

Abstract
T lymphocyte development and function are tightly regulated by the intrinsic death pathway through members of the Bcl-2 family. Genetic studies have demonstrated that the Bcl-2 family member Mcl-1 is an important anti-apoptotic protein in the development of multiple cell types including T lymphocytes. However, the expression pattern and anti-apoptotic roles of Mcl-1 in T lymphocytes at different developmental stages remain to be fully determined. In this study, we examined the expression pattern of Mcl-1 in different populations of T cells at the single cell level and found that Mcl-1 protein is constitutively expressed in all T cell populations and upregulated upon T cell antigen receptor (TCR) stimulation. We then investigated the role of Mcl-1 in the survival of these different populations by conditionally deleting Mcl-1 at various T cell stages. Our results show that Mcl-1 is required for the survival of double-negative (DN) and single-positive (SP) thymocytes as well as naïve and activated T cells. Furthermore, we demonstrate that Mcl-1 functions together with Bcl-xL to promote double-positive (DP) thymocyte survival. Thus, Mcl-1 is a critical anti-apoptotic factor for the survival of T cells at multiple stages in vivo.
Keywords: anti-apoptosis, Mcl-1, T lymphocyte survival
Introduction
The life of a T lymphocyte can be divided into multiple distinct stages (1). In the thymus, T lymphocytes are generated through several well-characterized steps marked by the expression of the co-receptors CD4 and CD8. Thymocyte precursors that are CD4−CD8− double negative (DN) mature into CD4+CD8+ double positive (DP) and then either CD4+ or CD8+ single positive (SP) thymocytes. SP thymocytes exit to the periphery as naïve T cells. When a naïve T cell encounters its cognate antigen it undergoes activation and proliferation and differentiates into effector T cells. After the peak of the immune response, 90–95% of the effector T cells are eliminated during the contraction phase, while the remaining 5–10% of effector cells become memory T cells (1).
Proteins of the Bcl-2 family function in the intrinsic death pathway, playing important roles in regulating the life and death of T lymphocytes (2–4). These proteins contain between one and four distinct regions of homology termed Bcl-2 homology (BH) domains. The members of the Bcl-2 family can be broadly divided into anti-apoptotic and pro-apoptotic groups. Anti-apoptotic Bcl-2 family members including Bcl-2, Bcl-xL, and Mcl-1 are expressed in T lymphocytes. For example, both Bcl-2 and Mcl-1 are expressed in DN thymocytes, whereas the expression of Bcl-xL is largely inversely correlated with that of Bcl-2 (5–9). Bcl-2 is downregulated in DP and upregulated in SP thymocytes, while Bcl-xL is strongly upregulated in DP but downregulated in SP cells (6–9). Furthermore, Bcl-2 is expressed at a relatively high level in naïve T cells and downregulated in activated/effector CD8+ T cells (10–12). In contrast, Bcl-xL is not expressed in naïve T cells but is rapidly upregulated in activated/effector T cells (11, 13). The complex pattern of expression of Bcl-2 and Bcl-xL may be related to observations that these two proteins function during distinct stages of T lymphocyte development.
Genetic studies using mice lacking Bcl-2 or conditionally lacking Bcl-x have demonstrated that these anti-apoptotic proteins promote the survival of T lymphocytes at different stages. Deletion of Bcl-2 results in rapid disappearance of naïve T cells (14–16) while deletion of Bcl-x impairs the survival of DP thymocytes (17–19). Although Bcl-xL is upregulated in activated T cells, our recent data show that the development of effector T cells is not impaired in Bcl-xL conditional knockout mice (19), suggesting that other anti-apoptotic proteins may promote the survival of activated T cells. A likely candidate for this function is Mcl-1. Mcl-1 was initially discovered as a gene induced early during differentiation of the myeloid cell line ML-1 (20). Genetic studies have demonstrated that Mcl-1 functions as an important anti-apoptotic protein in several different cell types. Deletion of Mcl-1 in mice leads to embryonic lethality owing to a failure of implantation of the blastocyst in the uterus (21). Conditional deletion studies reveal that Mcl-1 is required for early lymphoid development and naïve T cell survival (22). Furthermore, Mcl-1 also promotes the survival of neutrophils and hematopoietic stem cells (23, 24).
Although it is clear that Mcl-1 is an important molecule for T cell survival, many aspects of its role remain to be addressed. In the present study, we focused on two major questions: what is the expression pattern of Mcl-1 in the different T cell populations and at what stage of T cell development it is essential for cell survival. To answer these questions, we have developed a flow cytometry-based assay for Mcl-1 expression and generated novel genetic mouse models. Our results demonstrate that unlike Bcl-2 and Bcl-XL, Mcl-1 is expressed throughout T cell development and is upregulated by TCR stimulation. Conditional deletion of Mcl-1 resulted in impaired cell survival not only of DN thymocytes and naïve T cell as previously reported, but also of SP thymocytes and activated T cells (22). Although Mcl-1 was not essential for the survival of DP thymocytes, the combined deletion of Mcl-1 and Bcl-XL resulted in a dramatic reduction of the size of this population in vivo. Taken together, our data demonstrates that Mcl-1 plays a critical role for the survival of T cells throughout their development and functions.
Materials and methods
Mice
Mice containing conditional Mcl-1f/f or Bcl-x f/f alleles were generated in our laboratory as previously described (19, 23). Mcl-1 f/f and/or Bcl-x f/f mice were bred with Lck-Cre (25) (The Jackson Laboratory, Bar Harbor, MI), CD4-Cre (26) (Taconic Farms, Germantown, NY), and ER-Cre (27) mice. The phenotype of the Mcl-1f/f mice is indistinguishable from those of wild-type (Mcl-1+/+) C57BL/6 x 129 mice and these mice were used as controls throughout the study. For some of the experiments, Mcl-1f/f CD4Cre mice were backcrossed to C57BL/6 for 7 generations. Bcl-2 Tg mice (28) were provided by Dr. Motonari Kondo at Duke University. All mice were used at 6–10 weeks of age. Animal usage was conducted according to protocols approved by the Duke University Institutional Animal Care and Use Committee.
Flow cytometry
Splenocytes, thymocytes, and lymph node cells were obtained by disrupting the respective organs with a syringe plunger. Bone marrow cells were flushed out of the femurs and tibiae. This was followed by filtering through 90 μm nylon mesh (SEFAR Filtration, Depew, NY) to generate single cell suspensions. Single cell suspensions of thymus, spleen, lymph nodes, and BM were lysed of RBC by treatment with ACK buffer. FcRγ-receptor binding was blocked with 2.4G2 hybridoma supernatant. The following antibodies were used for cell surface staining: CD4, CD8, CD3, CD44, CD25, TCRβ, CD62L, CD69, Qa-2, B220, CD43, CD11b, GD11c, and Gr-1 conjugated to FITC, PE, PE/cy5, PE/cy7, APC, APC/cy7, or APC/Alexa Fluor 750 from Pharmingen (San Diego, CA), Biolegend (San Diego, CA), or eBioscience (San Diego, CA). The cells were resuspended in FACS buffer containing propidium iodide (PI) (Sigma, St. Louis, MO). Data were collected on a FACScan, FACStarPLUS, or FACSVantage SE DiVa flow cytometer (BD Biosciences, San Jose, CA) and analyzed using CellQuest software (BD Biosciences, CA). Apoptosis was measured by Annexin V and 7-aminoactinomycin D (7-AAD) staining using the Annexin V-PE kit (BD Pharmingen) according to the manufacturer’s instructions.
Intracellular staining for Mcl-1 was performed by first staining for cell surface markers following the above protocol. The cells were then washed twice and fixed with 2% paraformaldehyde (Sigma) in PBS for 20 min on ice. Permeabilization was achieved by incubating the cells in 0.1% Saponin (Sigma) in FACS buffer containing 5% donkey serum for 20 min on ice. Mcl-1 was detected with polyclonal rabbit anti-Mcl-1 antibody (Rockland Immunochemicals, Gilbertsville, PA) at 0.1 μg/106 cells in permeabilization buffer for 1 h on ice. Normal rabbit immunoglobulin (Sigma) was used as an isotype control. A secondary donkey anti-rabbit-FITC antibody (Jackson Immunoresearch) was used at 1:100 in permeabilization buffer for 20 min on ice for final detection.
Western blotting
5×106 cells were lysed with 100μl 1xSDS sample buffer. Equal amounts of protein (20 μg) were separated on 10% polyacrylamide gel and transferred to PVDF membrane (PerkinElmer, Wellesley, MA) for chemiluminescent detection. The membranes were blocked for 1 h at room temperature with 5% non-fat milk (Carnation, Solon, OH) in 0.02% Tween 20 (EMD Biosciences) in PBS (PBS-T) followed by overnight incubation at 4°C with the primary antibody diluted in 3% BSA (Sigma) in PBS-T. The primary antibodies were rabbit anti-Mcl-1 used at 1:10,000 dilution (Rockland Immunochemicals). Rabbit anti-ERK2 and goat anti-actin were used at 1:1000 dilution (Santa Cruz Biotechnology). The membranes were incubated with the secondary antibodies for 1–3 h at room temperature. The secondary antibodies were anti-rabbit-HRP or anti-goat-HRP (Santa Cruz Biotechnology) used at 1:10,000 dilution. Proteins were detected with West Pico Chemiluminescence substrate (Pierce, Rockford, IL) for 5 min at room temperature. The membranes were exposed on Sterling high speed X-ray film (BioWorld, Dublin, OH). For fluorescent Western blots, PVDF-FL membranes (Millipore) were used. After primary antibody incubation, membranes were incubated in anti-rabbit Alexa-Fluor 680 (Molecular Probes, Invitrogen, Carlsbad, CA) or anti-goat IRDye 800 (Rockland Immunochemicals) at 1:5,000 dilution for 1 h at room temperature. Proteins were detected using the Odyssey system (LI-COR, Lincoln, NE).
Cell stimulation and death assays
Total thymocytes were lysed of RBC and 5×105 cells were cultured in 200 μl complete RPMI-1640 (Gibco) medium for different periods of time in the presence of 2 ng/ml PMA, 20 ng/ml ionomycin, or 1 nM dexamethsone (all from Sigma). FACS sorted DP thymocytes were cultured at 1.5×105 cells in 200 μl complete medium for 24 or 48 h in the presence of 2 ng/ml PMA, 1 nM dexamethasone, or 0.6 μM Tunicamycin (Sigma), or plate-bound anti-CD3 (2C11) and anti-CD28 (BioLegend).
Mcl-1f/f-ER-Cre T cells were purified from total splenocytes using the EasySep T cell enrichment kit (Stem Cell Technologies, Vancouver, BC, Canada) and cultured at 106 cells/ml for different periods of time in RPMI-1640 complete medium. The cells were stimulated with plate-bound anti-CD3 (2C11) and anti-CD28 antibodies or 1 ng/ml IL-7. To induce deletion of Mcl-1 in vitro, Mcl-1f/fER-Cre T cells were treated with 0.2 μM 4-hydroxytamoxifen (4-OHT) (Sigma) dissolved in ethanol. Cell death was evaluated by PI staining.
Statistical analysis
Statistical analysis was performed using unpaired two-tailed Student’s t test using the GraphPad Prizm software (GraphPad, San Diego, CA).
Results
Expression of Mcl-1 at different stages of T cell development
The expression pattern of Mcl-1 was previously examined by RT-PCR and Western blot analysis at the total population level (22). This study showed that both T and B cells express Mcl-1. However, it was not known whether the expression level of Mcl-1 protein in T lymphocytes is differentially regulated throughout T cell development. To test this, we developed a flow cytometric assay to measure the intracellular levels of Mcl-1. We used Mcl-1-deficient T cell populations wherever possible as negative controls (DP, CD4+ SP, and CD8+ SP thymocytes from Mcl-1f/fCD4-Cre mice as shown in Figure 1A). The fluorescence intensity of Mcl-1-deficient T cells stained with anti-Mcl-1 antibody was similar to the background staining seen in cells stained with an isotype-matched control antibody (Figure 1A). As the degree of background staining varied in the different T cell populations, we used the ratio of the fluorescence intensity of Mcl-1 staining to that of the background staining to represent the expression levels of Mcl-1 (Figure 1).
Figure 1. Mcl-1 expression in different T cell populations as assessed by intracellular staining.

A. Expression of Mcl-1 (thick lines) in double-negative CD4−CD8−CD3− (DN), double-positive CD4+CD8+ (DP), CD4 single-positive CD4+CD8−TCRβhi (CD4SP), CD8 single-positive CD4−CD8+TCRβhi (CD8SP), immature single-positive CD4−CD8+TCRβ − (ISP), and positively selected DP CD4+CD8+CD69+TCRβhi thymocytes as assessed by FACS. B. Expression of Mcl-1 (thick lines) in different DN subpopulations: DN1 – CD44+CD25− DN2 – CD44+CD25+; DN3 – CD44−CD25+; DN4 – CD44−CD25− as assessed by FACS. C. Expression of Mcl-1 (thick lines) in peripheral T cells: naïve (CD44−CD62Lhi) CD4+ or CD8+ T cells; activated/effector memory (CD44+CD62Llo) CD4+ or CD8+ T cells and central memory (CD44+CD62Lhi) CD8+ T cells as assessed by FACS. (A–C) The shaded histograms depict isotype control staining. For DP, CD4SP, CD8SP, and positively selected DP cells the thin lines represent staining for Mcl-1 in these cells from Mcl-1f/fCD4-Cre mice. The numbers represent the ratios of the fluorescence intensity of Mcl-1 staining divided by the fluorescent intensity of the isotype control staining. Data are representative of four individual experiments.
Mcl-1 expression levels were similar in different populations of developing thymocytes including DN, DP, CD4+ or CD8+ SP (Figure 1A). Thymocytes at the transitional immature single positive (ISP) or post-positive selection stage (TCRβhiCD69+ DP) also express similar levels of Mcl-1 (Figure 1A). Within the DN compartment, Mcl-1 expression was the highest in DN1 cells while thymocytes at the DN2-DN4 stages expressed slightly lower levels of Mcl-1 (Figure 1B). Mcl-1 protein was also detected in different populations of peripheral T cells (Figure 1C). Naïve CD4+ and CD8+ (CD62Lhi CD44+) T cells express levels of Mcl-1 largely comparable to those detected in memory CD4+ (CD62Llo CD44+), effector memory CD8+ (CD62Llo CD44+), and central memory CD8+ (CD62Lhi CD44+) T cells (Figure 1C). Taken together, these data demonstrate that Mcl-1 protein is constitutively expressed by T lymphocytes at all developmental stages.
Thymocyte maturation from DN2/3 to DN4 stage depends on Mcl-1
A previous report using conditional knockout mice has demonstrated that Mcl-1 plays a critical role in the survival of DN thymocytes and naïve T lymphocytes (22). Given that Mcl-1 is expressed at all stages of T cell development, it is important to know whether Mcl-1 also promotes the survival of other T cell populations. For example, it is unknown whether Mcl-1 is required for the survival of SP thymocytes and activated effector cells, which also express other anti-apoptotic proteins, including Bcl-2 and Bcl-xL., To address this, we generated mice lacking Mcl-1 in Tlymphocytes by crossing Mcl-1f/f mice that were previously generated in our laboratory (23) to a transgenic line expressing the Cre recombinase under control of the proximal Lck promoter (25). The Lck promoter drives Cre expression in early stages of T cell development, starting around the DN1-DN2 stage. Thus, Lck-Cre-induced deletion of Mcl-1 will allow the assessment of its role in the early stages of T cell development. Consistent with previously published results (22), thymic cellularity was greatly reduced in Mcl-1f/fLck-Cre mice, as thymuses from these mice contained only ~5% of the number of cells observed in a control thymus (Figure 2A). FACS analysis revealed that the frequencies of DN and CD4+ SP thymocytes were relatively increased while the frequency of DP cells was decreased (Figure 2B). The total cell numbers of DP, CD4+ and CD8+ SP thymocytes from Mcl-1f/fLck-Cre mice were only 5–10% of the controls while the total number of DN thymocytes from Mcl-1f/fLck-Cre mice was ~40% of the controls (Figure 2A and 2B). When the DN compartment of Mcl-1f/fLck-Cre mice was analyzed, the frequencies of DN2/3 thymocytes were increased while the frequency of DN4 thymocytes was decreased (Figure 2B), suggesting a developmental blockade between the DN2/3 and DN4 stages. A likely explanation for this observation is that Mcl-1 promotes the survival of thymocytes undergoing the DN2/3 to DN4 transition. To test this, we examined the apoptotic rates of DN thymocytes from Mcl-1f/fLck-Cre and control mice. As shown in Figure 2C, the apoptotic rate of DN thymocytes from Mcl-1f/fLck-Cre mice was increased by ~2-fold (Figure 2C).
Figure 2. Blockade of DN thymocyte development in Mcl-1f/fLck-Cre mice.
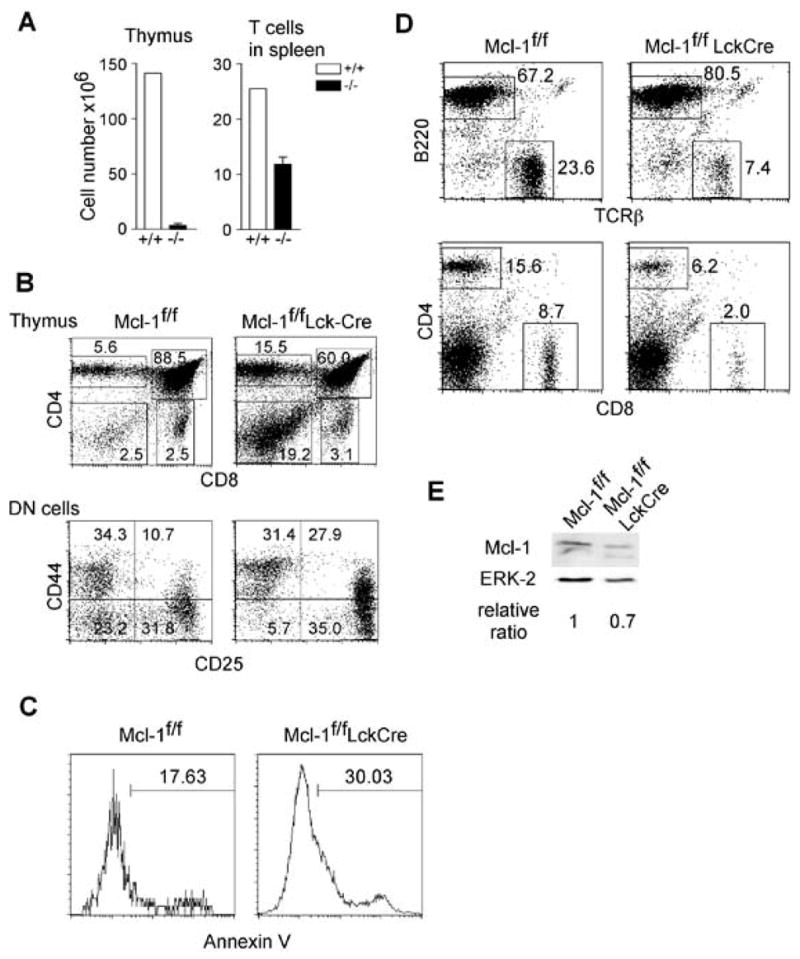
A. Total numbers of thymocytes and splenic T cells in Mcl-1f/fLck-Cre and control mice (n=3). Data are presented as mean+SD. B. FACS profiles of the thymus of Mcl-1f/fLck-Cre mice. The upper panels show total thymocytes and the lower panels show pre-gated CD3−CD4−CD8− DN cells. C. Apoptosis in the DN compartment of Mcl-1f/fLck-Cre mice as measured by Annexin V staining ex vivo. D. FACS profiles of the spleens of Mcl-1f/fLck-Cre and control mice. E. Mcl-1 protein expression in purified T cells from Mcl-1f/fLck-Cre and control mice as determined by Western blot. ERK-2 serves as a loading control.
We then examined the mature T cell compartment in Mcl-1f/fLck-Cre mice. The number of T cells in the spleen of Mcl-1f/fLck-Cre mice was 40–50% of that in control mice (Figure 2A and 2D). Given the dramatically reduced thymocyte number in Mcl-1f/fLck-Cre mice, it was surprising to observe this substantial number of mature T cells. These cells could have been derived through an Mcl-1-independent pathway or have escaped deletion of Mcl-1. To distinguish between these two possibilities, we purified mature T cells from the spleen of Mcl-1f/fLck-Cre mice and assessed them for Mcl-1 expression. Mature T cells in Mcl-1f/fLck-Cre mice expressed nearly the same level of Mcl-1 as control cells (Figure 2E), suggesting that the surviving T cells in the periphery had escaped deletion. Together, these results suggest that the requirement for Mcl-1 in the early stages of T cells development is essential for their further maturation.
Impaired SP thymocyte compartment in Mcl-1f/fCD4-Cre mice
The essential role of Mcl-1 in early stages of thymocyte development precluded our analysis of its function in thymocytes at later stages as mature T cells in Mcl-1f/fLck-Cre mice largely escaped Mcl-1 deletion. To circumvent this problem, we crossed Mcl-1 mice to another transgenic line expressing the Cre recombinase under control of the CD4 promoter (26). The CD4 promoter lacking the silencer element that restricts expression in CD8+ T cells drives Cre expression in all T cells from the onset of CD4 expression. Unlike Lck-driven Cre expression, CD4 promoter-driven Cre expression does not begin until the DN4 stage (26), bypassing the DN2/3 to DN4 transition.
In contrast to the severely reduced thymic cellularity observed in Mcl-1f/fLck-Cre mice, the number of total thymocytes in Mcl-1f/fCD4-Cre was comparable to that of control mice (Figure 3A). FACS analysis revealed that while the frequency of DN cells in Mcl-1f/fCD4-Cre mice was similar to that of control mice, there was a slight increase in the frequency of DP cells and a substantial decrease in the frequencies of CD4+ and CD8+ SP cells (Figure 3B). Mcl-1 appears to be especially critical for the development of CD8 SP thymocytes, as the frequency of CD8+ SP cells was reduced by >80% (Figure 3B). Furthermore, the majority of the CD8+CD4+ thymocytes did not express high levels TCRβ (Figure 3B), indicating that these cells are ISPs, an intermediate stage between DN and DP cells. Thus, the frequency of mature CD8+ SP thymocytes in Mcl-1f/fCD4-Cre mice was <0.2%, which represents a >10-fold reduction when compared to controls. We further assessed the maturation of SP thymocytes in Mcl-1f/fCD4-Cre mice using several other markers including CD69, HSA, CD62L, and Qa-2 (29). SP thymocytes that have just undergone positive selection are CD69hiHSAhiCD62LloQa-2lo. As they mature in the thymic medulla, SP cells up-regulate CD62L and Qa-2 and down-regulate CD69 and HSA (29). As shown in Figure 3C, both CD4+ and CD8+ SP thymocytes with a more mature phenotype (CD62LhiCD69loQa-2hi) were decreased in Mcl-1f/fCD4-Cre mice compared to controls, indicating an impaired mature SP thymocyte compartment.
Figure 3. Impaired SP compartment in Mcl-1f/fCD4-Cre mice.

A. Number of total thymocytes in Mcl-1f/fCD4-Cre and control mice (n=5). Shown are mean+SD. B. FACS profiles of thymocytes from Mcl-1f/fCD4-Cre and control mice on a mixed 129 x C57BL/6 background. The bottom plots represent TCRβ expression in CD8+ SP thymocytes. C. FACS profiles of CD4+ and CD8+ SP thymocytes stained with maturation markers CD62L, CD69, and Qa2. Cells were gated on CD4+ or CD8+ SP thymocytes. D. Expression of Mcl-1 protein in total thymocytes from Mcl-1f/fCD4-Cre and control mice as detected by Western blot. Erk2 serves as a loading control. E. Apoptotic rates of DP, CD4+ SP, and CD8+ SP thymocytes from Mcl-1f/fCD4-Cre and control mice as assessed by Annexin V and 7-AAD staining. F. Number of total T cells in the spleen of Mcl-1f/fCD4-Cre and control mice. T cells were enumerated by FACS staining with anti-CD3 and cell counting (n=5). G. FACS profiles of the spleens of Mcl-1f/f CD4-Cre and control mice as assessed by CD4, CD8, B220, and TCRβ staining. The data are representative of 3–5 individual experiments.
We next determined the efficiency of Mcl-1 deletion in total thymocytes of Mcl-1f/fCD4-Cre mice. Western blot analysis demonstrated that >95% of Mcl-1 was deleted in the thymocytes of Mcl-1f/fCD4-Cre mice (Figure 3D). As CD4 promoter-driven Cre expression starts at the DN4 stage, this deletion efficiency suggests that most of the thymocytes in Mcl-1f/fCD4-Cre mice have successfully deleted the Mcl-1 gene. Indeed, FACS analysis of intracellular Mcl-1 expression in DP, CD4+ SP, and CD8+ SP thymocytes from Mcl-1f/fCD4-Cre mice demonstrated that Mcl-1 was completely absent in these cells (Figure 1A).
We then examined the apoptotic rates among different thymocyte populations in Mcl-1f/fCD4-Cre mice. The frequencies of apoptotic DP or CD4+ SP thymocytes in Mcl-1f/fCD4-Cre mice were largely comparable to those of control thymocytes (Figure 3E). In contrast, the apoptotic rates of CD8+ SP thymocytes in the mutant mice were increased. That no increase in the apoptotic rate of CD4 thymocytes was observed in Mcl-1f/fCD4-Cre mice may be due to the fact that it is difficult to detect apoptotic cells in the thymus as the dead cells are cleared very rapidly by resident macrophages, and only dramatically enhanced apoptosis can be detected using this approach.
Lastly, we examined the peripheral T cell compartment in Mcl-1f/fCD4-Cre mice. The number of mature T cells in the spleen of Mcl-1f/fCD4-Cre mice was <10% of that observed in control mice (Figure 3F). FACS analysis showed that very few CD4 and CD8 T cells were detected in Mcl-1f/fCD4-Cre mice (Figure 3G). This dramatic reduction in the peripheral T cell compartment is consistent with the above observation that development of both CD4+ SP and CD8+ SP thymocytes depends on Mcl-1. Thus, Mcl-1 plays a critical role in the development of SP thymocytes.
Mcl-1 regulates DP thymocyte survival in vitro
The normal DP thymocyte compartment observed in Mcl-1f/fCD4-Cre mice suggests that Mcl-1 is dispensable for in vivo DP thymocyte survival. Given that Mcl-1 is expressed in DP thymocytes, we wondered whether Mcl-1 regulates DP thymocyte survival in vitro, as the in vitro environment may provide fewer pro-survival signals than the in vivo environment. We thus examined the survival of Mcl-1-deficient thymocytes in vitro in the presence or absence of apoptotic stimuli. We used several well-characterized chemical agents that induce thymocyte apoptosis. Some of them, such as Dexamethasone, Ionomycin and Tunicamycin are dependent on the BH3-only protein Bim, while phorbol myristate acetate (PMA) requires the BH3-only protein Puma for death induction (30, 31). In vitro cultured total thymocytes from Mcl-1f/fCD4-Cre mice underwent apoptosis at a rate dramatically higher than control cells (Figure 4A). After 2 days in culture in the absence of any stimulus, <5% of Mcl-1-deficient thymocytes survived while ~40% of control cells did (Figure 4A). As a positive control, thymocytes from Bcl-2 Tg mice survived better than wild type thymocytes (Figure 4A). Addition of PMA or ionomycin did not obviously change the death kinetics of these cells, while addition of dexamethasone dramatically facilitated the death of Mcl-1-deficient total thymocytes (Figure 4A).
Figure 4. Impaired survival of Mcl-1-deficient DP thymocytes in vitro.

A. Survival of Mcl-1-deficient thymocytes in response to different death-inducing stimuli compared to wild type and Bcl-2 transgenic cells. B. Survival of sorted DP Mcl-1-deficient thymocytes in vitro in response to different death-inducing stimuli. Cells were treated with dexamethasone, PMA, or tunicamycin for 24 h. Data are representative of three individual experiments.
To further confirm that in vitro cultured DP thymocytes depend on Mcl-1 for their survival, we sorted DP thymocytes from Mcl-1f/fCD4-Cre and control mice and cultured them in vitro. The survival of Mcl-1-deficient DP thymocytes cultured with or without stimulation was significantly decreased when compared to control thymocytes (Figure 4B). These data demonstrate that Mcl-1 is required for the survival of DP thymocytes in vitro and suggests that Mcl-1-deficient thymocytes are protected by other anti-apoptotic factor(s) in vivo.
DP thymocyte survival in vivo depends on both Mcl-1 and Bcl-xL
A likely candidate anti-apoptotic protein promoting DP thymocyte survival in vivo in Mcl-1f/fCD4-Cre mice is Bcl-xL as Bcl-xL but not Bcl-2 is highly expressed in DP thymocytes (6–9). Our previous data using Bcl-xL conditional knockout mice demonstrate that the DP thymocyte compartment is slightly reduced in the absence of -Bcl-x, and their in vitro survival of DP thymocytes depends on Bcl-x (19). These results suggest that Bcl-xL and Mcl-1 may function redundantly to promote DP thymocyte survival in vivo. To test this, we generated conditional knockout mice lacking both Mcl-1 and Bcl-x in T cells by crossing Mcl-1f/fand Bcl-xf/f mice to CD4-Cre mice. Analysis of CD4/CD8 profiles of Mcl-1f/f Bcl-xf/f CD4-Cre mice demonstrate that the frequency of DP thymocytes in these mice was reduced, as the DP compartment comprised ~85% of thymocytes in control or single knockout mice, but only ~45% of thymocytes in double knockout mice. (Figure 5A). Importantly, the absolute number of DP thymocytes in the double conditional knockout mice was only ~10% of that in the controls (Figure 5B). Deletion of either Mcl-1 or Bcl-x in thymocytes significantly impaired the development of CD8+ SP but not CD4+ SP thymocytes (Figure 5B and 5C). In contrast, deletion of both Mcl-1 and Bcl-x almost completely abolished the CD4+ and CD8+ SP compartments as judged by their number and the frequency of TCRβ+ cells (Figure 5B and 5C). These results unequivocally demonstrate that DP thymocyte survival is promoted by both Mcl-1 and Bcl-xL in vivo.
Figure 5. DP thymocyte survival in vivo depends on both Mcl-1 and Bcl-xL.

A. FACS profiles of thymocytes from mice conditionally lacking Mcl-1, Bcl-x, or both Mcl-1 and Bcl-xL. These mice have been crossed to C57BL/6 background for 7 generations. Numbers indicate the frequency of cells in each subset. B. Numbers of total thymocytes, DP, CD4+ SP, CD8+ SP, and ISP cells from mice conditionally lacking Mcl-1, Bcl-x or both. Shown are mean+SD (n=3–5). C. TCRβ expression in CD4+ SP or CD8+ SP thymocytes from mice conditionally lacking Mcl-1, Bcl-x, or both Mcl-1 and Bcl-xL. Numbers indicate the frequency of TCRβ+ cells within the SP thymocyte compartment.
Mcl-1 is required for the survival of activated T cells
Previous data have shown that Mcl-1 elicited in response to cytokine signaling is required for the survival of naïve T cells (22). Among T cell survival cytokines, IL-7 induces the highest level of Mcl-1 expression in naïve T cells (22). One important question remaining to be addressed is whether Mcl-1 expression is regulated by TCR engagement and whether Mcl-1 plays a role in the survival of activated T cells. To address these issues, we stimulated wild type T cells with anti-CD3 and examined Mcl-1 expression by Western blot. Cell lysate from IL-7-stimulated naïve T cells was used as a positive control. Consistent with the previous result (22), Mcl-1 expression was increased in naïve T cells stimulated with IL-7 (Figure 6A). Interestingly, anti-CD3 stimulation strongly upregulated Mcl-1 expression in T cells, and the extent of anti-CD3-induced upregulation was greater than that induced by IL-7 after 48 hr stimulation (Figure 6A). To further examine the kinetics of anti-CD3 induced Mcl-1 expression in T cells, we stimulated splenic T cells with anti-CD3 or IL-7 for 6–12 hr. Short duration stimulation with anti-CD3 or IL-7 also upregulated Mcl-1 expression in T cells (Figure 6B). The extent of Mcl-1 upregulation in T cells induced by IL-7 was higher than that induced by anti-CD3 (Figure 6B). These data suggest that other factors might be involved in anti-CD3 induced Mcl-1 upregulation at late time point. Alternatively, anti-CD3 induced Mcl-1 upregulation may require longer time to reach its peak level.
Figure 6. Survival of activated T cells depends on Mcl-1.

A. Mcl-1 expression in naïve (CD44−CD62Lhi) and memory (CD44+CD62Llo) T cells as well as in T cells after 48 h of stimulation with 10 ng/ml IL-7 or 5 μg/ml anti-CD3 as determined by Western blot. Actin serves as a loading control. B. Mcl-1 expression in T cells after 6–12 h of stimulation with 10 ng/ml IL-7 or 5 μg/ml anti-CD3 as determined by Western blot. Actin serves as a loading control. Numbers represent normalized expression levels of Mcl-1. C. Survival of purified Mcl-1f/f ERCre T cells after 48 h in response to cytokine withdrawal, 1 ng/ml IL-7, or plate-bound anti-CD3 + anti-CD28. Cells were treated with 0.2 μM 4-OHT or vehicle alone. Data mean±SD from a single experiment performed in triplicate are representative of 3 independent experiments.
To test whether Mcl-1 plays a role in the survival of activated T cells, we generated an inducible deletion system by crossing Mcl-1f/f mice to the ER-Cre line (27). ER-Cre mice express the Cre recombinase fused to a mutant form of the estrogen ligand-binding domain inserted into the ROSA26 locus. The mutation in the estrogen ligand-binding domain makes it insensitive to endogenous estrogen, but capable of activation by the drug tamoxifen. We treated purified T cells from Mcl-1f/fER-Cre and control mice (Mcl-1f/f) in vitro with 0.2 μM 4-OH tamoxifen (4-OHT), which did not demonstrate significant toxicity in vitro. A two-day treatment resulted in significantly reduced recovery of live cells (p<0.001) from Mcl-1f/fER-Cre T cell culture (Figure 6C). Importantly, T cell activation with anti-CD3/CD28 resulted in accumulation of live cells in the control cultures, but the cell numbers recovered from Mcl-1f/fER-Cre cultures treated with 4-OHT remained significantly lower (p<0.001) (Figure 6C). Similarly, IL-7 treatment did not rescue 4-OHT-treated Mcl-1f/fER-Cre T cells from death (p<0.05) (Figure 6C). As activated T cells strongly upregulate Bcl-xL expression, these results suggest that the upregulation of Bcl-xL is not sufficient to protect activated T cell from death, and that Mcl-1 plays a critical role in the survival of activated T cells.
Discussion
Our studies demonstrate that Mcl-1 promotes T lymphocyte survival at multiple stages. In contrast to the anti-apoptotic roles of Bcl-2 and Bcl-xL at specific T cell developmental stages, Mcl-1 appears to be a universal regulator of T cell survival. Genetic studies have established an anti-apoptotic role for Bcl-2 in naïve T cells and Bcl-xL in DP thymocytes (14–19). Our data shown here, together with a previous report (22), have established that Mcl-1 promotes the survival of DN, DP, SP, naïve, and activated T cells.
Our results confirm the previous finding that DN thymocyte development depends on Mcl-1 as deletion of Mcl-1 early in T cell development results in a blockade at the DN2/3 to DN4 transition. The major survival factors for T cells at this stage are IL-7R and pre-TCR signaling. IL-7R signaling is known to induce both Bcl-2 and Mcl-1 (22, 28, 32). Deletion of IL-7 or IL-7R components blocks thymocyte development at the DN2 stage, while deletion or mutation of pre-TCR components impairs developmental progression at the DN3 stage (33–36). The phenotype of Mcl-1-deficient mice suggests that both IL-7R and pre-TCR might rely on Mcl-1 to promote DN cell survival.
We also show that the survival of SP thymocytes depends on Mcl-1. Little is known about the molecular regulation of SP thymocyte survival in vivo. The severe impairment of the SP thymocyte compartment in Mcl-1f/fCD4-Cre mice suggests that the function of Mcl-1 cannot be compensated by Bcl-2 despite its expression in SP thymocytes. This suggests that perhaps these two proteins operate in distinct molecular pathways. Most likely the difference between the two anti-apoptotic molecules lies in their interaction partners (37). For example, only Mcl-1 but not Bcl-2 can interact with the pro-apoptotic molecule Bak. Conversely, only Bcl-2 but not Mcl-1 interacts with BAD.
Similar to DN and SP thymocytes, naïve T cell express high levels of Bcl-2, but are also critically dependent on Mcl-1 for their survival. This was first demonstrated by Opferman et al. using an inducible Cre driven by interferon-response elements (22). However, this system has a caveat in that it requires induction of high levels of IFNα/β that impact T cell survival (38). We used the ER-Cre inducible recombinase to show that naïve T cells require Mcl-1 for their survival. Naïve T cell survival requires IL-7 and TCR/MHC-peptide contact (39). As both Mcl-1 and Bcl-2 can be upregulated by IL-7 signaling while Bcl-2 expression is downregulated upon TCR engagement, our data suggest that TCR/MHC-peptide engagement may mediate its survival signal through Mcl-1. It is likely, however, that Mcl-1 and Bcl-2 operate in separate molecular pathways to ensure the survival of the cell, because loss of either protein cannot be compensated by the remaining one.
An interesting finding from our study is the anti-apoptotic role of Mcl-1 in DP thymocytes and activated T cells, in which the expression of Mcl-1 parallels that of Bcl-xL but not Bcl-2. Although DP thymocytes lacking either Mcl-1 or Bcl-x display some degree of survival defects both in vitro and in vivo, the deletion of both Mcl-1 and Bcl-x results in a drastic reduction of the DP thymocyte compartment and a nearly complete block of the development of SP thymocytes. Thus, Mcl-1 and Bcl-xL can largely compensate for each other in the DP compartment. In contrast, activated T cells from Mcl-1f/f ER-Cre mice undergo apoptosis, suggesting that Bcl-xL upregulation is not sufficient to prevent cell death induced by Mcl-1 deletion. In contrast, Bcl-xL is dispensable for the survival of activated T cells (19). As Bcl-xL functions similarly to Mcl-1 in terms of their interaction with the pro-apoptotic protein Bak (37), the death of activated T cells lacking Mcl-1 may reflect a dose-dependent effect. Alternatively, Mcl-1 may have a unique role that cannot be compensated by Bcl-xL. Further studies are required to establish the in vivo mechanisms by which Mcl-1 exerts its anti-apoptotic function throughout T cell development.
Acknowledgments
This work was supported by NIH grants CA92123 and AI54683.
We thank Claire Gordy for critically reading the manuscript.
References
- 1.Goldrath AW, Bevan MJ. Selecting and maintaining a diverse T-cell repertoire. Nature. 1999;402:255–262. doi: 10.1038/46218. [DOI] [PubMed] [Google Scholar]
- 2.Rathmell JC, Thompson CB. The central effectors of cell death in the immune system. Annu Rev Immunol. 1999;17:781–828. doi: 10.1146/annurev.immunol.17.1.781. [DOI] [PubMed] [Google Scholar]
- 3.Droin NM, Green DR. Role of Bcl-2 family members in immunity and disease. Biochim Biophys Acta. 2004;1644:179–188. doi: 10.1016/j.bbamcr.2003.10.011. [DOI] [PubMed] [Google Scholar]
- 4.Zhang N, Hartig H, Dzhagalov I, Draper D, He YW. The role of apoptosis in the development and function of T lymphocytes. Cell Res. 2005;15:749–769. doi: 10.1038/sj.cr.7290345. [DOI] [PubMed] [Google Scholar]
- 5.Mandal M, Borowski C, Palomero T, Ferrando AA, Oberdoerffer P, Meng F, Ruiz-Vela A, Ciofani M, Zuniga-Pflucker JC, Screpanti I, Look AT, Korsmeyer SJ, Rajewsky K, von Boehmer H, Aifantis I. The BCL2A1 gene as a pre-T cell receptor-induced regulator of thymocyte survival. J Exp Med. 2005;201:603–614. doi: 10.1084/jem.20041924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gratiot-Deans J, Ding L, Turka LA, Nunez G. bcl-2 proto-oncogene expression during human T cell development. Evidence for biphasic regulation. J Immunol. 1993;151:83–91. [PubMed] [Google Scholar]
- 7.Gratiot-Deans J, Merino R, Nunez G, Turka LA. Bcl-2 expression during T-cell development: early loss and late return occur at specific stages of commitment to differentiation and survival. Proc Natl Acad Sci U S A. 1994;91:10685–10689. doi: 10.1073/pnas.91.22.10685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Grillot DA, Merino R, Nunez G. Bcl-XL displays restricted distribution during T cell development and inhibits multiple forms of apoptosis but not clonal deletion in transgenic mice. J Exp Med. 1995;182:1973–1983. doi: 10.1084/jem.182.6.1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Veis DJ, Sentman CL, Bach EA, Korsmeyer SJ. Expression of the Bcl-2 protein in murine and human thymocytes and in peripheral T lymphocytes. J Immunol. 1993;151:2546–2554. [PubMed] [Google Scholar]
- 10.Grayson JM, Zajac AJ, Altman JD, Ahmed R. Cutting edge: increased expression of Bcl-2 in antigen-specific memory CD8+ T cells. J Immunol. 2000;164:3950–3954. doi: 10.4049/jimmunol.164.8.3950. [DOI] [PubMed] [Google Scholar]
- 11.Grayson JM, Murali-Krishna K, Altman JD, Ahmed R. Gene expression in antigen-specific CD8+ T cells during viral infection. J Immunol. 2001;166:795–799. doi: 10.4049/jimmunol.166.2.795. [DOI] [PubMed] [Google Scholar]
- 12.Hildeman DA, Zhu Y, Mitchell TC, Bouillet P, Strasser A, Kappler J, Marrack P. Activated T cell death in vivo mediated by proapoptotic bcl-2 family member bim. Immunity. 2002;16:759–767. doi: 10.1016/s1074-7613(02)00322-9. [DOI] [PubMed] [Google Scholar]
- 13.Boise LH, Minn AJ, Noel PJ, June CH, Accavitti MA, Lindsten T, Thompson CB. CD28 costimulation can promote T cell survival by enhancing the expression of Bcl-XL. Immunity. 1995;3:87–98. doi: 10.1016/1074-7613(95)90161-2. [DOI] [PubMed] [Google Scholar]
- 14.Veis DJ, Sorenson CM, Shutter JR, Korsmeyer SJ. Bcl-2-deficient mice demonstrate fulminant lymphoid apoptosis, polycystic kidneys, and hypopigmented hair. Cell. 1993;75:229–240. doi: 10.1016/0092-8674(93)80065-m. [DOI] [PubMed] [Google Scholar]
- 15.Nakayama K, Nakayama K, Negishi I, Kuida K, Shinkai Y, Louie MC, Fields LE, Lucas PJ, Stewart V, Alt FW, et al. Disappearance of the lymphoid system in Bcl-2 homozygous mutant chimeric mice. Science (New York, NY) 1993;261:1584–1588. doi: 10.1126/science.8372353. [DOI] [PubMed] [Google Scholar]
- 16.Wojciechowski S, Tripathi P, Bourdeau T, Acero L, Grimes HL, Katz JD, Finkelman FD, Hildeman DA. Bim/Bcl-2 balance is critical for maintaining naive and memory T cell homeostasis. J Exp Med. 2007;204:1665–1675. doi: 10.1084/jem.20070618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Motoyama N, Wang F, Roth KA, Sawa H, Nakayama K, Nakayama K, Negishi I, Senju S, Zhang Q, Fujii S, et al. Massive cell death of immature hematopoietic cells and neurons in Bcl-x-deficient mice. Science (New York, NY) 1995;267:1506–1510. doi: 10.1126/science.7878471. [DOI] [PubMed] [Google Scholar]
- 18.Ma A, Pena JC, Chang B, Margosian E, Davidson L, Alt FW, Thompson CB. Bclx regulates the survival of double-positive thymocytes. Proc Natl Acad Sci U S A. 1995;92:4763–4767. doi: 10.1073/pnas.92.11.4763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang N, He YW. The antiapoptotic protein Bcl-xL is dispensable for the development of effector and memory T lymphocytes. J Immunol. 2005;174:6967–6973. doi: 10.4049/jimmunol.174.11.6967. [DOI] [PubMed] [Google Scholar]
- 20.Kozopas KM, Yang T, Buchan HL, Zhou P, Craig RW. MCL1, a gene expressed in programmed myeloid cell differentiation, has sequence similarity to BCL2. Proc Natl Acad Sci U S A. 1993;90:3516–3520. doi: 10.1073/pnas.90.8.3516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rinkenberger JL, Horning S, Klocke B, Roth K, Korsmeyer SJ. Mcl-1 deficiency results in peri-implantation embryonic lethality. Genes Dev. 2000;14:23–27. [PMC free article] [PubMed] [Google Scholar]
- 22.Opferman JT, Letai A, Beard C, Sorcinelli MD, Ong CC, Korsmeyer SJ. Development and maintenance of B and T lymphocytes requires antiapoptotic MCL-1. Nature. 2003;426:671–676. doi: 10.1038/nature02067. [DOI] [PubMed] [Google Scholar]
- 23.Dzhagalov I, St John A, He YW. The antiapoptotic protein Mcl-1 is essential for the survival of neutrophils but not macrophages. Blood. 2007;109:1620–1626. doi: 10.1182/blood-2006-03-013771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Opferman JT, Iwasaki H, Ong CC, Suh H, Mizuno S, Akashi K, Korsmeyer SJ. Obligate role of anti-apoptotic MCL-1 in the survival of hematopoietic stem cells. Science (New York, NY) 2005;307:1101–1104. doi: 10.1126/science.1106114. [DOI] [PubMed] [Google Scholar]
- 25.Hennet T, Hagen FK, Tabak LA, Marth JD. T-cell-specific deletion of a polypeptide N-acetylgalactosaminyl-transferase gene by site-directed recombination. Proc Natl Acad Sci U S A. 1995;92:12070–12074. doi: 10.1073/pnas.92.26.12070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lee PP, Fitzpatrick DR, Beard C, Jessup HK, Lehar S, Makar KW, Perez-Melgosa M, Sweetser MT, Schlissel MS, Nguyen S, Cherry SR, Tsai JH, Tucker SM, Weaver WM, Kelso A, Jaenisch R, Wilson CB. A critical role for Dnmt1 and DNA methylation in T cell development, function, and survival. Immunity. 2001;15:763–774. doi: 10.1016/s1074-7613(01)00227-8. [DOI] [PubMed] [Google Scholar]
- 27.Shapiro-Shelef M, Lin KI, Savitsky D, Liao J, Calame K. Blimp-1 is required for maintenance of long-lived plasma cells in the bone marrow. J Exp Med. 2005;202:1471–1476. doi: 10.1084/jem.20051611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Akashi K, Kondo M, von Freeden-Jeffry U, Murray R, Weissman IL. Bcl-2 rescues T lymphopoiesis in interleukin-7 receptor-deficient mice. Cell. 1997;89:1033–1041. doi: 10.1016/s0092-8674(00)80291-3. [DOI] [PubMed] [Google Scholar]
- 29.McCaughtry TM, Wilken MS, Hogquist KA. Thymic emigration revisited. J Exp Med. 2007;204:2513–2520. doi: 10.1084/jem.20070601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bouillet P, Metcalf D, Huang DC, Tarlinton DM, Kay TW, Kontgen F, Adams JM, Strasser A. Proapoptotic Bcl-2 relative Bim required for certain apoptotic responses, leukocyte homeostasis, and to preclude autoimmunity. Science (New York, NY) 1999;286:1735–1738. doi: 10.1126/science.286.5445.1735. [DOI] [PubMed] [Google Scholar]
- 31.Villunger A, Michalak EM, Coultas L, Mullauer F, Bock G, Ausserlechner MJ, Adams JM, Strasser A. p53-and drug-induced apoptotic responses mediated by BH3-only proteins puma and noxa. Science (New York, NY) 2003;302:1036–1038. doi: 10.1126/science.1090072. [DOI] [PubMed] [Google Scholar]
- 32.von Freeden-Jeffry U, Solvason N, Howard M, Murray R. The earliest T lineage-committed cells depend on IL-7 for Bcl-2 expression and normal cell cycle progression. Immunity. 1997;7:147–154. doi: 10.1016/s1074-7613(00)80517-8. [DOI] [PubMed] [Google Scholar]
- 33.von Freeden-Jeffry U, Vieira P, Lucian LA, McNeil T, Burdach SE, Murray R. Lymphopenia in interleukin (IL)-7 gene-deleted mice identifies IL-7 as a nonredundant cytokine. J Exp Med. 1995;181:1519–1526. doi: 10.1084/jem.181.4.1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Peschon JJ, Morrissey PJ, Grabstein KH, Ramsdell FJ, Maraskovsky E, Gliniak BC, Park LS, Ziegler SF, Williams DE, Ware CB, Meyer JD, Davison BL. Early lymphocyte expansion is severely impaired in interleukin 7 receptor-deficient mice. J Exp Med. 1994;180:1955–1960. doi: 10.1084/jem.180.5.1955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Malissen M, Gillet A, Ardouin L, Bouvier G, Trucy J, Ferrier P, Vivier E, Malissen B. Altered T cell development in mice with a targeted mutation of the CD3-epsilon gene. Embo J. 1995;14:4641–4653. doi: 10.1002/j.1460-2075.1995.tb00146.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zhang W, Sommers CL, Burshtyn DN, Stebbins CC, DeJarnette JB, Trible RP, Grinberg A, Tsay HC, Jacobs HM, Kessler CM, Long EO, Love PE, Samelson LE. Essential role of LAT in T cell development. Immunity. 1999;10:323–332. doi: 10.1016/s1074-7613(00)80032-1. [DOI] [PubMed] [Google Scholar]
- 37.Willis SN, Chen L, Dewson G, Wei A, Naik E, Fletcher JI, Adams JM, Huang DC. Proapoptotic Bak is sequestered by Mcl-1 and Bcl-xL, but not Bcl-2, until displaced by BH3-only proteins. Genes Dev. 2005;19:1294–1305. doi: 10.1101/gad.1304105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tough DF, Borrow P, Sprent J. Induction of bystander T cell proliferation by viruses and type I interferon in vivo. Science (New York, NY) 1996;272:1947–1950. doi: 10.1126/science.272.5270.1947. [DOI] [PubMed] [Google Scholar]
- 39.Surh CD, Sprent J. Regulation of mature T cell homeostasis. Semin Immunol. 2005;17:183–191. doi: 10.1016/j.smim.2005.02.007. [DOI] [PubMed] [Google Scholar]