from total cell lysates of Hs68 fibroblasts using AbC169/182 were performed similarly to those in Figure 1. Lane 1, ∼20 μg of cell extract; lane C, 5 ng of rabbit skeletal muscle PP2Ac. Positions of molecular mass markers are indicated at the left. The bands revealed are competed by the antigenic peptide. (B) Hs68 fibroblasts were grown on glass coverslips and formalin fixed. Subsequently, they were costained for the catalytic subunit using AbC169/182, revealed by FITC (left panel), and vimentin using mAb V9, revealed by Texas Red, as described for Figure 7B. To visualize colocalization of PP2Ac with vimentin, double exposures of both signal were recorded (right panel). Note that PP2Ac colocalized with most of the vimentin (yellow), but some regions of vimentin were devoid of PP2Ac staining (red, arrowheads). (C and D) Hs68 fibroblasts, grown on glass coverslips, were microinjected with the antisense B55 RNA–expressing construct pECE-B55as. After 12 h, cells were fixed using formalin. In C cells were stained for vimentin-associated PP2Ac using AbC169/182 (left panel) and for the microinjection marker (right panel). In D cells were stained stained for total PP2Ac using AbCrecomb (left panel) and for the microinjection marker (right panel). Arrowheads indicate injected cells. (E) In similar experiments as in C and D, cells were microinjected with the control plasmid pECE or with pECE-B55as. Subsequently they were stained for the totality of cellular PP2Ac using AbCrecomb (Turowski et al., 1995), vimentin-associated PP2Ac using AbC169/182, or vimentin using V9. Subsequently, the fluorescence of each microinjected cell was quantitated using ImgCalc. The histogram shows average fluorescence as determined from at least 30 injected cells for each staining. Values found for control-injected cells were arbitrarily set to 100%. Bars, 5 μm.
DISCUSSION
Observations implicating PP2A in a variety of frequently antagonistic pathways were gathered in studies using inhibitors such as OA, mutants in yeast or Drosophila, or in vitro analyses of defined holoenzymes (for review, see Shenolikar, 1994; Wera and Hemmings, 1995). The question has remained, however, of which holoenzyme acts at a precise moment to dephosphorylated a given substrate in a living cell. We have used a combination of biochemical fractionation, activity assays, pharmacological inhibition, and antisense knockout of regulatory PP2A subunits in living cells to identify specific heterotrimeric PP2A holoenzymes in a given cellular process. In the current report we show that heterotrimeric PP2A containing the B55 regulatory subunit (PP2A1) dephosphorylates vimentin of living fibroblasts throughout interphase.
B55 Is a Cytoplasmic Regulator of PP2A
PP2A is a cytoplasmic and nuclear regulator in mammalian cells (Turowski et al., 1995). In the present report we show, using immunoblots of fractionated cell extracts and indirect immunofluorescence, that the B55-type regulatory subunit is a predominantly cytoplasmic protein in Hs68 fibroblasts. Its cytoplasmic localization is consistent with the absence of a nuclear localization signal in the primary structure of B55 isoforms (Mayer et al., 1991; Zolnierowicz et al., 1994). Moreover, all processes in which B55-containing PP2A holoenzymes have been implicated to date appear to be cytoplasmic (Healy et al., 1991; Mayer-Jaekel et al., 1993, 1994; Uemura et al., 1993; Lee et al., 1994; Pitcher et al., 1995; Hansra et al., 1996; Sontag et al., 1996). In contrast, members of the B56 and B72 families of regulatory subunits might mediate nuclear functions of PP2A (Hendrix et al., 1993a; McCright et al., 1996).
Our data also showed that a fraction of cytoplasmic B55 is associated with the IFs in mammalian fibroblasts. Mild detergent extraction of cells before fixation solubilized >90% of the cellular B55 as well as the PP2Ac and PR65, indicating that the majority of B55-containing heterotrimer (PP2A1) is present in a highly soluble form. The remaining material was found colocalized with vimentin IF. Using a similar pre-extraction technique, our laboratory has previously visualized the association of PP1 with actin microfilaments (Fernandez et al., 1990). Immunochemical analysis of purified vimentin, isolated under highly stringent conditions, further corroborated a physical association of PP2A1 with vimentin.
B55 has been reported to be associated with the microtubule network (Sontag et al., 1995). Under no circumstances have we observed an association of PP2A subunits with microtubules, not even when using cell fixatives favoring microtubule stabilization (our unpublished results). The filamentous staining pattern of AbCs, Ab65s, and Ab55s described above colocalized with the vimentin IF but not with microtubules. Also, during the cofractionation of PP2A and vimentin, tubulin and microtubule-associated proteins were solubilized because of the high-salt extraction performed on ice. Moreover, depleting fibroblasts of B55 by antisense microinjection did not cause any alterations in the microtubule network, suggesting no obvious function for B55 in maintaining interphase microtubule integrity. However, given previous reports, it remains feasible that PP2A is implicated in the reversible phosphorylation of microtubule-associated proteins in neuronal cells (Merrick et al., 1996; Sontag et al., 1996).
PP2A Targeted by B55 Dephosphorylates Vimentin In Vivo
Hyperphosphorylation of vimentin and other IF proteins leads to their disassembly, leading to bundling in vivo (for review, see Inagaki et al., 1996). In this report we have identified PP2A as a major phosphatase involved in vimentin modulation. First, the inhibition of type 2A protein phosphatases by OA in Hs68 fibroblasts induced marked hyperphosphorylation and reorganization of vimentin, in accordance with other reports (Yatsunami et al., 1991; Eriksson et al., 1992; Lee et al., 1992; Lai et al., 1993). OA is a very potent inhibitor of PP2A but appears to be equally potent in inhibiting other PP2A-related phosphatases (Brewis et al., 1993; Chen et al., 1994). However, the abundance of phosphovimentin would rather imply PP2A as major vimentin protein phosphatase and discount a role of less-abundant PP2A-related phosphatases. In support we could obtain similar disassembly of the vimentin by functional knockout of B55, which is a regulatory subunit of PP2A and not of any other protein phosphatases. The functional knockout of B55 also caused increased vimentin phosphorylation. This increase was less pronounced than that after OA treatment, and this difference probably reflects the direct and instant inhibition of PP2A by OA and the indirect and progressive action of B55 antisense knockout. The latter process is less efficient, because it depends on the balance between B55 and B55-antisense transcription rate, B55 protein stability, as well as B55 turnover in PP2A1 holoenzymes. Interestingly, in this respect, microinjection of affinity-purified anti-B55 antibodies could produce similar changes in vimentin organization within shorter incubation times (our unpublished results). The exclusive role B55 plays in vimentin dephosphorylation was further underlined by the observation that B56 antisense expression (the other class of PP2A regulatory subunits expressed in Hs68 fibroblasts; Turowski, Fernandez, and Lamb, manuscript in preparation) had no effect on vimentin organization.
To study vimentin dephosphorylation by defined phosphatase preparations, we examined phosphate release from highly phosphorylated vimentin isolated from OA-treated cells. Because such hyperphosphorylated vimentin was completely insoluble, we were constrained to measure dephosphorylation on phosphovimentin adsorbed to nitrocellulose strips. In these assays recombinant B55 increased the specificity of dimeric PP2A by approximately fivefold, an increase similar to that measured with histone H1, which is a highly specific substrate for B55-containing PP2A (Ferrigno et al., 1993). Further confirmation that PP2A could act on native vimentin was derived from an in situ assay, which is based on the assumption that vimentin phosphorylation induces its disassembly and bundling, whereas dephosphorylation would induce reassembly into filaments (Inagaki et al., 1996). In these in situ assays the presence of B55 was an absolute prerequisite for PP2A to induce vimentin filament reassembly. Other lines of evidence indicated that vimentin is a direct substrate for B55/PP2A. A fraction of PP2Ac, PR65, and B55 tightly associated with vimentin could be purified from mammalian fibroblasts. Furthermore, using antibody AbC169/182 we could measure a disappearance of vimentin-associated PP2Ac after B55 depletion. These results imply that B-type regulators do not only modulate the specificity of PP2A for its substrate but also play a role in targeting PP2A to a substrate. This is a concept that is already largely accepted and proven for PP1 (Hubbard and Cohen, 1993) but remained to be shown for PP2A. In conclusion, our observations, 1) hyperphosphorylation of vimentin by either OA treatment or B55 depletion, 2) physical association of PP2Ac, PR65, and B55 with vimentin, 3) enhanced in vitro dephosphorylation and in situ reassembly of vimentin in the presence of B55, and 4) the loss of PP2Ac from vimentin in B55-depleted cells, indicate an unequivocal role of B55 in directly targeting PP2A core dimer to vimentin.
Role of IF Dephosphorylation by PP2A
A number of observations suggest that PP2A might be a general IF phosphatase. We have previously reported that cytokeratins 8 and 18 become hyperphosphorylated when type 2A but not type 1 enzymes are inhibited (Favre et al., 1997). PP2A was also identified as a neurofilament phosphatase and, significantly, associates with this type of IF (Saito et al., 1995). Whether PP2A is always targeted to IF by B55 remains to be established.
What is the function of vimentin dephosphorylation by PP2A during interphase? IF proteins are major phosphoproteins within living cells and substrates for a variety of protein kinases such as cAMP- and cGMP-dependent protein, calcium- and calmodulin-dependent protein kinase II, PKC, and p34cdc2 (for review, see Inagaki et al., 1996). Whereas strong hyperphosphorylation and vimentin reorganization are observed when the intracellular levels of cAMP-dependent protein kinases were significantly increased by direct microinjection (Lamb et al., 1989), activation of endogenous cAMP-dependent protein kinase or PKC using cAMP analogues or phorbol esters generates only modest changes in IF phosphorylation (DePhilip and Kierszenbaum, 1982; Coca-Prados, 1985; Chou and Omary, 1991; Deery, 1993). The discrepancy between these results suggests the presence of excess IF phosphatase activity, explaining why inhibition of PP2A (Yatsunami et al., 1991; Eriksson et al., 1992; Lee et al., 1992; Lai et al., 1993) or interfering with its targeting induces such a marked hyperphosphorylation of vimentin (this report). From these data we favor the hypothesis that the main role of PP2A is to prevent inopportune reorganization of vimentin in response to interphase kinase activation, as also previously suggested by Lai et al. (1993). In this respect it was interesting to note that the colocalization of PP2A subunits did not extend to the totality of the vimentin network (also see Figures 7C and 8B). Indeed some regions of vimentin appeared completely devoid of PP2A. This might indicate that PP2A not only serves a “Sisyphus” role in turning phosphate over on vimentin but is also actively involved in regulating interphase IF dynamics. IF reorganization is necessary for morphological changes during differentiation, migration, and spreading (Fuchs and Weber, 1994). Our observations that B55/PP2A modulates vimentin phosphorylation might be a key in understanding how mutant alleles of B55 can lead to developmental defects in Drosophila (Uemura et al., 1993; Shiomi et al., 1994). Such a role in modulating IF dynamics may also explain the effects of mutant B55 alleles on mitotic transit in yeast and Drosophila (Healy et al., 1991; Mayer-Jaekel et al., 1993; Wang and Burke, 1997), in which IF proteins, such as vimentin and lamins, undergo significant, phosphorylation-induced reorganization.
ACKNOWLEDGMENTS
We thank Laure Lelievre for contributing to the initial studies on the localization of B55, D. Casanova and C. Lecardeur for immunizing rabbits, Celine Franckhauser for technical assistance, Dr. S. Courtneidge for the gift of antibody AbC302/309, Dr. M. Bollen for PP1 catalytic subunit, Dr. J. Goris for purified PP2A heterodimer, and Dr. J.-C. Labbé for anticyclin B antibodies and starfish extracts. The advice of Prof. J.-C. Cavadore for preparing peptide–protein conjugates was greatly appreciated. Prof. J. Demaille is thanked for continuous support. We also thank our colleagues Marie Vandromme and Anne Debant for critical reading of the manuscript. This work was supported in part by a postdoctoral fellowship from the Roche Research Foundation (to P.T., contract 95-161), by Human Frontiers Science Organization grant RG 533/96, by Biomed 2, by Association pour la Recherche contre le Cancer contract 1344 (to A.F.), and by La Ligue contre le Cancer (to N.J.C.L.).
Footnotes
OA and related toxins have been shown to exhibit a similar inhibitory potential toward certain PP1- and PP2A-related minor phosphatases than toward PP1 and PP2A themselves (Brewis et al., 1993; Chen et al., 1994). In this report we use the terms “type 1” for PP1 and PP1-related and “type 2A” for PP2A and PP2A-related enzymes. The terms PP1 and PP2A were reserved for the respective holoenzyme family.
REFERENCES
- Agostinis P, Goris J, Waelkens E, Pinna LA, Marchiori F, Merlevede W. Dephosphorylation of phosphoproteins and synthetic phosphopeptides. Study of the specificity of the polycation-stimulated and Mg-ATP-dependent phosphorylase phosphatases. J Biol Chem. 1987;262:1060–1064. [PubMed] [Google Scholar]
- Alessi DR, Andjelkovic M, Caudwell B, Cron P, Morrice N, Cohen P, Hemmings BA. Mechanism of activation of protein kinase B by insulin and IGF-1. EMBO J. 1996;15:6541–6551. [PMC free article] [PubMed] [Google Scholar]
- Andjelkovic N, Zolnierowicz S, Van-Hoof C, Goris J, Hemmings BA. The catalytic subunit of protein phosphatase 2A associates with the translation termination factor eRF1. EMBO J. 1996;15:7156–7167. [PMC free article] [PubMed] [Google Scholar]
- Blattler DP, Garner F, Van Slyke K, Bradley A. Quantitative electrophoresis in polyacrylamide gels of 2–40% J Chromatogr. 1972;64:147–155. [Google Scholar]
- Blose SH, Meltzer DI, Feramisco JR. 10-nm filaments are induced to collapse in living cells microinjected with monoclonal and polyclonal antibodies against tubulin. J Cell Biol. 1984;98:847–858. doi: 10.1083/jcb.98.3.847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brewis ND, Street AJ, Prescott AR, Cohen PT. PPX, a novel protein serine/threonine phosphatase localized to centrosomes. EMBO J. 1993;12:987–996. doi: 10.1002/j.1460-2075.1993.tb05739.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cegielska A, Shaffer S, Derua R, Goris J, Virshup DM. Different oligomeric forms of protein phosphatase 2A activate and inhibit SV40 DNA replication. Mol Cell Biol. 1994;14:4616–4623. doi: 10.1128/mcb.14.7.4616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen CA, Okayama H. Calcium phosphate-mediated gene transfer: a highly efficient transfection system for stably transforming cells with plasmid DNA. Biotechniques. 1988;6:632–638. [PubMed] [Google Scholar]
- Chen MX, McPartlin AE, Brown L, Chen YH, Barker HM, Cohen PT. A novel protein human serine/threonine phosphatase, which possesses four tetratricopeptide repeat motifs and localizes to the nucleus. EMBO J. 1994;15:4278–4290. doi: 10.1002/j.1460-2075.1994.tb06748.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou C-F, Omary MB. Phorbol acetate enhances the phosphorylation of cytokeratins 8 an 18 in human colonic epithelial cells. FEBS Lett. 1991;282:200–204. doi: 10.1016/0014-5793(91)80477-k. [DOI] [PubMed] [Google Scholar]
- Chou Y-H, Bischoff JR, Beach D, Goldman RD. Intermediate filament reorganization during mitosis is mediated by p34cdc2 phosphorylation of vimentin. Cell. 1990;62:1063–1071. doi: 10.1016/0092-8674(90)90384-q. [DOI] [PubMed] [Google Scholar]
- Coca-Prados M. Regulation of protein phosphorylation of the intermediate-sized filament vimentin in cilary epithelium of the mammalian eye. J Biol Chem. 1985;260:10332–10338. [PubMed] [Google Scholar]
- Csortos C, Zolnierowicz S, Bako E, Durbin SD, DePaoli-Roach AA. High complexity in the expression of the B′ subunit of protein phosphatase 2A0. Evidence for the existence of at least seven novel isoforms. J Biol Chem. 1996;271:2578–2588. doi: 10.1074/jbc.271.5.2578. [DOI] [PubMed] [Google Scholar]
- Deery WJ. Role of phosphorylation in keratin and vimentin filament integrity in cultured thyroid epithelial cells. Cell Motil Cytoskeleton. 1993;26:325–339. doi: 10.1002/cm.970260407. [DOI] [PubMed] [Google Scholar]
- DePhilip RM, Kierszenbaum AL. Hormonal regulation of protein synthesis, secretion and phosphorylation in cultured rat Sertoli cells. Proc Natl Acad Sci USA. 1982;79:6551–6555. doi: 10.1073/pnas.79.21.6551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis L, Clauser E, Morgan DO, Edery M, Roth R, Rutter WJ. Replacement of insulin receptor tyrosine residues 1162 and 1163 compromises insulin-stimulated kinase activity and uptake of 2-deoxyglucose. Cell. 1986;45:721–732. doi: 10.1016/0092-8674(86)90786-5. [DOI] [PubMed] [Google Scholar]
- Eriksson JE, Brautigan DL, Vallee R, Olmsted J, Fujiki H, Goldman RD. Cytoskeletal integrity in interphase cells requires protein phosphatase activity. Proc Natl Acad Sci USA. 1992;89:11093–11097. doi: 10.1073/pnas.89.22.11093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Favre B, Turowski P, Hemmings BA. Differential inhibition and posttranslational modification of protein phosphatase 1 and 2A in MCF7 cells treated with calyculin-A, okadaic acid, and tautomycin. J Biol Chem. 1997;272:13856–13863. doi: 10.1074/jbc.272.21.13856. [DOI] [PubMed] [Google Scholar]
- Favre B, Zolnierowicz S, Turowski P, Hemmings BA. The catalytic subunit of protein phosphatase 2A is carboxyl-methylated in vivo. J Biol Chem. 1994;269:16311–16317. [PubMed] [Google Scholar]
- Fernandez A, Brautigan DL, Lamb NJC. Protein phosphatase type 1 in mammalian cell mitosis: chromosomal localization and involvement in mitotic exit. J Cell Biol. 1992;116:1421–1430. doi: 10.1083/jcb.116.6.1421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez A, Brautigan DL, Mumby M, Lamb NJC. Protein phosphatase type-1, not type-2A, modulates actin microfilament integrity and myosin light chain phosphorylation in living nonmuscle cells. J Cell Biol. 1990;111:103–112. doi: 10.1083/jcb.111.1.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrigno P, Langan TA, Cohen P. Protein phosphatase 2A1 is the major enzyme in vertebrate cell extracts that dephosphorylates several physiological substrates for cyclin-dependent protein kinases. Mol Biol Cell. 1993;4:669–677. doi: 10.1091/mbc.4.7.669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franke WW, Weber K, Osborn M, Schmid E, Freudenstein C. Antibody to prekeratin: decoration of tonofilament-like arrays in various cells of epithelial character. Exp Cell Res. 1978;116:429–445. doi: 10.1016/0014-4827(78)90466-4. [DOI] [PubMed] [Google Scholar]
- Fuchs E, Weber K. Intermediate filaments: structure, dynamics, function and disease. Annu Rev Biochem. 1994;63:345–382. doi: 10.1146/annurev.bi.63.070194.002021. [DOI] [PubMed] [Google Scholar]
- Girard F, Strausfeld U, Cavadore J-C, Russel P, Fernandez A, Lamb NJC. cdc25 is a nuclear protein expressed constitutively throughout the cell cycle in nontransformed mammalian cells. J Cell Biol. 1992;118:785–794. doi: 10.1083/jcb.118.4.785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansra G, Bornancin F, Whelan R, Hemmings BA, Parker PJ. 12-O-Tetradecanoylphorbol-13-acetate-induced dephosphorylation of protein kinase Cα correlates with the presence of a membrane-associated protein phosphatase 2A heterotrimer. J Biol Chem. 1996;271:32785–32788. doi: 10.1074/jbc.271.51.32785. [DOI] [PubMed] [Google Scholar]
- Haystead TA, Sim AT, Carling D, Honnor RC, Tsukitani Y, Cohen P, Hardie DG. Effects of the tumour promoter okadaic acid on intracellular protein phosphorylation and metabolism. Nature. 1989;337:78–81. doi: 10.1038/337078a0. [DOI] [PubMed] [Google Scholar]
- Healy AM, Zolnierowicz S, Stapleton AE, Goebl M, DePaoli-Roach AA, Pringle JR. CDC55, a Saccharomyces cerevisiae gene involved in cellular morphogenesis: identification, characterization, and homology to the B subunit of mammalian type 2A protein phosphatase. Mol Cell Biol. 1991;11:5767–5780. doi: 10.1128/mcb.11.11.5767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hendrix P, Mayer-Jaekel RE, Cron P, Goris J, Hofsteenge J, Merlevede W, Hemmings BA. Structure and expression of a 72-kDa regulatory subunit of protein phosphatase 2A. J Biol Chem. 1993a;268:15267–15276. [PubMed] [Google Scholar]
- Hendrix P, Turowski P, Mayer-Jaekel RE, Goris J, Hofsteenge J, Merlevede W, Hemmings BA. Analysis of subunit isoforms in protein phosphatase 2A holoenzymes from rabbit and Xenopus. J Biol Chem. 1993b;268:7330–7337. [PubMed] [Google Scholar]
- Hubbard MJ, Cohen P. On target with a new mechanism for the regulation of protein phosphorylation. Trends Biochem Sci. 1993;18:172–177. doi: 10.1016/0968-0004(93)90109-z. [DOI] [PubMed] [Google Scholar]
- Inagaki M, Matsuoka Y, Tsujimura K, Ando S, Tokui T, Takahashi T, Inagaki N. Dynamic property of intermediate filaments: regulation by phosphorylation. Bioessays. 1996;18:481–487. [Google Scholar]
- Kamibayashi C, Estes R, Lickteig RL, Yang S-I, Craft C, Mumby MC. Comparison of heterotrimeric protein phosphatase 2A containing different B subunits. J Biol Chem. 1994;269:20139–20148. [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural protein during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Lai YK, Lee WC, Chen K-D. Vimentin serves as a phosphatase Sink During the apparent activation of protein kinases by okadaic acid in mammalian cells. J Cell Biochem. 1993;53:161–168. doi: 10.1002/jcb.240530209. [DOI] [PubMed] [Google Scholar]
- Lamb NJC, Fernandez A. Microinjection of antibodies into mammalian cells. Methods Enzymol. 1997;283:72–83. doi: 10.1016/s0076-6879(97)83008-x. [DOI] [PubMed] [Google Scholar]
- Lamb NJC, Fernandez A, Feramisco JR, Welch WJ. Modulation of vimentin containing intermediate filament distribution and phosphorylation in living fibroblasts by the cAMP-dependent protein kinase. J Cell Biol. 1989;108:2409–2422. doi: 10.1083/jcb.108.6.2409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee TH, Turck C, Kirschner MW. Inhibition of cdc2 activation by INH/PP2A. Mol Biol Cell. 1994;5:323–338. doi: 10.1091/mbc.5.3.323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee WC, Yu JS, Yang SD, Lai YK. Reversible hyperphosphorylation and reorganization of vimentin intermediate filaments by okadaic acid in 9L rat brain tumor cells. J Cell Biochem. 1992;49:378–393. doi: 10.1002/jcb.240490408. [DOI] [PubMed] [Google Scholar]
- Mayer RE, Hendrix P, Cron P, Matthies R, Stone SR, Goris J, Merlevede W, Hofsteenge J, Hemmings BA. Structure of the 55-kDa regulatory subunit of protein phosphatase 2A: evidence for a neuronal-specific isoform. Biochemistry. 1991;30:3589–3597. doi: 10.1021/bi00229a001. [DOI] [PubMed] [Google Scholar]
- Mayer-Jaekel RE, Ohkura H, Ferrigno P, Andjelkovic N, Shiomi K, Uemura T, Glover DM, Hemmings BA. Drosophila mutants in the 55 kDa regulatory subunit of protein phosphatase 2A show strongly reduced ability to dephosphorylate substrates of p34cdc2. J Cell Sci. 1994;107:2609–2616. doi: 10.1242/jcs.107.9.2609. [DOI] [PubMed] [Google Scholar]
- Mayer-Jaekel RE, Ohkura H, Gomes R, Sunkel CE, Baumgartner S, Hemmings BA, Glover DM. The 55 kd regulatory subunit of Drosophila protein phosphatase 2A is required for anaphase. Cell. 1993;72:621–633. doi: 10.1016/0092-8674(93)90080-a. [DOI] [PubMed] [Google Scholar]
- Mayer-Jaekel RE, Hemmings BA. Protein phosphatase 2A—a “menage a trois.”. Trends Cell Biol. 1994;4:287–291. doi: 10.1016/0962-8924(94)90219-4. [DOI] [PubMed] [Google Scholar]
- McCright B, Rivers AM, Audlin S, Virshup DM. The B56 family of protein phosphatase 2A (PP2A) regulatory subunits encodes differentiation-induced phosphoproteins that target PP2A to both nucleus and cytoplasm. J Biol Chem. 1996;271:22081–22089. doi: 10.1074/jbc.271.36.22081. [DOI] [PubMed] [Google Scholar]
- McCright B, Virshup DM. Identification of a new family of protein phosphatase 2A regulatory subunits. J Biol Chem. 1995;270:26123–26128. doi: 10.1074/jbc.270.44.26123. [DOI] [PubMed] [Google Scholar]
- Merrick SE, Demoise DC, Lee VM. Site-specific dephosphorylation of tau protein at Ser202/Thr205 in response to microtubule depolymerization in cultured human neurons involves protein phosphatase 2A. J Biol Chem. 1996;271:5589–5594. doi: 10.1074/jbc.271.10.5589. [DOI] [PubMed] [Google Scholar]
- Michelson S, Turowski P, Picard L, Goris J, Landini MP, Topilko A, Hemmings BA, Bessia C, Garcia A, Virelizier JL. Human cytomegalovirus carries serine/threonine protein phosphatase 1 and a host-cell derived PP2A. J Virol. 1996;70:1415–1423. doi: 10.1128/jvi.70.3.1415-1423.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Farrell PH. High resolution two-dimensional electrophoresis of proteins. J Biol Chem. 1975;250:4007–4021. [PMC free article] [PubMed] [Google Scholar]
- Pallas DC, Weller W, Jaspers S, Miller TB, Lane WS, Roberts TM. The third subunit of protein phosphatase 2A (PP2A), a 55-kilodalton protein which is apparently substituted for by T antigens in complexes with the 36- and 63-kilodalton PP2A subunits, bears little resemblance to T antigens. J Virol. 1992;66:886–893. doi: 10.1128/jvi.66.2.886-893.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitcher JA, Payne ES, Csortos C, DePaoli-Roach AA, Lefkowitz RJ. The G-protein-coupled receptor phosphatase: a protein phosphatase type 2A with a distinct subcellular distribution and substrate specificity. Proc Natl Acad Sci USA. 1995;92:8343–8347. doi: 10.1073/pnas.92.18.8343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saito T, Shima H, Osawa Y, Nagao M, Hemmings BA, Kishimoto T, Hisanaga S. Neurofilament-associated protein phosphatase 2A: its possible role in preserving neurofilaments in filamentous states. Biochemistry. 1995;34:7376–7384. doi: 10.1021/bi00022a010. [DOI] [PubMed] [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Shenolikar S. Protein serine/threonine phosphatases—new avenues for cell regulation. Annu Rev Cell Biol. 1994;10:55–86. doi: 10.1146/annurev.cb.10.110194.000415. [DOI] [PubMed] [Google Scholar]
- Shiomi K, Takeichi M, Nishida Y, Nishi Y, Uemura T. Alternative cell fate choice induced by low-level expression of a regulator of protein phosphatase 2A in the Drosophila peripheral nervous system. Development. 1994;120:1591–1599. doi: 10.1242/dev.120.6.1591. [DOI] [PubMed] [Google Scholar]
- Sontag E, Nunbhakdi-Craig V, Bloom GS, Mumby MC. A novel pool of protein phosphatase 2A is associated with microtubules and is regulated during the cell cycle. J Cell Biol. 1995;128:1131–1144. doi: 10.1083/jcb.128.6.1131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sontag E, Nunbhakdi-Craig V, Lee G, Bloom GS, Mumby MC. Regulation of the phosphorylation state and microtubule-binding activity of Tau by protein phosphatase 2A. Neuron. 1996;17:1201–1207. doi: 10.1016/s0896-6273(00)80250-0. [DOI] [PubMed] [Google Scholar]
- Stone SR, Hofsteenge J, Hemmings BA. Molecular cloning of cDNAs encoding two isoforms of the catalytic subunit of protein phosphatase 2A. Biochemistry. 1987;26:7215–7220. doi: 10.1021/bi00397a003. [DOI] [PubMed] [Google Scholar]
- Summers MD, Smith GE. A manual of methods for baculovirus vectors and insect cell culture procedures. Tex Agric Exp Stn Bull. 1987;1555:1–57. [Google Scholar]
- Tanabe O, Nagase T, Murakami T, Nozaki H, Usui H, Nishito Y, Hayashi H, Kagamiyama H, Takeda M. Molecular cloning of a 74-kDa regulatory subunit (B“ or delta) of human protein phosphatase 2A. FEBS Lett. 1996;379:107–111. doi: 10.1016/0014-5793(95)01500-0. [DOI] [PubMed] [Google Scholar]
- Tehrani MA, Mumby MC, Kamibayashi C. Identification of a novel protein phosphatase 2A regulatory subunit highly expressed in muscle. J Biol Chem. 1996;271:5164–5170. doi: 10.1074/jbc.271.9.5164. [DOI] [PubMed] [Google Scholar]
- Turowski P, Favre B, Campbell KS, Lamb NJC, Hemmings BA. Modulation of the enzymatic properties of protein phosphatase 2A catalytic subunit by the recombinant 65-kDa regulatory subunit PR65α. Eur J Biochem. 1997;248:200–208. doi: 10.1111/j.1432-1033.1997.t01-1-00200.x. [DOI] [PubMed] [Google Scholar]
- Turowski P, Fernandez A, Favre B, Lamb NJC, Hemmings BA. Differential methylation and altered conformation of cytoplasmic and nuclear forms of protein phosphatase 2A during cell cycle progression. J Cell Biol. 1995;129:397–410. doi: 10.1083/jcb.129.2.397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turowski P, Lamb NJC. Microinjection and immunological methods in the analysis of type 1 and 2A protein phosphatases from mammalian cells. Methods Mol Biol. 1998;93:117–136. doi: 10.1385/0-89603-468-2:117. [DOI] [PubMed] [Google Scholar]
- Uemura T, Shiomi K, Togashi S, Takeichi M. Mutation of twins encoding a regulator of protein phosphatase 2A leads to pattern duplication in Drosophila imaginal disks. Genes & Dev. 1993;7:429–440. doi: 10.1101/gad.7.3.429. [DOI] [PubMed] [Google Scholar]
- Wang Y, Burke DJ. Cdc55p, the B-type regulatory subunit of protein phosphatase 2A, has multiple functions in mitosis and is required for the kinetochore/spindle checkpoint in Saccharomyces cerevisiae. Mol Cell Biol. 1997;17:620–626. doi: 10.1128/mcb.17.2.620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wera S, Hemmings BA. Serine/threonine protein phosphatases. Biochem J. 1995;311:17–29. doi: 10.1042/bj3110017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yatsunami J, Fujiki H, Suganuma M, Yoshizawa S, Eriksson JE, Olson MO, Goldman RD. Vimentin is hyperphosphorylated in primary human fibroblasts treated with okadaic acid. Biochem Biophys Res Commun. 1991;177:1165–1170. doi: 10.1016/0006-291x(91)90662-q. [DOI] [PubMed] [Google Scholar]
- Zhao Y, Boguslawski G, Zitomer RS, DePaoli-Roach AA. Saccharomyces cerevisiae homologs of the mammalian B and B′ subunits of protein phosphatase 2A direct the enzyme to distinct cellular functions. J Biol Chem. 1997;272:8256–8262. doi: 10.1074/jbc.272.13.8256. [DOI] [PubMed] [Google Scholar]
- Zolnierowicz S, Csortos C, Bondor J, Verin A, Mumby MC, DePaoli-Roach AA. Diversity in the regulatory B-subunits of protein phosphatase 2A: identification of a novel isoform highly expressed in brain. Biochemistry. 1994;33:11858–11867. doi: 10.1021/bi00205a023. [DOI] [PubMed] [Google Scholar]
- Zolnierowicz S, Van Hoof C, Andjelkovic N, Cron P, Stevens I, Merlevede W, Goris J, Hemmings BA. The variable subunit associated with protein phosphatase 2A0 defines a novel multimember family of regulatory subunits. Biochem J. 1996;317:187–194. doi: 10.1042/bj3170187. [DOI] [PMC free article] [PubMed] [Google Scholar]