

Abstract
Insulin-like growth factor 1 (IGF-1) enhances thymopoiesis but given the broad distribution of IGF-1 receptors (IGF-1Rs), its mechanism of action has remained unclear. To identify points of thymic regulation by IGF-1, we examined its effects on T-cell precursors, thymocytes, and thymic epithelial cells (TECs) in normal and genetically altered mice. In thymus-intact but not thymectomized mice, IGF-1 administration increased peripheral naive and recent thymic emigrant (RTE) populations, demonstrating its effect on T-cell production, not peripheral expansion. IGF-1 administration increased bone marrow LSK (lineage−, Sca-1+, c-kit+) precursor proliferation and peripheral LSK populations, increased thymocyte populations in a sequential wave of expansion, and proportionately expanded TEC subpopulations and enhanced their chemokine expression. To separate IGF-1's effects on thymocytes and TECs, we generated mice lacking IGF-1R on thymocytes and T cells. Thymocyte and RTE numbers were decreased in these mice, but IGF-1 treatment produced comparable thymocyte numbers to similarly treated wild-type mice. We additionally separated thymic- from LSK-specific effects by demonstrating that IGF-1 increased thymocyte numbers despite impaired early thymic progenitor (ETP) importation in PSGL-1KO mice. These results indicate the critical point thymic function regulation by IGF-1 involves TEC expansion regulating thymocyte precursor entry and facilitating thymocyte development.
Introduction
Thymic function after lymphodepletion as seen in hematopoietic stem cell transplantation (HSCT), HIV infection, and aging is critical for maintaining a broad repertoire of T-cell responses. Impaired thymic function results in not only increased susceptibility to opportunistic infections but also increased risk of tumor relapse due to impaired tumor surveillance.1 Moreover, given the critical role of the thymus in negative selection and the generation of regulatory T cells, impaired thymic function may be a significant contributory factor in the development of graft-versus-host disease and autoimmunity.2,3 Strategies to preserve and/or enhance thymic function therefore may overcome these important clinical barriers.
Thymic regulation can occur at multiple levels. The thymus requires continuous replenishment of bone marrow–derived progenitors to maintain T-cell production. Thereafter, T-cell development occurs in an ordered temporal-spatial sequence within distinct developmental niches defined by interactions between thymocytes and thymic epithelial cells (TECs). TECs define overall thymic function by modulating thymocyte survival, proliferation, trafficking, and positive and negative selection.4 Consequently, increasing the number of available T-cell precursors for importation into the thymus,5,6 expanding developmental niches within the thymus by expanding of TEC populations,7,8 and manipulating interactions between TECs and thymocytes to increase thymopoietic throughput have all been implicated as potential points of thymic function regulation. Nevertheless, their relative importance in the regulation of overall thymic regulation remains unknown. In this regard, the actions of thymic regulators, which presumably act by one or more of these mechanisms, provide insights into the biology of thymic regulation.
Insulin-like growth factor 1 (IGF-1) is one of a family of neuroendocrine factors having multiple anabolic functions including increased cell proliferation, inhibition of apoptosis, and cell differentiation that confer positive effects on organ growth and recovery from injury.9,10 The proliferative and developmental effects of growth hormone (GH) on multiple organ systems, including the thymus and the peripheral immune system,11,12 are primarily mediated through IGF-1.13,14 Moreover, IGF-1 is produced by a wide variety of cells in a paracrine/autocrine fashion independently of GH regulation.15,16
Immunologically, IGF-1 has been shown to promote hematopoiesis,12,17,18 prolong lymphocyte survival,19,20 and modulate T-cell signaling.21 In the thymus, IGF-1 has been implicated as a positive thymic regulator based on early observations that age-related declines in thymic function paralleled declines in plasma concentrations of IGF-1.22 IGF-1 receptor (IGF-1R) is expressed on thymocytes and peripheral T cells.23,24 In murine fetal thymic organ cultures, inhibition of IGF-1 by antibody blockade resulted in significant changes in total thymocyte numbers and subset composition.25 TECs also express IGF-1R, and addition of IGF-1 to human TEC lines facilitated the adhesion between thymocytes and TECs through modulation of cell adhesion molecule expression and the production by TECs of extracellular matrix proteins.26 In addition, IGF-1 has been shown to induce proliferation in cultured human TEC lines.27 These in vitro studies illustrating the positive effects of IGF-1 on T-cell development are further supported by in vivo observations where IGF-1 has been shown to enhance thymic reconstitution in murine HSCT.28,29
Because the cellular mechanisms by which IGF-1 affects thymic function in vivo have not been completely elucidated, we undertook a systematic approach to examine the effect of exogenous IGF-1 in mice on hematopoietic precursor cell production, T-cell development and thymic export, and TEC distribution and proliferation. We show that exogenous IGF-1 has positive effects on thymic function through expansion of peripheral T-cell precursor populations as well as TEC populations. Using genetically altered mice to separate these points of regulation by IGF-1, we demonstrate that the predominant effect of IGF-1 on thymic function is through its effects on TEC numbers and function, which in turn supports enhanced T-cell development.
Methods
Animals
Eight- to 12-week-old thymus-intact and thymectomized C57BL/6 mice were purchased from the Animal Production Unit, National Cancer Institute (Frederick, MD). Completeness of thymectomy was confirmed by visual inspection at the time of killing. pLCK-Cre mice were purchased from Taconic (Germantown, NY). IGF-1R/loxP mice were kindly provided by D. LeRoith (National Institute of Diabetes and Digestive and Kidney Diseases [NIDDK], NIH). PSGL-1 knockout mice were purchased from The Jackson Laboratory (Bar Harbor, ME). All murine studies were carried out under approved NCI animal study protocols.
IGF-1 administration
Recombinant human IGF-1 (Tercica; National Hormone and Peptide Program, Torrance, CA) was reconstituted according to the manufacturer's instructions and resuspended in 5% sucrose with 0.1% C57Bl/6 normal mouse sera in phosphate-buffered saline. IGF-1 was administered by continuous infusion at 100 μg/day for 14 days via subcutaneous osmotic pumps (Durect, Cupertino, CA) implanted according to institutionally approved manufacturer's protocols. For 4-week IGF-1 administration, pumps were replaced after the first 2-week infusion of IGF-1 with a second pump containing freshly prepared IGF-1.
Flow cytometry
Single-cell suspensions were prepared from thymus, spleen, and pooled inguinal, axillary, cervical, and mesenteric lymph nodes of mice. Red blood cells were removed by ACK lysis (Quality Biological, Gaithersburg, MD). Lymphocytes were isolated from whole blood after 2 rounds of ACK lysis. Cells were then counted with a hemocytometer or using a Cellometer Auto T4 automated cell counter (Nexcelom Bioscience, Lawrence, MA).
For flow cytometry, cells were incubated with 2.4G2 blocking agent followed by directly conjugated and biotinylated monoclonal antibodies (mAbs), including CD4, CD8α, CD3ϵ, CD25, CD44, c-kit, and Sca-1 (BD Pharmingen, San Diego, CA). Biotinylated mAbs were developed with Pacific Blue–conjugated streptavidin (Invitrogen, Frederick, MD). For enumeration of peripheral and thymic progenitor populations, an antibody cocktail was used to identify lineage− cells, composed of mAbs to TER-119, CD3ϵ, CD8α, TCRβ, Gr-1, CD11b, CD11c, B220, IgM, CD19, and DX5. Intracellular staining of BrdU was performed using BrdU Flow Kits (BD Pharmingen). Multiparameter flow cytometry was performed using a LSRII flow cytometer (Becton Dickinson, San Jose, CA) and analyzed with FlowJo software (TreeStar, Eugene, OR).
TEC analysis
TECs were isolated based on the protocol of Gray et al.30 Pooled thymic fragments from 3 thymi were agitated with a magnetic stirrer in cold RPMI. Following repeated pipetting to remove additional thymocytes, thymic fragments were enzymatically digested with Liberase Blendzyme 4 and DNase I (Roche, Indianapolis, IN), both at a concentration of 0.125% wt/vol in RPMI, followed by mechanical dispersion with a 26-G needle. Digestions were then pooled and resuspended in Hanks buffered saline solution with 1% FCS, 5 mM EDTA, and 0.1% NaN3 wt/vol. Antibodies used for analysis and cell sorting included CD45 and Ly51 (BD Pharmingen) and FITC-conjugated UEA-1 (Vector Laboratories, Burlingame, CA). Cortical TECs (cTECs) (CD45−, Ly51+, UEA-1−) and medullary TECs (mTECs) (CD45−, Ly51−, UEA-1+) were sorted using a fluorescence-activated cell sorting (FACS) Vantage flow cytometer (Becton Dickinson).
T-cell receptor rearrangement excision circle enumeration
The procedure for quantitation of T-cell receptor rearrangement excision circle (TREC) molecules is described in detail elsewhere.31 Briefly, unseparated or CD4+ and CD8+ cells isolated by magnetic bead separation (Miltenyi Biotec, Auburn, CA) were digested with proteinase K (Boehringer Mannheim, Mannheim, Germany). Lysates were added to real-time quantitative polymerase chain reaction (PCR) reactions containing Platinum Quantitative PCR Supermix-UDG with ROX reference dye (Invitrogen) and primer/fluorescent probe sets (Biosynthesis, Lewisville, TX) for mδRec-ψJα–derived TRECs or the CD8β gene as a normalizing control. Amplifications were performed in triplicate on an ABI Prism 7700 Sequence Detection system (Perkin-Elmer, Waltham, MA). PCR conditions were 50°C for 2 minutes, 95°C for 5 minutes, then 40 cycles of 95°C for 15 seconds and 60°C for 1 minute.
Reverse-transcription–PCR
From sorted TEC populations, total RNA was isolated using the RNeasy Micro Kit (Qiagen, Valencia, CA). cDNA was generated from oligo dT primers using the SuperScript III First-Strand kit (Invitrogen). CXCL12 and CCL25 expression was measured using Taqman Gene Expression Assays (Applied Biosystems, Foster City, CA) normalized to mouse GAPDH. For each cytokine-GAPDH combination, validation experiments were performed on serially diluted cDNA prepared from total thymus RNA to confirm relative quantitation of cytokine transcript levels using the comparative CT method (User Bulletin no. 232; Applied Biosystems).
BrdU labeling
Mice were given a continuous oral BrdU (Sigma-Aldrich, St Louis, MO) in the drinking water at a concentration of 0.8 mg/mL, which was made fresh and changed daily. Untreated mice were used as negative controls for BrdU staining. For assessments of cell cycle entry status, mice were given 2 doses of BrdU at 1 mg/dose intraperitoneally at 2-hour intervals. Cells were isolated 2 hours following the second dose and assessed for BrdU incorporation by flow cytometry.
Histology
Frozen sections of thymus (6 μm each) were stained with hematoxylin and eosin and visualized by light microscopy. For analysis of TECs in thymic sections by confocal microscopy or laser scanning cytometry, the following primary antibodies were used: polyclonal rabbit anti–mouse cytokeratin 5 (K5; Covance, Denver, PA), monoclonal rat anti–mouse cytokeratin 8 (K8, Troma-1; Developmental Studies Hybridoma Bank, University of Iowa, Iowa City), polyclonal rabbit anti–mouse cytokeratin 14 (K14; Covance), and biotinylated UEA-1 (Vector Laboratories). Secondary antibodies included FITC-conjugated donkey anti–rabbit IgG, Texas Red–conjugated donkey anti–rat IgG (Jackson ImmunoResearch Laboratories, West Grove, PA), and Alexa-Fluor 633–conjugated streptavidin (Invitrogen). Slides were mounted in Vectashield mounting medium with DAPI (Vector Laboratories) for DNA staining.
Confocal images were acquired with a Zeiss LSM 510 NLO confocal system (Carl Zeiss, Heidelberg, Germany) with an Axiovert 200M inverted microscope and operating with a 2-photon laser tuned to 750 nm, 25-mW argon laser tuned to 488 nm, and a 1-mW HeNe laser tuned to 543 nm. For imaging, 10× Plan-Neofluar 0.3 NA, 20× Plan Apochromat NA 0.75, and 40× Plan Apochromat 1.3 NA oil-immersion objectives were used at various zoom settings. Images were collected using a multitrack configuration where the FITC, Texas Red, and DAPI signal was collected with BP 510/20 IR nm, 565-615 nm, and BP 390-465 nm filters after excitation with 488-nm, 543-nm, and 750-nm laser lines, respectively.
Analyses of stained sections by laser scanning cytometry were performed on the iCys Laser Scanning Cytometer (CompuCyte, Cambridge, MA) using 488-nm, 544-nm, and 405-nm excitation laser lines. Images were acquired at 1-μm resolution.
Statistical analysis
Quantitative data are presented as means plus or minus SEM. Statistical analysis was performed using the nonparametric Mann-Whitney test where indicated.
Results
Exogenous IGF-1 enhances thymic function and thymic output
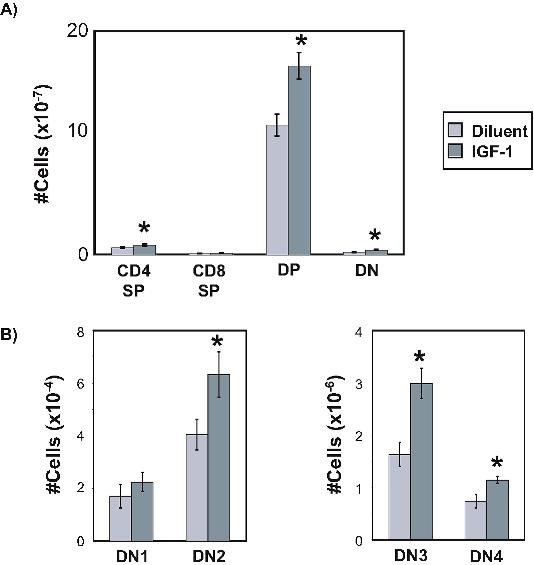
We first assessed the effect of exogenous IGF-1 on thymic function and thymic output by enumerating thymic and peripheral T-cell subsets. A 2-week course of continuous IGF-1 administration resulted in significant increases in thymic mass and cellularity that were symmetric across all thymocyte subsets, including the earliest thymocyte lineage-negative subsets (DN1-DN4). Furthermore, the total level of T-cell receptor excision circles (TRECs) increased in the thymus and periphery, indicating increased thymopoietic activity and export of recent thymic emigrants (Figures 1A-D,2C).
Figure 1.

Exogenous IGF-1 enhances thymic function, thymocyte proliferation, and thymopoietic throughput. Recombinant human IGF-1 was administered by continuous infusion using subcutaneously placed osmotic pumps to deliver a dose of 100 μg/day for a total of 2 weeks in 8- to 12-week-old thymus-intact female C57BL/6 mice. Mice were killed at the end of the 2-week infusion for analysis, or had pumps replaced to deliver another 2-week course of continuous IGF-1 at 100 μg/day for a total course of 4 weeks. Thymus data (A-D) were obtained after 2 weeks of IGF-1 treatment: (A) Normalized thymus weight to body weight ratios; (B) major thymocyte subsets defined by CD4+ and CD8+ staining patterns, from least mature to most mature: DN (CD4− CD8−), DP (CD4+ CD8+), and CD4SP (CD4+ CD8−) and CD8SP (CD4− CD8+); (C) early thymocyte subpopulations defined by lineage− and CD44 and CD25 costaining patterns, with maturation progression from DN1 through DN4; and (D) total thymus TRECs after 2 weeks of IGF-1 treatment. (E) Effect of IGF-1 on thymocyte proliferation. At the specified time points during IGF-1 administration, BrdU was administered intraperitoneally and BrdU uptake in the specified thymocyte subpopulations was assessed as a measure of cell-cycle entry. Shown are representative data of 2 independent experiments with 4 mice per group per time point. *P < .05 between IGF-1– and diluent-treated mice. (F) Effect of IGF-1 on thymopoietic throughput. IGF-1 was administered together with BrdU (0.8 mg/mL) spiked into the drinking water for continuous BrdU administration. At the specified days of IGF-1/BrdU coadministration, thymocyte populations were analyzed for BrdU uptake. Shown are representative data of 2 independent experiments with 4 mice per group per time point. P < .05 between IGF-1– and diluent-treated mice for all subsets at all time points. Error bars represent SEM.
Figure 2.

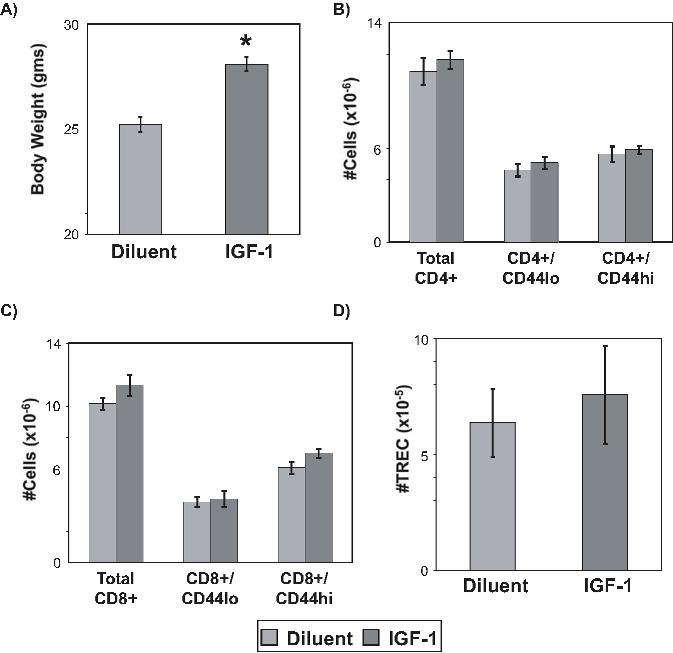
Exogenous IGF-1 enhances thymic output. Recombinant human IGF-1 was administered by continuous infusion using subcutaneously placed osmotic pumps to deliver a dose of 100 μg/day for a total of 2 weeks in 8- to 12-week-old thymus-intact female C57BL/6 mice. Pumps were replaced to deliver another 2-week course of continuous IGF-1 at 100 μg/day for a total course of 4 weeks. Combined spleen and lymph node data were obtained after 4 weeks of IGF-1 treatment: (A) CD4+ subsets; (B) CD8+ subsets; and (C) total TRECs from separated CD4+ and CD8+ T cells. Presented data are representative of 3 independent experiments, with 4 mice per group per experiment. *P < .05 between IGF-1– and diluent-treated mice. Error bars represent SEM.
The effect of IGF-1 administration on thymic activity was further assessed by BrdU incorporation experiments to evaluate thymocyte proliferation and thymopoietic throughput. Based on BrdU incorporation during short-term BrdU treatment, different thymocyte subsets exhibited increased proliferative activity with IGF-1 at different points in the time course of IGF-1 administration. DN2 thymocytes exhibited increased proliferation at day 4 of IGF-1 administration, whereas DP, CD4SP, and CD8SP thymocytes exhibited increased BrdU incorporation by day 7. Because DP thymocytes do not proliferate, we speculated that the increase in BrdU+ DP thymocytes was due to the rapid transit of dividing cells from the preceding immature CD8+CD3− single-positive (ISP) population, the BrdU uptake of which was also increased at this time point (data not shown). By day 10 of IGF-1 administration, no differences in cell proliferation were observed, suggesting that thymocyte proliferation by IGF-1 had equilibrated (Figure 1E).
To determine whether exogenous IGF-1 enhanced thymic function through alterations in thymopoietic throughput, we measured the rate of BrdU accumulation in thymocyte subsets in mice given continuous oral BrdU. Thymocytes undergoing cell division during the DN stage remain BrdU+ through subsequent maturational stages.33,34 The rate of BrdU acquisition in nondividing DP thymocytes therefore reflects the transition rates from DN to DP stages of thymocyte maturation. In CD4 and CD8 single-positive (SP) thymocytes, the accumulation of BrdU+ cells reflects a combination of proliferation of ISP thymocytes that are the developmental precursors to DP thymocytes, maturation of DP into SP thymocytes, and cell proliferation among SP thymocytes.35,36 Continuous administration of BrdU resulted in a greater accumulation of BrdU in all thymocyte subsets (Figure 1F) from mice treated with IGF-1, indicating that thymocyte proliferation and/or developmental throughput was increased. Together with the short-term BrdU experiments, these data are consistent with an IGF-1–induced wave of proliferative expansion initiated first during the DN stage and carrying through successive populations.
Enhancements of thymic function by IGF-1 resulted in an accumulation of peripheral naive T cells and recent thymic emigrants (RTEs) as assessed by TREC enumeration. No quantitative changes in peripheral T-cell and RTE populations were observed after 2 weeks of IGF-1 treatment (data not shown). With 4 weeks of IGF-1 treatment, significant increases in CD4+ and CD8+ naive cell populations, as well as total CD4 and CD8 TREC content, were observed (Figure 2).
Because peripheral T cells express IGF-1R, we determined whether the numeric changes in peripheral T-cell and RTE populations were a result of a thymic-independent effect on peripheral T-cell survival and proliferation versus the thymic effects of IGF-1. Administration of IGF-1 into age-matched thymectomized mice following the same schedule for thymus-intact mice showed no effect on peripheral T-cell and RTE numbers. Increases in body weight of IGF-1–treated animals confirmed its anabolic activity (Figure S1, available on the Blood website; see the Supplemental Materials link at the top of the online article). Assessment of peripheral T-cell proliferation by intracellular Ki67 staining showed no changes with IGF-1 (data not shown). Together, these data demonstrate that exogenous IGF-1 can enhance thymic function, which in turn leads to increased export of mature T cells into the periphery, but that direct effects of IGF-1 on increasing established T-cell populations by proliferation are limited.
Exogenous IGF-1 expands circulating hematopoietic precursors
Thymocyte precursors originating from the bone marrow and extramedullary sites enter the thymus to initiate T-cell development.6,37 Given the role of IGF-1 on hematopoietic stem cell proliferation and differentiation in vitro,17,38 the effects of exogenous IGF-1 on T-cell precursor production and its temporal relationship to thymic function enhancements were examined. IGF-1 administration resulted in a 4-fold increase in the number of peripheral T-cell precursor populations (LSK: lineage−, c-kit+, and Sca-1+)6 by day 4 of administration, which increased to a maximum 10-fold increase by day 7 of IGF-1 infusion, before declining to lower but still significantly greater numbers by day 14. The initial increase in peripheral LSK numbers preceded any numeric change in thymocyte populations (Figure 3A), suggesting that an increased pool of available T-cell precursors may contribute to the enhancement of thymic function by IGF-1. No changes in thymocyte populations, including early thymic progenitors (ETPs; lineage− CD25− CD44hi c-kit+) representing the earliest intrathymic progenitor population, were observed before day 7 of IGF-1 treatment. In contrast to the peripheral LSK population, bone marrow LSK numbers remained constant throughout the duration of IGF-1 administration.
Figure 3.

Exogenous IGF-1 expands peripheral LSK and increases cell cycle entry of bone marrow and peripheral LSK before numeric increases in thymocyte populations. (A) IGF-1 was administered by continuous infusion for 2 weeks into 8- to 10-week-old thymus-intact C57BL/6 mice. At days 4, 7, 10, and 14 of IGF-1 infusion, cohorts of mice were killed and analyzed for thymocyte and peripheral (combined spleen and lymph node) LSK enumeration. Shown are the quantitative fold differences for each cell population between IGF-1–treated and diluent-treated control mice at each time point during IGF-1 administration. Data points are based on the combined data of 2 independent experiments, with 6 mice per group per time point. Quantitative differences between IGF-1 and control mice were statistically significant (P < .05) for all populations except for bone marrow LSK at all time points and thymocyte subpopulations at day 4 only. Dotted lines indicate 1:1 ratio in cell numbers between IGF-1–treated and diluent-treated control mice. ETP indicates early thymic progenitor (lineage−, CD44hi, CD25−, c-kit+); LSK, lineage−, Sca-1+, c-kit+ progenitor cells. (B) BrdU uptake after intraperitoneal BrdU administration was measured in bone marrow and peripheral LSK at day 4 of IGF-1 administration in C57BL/6 mice. Shown are representative data from 3 independent experiments, with 4 mice per group. Error bars represent SEM. *P < .05 between IGF-1– and diluent-treated mice.
IGF-1 might increase circulating LSK populations by enhancing their release from the bone marrow or by enhancing their proliferation. In both bone marrow and peripheral LSK populations, short-term BrdU incorporation increased with IGF-1 administration (Figure 3B), indicating enhanced LSK proliferation by IGF-1. That bone marrow LSK numbers were unchanged despite the increase in proliferation is consistent with their exit into the circulation, which has been suggested to be linked to increased cell-cycle activity.39
Exogenous IGF-1 affects TEC numbers, proliferation, and chemokine expression
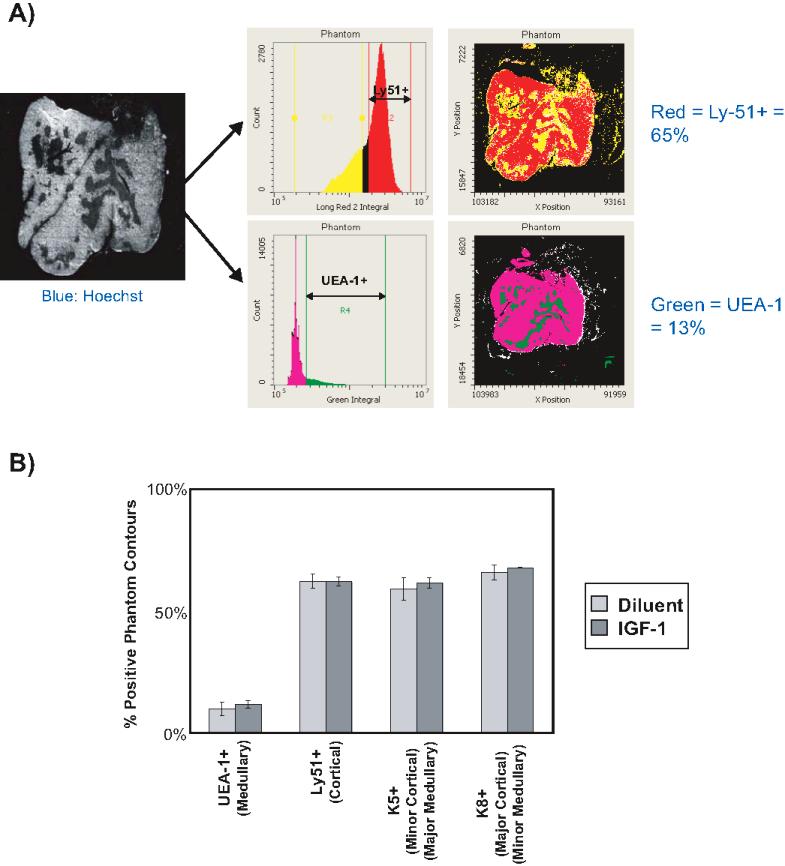
While the preceding observations clearly demonstrate an effect of IGF-1 on hematopoietic stem cell and thymocyte populations, documented responses of TECs to IGF-1 in vitro led to the hypothesis that IGF-1 would have similar effects on TEC populations in vivo. In addition, we sought to understand the effect of exogenous IGF-1 on TEC biology as it relates to overall thymic function. Despite significant increases in thymic weights with IGF-1 administration, there were no differences in thymic cytoarchitecture as assessed by light microscopy (Figure 4A,B). More detailed histologic analysis of TECs with confocal microscopy and laser scanning cytometry using antibody combinations to stain major and minor cortical and medullary TEC populations revealed no differences in the relative content or distribution of these populations (Figure 4B-D; Figure S2). Thus, IGF-1 increases thymic size proportionately, without demonstrated changes in TEC organization.
Figure 4.

Exogenous IGF-1 symmetrically expands TEC populations and increases their turnover. (A) Thymic weights of diluent-treated control mice and mice treated for 14 days with IGF-1. Shown are combined data of 4 experiments with 11 mice per group. Error bars represent SEM. *P < .05 between IGF-1– and diluent-treated mice. (B) Light microscopy image of thymus sections of diluent-treated and IGF-1–treated mice. Images (100× magnification) are representative of 3 mice per group. (C) Representative confocal images (200× magnification) of thymi from diluent-treated and IGF-1–treated mice identifying major cortical (red; K5− K8+), minor cortical (yellow; K5+ K8+), and major medullary (green; K5+ K8−) TEC subsets. (D) Representative confocal images (100× magnification) of thymi identifying minor cortical (red; K5+ K14− UEA-1−), major medullary (pink; K5− K14− UEA-1+), and minor medullary (green; K5− K14+ UEA-1−) TEC subsets. Images are representative of 4 to 6 sections per animal, 3 animals per experimental group. (E) TECs were isolated from thymi of IGF-1–treated and diluent-treated control mice by enzymatic digestion. By flow cytometry, cortical TECs (cTECs) were identified phenotypically as CD45− Ly51+ UEA-1−, and major medullary TECs (mTECs) were identified phenotypically as CD45− Ly51− UEA-1+. For each TEC population, the total number of TECs and their relative proportion to the entire nonhematopoietic CD45− population was calculated. These values were then compared with the total number of thymocytes to compare their relative changes with IGF-1 treatment. Graphs represent the relative fold changes between IGF-1–treated and diluent-treated control mice on day 4 and day 14 of IGF-1 administration in the following parameters: relative percentage of cortical and medullary TECs ( ), total cortical and medullary TECs (
), total cortical and medullary TECs ( ), and total thymocyte numbers (
), and total thymocyte numbers ( ). Shown are combined data from 3 independent experiments with TECs isolated from 3 thymi per group per experiment. (F) For measurements of TEC turnover, continuous BrdU was administered into mice for 5 days, from day 2 to day 7 and from day 9 to day 14, after which BrdU incorporation in sorted cortical and medullary TECs was measured. Shown are representative results from 2 independent experiments, with 3 thymi per group per experiment. Error bars represent SEM. *P < .05 between IGF-1– and diluent-treated mice.
). Shown are combined data from 3 independent experiments with TECs isolated from 3 thymi per group per experiment. (F) For measurements of TEC turnover, continuous BrdU was administered into mice for 5 days, from day 2 to day 7 and from day 9 to day 14, after which BrdU incorporation in sorted cortical and medullary TECs was measured. Shown are representative results from 2 independent experiments, with 3 thymi per group per experiment. Error bars represent SEM. *P < .05 between IGF-1– and diluent-treated mice.
To enumerate TEC populations, TECs were isolated from thymi of control and IGF-1 mice by enzymatic digestion and analyzed by flow cytometry. Relative and total cortical TEC (cTEC) and medullary TEC (mTEC) numbers were determined. At day 4 of IGF-1 administration, there were no differences in either the relative or absolute number of cTECs and mTECs that correlated with the absence of any changes in total thymocyte number (Figure 4E). However, by day 14 of IGF-1 administration, total cTEC and mTEC numbers were significantly increased with a concomitant increase in thymocyte number. Relative cTEC and mTEC numbers remained unchanged, consistent with the histologic observations. The expansion of cTEC and mTEC populations coincided with increased proliferative expansion throughout the period of IGF-1 administration as determined by BrdU uptake after continuous BrdU administration (Figure 4F). Similarly, CD45− CD31+ vascular endothelial cells exhibited increased expansion (data not shown). These data clearly demonstrate that IGF-1, in addition to acting on hematopoietic elements of T-cell development, also affects TEC populations in a highly coordinate fashion, reinforcing the principle that TEC and thymocyte development are closely interdependent.40
Chemokines are expressed by TECs to regulate directional migration of developing thymocytes through thymic developmental niches and ensuring ordered T-cell development. CXCL12 and CCL25 have been shown to play critical roles in thymocyte migration and T-cell development.41–43 To determine whether exogenous IGF-1 affected chemokine expression, we measured CXCL12 and CCL25 expression in sorted TEC populations at time points during IGF-1 administration corresponding to periods of dynamic changes in thymocyte population numbers and turnover. CXCL12 and CCL25 expression was increased in both cTECs and mTECs by day 4 of IGF-1 administration. By day 7 of IGF-1 administration, expression of both chemokines remained elevated in cTECs, but normalized in mTECs (Table 1). Surface expression of the cognate chemokine receptors, CXCR4 and CCR9, on thymocyte subpopulations was not altered (data not shown). Thus, in addition to expanding their numbers, exogenous IGF-1 also affects function of TECs, which, through expansion of developmental niches and expression of chemokines such as CXCL12 and CCL25, could serve to facilitate enhanced T-cell development.
Table 1.
Effect of IGF-1 on expression of CXCL12 and CCL25 in thymic epithelial cells
| Day of IGF-1 infusion | Relative expression, IGF/diluent |
|
|---|---|---|
| CXCL12 | CCL25 | |
| Cortical TECs (CD45−, Ly51+, UEA1−) | ||
| 4 | 3.20 (2.75-3.73) | 1.92 (1.26-2.93) |
| 7 | 2.19 (1.78-2.69) | 2.04 (1.38-2.50) |
| Medullary TECs (CD45−, Ly51−, UEA1+) | ||
| 4 | 7.18 (4.00-15.14) | 1.85 (1.51-2.77) |
| 7 | 1.44 (0.92-2.21) | 0.51 (0.38-0.69) |
Real-time quantitative RT-PCR was performed from cDNA isolated from sorted thymic epithelial cells pooled from 3 mice per group per time point of IGF-1 versus diluent controls using primer/probe sets specific for CXCL12, CCL25, and GAPDH as the endogenous control. Data presented are the means (with ranges in parentheses) of relative expression of CXCL12 and CCL25 in the specified TEC population between IGF-1– and diluent-treated mice at day 4 and day 7 of diluent/IGF-1 administration. Values were determined using the comparative CT method.
IGF-1's effects on TECs are sufficient for thymic function enhancement
Given the interplay between thymocytes and TECs in T-cell development and expression of IGF-1R in both populations, we sought to separate the effects of IGF-1 on these populations by generating mice that lack IGF-1R expression on thymocytes and T cells. Mice containing loxP sites flanking the third exon of the IGF-1R receptor encoding the high-affinity binding region to IGF-144 were crossed with mice containing cre recombinase driven by the proximal LCK promoter to generate T cell–specific IGF-1R knockout mice (T-IGF-1R−). As the proximal LCK promoter is activated at the DN1 stage of thymocyte development,45 the predicted deletion of exon 3 of IGF-1R in more mature thymocytes and peripheral T cells was confirmed by reverse-transcription (RT)–PCR. In contrast, TEC populations, as well as more immature LSK hematopoietic stem cells, did not undergo exon 3 deletion but instead retained an intact IGF-1R message (Figure 5A,B), confirming cre-mediated deletion tissue specificity.
Figure 5.

Abrogation of IGF-1R signaling in thymocytes results in a decrease in thymic function and in peripheral T-cell and RTE populations that is restored by IGF-1 administration. (A) Schematic representation of IGF-1R/loxP mice illustrating the generation of T-IGF-1R− mice upon cre-mediated deletion of exon 3 encoding the high-affinity binding site in cells expressing the proximal LCK promoter. Arrows indicate the location of PCR primers used in RT-PCR to confirm exon 3 deletion. (B) RT-PCR of sorted thymocyte and TEC populations confirming the loss of IGF-1R exon 3 expression in thymocytes in mice expressing the cre recombinase (−), but not in LSK, vascular endothelium (CD45− CD31+), cTECs (CD45− Ly51+ UEA-1−), or mTECs (CD45− Ly51− UEA-1+; +). (C) Decreased thymocyte populations in T-IGF-1R− mice are associated with the loss of splenic naive CD4+ (D), CD8+ (E), and RTE (F) populations. n = 16 to 22 mice per experimental group combined from 4 independent experiments. *P < .05 between IGF-1– and diluent-treated mice. (G-I) T-IGF-1R− mice and wild-type littermates lacking cre expression (T-IGF-1R+) were given a 2-week course of IGF-1 at a dose of 100 μg/day. (G) Thymocyte populations, (H) thymic TRECs (*P < .05 between each IGF-1 treatment group and both diluent control groups; #, P = .07 between diluent-treated T-IGF-1R+ and T-IGF-1R− groups), and (I) peripheral LSK (*P < .05 between each IGF-1 treatment group and both diluent control groups) were enumerated. Shown are composite data of 2 independent experiments with 7 to 8 mice per experimental group. Error bars represent SEM.
At baseline steady state, T-IGF-1R− mice had significantly fewer thymocytes than wild-type littermates, resulting predominantly from decreased numbers of DP and CD4SP thymocytes (Figure 5C). In addition, peripheral naive CD44lo CD4+ and CD8+ T cells as well as RTEs were significantly decreased in T-IGF-1R− mice (Figure 5D-F), consistent with decreased thymic output. These data demonstrate that the production and maintenance of T cells are regulated in part by IGF-1R signaling on thymocytes.
We then sought to determine whether IGF-1R signaling in TECs could overcome the deleterious effects of loss of IGF-1R in thymocytes by examining the effect of exogenous IGF-1 on the restoration of thymic function in the T-IGF-1R− mice. Despite the absence of IGF-1R expression in thymocyte populations following the DN stage, exogenous IGF-1 increased thymocyte cellularity and function to levels comparable to similarly treated T-IGF-1R+ mice. Moreover, in both T-IGF-1R+ and T-IGF-1R− mice, peripheral LSK populations were significantly increased (Figure 5G-I). These experiments strongly suggest that IGF-1 controls thymic function through its effect on TECs. The possibility remains, however, that the increased availability of thymocyte precursor LSK numbers induced by IGF-1 and their subsequent importation into the thymus may also play a role in restoring thymic function.
To address this latter possibility, IGF-1 was administered in mice in which thymocyte precursor importation was severely limited. In mice lacking the P-selectin ligand PSGL-1 (PSGL-1KO), ETP homing into the thymus is significantly impaired, leading to an availability of niches for ETP importation.46 Increases in thymic function when ETP import is held to low levels would indicate that the IGF-1–induced TEC expansion alone is sufficient for enhancements of overall thymic function. IGF-1 was therefore administered into wild-type and age-matched PSGL-1KO mice, after which peripheral LSK and ETP populations were enumerated. Compared with age-matched wild-type nonlittermates, total thymocyte numbers in PSGL-1KO mice were modestly but significantly decreased. IGF-1 administration increased thymic cellularity to levels comparable to those of wild-type mice. Thymic TREC in IGF-1–treated PSGL-1KO mice was also increased over that of diluent-treated control mice. On the other hand, ETP frequency in IGF-1–treated PSGL-1KO mice was substantially less than that of wild-type mice (Figure 6A-C; Figure S3). Although ETP number was statistically increased in IGF-1–treated PSGL-1KO mice over diluent-treated controls, it too was significantly less than that of wild-type mice. Peripheral LSK frequency and number, as expected, were increased with IGF-1 administration in both wild-type and PSGL-1KO mice (Figure 6D and data not shown), and the impairment of precursor importation in PSGL-1KO mice is reflected by the increase in the LSK/ETP ratio. Importantly, the LSK/ETP ratio in IGF-1–treated mice normalized to diluent-treated mice was roughly equivalent between PSGL-1KO and wild-type mice. For both wild-type and PSGL-1KO mice, IGF-1 administration resulted in a greater than 2-fold increase in the normalized IGF-1/diluent ratio of LSK/ETP (Figure 6E). That IGF-1 administration led to increased thymic function in the presence of impaired ETP homing and that the expansion in peripheral LSK with IGF-1 treatment did not lead to a disproportionate increase in ETPs is strong evidence that the IGF-1 effect on TECs is sufficient to positively regulate thymic function.
Figure 6.

Exogenous IGF-1 enhances thymic function in PSGL-1KO mice without a disproportionate increase in ETPs. PSGL-1KO mice and age-matched wild-type controls were given a 2-week course of IGF-1, after which thymocyte subsets, ETPs, and peripheral (combined spleen and lymph node) LSK were enumerated. (A) Changes in thymocyte number and thymic TREC number in PSGL-1KO mice treated with IGF-1. (B) ETP numbers in wild-type mice compared with diluent- and IGF-1–treated PSGL-1KO mice. (C,D) changes in frequencies of ETPs (C) and peripheral LSK (D) in diluent- and IGF-1–treated wild-type and PSGL-1KO mice. (E) LSK/ETP ratios in diluent- and IGF-1–treated wild-type and PSGL-1KO mice. The normalized IGF-1/diluent ratios for LSK/ETP ratios in B6 and PSGL-1KO mice are indicated. n = 6 to 11 mice per group combined from 3 independent experiments (*P < .05). Error bars represent SEM.
Discussion
Regulation of thymic function could conceivably occur at 3 levels. The first involves regulating the availability of bone marrow–derived precursors and their entry into the thymus. The second involves regulating the inherent proliferation and differentiation characteristics of developing thymocytes. The third involves regulating TEC number and function, which form the functional platforms on which thymic T-cell development occurs. Given the high level of interdependence between thymocyte and TEC development,40 quantitative and functional changes in one compartment often affect another. For agents such as keratinocyte growth factor, the cognate receptor for which is expressed exclusively on TECs,8 the role of TEC expansion in enhancing thymopoiesis is clear. In contrast, for IGF-1, the receptor for which is expressed on both hematopoietic and epithelial cells, dissecting the mechanisms by which IGF-1 enhances thymic function is more difficult. Although IGF-1 has been implicated in individual studies to positively affect thymocytes, hematopoietic stem cells, and TECs, the relative contributions of these effects regarding thymic function in vivo remained unclear. In studies assessing the effect of IGF-1 on thymic reconstitution after HSCT, whether this was due to a direct effect of IGF-1 on thymocytes themselves, a secondary effect on TECs, and/or expansion of T-cell precursor populations was not studied.28,29 Finally, given IGF-1's antiapoptotic effects on peripheral T cells,19,20 the possibility remained that the expansion of peripheral T cells with short-term IGF-1 was independent of IGF-1's thymic effects.
Our results demonstrate that short-term administration of IGF-1 results in enhancements of thymic function through effects on both thymocytes and TECs, ultimately leading to increased RTE export. IGF-1–induced changes are thymic dependent; no changes were observed in the peripheral T-cell populations in IGF-1–treated thymectomized mice. IGF-1 administration resulted in a substantial expansion in peripheral LSK as well as increased turnover in both peripheral and bone marrow LSK. The changes in LSK populations preceded any numeric changes in thymocyte subset numbers, leading to the possibility that increasing the availability of thymocyte precursors may contribute to the increase in overall thymic function. These observations confirm clinical and preclinical observations that the speed of immune recovery is influenced by the hematopoietic stem cell dose in HSCT5 and provide strong support for the use of IGF-1 as a stem cell–mobilizing agent in clinical HSCT, particularly in nonmalignant settings that avoid the risk of growth and metastasis of IGF-1R+ tumors.47
In addition to its effects on thymocytes and their precursors, IGF-1 also enhances TEC numbers and proliferation, supporting the alternative, although not mutually exclusive, possibility that expansions of TECs rather than precursor availability are required to enhance thymic function. Because thymic T-cell development occurs along a linear pathway of specific functional and geographic developmental niches,48 enhancement of thymic function would presumably require the availability of increased spatial or functional niche size at all stages of development in addition to the precursor expansion stage. That we observed differences in the expression of the chemokines CXCL12 and CCL25 by sorted cTECs and mTECs from IGF-1–treated mice suggests that modulation of chemokine expression by TECs could play an important role in regulating thymic T-cell development by modulating the movement of thymocytes from one developmental niche to another. CXCL12, in particular, may play an important role in this, as it has been demonstrated to have a role in both chemotaxis and chemofugetaxis of thymocytes,41,42 suggesting roles in the migration of DN thymocytes from the corticomedullary junction toward the subcapsular epithelium and the subsequent migration of DP thymocytes from the subcapsular epithelium back to the medulla. CCL25 is critical for the importation of T-cell precursors into the thymus.49 In this context, the importation of thymic precursors is dependent on the availability of open thymic niches that regulate thymic homing and T-cell development in a gated fashion; progression of thymocytes from one developmental stage to the next is dictated by the availability of these niches.50 Finally, the aforementioned studies relating stem cell dose to T-cell reconstitution were performed in the context of HSCT, where the developmental niches in the thymus are devoid of thymocytes due to radiation and chemotherapy effects, and therefore may be more dependent on the availability of circulating progenitors for their recovery. Consistent with this, titrating increasing numbers of donor congenic bone marrow cells into nonirradiated thymus-intact mice increased the relative number of donor-derived thymocytes, but did not increase overall thymocyte numbers (Y.-W.C., unpublished data, June 2007).
To separate the differential effects of IGF-1 among T-cell precursors, thymocytes, and TECs, we generated mice in which IGF-1R is absent on most thymocytes and mature T cells, but present on TEC and LSK populations (T-IGF-1R− mice). These mice had decreased thymocyte, peripheral naive T-cell, and RTE populations at baseline, consistent with the role of IGF-1 in maintaining these populations. That the increased susceptibility to apoptosis as a result of the loss of antiapoptotic functions conferred by IGF-1R signaling may further contribute to the lower peripheral T-cell numbers including RTEs remains to be formally tested. With exogenous IGF-1, however, thymocyte numbers and function were completely restored, suggesting that the positive effect of IGF-1 on IGF-1R+ TECs was sufficient to overcome the absence of IGF-1R signaling in thymocytes. Because IGF-1 administration into T-IGF-1R− mice does not rule out the possibility of enlarged precursor populations positively regulating thymic function, we also examined the effect of IGF-1 on thymic function in PSGL-1KO mice, where thymic homing of ETPs is significantly impaired,46 and compared relative changes in peripheral LSK and ETP populations to that observed in normal mice. If the expansion of these precursor populations by IGF-1 was primarily responsible for enhancing thymic function, it would be predicted that a disproportionate number of ETPs would accumulate in these mice. In this scenario, not only are peripheral LSK numbers significantly increased, but the ETP niche within the thymus is empty and increasingly receptive to the importation of circulating T progenitors due to presumptive up-regulation of P-selectin.46 This, however, was not observed experimentally. Although thymic cellularity was increased, ETP numbers were not disproportionately increased in IGF-1–treated PSGL-1KO mice despite the disproportionate increase in peripheral LSK numbers. Furthermore, the LSK/ETP ratios of IGF-1–treated normalized to diluent-treated PSGL-1KO mice were equivalent to that of wild-type mice. Disproportionate increases in precursor importation in IGF-1–treated PSGL-KO mice that would be the consequence of increased precursor availability would result in a decrease of the normalized ratio. Together with the findings in IGF-1–treated T-IGF-1-R− mice, this finding therefore supports the notion that importation of T-cell progenitors from the circulation is a TEC-mediated phenomenon, and that IGF-1's effects on TECs are sufficient to positively regulate thymic function, whereas the effect of increased LSK populations on the functional status in the nondepleted thymus is secondary under these experimental conditions. As was stated before, these effects are not necessarily mutually exclusive. In the setting of T-cell reconstitution after acute lymphodepletion, where thymic developmental niches are “empty,” both effects of IGF-1 may play an important role in accelerating thymus function recovery. Further dissection of these individual effects of IGF-1 in acute lymphodepletion models requires further investigation.
In summary, these studies with IGF-1 identify entry of thymocyte precursors into the thymus and the proportionate expansion of developmental niches through alterations in TEC numbers and function as critical points of regulation in thymic productivity, and establish IGF-1 as a positive thymic regulator. Because the signaling pathway through the IGF-1R is distinct from those of other positive thymic regulators such as KGF,51 the possibility exists for synergistic enhancing effects on thymic function. However, the use of these agents in the context of HSCT for the treatment of cancer must proceed with caution, as certain tumors are dependent on IGF-1R signaling for their survival, proliferation, and metastasis.47
Supplementary Material
Acknowledgments
The authors thank Frances Hakim for critical review of the paper and Nyana Singh for technical assistance.
This work was entirely supported by intramural funding through the National Cancer Institute.
Footnotes
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Authorship
Contribution: Y.-W.C. designed and performed the research, analyzed data, and wrote the paper; S.S., B.C., V.K., and D.H. performed research; W.T. and S.G. contributed critical analytical tools and expertise; and R.E.G. designed the research and wrote the paper.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Yu-Waye Chu, Experimental Immunology Branch, Center for Cancer Research, NIH, Building 10CRC/Rm 3-3288, 10 Center Drive, Bethesda, MD 20892-1360; e-mail: chuy@mail.nih.gov.
References
- 1.Williams KM, Hakim FT, Gress RE. T cell immune reconstitution following lymphodepletion. Semin Immunol. 2007;19:318–330. doi: 10.1016/j.smim.2007.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hakim FT, Gress RE. Thymic involution: implications for self-tolerance. Methods Mol Biol. 2007;380:377–390. doi: 10.1007/978-1-59745-395-0_24. [DOI] [PubMed] [Google Scholar]
- 3.Anderson G, Lane PJ, Jenkinson EJ. Generating intrathymic microenvironments to establish T-cell tolerance. Nat Rev Immunol. 2007;7:954–963. doi: 10.1038/nri2187. [DOI] [PubMed] [Google Scholar]
- 4.Takahama Y. Journey through the thymus: stromal guides for T-cell development and selection. Nat Rev Immunol. 2006;6:127–135. doi: 10.1038/nri1781. [DOI] [PubMed] [Google Scholar]
- 5.Chen BJ, Cui X, Sempowski GD, Domen J, Chao NJ. Hematopoietic stem cell dose correlates with the speed of immune reconstitution after stem cell transplantation. Blood. 2004;103:4344–4352. doi: 10.1182/blood-2003-07-2534. [DOI] [PubMed] [Google Scholar]
- 6.Bhandoola A, von Boehmer H, Petrie HT, Zuniga-Pflucker JC. Commitment and developmental potential of extrathymic and intrathymic T cell precursors: plenty to choose from. Immunity. 2007;26:678–689. doi: 10.1016/j.immuni.2007.05.009. [DOI] [PubMed] [Google Scholar]
- 7.Berzins SP, Godfrey DI, Miller JF, Boyd RL. A central role for thymic emigrants in peripheral T cell homeostasis. Proc Natl Acad Sci U S A. 1999;96:9787–9791. doi: 10.1073/pnas.96.17.9787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rossi SW, Jeker LT, Ueno T, et al. Keratinocyte growth factor (KGF) enhances postnatal T-cell development via enhancements in proliferation and function of thymic epithelial cells. Blood. 2007;109:3803–3811. doi: 10.1182/blood-2006-10-049767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Jones JI, Clemmons DR. Insulin-like growth factors and their binding proteins: biological actions. Endocr Rev. 1995;16:3–34. doi: 10.1210/edrv-16-1-3. [DOI] [PubMed] [Google Scholar]
- 10.LeRoith D, Yakar S. Mechanisms of disease: metabolic effects of growth hormone and insulin-like growth factor 1. Nat Clin Pract Endocrinol Metab. 2007;3:302–310. doi: 10.1038/ncpendmet0427. [DOI] [PubMed] [Google Scholar]
- 11.Clark R. The somatogenic hormones and insulin-like growth factor-1: stimulators of lymphopoiesis and immune function. Endocr Rev. 1997;18:157–179. doi: 10.1210/edrv.18.2.0296. [DOI] [PubMed] [Google Scholar]
- 12.Clark R, Strasser J, McCabe S, Robbins K, Jardieu P. Insulin-like growth factor-1 stimulation of lymphopoiesis. J Clin Invest. 1993;92:540–548. doi: 10.1172/JCI116621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liu JL, LeRoith D. Insulin-like growth factor I is essential for postnatal growth in response to growth hormone. Endocrinology. 1999;140:5178–5184. doi: 10.1210/endo.140.11.7151. [DOI] [PubMed] [Google Scholar]
- 14.Behringer RR, Lewin TM, Quaife CJ, Palmiter RD, Brinster RL, D'Ercole AJ. Expression of insulin-like growth factor I stimulates normal somatic growth in growth hormone-deficient transgenic mice. Endocrinology. 1990;127:1033–1040. doi: 10.1210/endo-127-3-1033. [DOI] [PubMed] [Google Scholar]
- 15.Brocardo MG, Schillaci R, Galeano A, et al. Early effects of insulin-like growth factor-1 in activated human T lymphocytes. J Leukoc Biol. 2001;70:297–305. [PubMed] [Google Scholar]
- 16.Arkins S, Rebeiz N, Biragyn A, Reese DL, Kelley KW. Murine macrophages express abundant insulin-like growth factor-I class I Ea and Eb transcripts. Endocrinology. 1993;133:2334–2343. doi: 10.1210/endo.133.5.8404686. [DOI] [PubMed] [Google Scholar]
- 17.Huang S, Terstappen LW. Lymphoid and myeloid differentiation of single human CD34+, HLA-DR+, CD38- hematopoietic stem cells. Blood. 1994;83:1515–1526. [PubMed] [Google Scholar]
- 18.Zumkeller W, Burdach S. The insulin-like growth factor system in normal and malignant hematopoietic cells. Blood. 1999;94:3653–3657. [PubMed] [Google Scholar]
- 19.Walsh PT, O'Connor R. The insulin-like growth factor-I receptor is regulated by CD28 and protects activated T cells from apoptosis. Eur J Immunol. 2000;30:1010–1018. doi: 10.1002/(SICI)1521-4141(200004)30:4<1010::AID-IMMU1010>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- 20.Walsh PT, Smith LM, O'Connor R. Insulin-like growth factor-1 activates Akt and Jun N-terminal kinases (JNKs) in promoting the survival of T lymphocytes. Immunology. 2002;107:461–471. doi: 10.1046/j.1365-2567.2002.01525.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bernabei P, Bosticardo M, Losana G, et al. IGF-1 down-regulates IFN-gamma R2 chain surface expression and desensitizes IFN-gamma/STAT-1 signaling in human T lymphocytes. Blood. 2003;102:2933–2939. doi: 10.1182/blood-2003-01-0100. [DOI] [PubMed] [Google Scholar]
- 22.Kelley KW, Meier WA, Minshall C, et al. Insulin growth factor-I inhibits apoptosis in hematopoietic progenitor cells: implications in thymic aging. Ann N Y Acad Sci. 1998;840:518–524. doi: 10.1111/j.1749-6632.1998.tb09590.x. [DOI] [PubMed] [Google Scholar]
- 23.Gagnerault MC, Postel-Vinay MC, Dardenne M. Expression of growth hormone receptors in murine lymphoid cells analyzed by flow cytofluorometry. Endocrinology. 1996;137:1719–1726. doi: 10.1210/endo.137.5.8612507. [DOI] [PubMed] [Google Scholar]
- 24.Kooijman R, Scholtens LE, Rijkers GT, Zegers BJ. Type I insulin-like growth factor receptor expression in different developmental stages of human thymocytes. J Endocrinol. 1995;147:203–209. doi: 10.1677/joe.0.1470203. [DOI] [PubMed] [Google Scholar]
- 25.Kecha O, Brilot F, Martens H, et al. Involvement of insulin-like growth factors in early T cell development: a study using fetal thymic organ cultures. Endocrinology. 2000;141:1209–1217. doi: 10.1210/endo.141.3.7360. [DOI] [PubMed] [Google Scholar]
- 26.de Mello-Coelho V, Villa-Verde DM, Dardenne M, Savino W. Pituitary hormones modulate cell-cell interactions between thymocytes and thymic epithelial cells. J Neuroimmunol. 1997;76:39–49. doi: 10.1016/s0165-5728(97)00031-3. [DOI] [PubMed] [Google Scholar]
- 27.Timsit J, Savino W, Safieh B, et al. Growth hormone and insulin-like growth factor-I stimulate hormonal function and proliferation of thymic epithelial cells. J Clin Endocrinol Metab. 1992;75:183–188. doi: 10.1210/jcem.75.1.1619008. [DOI] [PubMed] [Google Scholar]
- 28.Alpdogan O, Muriglan SJ, Kappel BJ, et al. Insulin-like growth factor-I enhances lymphoid and myeloid reconstitution after allogeneic bone marrow transplantation. Transplantation. 2003;75:1977–1983. doi: 10.1097/01.TP.0000070167.81584.A2. [DOI] [PubMed] [Google Scholar]
- 29.Montecino-Rodriguez E, Clark R, Dorshkind K. Effects of insulin-like growth factor administration and bone marrow transplantation on thymopoiesis in aged mice. Endocrinology. 1998;139:4120–4126. doi: 10.1210/endo.139.10.6263. [DOI] [PubMed] [Google Scholar]
- 30.Gray DH, Chidgey AP, Boyd RL. Analysis of thymic stromal cell populations using flow cytometry. J Immunol Methods. 2002;260:15–28. doi: 10.1016/s0022-1759(01)00493-8. [DOI] [PubMed] [Google Scholar]
- 31.Chu YW, Memon SA, Sharrow SO, et al. Exogenous IL-7 increases recent thymic emigrants in peripheral lymphoid tissue without enhanced thymic function. Blood. 2004;104:1110–1119. doi: 10.1182/blood-2003-10-3635. [DOI] [PubMed] [Google Scholar]
- 32.Applied Biosystems. User Bulletin #2. [Accessed February 1, 2008]; http://www3.appliedbiosystems.com/cms/groups/mcb_support/documents/generaldocuments/cms_040980.pdf.
- 33.Vasseur F, Le Campion A, Penit C. Scheduled kinetics of cell proliferation and phenotypic changes during immature thymocyte generation. Eur J Immunol. 2001;31:3038–3047. doi: 10.1002/1521-4141(2001010)31:10<3038::aid-immu3038>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
- 34.Penit C, Vasseur F. Sequential events in thymocyte differentiation and thymus regeneration revealed by a combination of bromodeoxyuridine DNA labeling and antimitotic drug treatment. J Immunol. 1988;140:3315–3323. [PubMed] [Google Scholar]
- 35.Le Campion A, Vasseur F, Penit C. Regulation and kinetics of premigrant thymocyte expansion. Eur J Immunol. 2000;30:738–746. doi: 10.1002/1521-4141(200003)30:3<738::AID-IMMU738>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 36.Pénit C, Vasseur F. Expansion of mature thymocyte subsets before emigration to the periphery. J Immunol. 1997;159:4848–4856. [PubMed] [Google Scholar]
- 37.Maillard I, Schwarz BA, Sambandam A, et al. Notch-dependent T-lineage commitment occurs at extrathymic sites following bone marrow transplantation. Blood. 2006;107:3511–3519. doi: 10.1182/blood-2005-08-3454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.McCubrey JA, Steelman LS, Mayo MW, Algate PA, Dellow RA, Kaleko M. Growth-promoting effects of insulin-like growth factor-1 (IGF-1) on hematopoietic cells: overexpression of introduced IGF-1 receptor abrogates interleukin-3 dependency of murine factor-dependent cells by a ligand-dependent mechanism. Blood. 1991;78:921–929. [PubMed] [Google Scholar]
- 39.Morrison SJ, Wright DE, Weissman IL. Cyclophosphamide/granulocyte colony-stimulating factor induces hematopoietic stem cells to proliferate prior to mobilization. Proc Natl Acad Sci U S A. 1997;94:1908–1913. doi: 10.1073/pnas.94.5.1908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Anderson G, Jenkinson EJ. Lymphostromal interactions in thymic development and function. Nat Rev Immunol. 2001;1:31–40. doi: 10.1038/35095500. [DOI] [PubMed] [Google Scholar]
- 41.Plotkin J, Prockop SE, Lepique A, Petrie HT. Critical role for CXCR4 signaling in progenitor localization and T cell differentiation in the postnatal thymus. J Immunol. 2003;171:4521–4527. doi: 10.4049/jimmunol.171.9.4521. [DOI] [PubMed] [Google Scholar]
- 42.Poznansky MC, Olszak IT, Evans RH, et al. Thymocyte emigration is mediated by active movement away from stroma-derived factors. J Clin Invest. 2002;109:1101–1110. doi: 10.1172/JCI13853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Svensson M, Marsal J, Uronen-Hansson H, et al. Involvement of CCR9 at multiple stages of adult T lymphopoiesis. J Leukoc Biol. 2008;83:156–164. doi: 10.1189/jlb.0607423. [DOI] [PubMed] [Google Scholar]
- 44.Holzenberger M, Dupont J, Ducos B, et al. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature. 2003;421:182–187. doi: 10.1038/nature01298. [DOI] [PubMed] [Google Scholar]
- 45.Shimizu C, Kawamoto H, Yamashita M, et al. Progression of T cell lineage restriction in the earliest subpopulation of murine adult thymus visualized by the expression of lck proximal promoter activity. Int Immunol. 2001;13:105–117. doi: 10.1093/intimm/13.1.105. [DOI] [PubMed] [Google Scholar]
- 46.Rossi FM, Corbel SY, Merzaban JS, et al. Recruitment of adult thymic progenitors is regulated by P-selectin and its ligand PSGL-1. Nat Immunol. 2005;6:626–634. doi: 10.1038/ni1203. [DOI] [PubMed] [Google Scholar]
- 47.LeRoith D, Roberts CT., Jr The insulin-like growth factor system and cancer. Cancer Lett. 2003;195:127–137. doi: 10.1016/s0304-3835(03)00159-9. [DOI] [PubMed] [Google Scholar]
- 48.Savino W, Dardenne M, Carnaud C. The conveyor belt model for intrathymic T-cell migration. Immunol Today. 1996;17:97–98. doi: 10.1016/0167-5699(96)80587-1. [DOI] [PubMed] [Google Scholar]
- 49.Schwarz BA, Sambandam A, Maillard I, Harman BC, Love PE, Bhandoola A. Selective thymus settling regulated by cytokine and chemokine receptors. J Immunol. 2007;178:2008–2017. doi: 10.4049/jimmunol.178.4.2008. [DOI] [PubMed] [Google Scholar]
- 50.Goldschneider I. Cyclical mobilization and gated importation of thymocyte progenitors in the adult mouse: evidence for a thymus-bone marrow feedback loop. Immunol Rev. 2006;209:58–75. doi: 10.1111/j.0105-2896.2006.00354.x. [DOI] [PubMed] [Google Scholar]
- 51.de Giorgi V, Sestini S, Massi D, Ghersetich I, Lotti T. Keratinocyte growth factor receptors. Dermatol Clin. 2007;25:477–485. doi: 10.1016/j.det.2007.06.017. vii. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}