

Abstract
To find out whether attentional capture by irrelevant but salient visual objects is an exogenous bottom-up phenomenon, or can be modulated by current task set, two experiments were conducted where the N2pc component was measured as an electrophysiological marker of attentional selection in response to spatially uninformative colour singleton cues that preceded target arrays. When observers had to report the orientation of a uniquely coloured target bar among distractor bars (colour task), behavioural spatial cueing effects were accompanied by an early cue-induced N2pc, indicative of rapid attentional capture by colour singleton cues. In contrast, when they reported the orientation of target bars presented without distractors (onset task), no behavioural cueing effects were found, and no early N2pc was triggered to physically identical cue arrays. Experiment 2 ruled out an alternative interpretation of these N2pc differences in terms of distractor inhibition. These results do not support previous claims that attentional capture is initially unaffected by top-down intention, and demonstrate the central role of task set in involuntary attentional orienting.
Introduction
Selective attention is the mechanism by which organisms bias perceptual and cognitive processing in favour of objects and events that are relevant to their behavioural goals. However, attention can also be attracted by salient perceptual stimuli that are unrelated to current intentions. Ever since James (1890), the dichotomy between an endogenous, voluntary, and controlled mode of allocating attention, and the exogenous, involuntary, and automatic capture of attention has been regarded as a fundamental distinction (e.g., Jonides, 1981; Müller & Rabbitt, 1989).
Evidence for exogenous attentional capture comes from visual search studies where salient non-targets (uniquely coloured “colour singleton” stimuli) were presented together with less salient shape singleton targets. The presence of colour singletons delayed target detection, even though colour was known to be task-irrelevant (e.g., Theeuwes, 1991), suggesting that colour singletons capture attention in a bottom-up fashion, independently of current task sets. However, other studies (Folk, Remington, & Johnston, 1992; Folk, Remington, & Wright, 1994; Folk & Remington, 1998) have challenged this claim. In these experiments, spatially non-predictive peripheral cues were presented 150 ms prior to the onset of a target display. Responses were faster when targets appeared at the cued location, indicative of attentional capture, but, crucially, such spatial cueing effects were only observed when cue properties matched the current task set. When observers had to detect a colour singleton target among non-targets, spatial cueing effects were found when target displays were preceded by colour singleton cues. In contrast, physically identical cue displays failed to produce cueing effects when targets were abrupt onset items. Conversely, with abrupt onset cues, spatial cueing effects were obtained for an onset target task, but not when targets were colour singletons.
The results obtained by Theeuwes (1991) and the findings reported by Folk and colleagues have led to diametrically opposed interpretations with respect to the nature of attentional capture. While the former suggest that capture is a stimulus-driven bottom-up phenomenon that is determined exclusively by the saliency of visual singleton objects, the latter imply that top-down task sets play an important role in the control of exogenous attentional capture. According to the contingent involuntary orienting hypothesis (Folk et al., 1992), attentional capture by salient visual objects is not triggered in a purely automatic bottom-up fashion, but is contingent upon whether or not these objects match task-relevant attributes as defined in the current task set. If this hypothesis is correct, the fact that Theeuwes (1991) found substantially delayed reaction times (RTs) when visual search arrays also contained a task-irrelevant colour singleton needs to be explained without assuming that such colour singletons capture attention in a bottom-up fashion. Folk and Remington (1998) have provided such an explanation by assuming that these RT costs are due to the increased visual complexity of search arrays that contain two singleton stimuli, which extends the duration of pre-attentive processing (filtering costs). In other words, the presence of an additional irrelevant colour singleton delays RTs not because it captures attention in a bottom-up fashion, but because attentional shifts to the target singleton are delayed.
On the other hand, the hypothesis that attention is captured in a completely bottom-up fashion by salient visual singletons has to account for the observation by Folk et al. (1992) that spatial cueing effects are eliminated when singleton cues do not share attributes with targets. Theeuwes, Atchley, and Kramer (2000) have argued that salient singleton stimuli (such as the cues used by Folk and colleagues) always capture attention independently of current task sets. However, attention is then rapidly disengaged from stimuli that have no task-set relevant attributes. In other words, although attention is initially captured in a bottom-up fashion by colour singleton cues, its rapid disengagement from irrelevant cues prior to the presentation of subsequent targets prevents the emergence of behavioural spatial cueing effects.
The question whether involuntary attentional capture is determined by current task set, or is initially triggered in a purely bottom-up fashion remains unresolved. Until now, this debate has been almost exclusively based on behavioural measures. Because overt performance reflects the end result of a long sequence of processing stages, observed behavioural effects are often compatible with competing theoretical claims about underlying cognitive processes. The controversy between the hypothesis that intentional task set can prevent attentional capture (Folk et al., 1992), and the alternative claim that capture occurs regardless of task set, but that attention is rapidly disengaged from task-set irrelevant objects (Theeuwes et al., 2000), is based on specific assumptions about the temporal dynamics of attentional capture that are difficult to dissociate on the basis of behavioural measures alone.
Scalp-recorded event-related brain potentials (ERPs) offer a more precise measure of the time course of perceptual and cognitive processes. In particular, the N2pc component provides a unique temporal marker for the allocation of attention in visual space, and is therefore ideally suited to distinguish between conflicting hypotheses about the nature and time course of attentional capture effects. The N2pc component is characterised by an enhanced negativity over posterior scalp electrodes contralateral to the side of an attended stimulus, is typically elicited between 180 ms and 300 ms after the onset of a visual search array, and is assumed to reflect the attentional selection of candidate target items that are presented among distractors in a visual search task, and/or the attentional inhibition of distractors (Luck & Hillyard, 1994a, 1994b; Eimer, 1996; Girelli & Luck, 1997; Woodman & Luck, 1999). In a recent ERP investigation of attentional capture, Hickey, McDonald, and Theeuwes (2006) measured the N2pc to search arrays that contained a salient non-target colour singleton and a less salient target shape singleton, analogous to the procedures first employed by Theeuwes (1991). When target and non-target singletons were presented on opposite sides, an N2pc elicited by the non-target colour singleton preceded the N2pc elicited by the shape target, suggesting that attentional shifts towards the more salient non-target singleton preceded shifts of attention towards the target. These findings indicate that stimulus-driven bottom-up factors may be more important for attentional capture than top-down tasks sets.
The aim of the present study was to investigate the impact of the manipulation of current task sets on attentional capture, using the N2pc as an indicator of spatially selective attentional processing. In contrast to Hickey et al. (2006), who measured the N2pc in response to visual search displays with target and non-target singletons, we recorded ERPs in a spatial cueing paradigm that was closely modelled on the study by Folk et al. (1992). Each trial started with a 50 ms presentation of a cue array consisting of six sets of small dots arranged in a circular fashion (Figure 1). In Experiment 1, one set of dots was red, all others were grey, and the location of the red colour singleton cues was unrelated to the position of upcoming target stimuli. Target stimulus arrays were presented 200 ms after cue onset. A colour task, where participants reported the orientation of a red target bar presented among grey non-target bars, was compared to an onset task, where the orientation of a single grey target bar had to be reported. Analogous to Folk et al. (1992), behavioural spatial cueing effects indicative of attentional capture were expected for the colour task. In contrast, no such cueing effects should be observed for the onset task, where colour singleton cues were task-set irrelevant.
Figure 1.
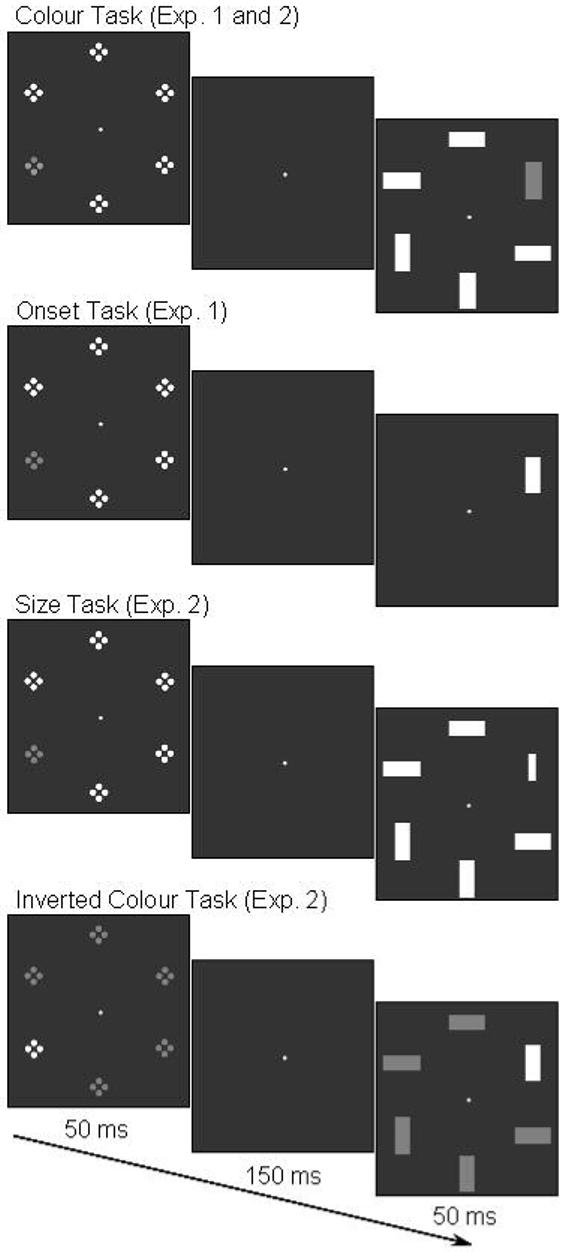
Illustration of the different task conditions used in Experiments 1 and 2. In all tasks, a colour singleton cue array was presented for 50 ms and was followed after a blank interval of 150 ms by a target array (50 ms duration), and participants had to report the orientation of the unique stimulus in the target array. This target stimulus was a red bar among grey bars in the colour task, a single grey bar in the onset task, and a small grey bar among larger grey bars in the size task. In the inverted colour task, the cue singleton was grey among red items, and the unique target stimulus was a grey bar presented among red bars. Red stimuli are shown in dark grey, grey stimuli in white.
To find out whether and how attentional capture by physically identical colour singleton cues was modulated by the current task set, the N2pc component was measured in response to these colour singleton cues, separately for the colour and the onset tasks. The 200 ms interval between cue and target array onset was chosen to ensure that the N2pc triggered by the cue was elicited prior to the emergence of any visual ERP components in response to the subsequent target array. A reliable N2pc component was expected in response to colour singleton cues for the colour task, where these cues shared a task-relevant attribute (colour) with the targets, and thus should capture attention even though they were spatially non-predictive. The critical question was whether and when an N2pc would be triggered by the cues in the onset task. If salient colour singletons capture attention regardless of task set, the N2pc should initially be identical for both tasks. If attention is rapidly disengaged from colour singletons when colour is task-irrelevant, as claimed by Theeuwes et al. (2000), the N2pc should be short-lived in the onset task, but remain present in a more sustained fashion in the colour task. In contrast, if attentional capture is genuinely contingent on task set (Folk et al., 1992), colour singleton cues should not capture attention at all in the onset task, and no N2pc should be observed.
EXPERIMENT 1
Materials and Methods
Participants
Twelve volunteers (mean age 25.3 years, 6 males) were paid to participate in Experiment 1. One of the participants was left-handed, the others were right-handed, and all had normal or corrected-to-normal vision.
Stimuli and Procedure
On each trial, a cue display (50 ms duration) was followed after a 150 ms interval by a target display (50 ms duration). The cue display consisted of a circular array of six sets of four closely aligned dots (each set subtending 0.8° × 0.8° visual angle), presented at a constant distance of 4.4° from central fixation. One set of dots was red (colour singleton cue), while all others were grey (Figure 1). Singleton cues were presented randomly and equiprobably at one of the four left and right locations, but never at the top or bottom. In the colour task (six successive blocks of 64 trials), a red bar was presented at one of the four lateral locations and was accompanied by five grey bars. The size of each bar was 1.3° × 0.5°, and its orientation (horizontal / vertical) varied randomly across trials. Participants had to report the orientation of the red target bar by pressing a left or right response key. In the onset task (six blocks of 64 trials), a single grey target bar was presented randomly at one lateral position and its orientation had to be reported. Colour singleton cues were uninformative with respect to target location (25% validity). All red and grey stimuli were isoluminant (11 cd/m2), and were presented on a 17″ computer monitor against a black background. A small grey fixation dot was continuously present at the screen centre throughout the experimental blocks. Intertrial interval was 1500 ms. Task order and mappings of target orientation to response hand were counterbalanced across participants.
Electrophysiological recording and data analysis
EEG activity was DC-recorded from 23 scalp sites with a sampling rate of 250 Hz (Cz reference, re-referenced offline to common average reference). Impedances were kept below 5 kΩ. EEG was epoched into 700 ms segments from 100 ms prior to cue onset to 600 ms after cue onset. Trials with saccades (voltage exceeding ±30 μV in the HEOG channel), eye blinks (exceeding ±60 μV at Fpz), or muscular artefacts (exceeding ±80 μV at any other electrode) were excluded from analysis. ERPs were averaged relative to a 100 ms pre-cue baseline for each combination of task (colour task versus onset task) and cue position (left versus right hemifield), collapsed across all four possible target locations. The N2pc was quantified by measuring mean ERP amplitudes at lateral posterior electrodes PO7/8 contralateral and ipsilateral to the side of the colour singleton cue within two successive post-cue time windows (early N2pc: 180-235 ms; late N2pc: 235-290 ms). Mean amplitudes were analysed in three-way ANOVAs for the factors task, contralaterality (hemisphere contralateral versus ipsilateral to the colour singleton cue), and cue position. RTs and error rates were analysed in a two-way ANOVA with the factors task and cue-target position (same, different).
Results
Behaviour
Figure 2 shows RTs and error rates in the colour and onset tasks for trials where colour singleton cues and targets were presented at same or at different locations. As expected, spatial cueing effects were present in the colour task, but not in the onset task. A main effect of task (F(1,11)=11.9, p<.01), reflecting faster responses in the onset task, was accompanied by an effect of cue-target position (F(1,11)=27.2, p<.001), and, most importantly, by an interaction of task with cue-target position (F(1,11)=24.5, p<.001). In the colour task, RTs were 37 ms faster when cues and targets were presented at the same location relative to different-location trials (t(11)=5.5, p<.001), while there was no such difference for the onset task (t<1). The same pattern was also present for error rates. A main effect of task (F(1,11)=6.0, p<.032), reflecting superior accuracy in the onset task, was accompanied by an effect of cue-target position (F(1,11)=12.2, p<.01), and, notably, an interaction of task with cue-target position (F(1,11)=14.0, p<.003). In the colour task, accuracy was higher on same-location trials (t(11)=5.0, p<.001), whereas no such difference was present in the onset task (t<1).
Figure 2.

Response times (line graphs) and error rates (bar graphs) in the colour and onset tasks of Experiment 1 for trials where colour singleton cues and subsequent targets were presented at the same location or at different locations.
ERPs
To demonstrate that cue-induced and target-induced ERP activity can be reliably dissociated in time, Figure 3 shows ERPs obtained in the 600 ms interval after cue onset in the colour task at lateral posterior electrodes PO7/PO8, collapsed across all cue and target locations. Cue-induced P1, N1, and N2 components were followed by P1 and N1 components triggered by the subsequent target array, and the onset of the target P1 occurred almost exactly 300 ms after cue onset. Figure 4 shows ERPs elicited in the colour and onset tasks by physically identical cue arrays at PO7/8 contralateral and ipsilateral to the side of a colour singleton cue, together with difference waveforms obtained by subtracting ipsilateral from contralateral activity (bottom panel). A solid N2pc with an onset latency of about 180 ms was triggered in the colour task, while no N2pc was present within this time range in the onset task.
Figure 3.

ERPs elicited in the 600 ms interval after cue onset in the colour task of Experiment 1, collapsed across lateral posterior electrodes PO7/PO8, and across all cue and target locations. P1, N1, and N2 components triggered by the cue array are followed by P1 and N1 components elicited in response to the target array - P1(t) and N1(t). ‘T’ marks the onset of the target array.
Figure 4.

ERPs elicited in the 300 ms interval after cue onset in the colour task (top left) and in the onset task (top right) of Experiment 1 at posterior electrode sites PO7/8 contralateral (solid lines) and ipsilateral (dashed lines) to the visual hemifield where the colour singleton cue was presented. The bottom panel shows difference waves obtained by subtracting ipsilateral from contralateral ERPs in the colour task (solid line) and onset task (dashed line).
The analysis of ERP mean amplitudes obtained during the early N2pc time window (180-235 ms post-cue) revealed main effects of task and contralaterality (F(1,11)=12.1 and 8.5, both p<.015). The interaction of task with contralaterality was also significant (F(1,11)=14.4, p<.003). Separate analyses for both tasks found a main effect of contralaterality in the colour task (F(1,11)=13.3, p<.004), reflecting the presence of the N2pc, whereas no such effect was present in the onset task (F(1,11)=1.6, p=.239). Figure 7 (top left panel) shows the scalp topography of the N2pc elicited between 180 and 235 ms after cue onset in the colour task. In the subsequent late N2pc time window (235-290 ms after cue onset), a main effect of contralaterality was obtained (F(1,11)=6.0, p<.033). The interaction of task and contralaterality was only marginally significant, F(1,11)=3.4, p<.09. Separate analyses showed again a main effect of contralaterality for the colour task (F(1,11)=5.2, p<.043), but not for the onset task (F(1,11)=2.8, p=.124).
Figure 7.
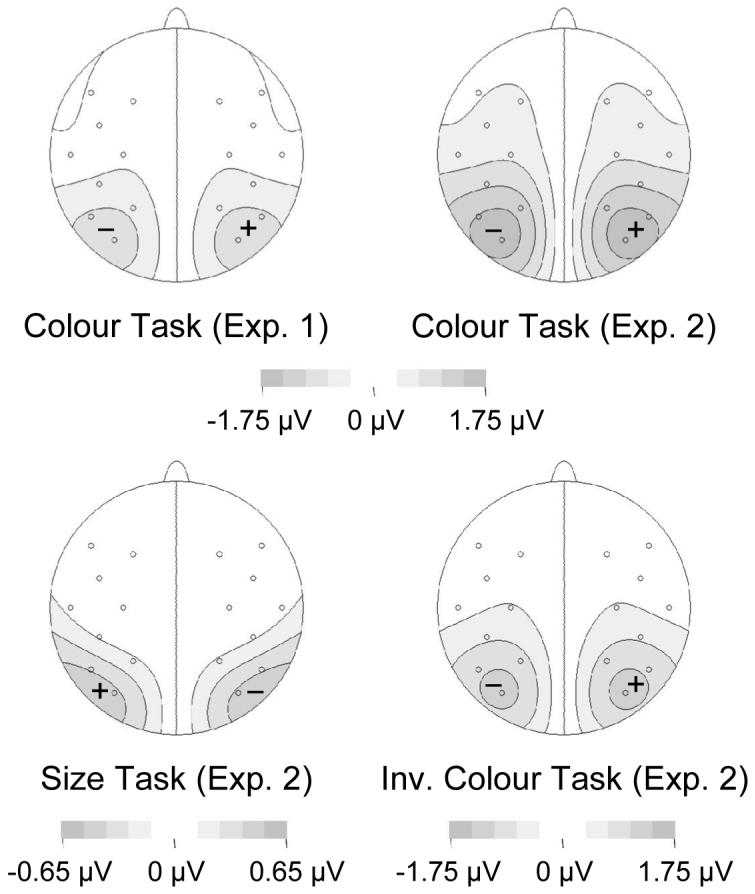
Topographical maps of cue-induced N2pc scalp distributions obtained during the 180-235 ms time interval after cue onset. These maps were constructed by spherical spline interpolation (see Perrin, Pernier, Bertrand, & Echallier, 1989) after mirroring the ipsilateral-contralateral difference waveforms to obtain symmetrical voltage values for both hemispheres. As a result of the mirroring procedure, the N2pc appears as negative voltage (-) over the left hemisphere and as positive voltage (+) over the right hemisphere. Note the different scale used for the size task of Experiment 2 (bottom left panel), and the inverted polarity of the N2pc effect in this task, representing an enhanced negativity ipsilateral to the side of the colour singleton cue.
Discussion
In Experiment 1, the N2pc component was measured as an electrophysiological marker of the spatial allocation of attention to address the current controversy about the role of current task-sets in attentional capture. Does the absence of spatial cueing effects for singleton cues that lack task-relevant features demonstrate that such cues fail to attract attention automatically (Folk et al., 1992), or is attention initially captured, but then rapidly disengaged (Theeuwes et al., 2000)? The results obtained in Experiment 1 appear to comprehensively rule out the latter claim. In the colour task, where singleton cues shared task-relevant features with the targets, behavioural spatial cueing effects were accompanied by a solid N2pc component with an onset latency of about 180 ms after cue presentation. This demonstrates that although spatially non-informative, colour singleton cues did capture attention under conditions where targets were also colour singletons. In marked contrast, no behavioural cueing effects were found in the onset task, thus replicating the results reported by Folk et al. (1992). Crucially, no N2pc was triggered in response to physically identical cue arrays during the critical early N2pc time window (180-235 ms post-cue). If colour singleton cues had initially captured attention in a bottom-up fashion and irrespective of current task set, as argued by Theeuwes et al. (2000), the early phase of the N2pc should have been very similar for both tasks. This was clearly not the case; in fact no reliable N2pc was observed in the onset task in either the early or the late N2pc time window. However, Figure 4 suggests that a small and delayed N2pc may have been present beyond 260 ms post-stimulus in the onset task. This observation was statistically confirmed (main effect of contralaterality for the 265-300 ms interval in the onset task: F(1,11)=15.4, p<.002), and suggests that colour singleton cues did attract attention on some trials during the onset task, although substantially later than in the colour task. This N2pc delay is not in line with the claims of Theeuwes et al. (2000), who would have predicted identical early N2pc onsets for both tasks. Instead, the presence of an attenuated and late N2pc in the onset task suggests that although intentional task sets may not always completely prevent attentional capture by task-irrelevant singleton objects, they substantially delay the onset of any involuntary attention shifts.
The results obtained in Experiment 1 strongly suggest that attentional capture is dependent on current task set, and is mediated by whether or not salient visual singletons possess task-relevant attributes. One could argue that even though all stimuli were isoluminant, red items may still have been more salient than grey items, and that the N2pc observed in response to colour singleton cues was at least in part due to the bottom-up saliency of red singletons. Since cue arrays were physically identical in the colour and onset tasks, this argument cannot account for the marked N2pc differences between the two tasks. Nevertheless, the possibility that the bottom-up saliency of red colour singleton cues may have contributed to the cue-elicited N2pc in the colour task will be further investigated in Experiment 2.
There is however an alternative explanation of the N2pc differences between the colour and onset tasks observed in Experiment 1 that does not imply task-set contingent attentional capture. Although the cue arrays were identical in both tasks, target arrays were very different, and this difference may have had a systematic effect on the cue-elicited ERPs. In the colour task, a red target was presented among grey distractors, and target selection may thus have involved the suppression of distractor items. In contrast, no suppression was required in the onset task, where targets were not accompanied by distractors. If participants had adopted a generalized search strategy that involved distractor suppression in the colour task, suppression may also have been applied to grey items in the cue array. The N2pc has in fact previously been linked to the attentional suppression of distractors that are presented close to the target (Luck & Hillyard, 1994b; but see Eimer, 1996, for N2pc results that appear inconsistent with this distractor suppression account). Its presence in the colour task could thus be linked to the suppression of task-set irrelevant cue elements, while its absence in the onset task might indicate the lack of suppression. In other words, the N2pc differences observed between these two tasks may not be related to the presence versus absence of rapid attentional capture, but instead to the presence versus absence of to-be-suppressed distractor items in the target array. Experiment 2 was conducted to test this alternative explanation.
EXPERIMENT 2
In Experiment 1, the N2pc observed for colour singleton cues in the colour task was interpreted as evidence for attentional capture, and its absence in the onset task as a demonstration that attentional capture is task-set contingent. To investigate the alternative hypothesis that this N2pc difference between tasks reflects the need to suppress distractor items in the colour task, and the absence of distractor suppression in the onset task, Experiment 2 included a new task where target items were presented among non-target distractors (analogous to the colour task of Experiment 1), but colour singleton cues were task-set irrelevant (analogous to the onset task of Experiment 1). In this task, red colour singleton cues were followed by target arrays that contained a single smaller grey bar among larger grey distractor bars, and participants had to discriminate the orientation of this size singleton target (size task; see Figure 1). Because colour singleton cues were task-set irrelevant in this task, no behavioural spatial cueing effects were expected, analogous to the onset task in Experiment 1. The critical question was whether an N2pc would still be elicited in response to colour singleton cues in this size task, where target items were presented among non-target distractors. To investigate this, the cue-elicited N2pc was measured for this task and then compared to the N2pc observed for a colour task that was identical to Experiment 1. If the N2pc observed in Experiment 1 for the colour task was primarily due to a generalized distractor suppression mode, it should be present for both colour and size tasks in Experiment 2, as targets were always presented among distractors in these two tasks. In contrast, if the cue-elicited N2pc reflected task-set contingent attentional capture, it should be present for the colour task, but not for the size task where colour singletons cues did not have target-defining features.
In addition, Experiment 2 investigated the possibility that the cue-elicited N2pc observed in Experiment 1 was at least in part due to the bottom-up saliency of red colour singleton cues by directly comparing behavioural cueing effects and cue-induced N2pc components in the colour task and in an ‘inverted colour task’ where cue and target arrays contained a grey singleton item that was presented among red distractors (see Figure 1). Any difference in the bottom-up saliency between red and grey colour singletons should be reflected by systematic differences in behavioural cueing effects and cue-induced N2pc components between the colour and inverted colour tasks.
Materials and Methods
Participants
Twelve volunteers (mean age 27.8 years, 3 males) were paid to participate in Experiment 2. Again, one of the participants was left-handed, the other participants were right-handed, and all had normal or corrected-to-normal vision.
Stimuli, Procedure, EEG recording, and analyses
The experiment consisted of three tasks (colour task, size task, and inverted colour task), with six successively presented blocks of 64 trials for each task. Task order was counterbalanced across participants. The colour task was identical to Experiment 1. The size task was identical to the colour task, except that colour singleton targets were replaced by size singleton targets (see Figure 1). These were horizontally or vertically oriented grey bars that were smaller (0.9° × 0.2°) than the grey distractor bars (1.3° × 0.5°). Participants had to report the orientation of these size singleton targets by pressing the left or right response key. The inverted colour task was identical to the colour task, except that the cue array now contained one grey set of dots among red sets of dots, and the target array contained a grey singleton target among red distractors (see Figure 1). Participants had to respond to the orientation of the grey singleton target bar. In all other respects, stimuli and procedures were identical to Experiment 1.
EEG recording and analysis procedures were identical to Experiment 1, except that the N2pc was now quantified separately for three tasks. In the overall ANOVAs, the factor task now had three levels (colour task versus size task versus inverted colour task). Greenhouse-Geisser corrections for nonsphericity were applied where appropriate.
Results
Behaviour
Figure 5 shows RTs and error rates in the colour, size, and inverted colour tasks, for trials where colour singleton cues and subsequent targets were presented at same or at different locations. Spatial cueing effects were present in the colour and inverted colour tasks, but not in the size task. A main effect of task (F(2,22)=11.2, p<.001) was obtained, and follow-up analyses using two-tailed t-tests revealed that responses tended to be generally slower in the size task relative to the colour task (t(11)=2.3, p<.05) and inverted colour task (t(11)=1.8, p=.095), whereas RTs did not differ between colour and inverted colour tasks (t(11)<1.0). More importantly, a main effect of cue-target position (F(1,11)=94.1, p<.001) was accompanied by an interaction of task with cue-target position (F(2,22)=39.6, p<.001). In both colour and inverted colour tasks, mean RTs were on average 48 ms faster when cues and targets were presented at the same location relative to different-location trials (t(11)=8.3 and 8.7, respectively, both p<.001). In contrast, mean RTs were on average 5 ms faster for different-location trials in the size task, although this tendency was not statistically significant (t(11)=1.4, p=.18). An analogous pattern was observed for error rates (see Figure 5). A main effect of task (F(2,22)=8.1, p<.003), reflecting slightly lower accuracy in the size task, was accompanied by an effect of cue-target position (F(1,11)=22.1, p<.001), and, notably, an interaction of task with cue-target position, F(2,22)=5.0, p<.029. In the colour and inverted colour tasks, accuracy was higher on same-location relative to different-location trials (t(11)=4.9 and 3.9, both p<.003), whereas no such difference was present in the size task (t<1).
Figure 5.

Response times (line graphs) and error rates (bar graphs) in the colour, size, and inverted colour tasks of Experiment 2 for trials where colour singleton cues and subsequent targets were presented at the same location or at different locations.
ERPs
Figure 6 shows ERPs elicited in the colour, size, and inverted colour tasks in response to colour singleton cues at PO7/8 contralateral and ipsilateral to the side of a singleton cue, together with difference waveforms obtained by subtracting ipsilateral from contralateral activity (bottom panel). As in Experiment 1, a solid N2pc with an onset latency of about 180 ms was triggered in the colour task. An N2pc was also clearly elicited in the inverted colour task, although it was slightly delayed relative to the N2pc observed in the colour task. In contrast, there was no evidence for any contralaterally enhanced negativity in the size task. If anything, ERPs tended to be slightly more negative ipsilateral to the side of the colour singleton cue in this task (see Figure 6).
Figure 6.

ERPs elicited in the 300 ms interval after cue onset in the colour task, size task, and inverted colour tasks of Experiment 2 at posterior electrode sites PO7/8 contralateral (solid lines) and ipsilateral (dashed lines) to the visual hemifield where the colour singleton cue was presented. The bottom panel shows difference waves obtained by subtracting ipsilateral from contralateral ERPs in the colour task (solid black line), size task (dashed line), and inverted colour task (solid grey line).
In the early N2pc time window (180-235 ms after cue onset), a main effect of contralaterality (F(1,11)=6.6, p<.026) was accompanied by an interaction of task and contralaterality (F(2,22)=8.0, p<.016). Separate analyses for each task revealed a significant main effect of contralaterality for the colour task (F(1,11)=9.8, p<.01) as well as for the inverted colour task (F(1,11)=5.0, p<.05), reflecting the presence of an N2pc in response to colour singleton cues in both tasks. The scalp distribution of this early N2pc is shown in Figure 7 for the colour task (top right panel) and the inverted colour task (bottom right panel). Interestingly, there was also a significant effect of contralaterality in the size task (F(1,11)=9.8, p<.01), which was due to the small but reliable inverted N2pc elicited in this task. As can be seen in Figure 7 (bottom left panel), the scalp distribution of this lateralised effect was similar to the cue-induced N2pc observed for the other tasks, although its polarity was reversed. In the late N2pc time window (235-290 ms post-cue), main effects of task (F(2,22)=6.0, p<.016) and contralaterality (F(1,11)=19.0, p<.002) were accompanied by a task by contralaterality interaction (F(2,22)=15.7, p<.001). Separate analyses confirmed main effects of contralaterality for the colour and inverted colour tasks (F(1,11)=15.1 and 19.4, both p<.003), whereas the inverted N2pc effect in the size task did not reach significance (F(1,11)=3.5, p=.089).
Figure 6 suggests that while reliable N2pc components were triggered contralateral to the colour singleton cue in both the colour and inverted colour tasks, the N2pc peak was slightly delayed in the inverted colour task. To investigate this latency difference, N2pc peak latencies (i.e., the latency of the maximal negative value obtained in contralateral-ipsilateral difference waveforms at PO7/PO8 in the 180-290 ms post-cue time window) were computed for each participant in both tasks. In the inverted colour task, the N2pc peak was indeed significantly delayed relative to the colour task (257 vs. 242 ms, t(11)=7.2, p<.001).
Discussion
A cue-elicited N2pc component was triggered in the colour task of Experiment 2, replicating the findings from Experiment 1. An N2pc was also elicited in response to grey singleton cues presented among red items in the inverted colour task where target singletons were grey among red bars. This demonstrates that the N2pc to the cues does not depend on the presence of salient red singletons among grey distractors, but can also be observed when this colour assignment is reversed. This conclusion was further underlined by the fact that identical behavioural spatial cueing effects were obtained in the colour and inverted colour tasks. However, the N2pc peaked slightly earlier in the colour task than in the inverted colour task (see Figure 6). This small but reliable latency effect could be due to the continuous presence of the fixation dot at the screen centre. The fact that this fixation dot was grey in all task conditions (as in Experiment 1) may have somewhat reduced the singleton status of grey items in the inverted colour task.
The principal purpose of Experiment 2 was to test the claim that the cue-triggered N2pc observed in the colour task of Experiment 1 is not related to attentional capture, but instead to the necessity to suppress distractor items in the target array. According to this hypothesis, cue-elicited N2pc components should have been similar in the colour and size tasks of Experiment 2, as targets were presented among non-target distractors in both tasks. However, this prediction was not confirmed. In the size task, where targets were small singleton bars presented among uniform larger bars, no N2pc was elicited contralateral to the side of colour singleton cues. As colour was task-irrelevant in the size task, this is exactly what is predicted by the hypothesis that attentional capture is contingent upon top-down task sets. In contrast, this observation rules out the alternative distractor suppression account, which had predicted similar N2pc effects for the colour and size tasks. Additional support for contingent capture is provided by the absence of behavioural spatial cueing effects by colour singleton cues in the size task. This finding extends the behavioural results reported by Folk et al. (1992), and suggests that when stimulus size is task-relevant, colour singletons do not capture attention in a bottom-up fashion.
An unexpected finding was the presence of a small but reliable inverted cue-elicited effect in the size task, with enhanced negativities in the N2pc time range ipsilateral to the colour singleton cue (see Figure 6). As can be seen in Figure 7 (bottom left panel), this lateralised effect showed the typical posterior scalp distribution of an N2pc, except for its reversed polarity with respect to the colour singleton location. The presence of an inverted N2pc in the size task appears to suggest a tendency for participants to direct attention away from the side of the colour singleton cue, and towards the opposite side of the cue array. Although this effect was not predicted, the contingent attentional capture hypothesis may offer a tentative explanation. The grey items in the cue array may have been sufficiently similar in terms of their colour and size to the target items in the size task (small grey singleton bars) to elicit some task-set contingent attentional capture. This may have resulted in a small net attentional bias away from the side of the cue array that contained the red colour singleton item and one grey item, and towards the side with two grey items. According to this explanation, which will be tested in future studies, the N2pc is sensitive to subtle (and in this case purely inadvertent) variations in the capacity of irrelevant visual stimuli to attract attention in a task-set contingent fashion, even when these are too small to produce reliable behavioural effects. An alternative possibility is that participants may have actively directed attention away from colour singleton cues in the size task, perhaps to reduce interference between these cues and subsequent size target stimuli at cued locations. The presence of a non-significant trend for RTs to be faster on different-location trials in the size task is consistent with both of these accounts.
While the presence of a small inverted N2pc in the size task was unexpected, it only serves to further emphasize the systematic effects of task set on N2pc components triggered in response to physically identical cues in the colour and size tasks. These differences cannot be accounted for by the presence versus absence of to-be-suppressed distractors, and thus provide further compelling evidence for task-set contingent attentional capture.
General Discussion
In the present study, the N2pc was used as an electrophysiological marker of selective attentional processing to investigate whether rapid attentional capture by spatially uninformative singleton cues is contingent upon top-down task sets. In Experiment 1, colour singleton cues triggered an N2pc when targets were also colour singletons, but not when targets were onset items, in line with the contingent attentional capture hypothesis. Experiment 2 ruled out an alternative interpretation of this difference in terms of distractor inhibition by demonstrating that under conditions where targets were always presented among non-target distractors, an N2pc was elicited contralateral to colour singleton cues only when colour was the relevant target-defining feature, but not when targets were defined by their size. In both experiments, the pattern of behavioural spatial cueing effects mirrored these electrophysiological results, which provide new and compelling evidence in support of the hypothesis that attentional capture is mediated by top-down task sets.
According to Theeuwes et al. (2000), the absence of behavioural spatial cueing effects for task-irrelevant singleton cues are due to the fact that although attention is initially captured in a bottom-up fashion by these cues, it is rapidly disengaged from cues that lack target-defining features. If this assumption was correct, an N2pc should have initially been triggered by all singleton cues irrespective of task instructions, whereas the later phase of the N2pc may have been differentially affected by top-down task set. The current results do not support this prediction. Cue-elicited N2pc components emerged early in the colour task (Experiment 1 and 2) and inverted colour task (Experiment 2), as would be expected under conditions where salient singleton cues share target-defining features. In marked contrast, no N2pc was triggered at all during the critical early N2pc time window (180-235 ms post-cue) in the onset task (Experiment 1), and a small but reliable inverted N2pc was observed in the size task (Experiment 2) where physically identical colour singleton cues were task-irrelevant. These strong effects of task set on the early phase of the N2pc to colour singleton cues does not support the assumptions of Theeuwes et al. (2000; see also Folk & Remington, 2006, for additional behavioural evidence against the rapid disengagement account). They are however fully consistent with the hypothesis that attentional capture is mediated by top-down task-set (Folk et al., 1992).
The N2pc to colour singleton cues emerged relatively early (at about 180 ms post-stimulus) in the present study, and overlapped with the peak of the N1 component (see Figures 4 and 6). In contrast, an N2pc to colour singletons started about 40 ms later in a previous study (Hickey et al., 2006) when they were accompanied by salient shape singleton targets in the opposite hemifield. This latency difference is likely due to the fact that the cue arrays used in the current experiments contained a colour singleton on the left or right side without another salient singleton item on the other side. As identical cue arrays were used in all task conditions investigated here, this factor cannot account for any of the marked N2pc differences that were observed between these conditions.
One could argue that while the present results have demonstrated that the N2pc is eliminated when colour singleton cues do not share target features, the absence of an N2pc does not necessarily imply the absence of attentional capture. In other words, a transient attention shift might have been triggered to the location of colour singleton cues in the onset task of Experiment 1, and in the size task of Experiment 2, followed by rapid attentional disengagement (as postulated by Theeuwes et al., 2000), without this being reflected by an N2pc. Such a scenario might be plausible if the N2pc was not linked to the initial allocation of spatial attention, but primarily to the subsequent attentional analysis of candidate target events at specific locations. However, recent studies have found reliable N2pc components to task-irrelevant colour singletons that were presented together with shape singleton targets (Hickey et al., 2006), as well as under conditions where non-targets were presented at locations that were previously cued as potentially task-relevant (Kiss, Van Velzen, & Eimer, in press), suggesting that the N2pc is elicited even when attentional allocation is not followed by further in-depth processing (see also Mazza, Turatto, Umiltà, & Eimer, 2007, for further evidence). Given these observations, it appears reasonable to assume that in the present study, rapid attentional capture should have been reflected by the N2pc.
This assumption is further supported by the fact that numerous previous studies have demonstrated firm links between the N2pc and the attentional selection of candidate target events (e.g., Luck & Hillyard, 1994a, 1994b; Eimer, 1996; Girelli & Luck, 1997; Woodman & Luck, 1999; Brisson & Jolicoeur, 2006; Jolicoeur, Sessa, Dell’Acqua, & Robitaille, 2006; Kiss et al., 2007; Mazza et al., 2007). In fact, results from several earlier N2pc studies are consistent with contingent attentional capture. For example, Luck and Hillyard (1994b, Experiment 1) studied pop-out visual search where targets or non-target singleton stimuli were presented together with homogeneous distractors. Targets were defined by a specific combination of colour, size, and orientation, and non-target singleton items either shared two of these features with the targets (‘difficult’ non-targets) or differed from the targets on all three dimensions (‘easy’ non-targets). While an N2pc was elicited in response to targets as well as difficult non-targets, no N2pc was found for easy non-targets. Luck and Hillyard (1994b) interpreted these results as reflecting the attentional processing of non-targets that require careful analysis to be distinguished from targets, without referring explicitly to task-set contingent attentional capture. However, the contingent capture hypothesis is perfectly consistent with their findings: An N2pc is triggered in response to difficult non-targets because these share target-defining features, but not to easy non-targets that lack any task-relevant features (see also Luck & Hillyard, 1994a, and Girelli & Luck, 1997, for further evidence that the N2pc to singleton items in pop-out visual search is determined by top-down task set).
While the present findings strongly support the view that attentional capture is mediated by top-down task sets, results from a recent ERP study that also employed the N2pc as an indicator of selective attentional processing (Hickey et al., 2006) have led to the conclusion that attentional capture is at least partially determined by bottom-up saliency. When a target shape singleton and a more salient non-target colour singleton were presented in opposite hemifields together with uniform distractor items, a small but reliable N2pc was triggered contralateral to the colour singleton between 220 and 265 ms post-stimulus, prior to the emergence of an N2pc to the shape target. This suggests that attention was initially captured in a bottom-up fashion by the irrelevant colour singleton before it was directed to the shape target. It is possible that procedural differences between the spatial cueing paradigm used in the present study and the two-singleton visual search task employed by Hickey et al. (2006) may account for these discrepant results. When attentional capture is investigated with cueing procedures (see also Folk et al., 1992), irrelevant singletons (cues) and relevant singletons (targets) are separated in time, whereas they are presented simultaneously in visual search arrays as used by Hickey et al. (2006) and Theeuwes (1991). Thus, these two paradigms differ substantially with respect to the impact of temporal attention, as well as with regard to the possibility of direct perceptual competition between simultaneously presented singleton items. It is worth noting that behavioural evidence for task-set contingent attentional capture has come primarily from studies that used spatial cueing procedures, whereas evidence for bottom-up driven capture effects has been found predominantly with two-singleton visual search displays (see Ruz & Lupiáñez, 2002, for more detailed discussion), suggesting that these two procedures may not be equivalent with respect to their susceptibility to top-down versus bottom-up attentional capture. Future studies need to employ the N2pc as a measure of attentional capture in order to directly compare and contrast the role of top-down and bottom-up factors for attentional capture in spatial cueing and visual search experiments. In addition, further spatial cueing experiments should also investigate contingent attentional capture with a fully crossed design (as in Folk et al., 1992), by orthogonally manipulating cue type (e.g., colour cue versus size cue) and top-down task set (e.g., colour task versus size task).
In summary, the present study has used the N2pc as electrophysiological marker of selective attentional processing to demonstrate that attentional capture by salient visual events is strongly dependent on top-down control, and is not triggered in a bottom-up fashion, irrespective of current task sets. These results provide new evidence in favour of the contingent involuntary orienting hypothesis.
Acknowledgements
This research was supported by a grant from the Biotechnology and Biological Sciences Research Council (BBSRC), UK. M.E. holds a Royal Society-Wolfson Research Merit Award. The authors thank Silvia Dalvit and Heijo Van de Werf for technical assistance.
REFERENCES
- Brisson B, Jolicoeur P. Electrophysiological evidence of central interference on the control of visual-spatial attention. Psychonomic Bulletin & Review. 2007;14:126–132. doi: 10.3758/bf03194039. [DOI] [PubMed] [Google Scholar]
- Eimer M. The N2pc component as an indicator of attentional selectivity. Electroencephalography and Clinical Neurophysiology. 1996;99:225–234. doi: 10.1016/0013-4694(96)95711-9. [DOI] [PubMed] [Google Scholar]
- Folk CL, Remington RW. Selectivity in distraction by irrelevant featural singletons: Evidence for two forms of attentional capture. Journal of Experimental Psychology: Human Perception and Performance. 1998;24:847–858. doi: 10.1037//0096-1523.24.3.847. [DOI] [PubMed] [Google Scholar]
- Folk CL, Remington RW. Top-down modulation of preattentive processing: Testing the recovery account of contingent capture. Visual Cognition. 2006;14:445–465. [Google Scholar]
- Folk CL, Remington RW, Johnston JC. Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance. 1992;18:1030–1044. [PubMed] [Google Scholar]
- Folk CL, Remington RW, Wright JH. The structure of attentional control: Contingent attentional capture by apparent motion, abrupt onset, and colour. Journal of Experimental Psychology: Human Perception and Performance. 1994;20:317–329. doi: 10.1037//0096-1523.20.2.317. [DOI] [PubMed] [Google Scholar]
- Girelli M, Luck SJ. Are the same attentional mechanisms used to detect visual search targets defined by color, orientation, and motion? Journal of Cognitive Neuroscience. 1997;9:238–253. doi: 10.1162/jocn.1997.9.2.238. [DOI] [PubMed] [Google Scholar]
- Hickey C, McDonald JJ, Theeuwes J. Electrophysiological evidence of the capture of visual attention. Journal of Cognitive Neuroscience. 2006;18:604–613. doi: 10.1162/jocn.2006.18.4.604. [DOI] [PubMed] [Google Scholar]
- James W. The Principles of Psychology. Vol. 1. New York: Dover; 1890. [Google Scholar]
- Jolicoeur P, Sessa P, Dell’Acqua R, Robitaille N. Attentional control and capture in the attentional blink paradigm: Evidence from human electrophysiology. European Journal of Cognitive Psychology. 2006;18:560–578. [Google Scholar]
- Jonides J. Voluntary vs. automatic control over the mind’s eye’s movement. In: Long JB, Baddeley AD, editors. Attention and Performance IX. Hillsdale, NJ: Erlbaum; 1981. pp. 187–203. [Google Scholar]
- Kiss M, Goolsby BA, Raymond JE, Shapiro KL, Silvert L, Nobre AC, Fragopanagos N, Taylor JG, Eimer M. Efficient attentional selection predicts distractor devaluation: ERP evidence for a direct link between attention and emotion. Journal of Cognitive Neuroscience. 2007;19:1316–1322. doi: 10.1162/jocn.2007.19.8.1316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiss M, Van Velzen J, Eimer M. The N2pc component and its links to attention shifts and spatially selective visual processing. Psychophysiology. doi: 10.1111/j.1469-8986.2007.00611.x. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luck SJ, Hillyard SA. Electrophysiological correlates of feature analysis during visual search. Psychophysiology. 1994a;31:291–308. doi: 10.1111/j.1469-8986.1994.tb02218.x. [DOI] [PubMed] [Google Scholar]
- Luck SJ, Hillyard SA. Spatial filtering during visual search: Evidence from human electrophysiology. Journal of Experimental Psychology: Human Perception and Performance. 1994b;20:1000–1014. doi: 10.1037//0096-1523.20.5.1000. [DOI] [PubMed] [Google Scholar]
- Mazza V, Turatto M, Umiltà C, Eimer M. Attentional selection and identification of visual objects are reflected by distinct electrophysiological responses. Experimental Brain Research. 2007;181:531–536. doi: 10.1007/s00221-007-1002-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller HJ, Rabbitt PM. Reflexive and voluntary orienting of attention: Time course of activation and resistance to interruption. Journal of Experimental Psychology: Human Perception and Performance. 1989;15:315–330. doi: 10.1037//0096-1523.15.2.315. [DOI] [PubMed] [Google Scholar]
- Perrin F, Pernier J, Bertrand O, Echallier J. Spherical splines for scalp potential and current density mapping. Electroencephalography & Clinical Neurophysiology. 1989;72:184–187. doi: 10.1016/0013-4694(89)90180-6. [DOI] [PubMed] [Google Scholar]
- Ruz M, Lupiáñez J. A review of attentional capture: On its automaticity and sensitivity to endogenous control. Psychológica. 2002;23:283–309. [Google Scholar]
- Theeuwes J. Cross-dimensional perceptual selectivity. Perception & Psychophysics. 1991;50:184–193. doi: 10.3758/bf03212219. [DOI] [PubMed] [Google Scholar]
- Theeuwes J, Atchley P, Kramer AF. On the time course of top-down and bottom-up control of visual attention. In: Monsell S, Driver J, editors. Attention and Performance XVIII. Cambridge, MA: MIT Press; 2000. pp. 105–124. [Google Scholar]
- Woodman GF, Luck SJ. Electrophysiological measurement of rapid shifts of attention during visual search. Nature. 1999;400:867–869. doi: 10.1038/23698. [DOI] [PubMed] [Google Scholar]