

Abstract
Development of suitable tools to assess enzyme activity directly from their complex cellular environment has a dramatic impact on understanding the functional roles of proteins as well as on the discovery of new drugs. In this study, a novel fluorescence-based chemosensor strategy for the direct readout of dipeptidase activities within intact living cells is described. Selective activity-based probes were designed to sense two important type II transmembrane serine proteases, Fibroblast activation protein (FAP) and Dipeptidyl peptidase IV (DPP-IV). These serine proteases have been implicated in diverse cellular activities, including blood coagulation, digestion, immune responses, wound healing, tumor growth, tumor invasion and metastasis. We here validated that Ac-GPGP-2SBPO and GPGP-2SBPO probes are excellent reporters of both proteolytic activities. Furthermore, the novel probes can differentiate between FAP and DPP-IV proteolytic activities in cellular assay. Potentially, this assay platform is immediately useful for novel drug discovery.
Keywords: Fibroblast activation protein, dipeptidyl peptidase IV, dipeptidyl peptidase, fluorogenic substrate, fluorescence
Introduction
Fibroblast activation protein (FAP; EC: 3.4.21.B28) and Dipeptidyl peptidase IV (DPP-IV; EC: 3.4.14.5) belong to a type II transmembrane serine protease family, and are capable of cleaving N-terminal dipeptides after a proline residue (1, 2). Structurally, both proteases have a short cytoplasmic tail, a transmembrane domain and a large C-terminal extracellular domain (ECD). The ECD domain consists of two characteristic sub-domains, a β-propeller sub-domain with several sites of N-linked glycosylation at the N-terminus and an α/β-hydrolase sub-domain with the catalytic triad of serine, aspartate and histidine residues at the C-terminus. The human FAP gene maps on chromosome 2 at 2q23 and the DPP-IV gene maps on the same chromosome at 2q24.3. Although FAP and DPP-IV share a high sequence identity and may have arisen by gene duplication, it was reported that each has distinct enzyme activity profile. Apart from having prolyl exopeptidase activity like its DPP-IV homologue, FAP also functions as a prolyl endopeptidase (3, 4) and has a collagenolytic activity capable of degrading gelatin and type I collagen (5, 6).
In terms of expression, FAP is selectively expressed on reactive stromal fibroblasts in over 90% of common human epithelial cancers, such as colorectal, breast and lung cancers (7). Expression of FAP is not detected in fibroblasts of benign epithelial tumors or normal adult tissues. High expression of FAP in tumors correlates with increased FAP activity relative to normal tissues (4, 8, 9). It has been suggested that FAP promotes tumor growth and proliferation (10). Currently, FAP inhibitor PT-100 (Val-boroPro) (11) is being tested in clinical trials to evaluate its efficacy to treat non-small cell lung and other cancers (12). In contrast to FAP, DPP-IV is ubiquitously expressed in normal epithelial cells and melanocytes. It was originally identified as a protein expressed on the surface of T cells and its expression is lost in various cancers (13-15). These suggest the involvement of DPP-IV in immunology, oncology, and other areas. DPP-IV has been recently validated as a therapeutic target for type 2 diabetics with the approval of the DPP-IV inhibitor sitagliptin phosphate (16). The proteolytic functions of these two proteins and their distinct expression distribution make them promising and attractive targets for new indications (8, 17-21).
Protease-targeted fluorogenic substrates using 7-amino-4-methylcoumarin (AMC) or 7-amino-4-trifluoromethyl coumarin (AFC) as a reporter have commonly been used for detecting protease activity in vitro (4-6). The fluorescence property of the coumarin reporters is masked by the peptide amide linkage. Upon proteolytic conversion of the probes, the reporters are released as fluorescent free dyes. This type of probes has been widely used in cell-free assays; however, it cannot be directly applied to complex cellular environments. Limitations of these probes are attributed to two major factors. First, high autofluorescence background can be caused by the illumination of cells. Second, the poor aqueous solubility of the fluorophores often required DMSO as a co-solvent, which could lead to reduced enzymatic activity (22).
To develop methods that allow more direct and reliable assessment of enzyme activity in vivo, a water-soluble and far-red fluorescence-based probe having emission maxima between 650 and 900 nm is preferred. We recently validated the use of a new water-soluble far-red fluorogenic dye containing a disulfonated benzo[a]phenoxazine (2SBPO) scaffold for enzyme sensing (23). 2SBPO conjugated to a GPGP tetrapeptide is weakly fluorescent but displays strong fluorescence emission at 670 nm upon cleavage by DPP-IV. Here we further demonstrate that the distinct proteolytic activities between FAP and DPP-IV could be assessed using pre-selected probes. The sensitivity and selectivity of these fluorescence-based chemosensors are sufficient to measure the prolyl-endopeptidase activities of FAP and the prolyl-exopeptidase activity of DPP-IV in living cells.
Materials and methods
Synthesis of the fluorescence-based chemosensors
The parent dye, 9-di-3-sulfonyl-propylaminobenzo[a]phenoxazonium perchlorate (2SBPO), was synthesized as previously described (24). Amino acids were purchased from Novabiochem (San Diego, CA). All peptide substrates were prepared by solution phase synthesis using N-Boc- protected amino acids and Ac-glycine for the acetyl-protected peptides using the same procedure as described for GPGP-2SBPO (23). The peptide substrates were purified on a C-18 Sep-Pak cartridge. Electrospray mass spectra were obtained on a Platform LCT mass spectrometer (Micromass, Beverly, MA) and MALDI mass spectra were measured on a Voyager linear mass spectrometer (Applied Biosystems; Framingham, MA). GPGP-2SBPO: HRMS (ES+/TOF) calcd for C36H44N7O11S2 (M)+: 814.2540, found 814.2538; Ac-GPGP-2SBPO: HRMS (ES+/TOF) calcd for C38H46N7O12S2 (M)+: 856.2640, found 856.2530; APAP-2SBPO: HRMS (ES+/TOF) calcd for C38H48N7O11S2 (M)+: 842.2848, found 842.2856. Ac-GP-2SBPO: MS (MALDI-TOF) calcd for C31H36N5O10S2 (M)+: 702.2, found 701.9; TSGP-2SBPO: MS (MALDI-TOF) calcd for C36H45N6O14S2 (M)+: 702.2, found 701.9.
Cell lines
HT-29, human colon cancer cell line was obtained from American Type Culture Collection (Rockville, MD). To detect the FAP activity in culture assay, human embryonic kidney cell line, HEK293, stably transfected with full length wild-type murine FAP (HEK-FAP) were used and HEK293 cells stably transfected with empty vector alone (HEK-vector) were used as a control cell line (9). For recombinant murine FAP preparation, HEK293 stably transfected with murine FAP-ECD cDNA was used (HEK- FAP-ECD) (9, 10). All cell lines were maintained in DMEM medium with 10% FBS.
In vitro and cell-based enzymatic assay
Recombinant murine DPP-IV was purchased from R&D Systems (Minneapolis, MN), while murine FAP was purified from HEK-FAP-ECD cells as previously described (9, 10). Purification of the FAP protein was confirmed by SDS-PAGE electrophoresis and staining with Gelcode blue stain reagent (Pierce, Rockford, IL). Concentration of the FAP protein was determined by using a BCA protein assay kit (Pierce, Rockford, IL).
All of the enzymatic assays were performed in triplicate on 96-well assay plates (Corning Inc. NY). Various substrates at 50 μM were tested in vitro with 10 nM of the respective recombinant murine DPP-IV and FAP enzymes under physiological conditions. A total of 200 μL of 10 mM PBS (pH 7.4) with substrate and enzyme, or with substrate alone as control, was incubated in the 96-well plate at 37°C for 2 hours. Fluorescence emission at 670 nm and excitation at 630 nm were measured by a fully modular monochromator-based plate reader (Safire2, Tecan, San Jose, CA). For cell-based enzymatic assays, the cells were first seeded at 50,000 cells/well in the 96-well assay plate. A day later, the serum-containing medium was removed and changed to 10 mM PBS (pH7.4) with 20 μM of the substrate. Cells were then incubated with substrates at 37°C for 3 to 5 hours prior to fluorescence measurement.
Western blot analyses
HEK-vector, HEK-FAP and HT-29 cells were lysed with M-PER mammalian protein extraction reagent (Pierce, Rockford, IL). Cell debris was removed by centrifugation and clear cell lysates were transferred to new microfuge tubes. The protein concentrations were determined by using Bradford reagent (Sigma-Aldrich, St, Louis, MO). The cell lysates (50 μg) were loaded onto 10% SDS-PAGE gels and transferred to Immun-blot PVDF membranes (Bio-Rad, Hercules, CA). Anti-murine FAP monoclonal antibody (6E1) (9) was custom made and used at a 1:100 dilution with 5% non-fat milk in PBS + 0.1% Tween. Anti-human DPP-IV polyclonal antibody was purchased from EM Biosciences (San Diego, CA) and used at a 1:1000 dilution. As a control for protein loading, actin antibody (C-11) from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA) was used at a 1:1000 dilution. All immunoblots were analyzed by using the ECL detection system (Amersham Biosciences, UK).
Cell proliferation assay
MTS tetrazolium compound from Promega (Madison, WI) was used as a colorimetric indicator for determining the number of viable cells in proliferation or cytotoxicity assays. Assays were performed by adding 20 μL of MTS compound to each well containing the cells in 100 μL of assay medium. Absorbance at 490 nm was recorded after 1-hour incubation at 37 0C in a humidified chamber with 5% CO2.
Results and Discussion
Fluorogenic substrate design
Recent study in substrate development has indicated that glycylprolyl dipeptide is an excellent substrate for DPP-IV; unfortunately, it can also be recognized by FAP (3). Result from a systematic screening of proline containing dipeptide library has suggested a broad substrate exopeptidase selectivity of these two proteases, especially DPP-IV (5). The best, maybe the only, approach to distinguish these two proteases is by measuring the exopeptidase and endopeptidase activity of DPP-IV and FAP, respectively (6).
A water-soluble DPP-IV sensitive probe, GPGP-2SBPO, was previously synthesized to measure dipeptidyl peptidase activity (23). The rationale of having a tetrapeptide to report the dipeptidase activity was the instability of the dipeptide and fluorogenic dye conjugate. GP-2SBPO underwent spontaneous non-enzymatic hydrolysis via intramolecular cyclization to give cyclo[Gly-Pro] and free 2SBPO in physiological conditions. To overcome the problem of this unexpected non-enzymatic hydrolysis, a second glycylprolyl dipeptide was added to the N-terminus of GP-2SBPO to generate the tetrapeptide substrate, GPGP-2SBPO, which was subsequently proven to be useful for DPP-IV sensing. Based on this finding, several tetrapeptide substrates were synthesized to detect exo and endo dipeptidyl protease activity (Fig 1).
1.

Structure and sequences of the synthesized fluorogenic probes.
Substrate selectivity and in vitro enzymatic activity
To evaluate the selectivity of these fluorogenic probes, recombinant murine FAP and DPP-IV enzymes were assayed for proteolytic activity in vitro. GPGP-2SBPO and APAP-2SBPO were used to compare the specific exopeptidase activity of FAP with DPP-IV. Fluorescence signals released from the substrates upon activation by their respective enzyme were monitored simultaneously in a 96-well assay plate. The fluorescence of control referred to the background signal where no enzyme was added. Although both proteases exhibit exopeptidase activity, as shown in Fig 2A, FAP preferred APAP-2SBPO substrate but showed no activity with the GPGP-2SBPO substrate. In contrast, DPP-IV readily cleaved both substrates (Fig 2B) and a greater than 6-fold increase in fluorescence signal was observed with GPGP-2SBPO. These results are consistent with the published data indicating that DPP-IV has a broader substrate specificity for prolyl exopeptidase activity (5).
2.

Substrate selectivity of FAP (A) and DPP-IV (B). The substrates (50 μM) were incubated with either protease (10 nM) in 10 mM PBS (pH 7.4) for 2 hours. The measurement was collected with 630 nm excitation and 670 nm emission. The dotted lines indicate the background signal.
It has been reported that FAP, but not DPP-IV, possesses endopeptidase activity (3, 5, 6). To confirm the endopeptidase specificity of the protease, we assayed FAP and DPP-IV against TSGP-2SPO, Ac-GP-2SBPO, and Ac-GPGP-2SBPO (Fig.2). As expected, DPP-IV showed no activity against any of these substrates (Fig. 2B). However, FAP showed a greater catalytic efficiency for Ac-GPGP-2SBPO than for Ac-GP-2SBPO and TSGP-2SBPO under the same assay reaction conditions.
Ac-GPGP-2SBPO was the most efficient substrate for FAP activation. Its cleavage yielded a 7-fold increase in fluorescence signal (Fig 2). Similar to our previous finding (23), following the specific FAP endopeptidase cleavage between the Pro and Gly residues, a spontaneous non-enzymatic hydrolysis of the resulting GP-2SBPO was happened to free 2SBPO and emit fluorescence. In contrast, the smaller increases in fluorescence signal for Ac-GP-2SBPO and TSGP-2SBPO may be attributable to sequence preference.
To demonstrate the dose response of FAP enzymatic activity, 10 μM Ac-GPGP-2SBPO substrate was mixed with four different concentrations of FAP enzyme and fluorescence signals were measured over a period of 2 hours. The reaction progress curves, shown in Fig. 3, approach a similar maximum plateau fluorescence value. This is an indication that FAP enzyme remains stable under the conditions tested. A near linear dose response (R2 = 0.98) was observed in the first 10 min. These curves were also used to define the amount of FAP enzyme that was needed to maintain initial velocity conditions.
3.
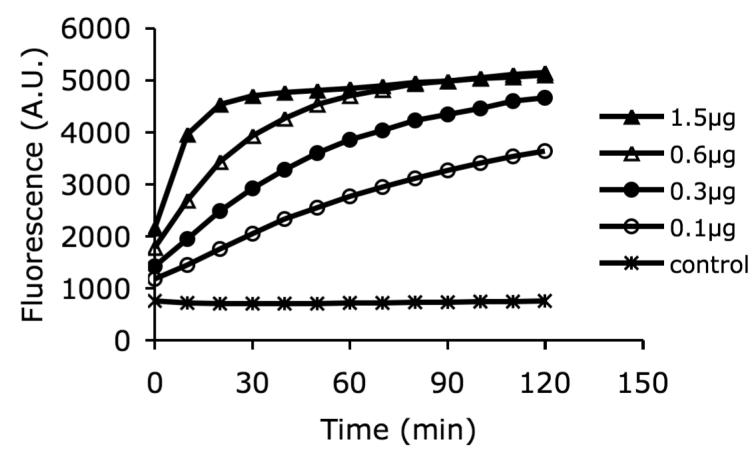
Dose response of fluorescence signal. The fluorescence signal was measured at various times with five different concentrations of FAP and a single Ac-GPGP-2SBPO substrate concentration at 10 uM in PBS. The increase in fluorescence emission was found to be enzyme concentration dependent. The fluorescence of control referred to the background signal where no enzyme was added.
Cell-based enzymatic studies
Having identified Ac-GPGP-2SBPO and GPGP-2SBPO probes as excellent reporter substrates of FAP and DPP-IV, respectively, we were interested to know whether these probes permit a direct and simple readout of the enzyme activity in intact mammalian cells. Three cell lines, i.e., (i) human colon cancer cells HT-29, (ii) human embryonic kidney HEK293 cells stably transfected with full length wild-type murine FAP (HEK-FAP) and (iii) HEK293 cells stably transfected with empty vector (HEK-vector), were used to monitor the selectivity of these two fluorogenic probes.
According to Western blot analyses, the HT-29 cell line expresses DPP-IV but not FAP (Fig 4A). On the other hand, HEK-FAP transfected cells show exogenous expression of FAP. For the HEK-vector control cell line, FAP expression is undetectable. It is also noteworthy that neither of these HEK cell lines express DPP-IV, as seen in the blot. Although FAP is not expressed in the HEK-vector control cell line, interestingly we observed an increase in fluorescence signal when Ac-GPGP-2SBPO substrates were directly incubated with HEK-vector cells in the growth medium which contains a 10% fetal bovine serum (data not shown). Recent studies have reported the identification of soluble forms of FAP from human blood and bovine serum (25, 26). In order to eliminate the interference and be able to monitor the transmembrane proteases in intact cells directly, a phosphate-buffered solution (10 mM) at pH 7.4 was used for subsequent cellular assays.
4.
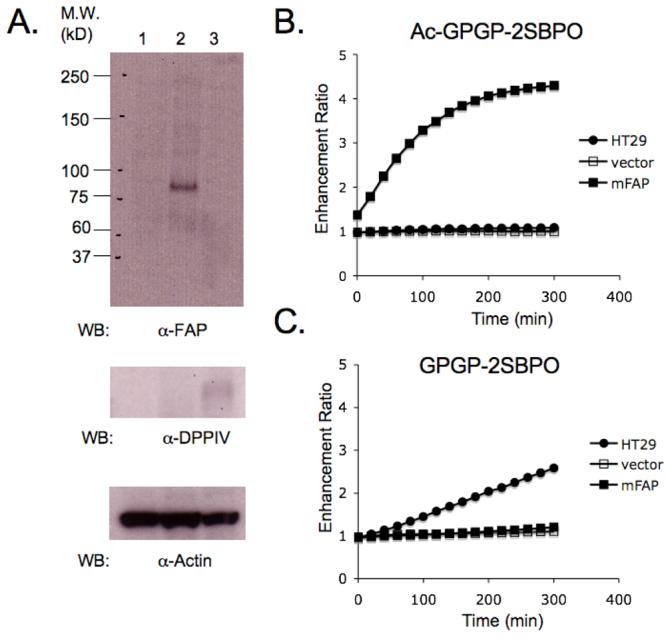
Western blot analysis and detection of dipeptidase activity by cell-based assays. (A) Western blots of cell lysates collected from HEK-vector (column 1), HEK-FAP (column 2) and HT-29 (column 3). (B) Activation of Ac-GPGP-2SBPO in 5 hours. Only HEK-FAP was able to activate the probe. (C) Activation of GPGP-2SBPO in 5 hours. Only HT-29 was able to activate the probe.
As shown in Fig. 4B and 4C, Ac-GPGP-2SBPO probe showed a 4-fold increase in fluorescence after direct incubation with HEK-FAP cells for 5 hours at 37 °C. This result demonstrated that Ac-GPGP-2SBPO possesses a significant selectivity for FAP in HEK-FAP cells as compared with HEK-vector control cells and HT-29 cells. In the case of GPGP-2SBPO, the substrate was selectively cleaved by the endogenous DPP-IV in HT-29 living cells and showed an approximately 3-fold increase in fluorescence in 5 hours. The smaller increase in fluorescence may suggest that the level of activity corresponds to its lower expression levels of DPP-IV within the cells, as supported by Western blot expression data.
Cell proliferation MTS assay was further performed to ensure the probes are non-toxic to cells. HEK-FAP cells were treated with various concentration of Ac-GPGP-2SBPO for 5 hours. Then, MTS assay medium was added into each well and incubated for another 1 hour. The absorbance readings at 490 nm showed no significant difference suggesting that 2SBPO fluorogenic dye is non-toxic under the experimental condition (Fig 5).
5.
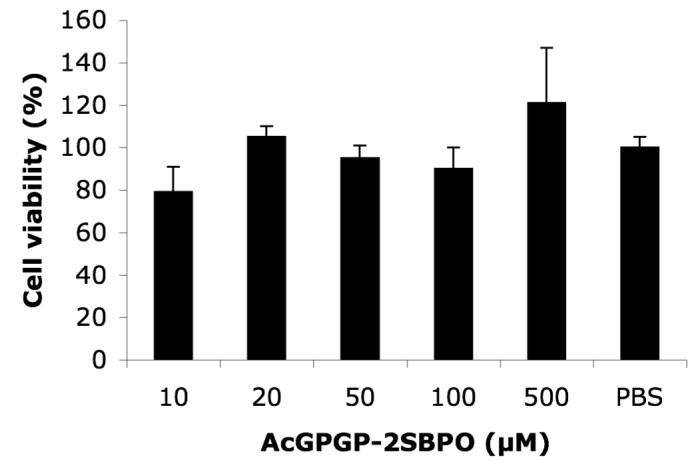
Effects of Ac-GPGP-2SBPO on cell toxicity. HEK-FAP cells were incubated with different concentrations of Ac-GPGP-2SBPO at 37 °C for 1 hr. Experiments were performed in triplicate, mean ± SE.
Conclusions
In summary, a novel near-infrared optical probe has been developed to detect specific FAP endopeptidase activity by either in vitro or in cellular assays. Ac-GPGP-2SBPO is a water-soluble acetylated tetrapeptide fluorescent substrate with the excitation wavelength at 630 nm and emission wavelength at 670 nm. It contains the Gly-Pro dipeptide motif for the FAP-selective endopeptidase cleavage site. We also validated that Ac-GPGP-2SBPO and GPGP-2SBPO probes are excellent reporters for studying FAP and DPP-IV prolyl aminopeptidase activities in cell culture and can be readily adapted for high-throughput drug screening format.
Acknowledgement
This research was supported in part by grants from NIH (CA86355, CA099385 and CA114149 to CT; CA090468, CA103991 and CA006927 to JC), and an appropriation from the Commonwealth of Pennsylvania (JC).
References
- 1.Scanlan MJ, Raj BK, Calvo B, Garin-Chesa P, Sanz-Moncasi MP, Healey JH, Old LJ, Rettig WJ. Molecular cloning of fibroblast activation protein alpha, a member of the serine protease family selectively expressed in stromal fibroblasts of epithelial cancers. Proc Natl Acad Sci U S A. 1994;91:5657–5661. doi: 10.1073/pnas.91.12.5657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cheng JD, Weiner LM. Tumors and their microenvironments: tilling the soil. Clin Cancer Res. 2003;9:1590–1595. [PubMed] [Google Scholar]
- 3.Aertgeerts K, Levin I, Shi L, Snell GP, Jennings A, Prasad GS, Zhang Y, Kraus ML, Salakian S, Sridhar V, Wijnands R, Tennant MG. Structural and kinetic analysis of the substrate specificity of human fibroblast activation protein alpha. J Biol Chem. 2005;280:19441–19444. doi: 10.1074/jbc.C500092200. [DOI] [PubMed] [Google Scholar]
- 4.Park JE, Lenter MC, Zimmermann RN, Garin-Chesa P, Old LJ, Rettig WJ. Fibroblast activation protein, a dual specificity serine protease expressed in reactive human tumor stromal fibroblasts. J Biol Chem. 1999;274:36505–36512. doi: 10.1074/jbc.274.51.36505. [DOI] [PubMed] [Google Scholar]
- 5.Edosada CY, Quan C, Wiesmann C, Tran T, Sutherlin D, Reynolds M, Elliott JM, Raab H, Fairbrother W, Wolf BB. Selective inhibition of fibroblast activation protein protease based on dipeptide substrate specificity. J Biol Chem. 2006;281:7437–7444. doi: 10.1074/jbc.M511112200. [DOI] [PubMed] [Google Scholar]
- 6.Edosada CY, Quan C, Tran T, Pham V, Wiesmann C, Fairbrother W, Wolf BB. Peptide substrate profiling defines fibroblast activation protein as an endopeptidase of strict Gly(2)-Pro(1)-cleaving specificity. FEBS Lett. 2006;580:1581–1586. doi: 10.1016/j.febslet.2006.01.087. [DOI] [PubMed] [Google Scholar]
- 7.Garin-Chesa P, Old LJ, Rettig WJ. Cell surface glycoprotein of reactive stromal fibroblasts as a potential antibody target in human epithelial cancers. Proc Natl Acad Sci U S A. 1990;87:7235–7239. doi: 10.1073/pnas.87.18.7235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dolznig H, Schweifer N, Puri C, Kraut N, Rettig WJ, Kerjaschki D, Garin-Chesa P. Characterization of cancer stroma markers: in silico analysis of an mRNA expression database for fibroblast activation protein and endosialin. Cancer Immun. 2005;5:10. [PubMed] [Google Scholar]
- 9.Cheng JD, Valianou M, Canutescu AA, Jaffe EK, Lee HO, Wang H, Lai JH, Bachovchin WW, Weiner LM. Abrogation of fibroblast activation protein enzymatic activity attenuates tumor growth. Mol Cancer Ther. 2005;4:351–360. doi: 10.1158/1535-7163.MCT-04-0269. [DOI] [PubMed] [Google Scholar]
- 10.Cheng JD, Dunbrack RL, Jr., Valianou M, Rogatko A, Alpaugh RK, Weiner LM. Promotion of tumor growth by murine fibroblast activation protein, a serine protease, in an animal model. Cancer Res. 2002;62:4767–4772. [PubMed] [Google Scholar]
- 11.Coutts SJ, Kelly TA, Snow RJ, Kennedy CA, Barton RW, Adams J, Krolikowski DA, Freeman DM, Campbell SJ, Ksiazek JF, Bachovchin WW. Structure-activity relationships of boronic acid inhibitors of dipeptidyl peptidase IV. 1. Variation of the P2 position of Xaa-boroPro dipeptides. J Med Chem. 1996;39:2087–2094. doi: 10.1021/jm950732f. [DOI] [PubMed] [Google Scholar]
- 12.Nemunaitis J, Vukelja SJ, Richards D, Cunningham C, Senzer N, Nugent J, Duncan H, Jones B, Haltom E, Uprichard MJ. Phase I Trial of PT-100, A Cytokine-Inducing Small Molecule, Following Chemotherapy for Solid Tumor Malignancy. Cancer Invest. 2006;24:553–561. doi: 10.1080/07357900600894732. [DOI] [PubMed] [Google Scholar]
- 13.Wesley UV, Tiwari S, Houghton AN. Role for dipeptidyl peptidase IV in tumor suppression of human non small cell lung carcinoma cells. Int J Cancer. 2004;109:855–866. doi: 10.1002/ijc.20091. [DOI] [PubMed] [Google Scholar]
- 14.Kelly T. Fibroblast activation protein-alpha and dipeptidyl peptidase IV (CD26): cell-surface proteases that activate cell signaling and are potential targets for cancer therapy. Drug Resist Updat. 2005;8:51–58. doi: 10.1016/j.drup.2005.03.002. [DOI] [PubMed] [Google Scholar]
- 15.Gorrell MD. Dipeptidyl peptidase IV and related enzymes in cell biology and liver disorders. Clin Sci (Lond) 2005;108:277–292. doi: 10.1042/CS20040302. [DOI] [PubMed] [Google Scholar]
- 16.Miller S, Onge EL. Sitagliptin: a dipeptidyl peptidase IV inhibitor for the treatment of type 2 diabetes. Ann Pharmacother. 2006;40:1336–1343. doi: 10.1345/aph.1G665. [DOI] [PubMed] [Google Scholar]
- 17.Rosenblum JS, Kozarich JW. Prolyl peptidases: a serine protease subfamily with high potential for drug discovery. Curr Opin Chem Biol. 2003;7:496–504. doi: 10.1016/s1367-5931(03)00084-x. [DOI] [PubMed] [Google Scholar]
- 18.Adams S, Miller GT, Jesson MI, Watanabe T, Jones B, Wallner BP. PT-100, a small molecule dipeptidyl peptidase inhibitor, has potent antitumor effects and augments antibody-mediated cytotoxicity via a novel immune mechanism. Cancer Res. 2004;64:5471–5480. doi: 10.1158/0008-5472.CAN-04-0447. [DOI] [PubMed] [Google Scholar]
- 19.Hu Y, Ma L, Wu M, Wong MS, Li B, Corral S, Yu Z, Nomanbhoy T, Alemayehu S, Fuller SR, Rosenblum JS, Rozenkrants N, Minimo LC, Ripka WC, Szardenings AK, Kozarich JW, Shreder KR. Synthesis and structure-activity relationship of N-alkyl Gly-boro-Pro inhibitors of DPP4, FAP, and DPP7. Bioorg Med Chem Lett. 2005;15:4239–4242. doi: 10.1016/j.bmcl.2005.06.075. [DOI] [PubMed] [Google Scholar]
- 20.Lee J, Fassnacht M, Nair S, Boczkowski D, Gilboa E. Tumor immunotherapy targeting fibroblast activation protein, a product expressed in tumor-associated fibroblasts. Cancer Res. 2005;65:11156–11163. doi: 10.1158/0008-5472.CAN-05-2805. [DOI] [PubMed] [Google Scholar]
- 21.Shreder KR, Wong MS, Corral S, Yu Z, Winn DT, Wu M, Hu Y, Nomanbhoy T, Alemayehu S, Fuller SR, Rosenblum JS, Kozarich JW. Boro-norleucine as a P1 residue for the design of selective and potent DPP7 inhibitors. Bioorg Med Chem Lett. 2005;15:4256–4260. doi: 10.1016/j.bmcl.2005.06.076. [DOI] [PubMed] [Google Scholar]
- 22.Grant SK, Sklar JG, Cummings RT. Development of novel assays for proteolytic enzymes using rhodamine-based fluorogenic substrates. J Biomol Screen. 2002;7:531–540. doi: 10.1177/1087057102238627. [DOI] [PubMed] [Google Scholar]
- 23.Ho NH, Weissleder R, Tung CH. Development of a dual fluorogenic and chromogenic dipeptidyl peptidase IV substrate. Bioorg Med Chem Lett. 2006;16:2599–2602. doi: 10.1016/j.bmcl.2006.02.045. [DOI] [PubMed] [Google Scholar]
- 24.Ho NH, Weissleder R, Tung CH. Development of water-soluble far-red fluorogenic dyes for enzyme sensing. Tetrahedron. 2006;62:578–585. [Google Scholar]
- 25.Collins PJ, McMahon G, O'Brien P, O'Connor B. Purification, identification and characterisation of seprase from bovine serum. Int J Biochem Cell Biol. 2004;36:2320–2333. doi: 10.1016/j.biocel.2004.05.006. [DOI] [PubMed] [Google Scholar]
- 26.Lee KN, Jackson KW, Christiansen VJ, Lee CS, Chun JG, McKee PA. Antiplasmin-cleaving enzyme is a soluble form of fibroblast activation protein. Blood. 2006;107:1397–1404. doi: 10.1182/blood-2005-08-3452. [DOI] [PubMed] [Google Scholar]