

Abstract
Cell penetrating peptides and proteins (CPPs) are important tools for the delivery of impermeable molecules into living mammalian cells. To enable these cells to internalize proteins fused to common oligohistidine affinity tags, we synthesized an artificial cell surface receptor comprising an N-alkyl derivative of 3β-cholesterylamine linked to the metal chelator nitrilotriacetic acid (NTA). This synthetic receptor inserts into cellular plasma membranes, projects NTA headgroups from the cell surface, and rapidly cycles between the plasma membrane and intracellular endosomes. Jurkat lymphocytes treated with the synthetic receptor (10 micromolar) for one hour displayed ~ 8,400,000 NTA groups on the cell surface. Subsequent addition of the green fluorescent protein AcGFP fused to hexahistidine or decahistidine peptides (3 micromolar) and Ni(OAc)2 (100 micromolar) enhanced the endocytosis of AcGFP by 150-fold (hexahistidine fusion protein) or 600-fold (decahistidine fusion protein) within four hours at 37 °C. No adverse effects on cellular proliferation or morphology were observed under these conditions. By enabling common oligohistidine affinity tags to function as cell penetrating peptides, this metal-chelating cell surface receptor provides a useful tool for studies of cellular biology.
Cell penetrating peptides and proteins (CPPs) have emerged as important new tools for the delivery of impermeable molecules into living mammalian cells. These delivery systems have been constructed from basic segments of HIV Tat,1 oligoguanidine-containing peptoids,2 β-peptides,3 and other oligocationic motifs.4 The mechanism of cellular penetration by CPPs is primarily thought to involve endocytosis,5 particularly with larger cargo,6 but varies depending on the delivery system and the cell type.7
Oligohistidine peptides are often fused to proteins to provide affinity tags (“His tags”) that facilitate protein purification. To enable these peptides to function as CPPs, we synthesized an artificial cell surface receptor (1) comprising the plasma membrane anchor N-alkyl-3β-cholesterylamine linked to the metal chelator nitrilotriacetic acid (NTA). N-Alkyl-3β-cholesterylamine derivatives can function as prosthetic molecules active on the surface of living mammalian cells because this steroid can insert into cellular plasma membranes, project linked headgroups from the cell surface, and rapidly cycle between the plasma membrane and intracellular endosomes, similar to many naturally-occurring cell surface receptors.8,9 The NTA motif binds tightly to nickel, cobalt, copper, and zinc dications, and immobilized metal chelate chromatography (IMAC) with NTA-linked supports is widely used to purify proteins fused to His tags.10 Other lipids11,12 and polymers13 linked to NTA headgroups have also been reported.
Receptor 1 was synthesized from 3β-cholesterylamine (2)9 as shown in Scheme 1. Alkylation of 2 with ethyl 5-bromovalerate, followed by Boc protection to afford 3, provided an improved route to protected N-alkyl derivatives of the membrane anchor. Addition of a linker shown to increase the population of these compounds on the cell surface,9 acylation with a protected NTA derivative, and final deprotection afforded 1 in 47% overall yield.
Scheme 1.
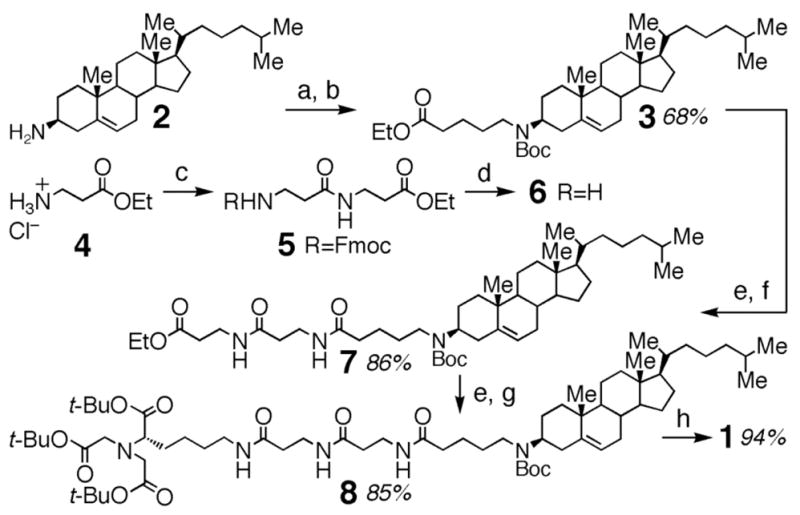
Reagents and conditions: (a) ethyl 5-bromovalerate, K2CO3, DMF, 60 °C; (b) (Boc)2O, DIEA, CH2Cl2; (c) N-Fmoc-β-Ala, EDC, HOBt, DIEA, CH2Cl2; (d) piperidine, DMF; (e) LiOH·H2O, MeOH/THF/H2O (3:2:1); (f) 6, EDC, HOBt, CH2Cl2; (g) Nα, Nα-bis[(tert-butoxycarbonyl)methyl]-L-Lys tert-butyl ester, EDC, HOBt, CH2Cl2; (h) TFA, CH2Cl2.
As representative His-tagged proteins, we overexpressed in E. coli the monomeric green fluorescent protein AcGFP fused to C-terminal (His)6 and (His)10 peptides and purified these proteins by IMAC. The (His)10 peptide binds Ni-NTA complexes ~ 6-fold more tightly than (His)6. For example, (His)6 and (His)10 peptides fused to 5HT3R serotonin receptors exhibit affinities for Ni-NTA derivatives of Kd = 1.05 μM and Kd = 166 nM, respectively.14
To qualitatively examine the ability of receptor 1 to mediate the cellular uptake of His-tagged proteins, human Jurkat T-lymphocytes were treated with 1 (10 μM) for one hour to load this compound into the outer leaflet of the cellular plasma membrane. These cells were washed and subsequently treated with a solution of AcGFP(His)10 (3.2 μM) and Ni(OAc)2 (100 μM) for an additional four hours. As shown in Figure 2 (panel A), examination of these cells by confocal laser scanning microscopy revealed fluorescent protein both on cell surface and in defined intracellular compartments. Washing the cells with disodium NTA (400 μM) as a chelator removed the bound protein from the cell surface, facilitating examination of the intracellular distribution (Figure 2, panel B). Delivery of AcGFP(His)6 under these conditions provided similar results, but the cells were 4-fold less fluorescent (Figure 3, Panel D). The fate of internalized AcGFP(His)10 was examined by coadministration with DiI-loaded low density lipoprotein (LDL), a protein internalized by receptor-mediated endocytosis, as a red fluorescent marker of endosomes and lysosomes.15 Substantial colocalization of red and green fluorophores was observed under these conditions (supporting information). This delivery to endosomes and lysosomes represents a common destination of proteins fused to CPPs.16
Figure 2.
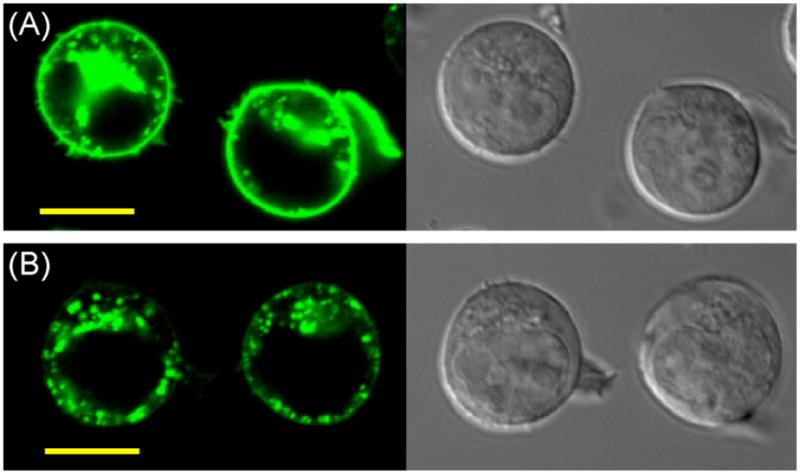
Confocal laser scanning (left) and differential interference contrast (right) micrographs of living Jurkat lymphocytes. Cellular plasma membranes were preloaded with receptor 1 (10 μM) for 1 h at 37 °C, cells were washed with fresh media, and media containing AcGFP-His10 (3.2 μM) and Ni(OAc)2 (100 μM) was added for an additional 4 h at 37 °C. Cells shown in Panel A were imaged immediately after this treatment. Cells shown in Panel B were washed with NTA (400 μM, 30 min) prior to microscopy to remove non-internalized protein from the cell surface. Scale bar = 10 μm.
Figure 3.

Analysis of cellular uptake of His-tagged AcGFP proteins by flow cytometry. Each bar represents the average fluorescence of 10,000 cells. Unless otherwise noted, cellular plasma membranes of Jurkat lymphocytes were preloaded with receptor 1 (10 μM) for 1 h at 37 °C, cells were washed with fresh media, and His-tagged AcGFP (3.2 μM)/metal diacetate solution (100 μM) was added for 4 h at 37 °C. Prior to analysis, cells were washed with NTA (400 μM, 30 min) in PBS (pH 7.4) to remove bound surface protein. Panel A: Dose dependence of receptor-mediated uptake. Premix conditions: a solution containing receptor 1, Ni(OAc)2, and AcGFP(His)10 was added to cells for 4 h. Panel B: Dependence on [Ni(OAc)2]. Panel C: Omission control experiments. Panel D; Uptake of (His)6 and (His)10 fusion proteins promoted by different metal diacetates.
Quantitative analysis by flow cytometry of synthetic receptor-mediated uptake is shown in Figure 3. Intracellular delivery of AcGFP(His)10 > 600-fold above basal levels of endocytosis was achieved by preloading the cellular plasma membrane with receptor 1 (10 μM) for 1 h followed by addition of the protein (3.2 μM) and Ni(OAc)2 (100 μM). Higher receptor concentrations, addition of disodium NTA as a competitor, or premixing the receptor with the protein and metal resulted in lower levels of uptake (panel A). Preloading the plasma membrane may facilitate uptake due to the high surface density of immobilized metal chelate.17,18 Significantly higher or lower concentrations of Ni(OAc)2 were less effective mediators of uptake (panel B), and omission control experiments confirmed the importance of all three components (panel C). Comparison of diacetates of Ni, Co, Cu, and Zn revealed that Ni(OAc)2 was the most effective metal for delivery of both (His)6 and (His)10 fusion proteins.
Consistent with the mechanism of synthetic receptor-mediated endocytosis,9 uptake of His-tagged AcGFP proteins was blocked by cooling cells to 4 °C (data provided in the supporting information). These conditions enabled quantification of the number of synthetic receptors on the cell surface. Jurkat lymphocytes were treated with 1 (10 μM) for 1 h at 37 °C, the cells were washed, chilled to 4 °C, and excess AcGFP(His)10 (19 μM) and Ni(OAc)2 (100 μM) was added to saturate the surface-bound NTA groups. Comparison of these cells with fluorescent bead standards by flow cytometry revealed an average of ~ 8,400,000 synthetic receptors per cell surface. Previous studies of related compounds suggest that a similar population of synthetic receptors resides in intracellular endosomes.9
Heavy metals such as Ni2+ exhibit dose-dependent toxicity to biological systems. To examine the toxicity of this metal acetate and protein delivery, cellular viability was examined by flow cytometry. In these experiments, Jurkat lymphocytes were treated with receptor 1 (10 μM, 1 h), followed by AcGFP(His)10 (3.2 μM), and Ni(OAc)2 (100 μM, 4 h) to enhance protein uptake by 600-fold. Cells were washed with disodium NTA to remove cell surface protein and were cultured for an additional 48 h. Cellular viability was determined by light scattering and staining the nuclei of membrane-compromised dead cells with the fluorophore propidium iodide. Under these conditions, 97% of the cells were viable, compared with 99% of untreated controls, and no effects on cellular morphology were observed. Other studies have similarly reported low toxicity of nickel salts at ≤ 160 μM in cell culture.19 By enabling common oligohistidine affinity tags to function as cell penetrating peptides, synthetic receptor 1 provides a potentially versatile new probe of cellular biology.
Supplementary Material
Supporting figures (S1–S3), compound characterization data and experimental procedures. This material is available at http://pubs.acs.org.
Figure 1.
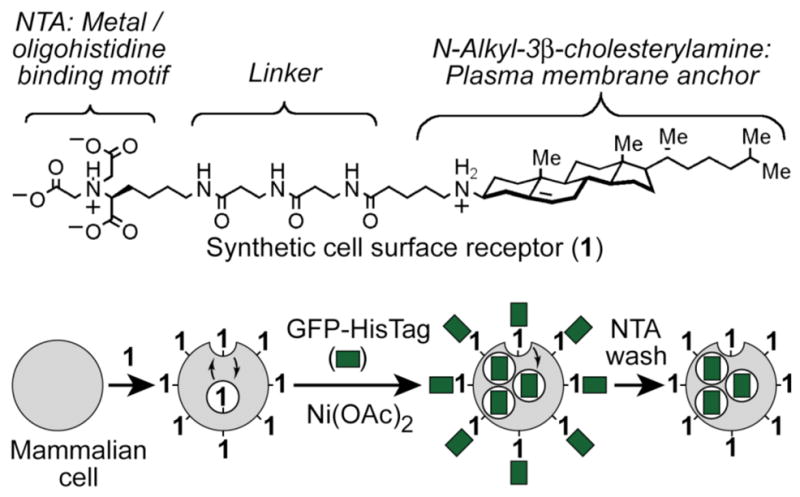
Top: Structure of the synthetic cell surface receptor (1) that binds metals and His tags. Bottom: Strategy for delivery of His-tagged proteins into mammalian cells by synthetic receptor-mediated endocytosis.
Acknowledgments
We thank the NIH (R01-CA83831) for financial support.
References
- 1.Brooks H, Lebleu B, Vives E. Adv Drug Deliv Rev. 2005;57:559–577. doi: 10.1016/j.addr.2004.12.001. [DOI] [PubMed] [Google Scholar]
- 2.Wender PA, Mitchell DJ, Pattabiraman K, Pelkey ET, Steinman L, Rothbard JB. Proc Natl Acad Sci USA. 2000;97:13003–13008. doi: 10.1073/pnas.97.24.13003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Umezawa N, Gelman MA, Haigis MC, Raines RT, Gellman SH. J Am Chem Soc. 2002;124:368–369. doi: 10.1021/ja017283v. [DOI] [PubMed] [Google Scholar]
- 4.Jiang H, O’Neil EJ, Divittorio KM, Smith BD. Org Lett. 2005;7:3013–3016. doi: 10.1021/ol0510421. [DOI] [PubMed] [Google Scholar]
- 5.Fuchs SM, Raines RT. Biochemistry. 2004;43:2438–2444. doi: 10.1021/bi035933x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Maiolo JR, Ferrer M, Ottinger EA. Biochim Biophys Acta. 2005;1712:161–172. doi: 10.1016/j.bbamem.2005.04.010. [DOI] [PubMed] [Google Scholar]
- 7.Rothbard JB, Jessop TC, Wender PA. Adv Drug Deliv Rev. 2005;57:495–504. doi: 10.1016/j.addr.2004.10.003. [DOI] [PubMed] [Google Scholar]
- 8.Peterson BR. Org Biomol Chem. 2005;3:3607–3612. doi: 10.1039/b509866a. [DOI] [PubMed] [Google Scholar]
- 9.Boonyarattanakalin S, Martin SE, Dykstra SA, Peterson BR. J Am Chem Soc. 2004;126:16379–16386. doi: 10.1021/ja046663o. [DOI] [PubMed] [Google Scholar]
- 10.Hochuli E, Dobeli H, Schacher A. J Chromatogr. 1987;411:177–184. doi: 10.1016/s0021-9673(00)93969-4. [DOI] [PubMed] [Google Scholar]
- 11.Hodges HB, Zhou M, Haldar S, Anderson JL, Thompson DH, Hrycyna CA. Bioconjug Chem. 2005;16:490–493. doi: 10.1021/bc050027d. [DOI] [PubMed] [Google Scholar]
- 12.van Broekhoven CL, Parish CR, Vassiliou G, Altin JG. J Immunol. 2000;164:2433–2443. doi: 10.4049/jimmunol.164.5.2433. [DOI] [PubMed] [Google Scholar]
- 13.Griffith BR, Allen BL, Rapraeger AC, Kiessling LL. J Am Chem Soc. 2004;126:1608–1609. doi: 10.1021/ja037646m. [DOI] [PubMed] [Google Scholar]
- 14.Guignet EG, Hovius R, Vogel H. Nat Biotechnol. 2004;22:440–444. doi: 10.1038/nbt954. [DOI] [PubMed] [Google Scholar]
- 15.Ghosh RN, Gelman DL, Maxfield FR. J Cell Sci. 1994;107:2177–2189. doi: 10.1242/jcs.107.8.2177. [DOI] [PubMed] [Google Scholar]
- 16.Young JC, Moarefi I, Hartl FU. J Cell Biol. 2001;154:267–273. doi: 10.1083/jcb.200104079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nieba L, Nieba-Axmann SE, Persson A, Hamalainen M, Edebratt F, Hansson A, Lidholm J, Magnusson K, Karlsson AF, Pluckthun A. Anal Biochem. 1997;252:217–228. doi: 10.1006/abio.1997.2326. [DOI] [PubMed] [Google Scholar]
- 18.Sigal GB, Bamdad C, Barberis A, Strominger J, Whitesides GM. Anal Chem. 1996;68:490–497. doi: 10.1021/ac9504023. [DOI] [PubMed] [Google Scholar]
- 19.Shiao YH, Lee SH, Kasprzak KS. Carcinogenesis. 1998;19:1203–1207. doi: 10.1093/carcin/19.7.1203. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting figures (S1–S3), compound characterization data and experimental procedures. This material is available at http://pubs.acs.org.