

Abstract
The first chemical inhibitor of the 19S Regulatory Particle (RP) of the proteasome is described. The molecule was identified by screening a library of nucleoside-capped peptoids for binding to the yeast 26S proteasome in a crude extract. The hit was resynthesized and shown to block 19S RP-mediated protein unfolding in vitro and proteasome-mediated turnover of p27 in HeLa cells.
The 26S proteasome is an approximately 2.5 MDa multicatalytic protease complex that is responsible for most non-lysosomal protein degradation in eukaryotic cells. It is composed of a barrel-like catalytic 20S core particle (CP) that consists of four stacked hetero-heptameric rings capped on each side by a 19S regulatory particle (RP) (Fig. 1)1, which contains a putative heterohexameric ring of ATPases (Rpt1-6) as well as several other proteins. The RP binds polyubiquitylated proteins, unfolds them, and feeds the polypeptide chain into the interior of the barrel where the proteins are degraded into small peptides2. Inhibition of the 20S CP proteolytic activity has emerged as an attractive pharmacological target for cancer and inflammatory diseases3,4 as well as useful as mechanistic probes of proteasome function in a variety of biological processes3.
Figure 1.
A cartoon of the 26S proteasome. The black line represents a substrate protein in the process of being degraded. The 19S RP (pink and blue shapes) that would cap the other end of the 20S CP (grey) has been omitted for clarity.
Recently, a number of pioneering studies have revealed that stimulating proteasome-mediated proteolysis is but one of several activities of the Rpt proteins. It is now clear that these ATPases play non-proteolytic roles in RNA polymerase II transcription, DNA repair, structural modification of chromatin and other nuclear processes5-10. Therefore, pharmacological inhibitors of the proteasomal ATPases (Rpts) would be of great interest. Here we report the isolation of the first compound of this type from a library of nucleoside-capped peptoids. We show that this peptoid derivative inhibits the protein unfolding activity of the Rpt proteins in vitro and inhibits the proteasome activity in living cells.
A “one bead one compound” peptoid library was constructed by split and pool synthesis (see Supp. Mat. Fig. S1)11. Each peptoid molecule was capped with a purine analogue (Fig. 2A) in hope of biasing the library towards targeting one of the ATPases. The theoretical diversity of the library was 85 (32,768 compounds). Approximately 100,000 beads, representing about 3-fold coverage of the library, were used in the screen for compounds that bind to the proteasome. The screen employed a whole cell extract12 prepared from a yeast strain that expressed a FLAG-tagged β4 proteasome sub-unit (one of the 20S proteins). The beads were exposed to the extract, washed thoroughly (see Supplementary Material), probed with anti-FLAG monoclonal antibody and then washed again. Finally, putative proteasome-binding peptoids were visualized by addition of a secondary anti-IgG antibody labeled with red-emitting quantum-dots13.
Fig. 2.
Isolation and characterization of RIP-1. (A) General structure of the purine-capped peptoid library. (B) A fluorescence micrograph of a bead scored as a “hit” (red halo) in the midst of large number of beads scored as negatives (blue). (C) The structure of the hit, RIP-1, as determined by Edman sequencing and C2, a control peptoid not selected for binding to the proteasome.
Three beads evinced an obvious red halo, consistent with retention of the quantum dot-conjugated secondary antibody (Fig. 2B). Note that the library had been pre-screened to eliminate peptoids that bound directly to the antibodies or the quantum dot before exposing it to the yeast extract. Edman degradation revealed that all three hits were identical, indicating the presence of a single proteasome-binding peptoid in the library with sufficient affinity and specificity to register under these relatively demanding conditions. We called this compound RIP-1 (Regulatory Particle Inhibitor Peptoid-1; Fig. 2C).
To examine the effect of the peptoid-purine conjugate RIP-1 on the unfolding activity of the 19S regulatory particle, we employed an assay recently developed in our laboratory10 that involves exposing Gal4-VP16 protein bound to an immobilized DNA containing five Gal4 binding sites. The base of the 19S RP binds to the VP16 activation domain7 and thus engages the fusion protein as a substrate for unfolding, resulting in the ATP-dependent disruption of the protein-DNA complex (Fig. 3A)10. An excess of soluble DNA is included in the assay to prevent reassociation of Gal4-VP16 with the immobilized DNA and the amount of protein remaining on the bead-bound DNA thus reflects the rate of ATPase-mediated “stripping” of the activator. A control lacking the proteasome allows correction for the intrinsic rate of dissociation of the complex. The proteolytic activity of the 20S CP is not involved in this process and, in fact, purified 19S RP lacking the 20S core is quite active10. As seen in Fig. 3B, RIP-1 inhibited the proteasome-mediated stripping of the Gal4-VP16 protein from the immobilized DNA with an IC50 of approximately 3 μM. A control peptoid, called C2 (Fig. 1), did not affect the chaperonin activity (see Supp. Mat. Fig. S2). The purine cap alone lacking a peptoid also had no effect. The peptoid lacking the purine cap did show some activity, though it was a weaker inhibitor than RIP-1 (not shown). A more complete structure activity analysis of this system will be presented in a subsequent full paper.
Fig. 3.
RIP-1 inhibits in vitro chaperonin activity. (A) A cartoon describing the proteasome-mediated stripping of the Gal4-VP16 protein from the immobilized DNA. (B) Inhibition of proteasome-mediated destabilization of the activator-DNA complex by RIP-1. The graph shows the average amount and standard error of the mean of GST-Gal4-VP16 remaining on DNA for three replicates. Increasing activator remaining on the DNA indicates decreased chaperonin activity of the ATPases.
Peptoids are generally more cell permeable than peptides14 and thus we hoped that RIP-1 might have activity against the proteasome in cellular assays. To examine this point, the effect of RIP-1 on steady state levels of p27, which is turned over via the ubiquitin-proteasome pathway15, was examined. HeLa cells were incubated with the indicated concentrations of RIP-1 (Fig. 4) or, as controls, the known proteasome inhibitor MG132, compound C2 or DMSO carrier alone (Fig. 4). RIP-1 induced increased accumulation of p27 in a dose-responsive fashion. The effect of 100 μM RIP-1 was similar to that of 1 μM MG132. Neither the control compound C2 or the DMSO carrier affected p27 levels significantly.
Fig. 4.
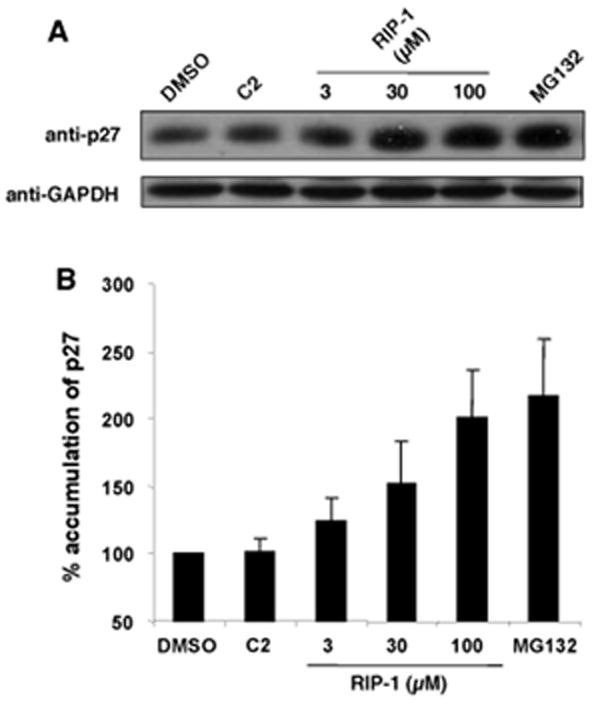
RIP-1 induces the accumulation of p27 in HeLa cells. (A) HeLa cells were treated with C2 (100 μM), the indicated concentrations of RIP-1, or MG132 (1 μM) for 6h. Lysates were subjected to immunoblot analysis with anti-p27 antibody. The results are representative of three independent experiments. (B) Immunoblot intensity normalized to the DMSO control for Fig. 4A. Error bars represent standard deviation from three independent experiments.
These data show clearly that RIP-1 inhibits the protein unfolding activity of the 19S RP with low micromolar potency (Fig. 3) and 26S-mediated proteolysis of p27 in living cells with an IC50 of about 30-50 μM (Fig. 4). Since the protein unfolding activity is mediated by the 19S RP and does not require the activity of the 20S CP, these results argue strongly that RIP-1 is indeed an inhibitor of the 19S RP, not the proteolytic activity of the 20S CP. As will be reported elsewhere (H.-S.L., et al., in preparation), this model is supported by the finding that RIP-1 does not inhibit peptidolysis catalyzed by the 20S RP in the absence of the 19S RP. Finally, as will also be reported elsewhere data from cross-linking experiments are consistent with the 19S RP being the target of RIP-1 and specifically implicate one of the ATPases, Sug2/Rpt4 as the direct receptor of the peptoid.
Supplementary Material
Detailed experimental procedures and supplementary figures are available free of charge at http://pubs.acs.org.
Acknowledgments
This work was supported by a contract from the National Heart, Lung, and Blood Institute (NO1-HV-28185) and a grant from the Welch Foundation (I-1299). H.S.L. was partially supported by a Korea Research Foundation Grant (M01-2004-000-20395-0).
References
- 1.Baumeister W, Walz J, Zuhl F, Seemuller E. Cell. 1998;92:367–380. doi: 10.1016/s0092-8674(00)80929-0. [DOI] [PubMed] [Google Scholar]
- 2.DeMartino G, Slaughter CA. J Biol Chem. 1999;274:22123–22126. doi: 10.1074/jbc.274.32.22123. [DOI] [PubMed] [Google Scholar]
- 3.Kisselev AF, Goldberg A. Chem & Biol. 2001;8:739–758. doi: 10.1016/s1074-5521(01)00056-4. [DOI] [PubMed] [Google Scholar]
- 4.Adams J. Cancer Cell. 2004;5:417–421. doi: 10.1016/s1535-6108(04)00120-5. [DOI] [PubMed] [Google Scholar]
- 5.Russell SJ, Reed SH, Huang W, Friedberg EC, Johnston SA. Mol Cell. 1999;5:687–696. doi: 10.1016/s1097-2765(01)80001-0. [DOI] [PubMed] [Google Scholar]
- 6.Ferdous A, Gonzalez F, Sun L, Kodadek T, Johnston SA. Mol Cell. 2001;7:981–991. doi: 10.1016/s1097-2765(01)00250-7. [DOI] [PubMed] [Google Scholar]
- 7.Gonzalez F, Delahodde A, Kodadek T, Johnston SA. Science. 2002;296:548–550. doi: 10.1126/science.1069490. [DOI] [PubMed] [Google Scholar]
- 8.Ezhkova E, Tansey WP. Mol Cell. 2004;13:435–442. doi: 10.1016/s1097-2765(04)00026-7. [DOI] [PubMed] [Google Scholar]
- 9.Collins GA, Tansey WP. Curr Op In Gentics & Dev. 2006;16:197–202. doi: 10.1016/j.gde.2006.02.009. [DOI] [PubMed] [Google Scholar]
- 10.Ferdous A, Sikder D, Gillette TG, Nalley K, Kodadek T, Johnston SA. Genes & Dev. 2007;21:112–123. doi: 10.1101/gad.1493207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Alluri PG, Reddy MM, Bacchawat-Sikder K, Olivos HJ, Kodadek T. J Amer Chem Soc. 2003;125:13995–14004. doi: 10.1021/ja036417x. [DOI] [PubMed] [Google Scholar]
- 12.Bradner JE, McPherson OM, Mazischek R, Barnes-Seeman D, Shen JP, Dhaliwal J, Stevenson KE, Duffner JL, Park SB, Neuberg DS, Nghiem P, Schreiber SL, Kohler AN. Chem & Biol. 2006;13:493–504. doi: 10.1016/j.chembiol.2006.03.004. [DOI] [PubMed] [Google Scholar]
- 13.Olivos HJ, Baccawat-Sikder K, Kodadek T. ChemBiochem. 2003;4:1242–1245. doi: 10.1002/cbic.200300712. [DOI] [PubMed] [Google Scholar]
- 14.Kwon YU, Kodadek T. J Amer Chem Soc. 2007;129:1508–1509. doi: 10.1021/ja0668623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Drexler HC. Cell Cycle. 2003;2(5):438–441. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Detailed experimental procedures and supplementary figures are available free of charge at http://pubs.acs.org.