

Abstract
To examine the possibility of active recycling of Emp24p between the endoplasmic reticulum (ER) and the Golgi, we sought to identify transport signal(s) in the carboxyl-terminal region of Emp24p. Reporter molecules were constructed by replacing parts of a control invertase-Wbp1p chimera with those of Emp24p, and their transport rates were assessed. The transport of the reporter was found to be accelerated by the presence of the cytoplasmic domain of Emp24p. Mutational analyses revealed that the two carboxyl-terminal residues, leucine and valine (LV), were necessary and sufficient to accelerate the transport. The acceleration was sequence specific, and the terminal valine appeared to be more important. The LV residues accelerated not only the overall transport to the vacuole but also the ER to cis-Golgi transport, suggesting its function in the ER export. Hence the LV residues are a novel anterograde transport signal. The double-phenylalanine residues did not affect the transport by itself but attenuated the effect of the anterograde transport signal. On the other hand, the transmembrane domain significantly slowed down the ER to cis-Golgi transport and effectively counteracted the anterograde transport signal at this step. It may also take part in the retrieval of the protein, because the overall transport to the vacuole was more evidently slowed down. Consistently, the mutation of a conserved glutamine residue in the transmembrane domain further slowed down the transport in a step after arriving at the cis-Golgi. Taken together, the existence of the anterograde transport signal and the elements that regulate its function support the active recycling of Emp24p.
INTRODUCTION
The endoplasmic reticulum (ER)1 is the first destination for most newly synthesized secretory and membrane proteins. The proteins are inserted into the lumen or the membrane of the ER and transported to their final destinations by vesicular transport traversing several membrane-bound organelles, including the Golgi apparatus. The molecular mechanisms of vesicular transport have been extensively described (for review, see Schekman and Orci, 1996).
Transport between the ER and the Golgi is mediated by coatomer protein I (COP I)- and COP II-coated vesicles (Schekman and Orci, 1996). Recent experiments in yeast and mammalian cell systems suggest that the export of cargo from the ER is mediated by COP II (Barlowe et al., 1994; Aridor et al., 1995), whereas COP I mainly participates in the recycling of proteins back to the ER (Cosson and Letourneur, 1994; Letourneur et al., 1994). Participation of COP I in the cargo export from the ER is controversial (Bednarek et al., 1995; Schekman and Orci, 1996).
Proteins were thought to be transported out of the ER by default unless they have a specific retention or targeting signal. Now, accumulating evidence supports the existence of a cargo selection mechanism during the export from the ER (Schekman and Orci, 1996): proteins may be actively packaged into or excluded from budding vesicles at the ER (Kuehn and Schekman, 1997).
Emp24p is the first protein reported to function in active cargo selection. It is a type I membrane protein with a large luminal domain, a transmembrane domain (TMD), and a short cytoplasmic tail domain and was found to be enriched in COP II-coated vesicles isolated from yeast cells (Schimmöller et al., 1995). Deletion of the gene EMP24 is not lethal, indicating the preserved function of a basic level of the secretory pathway, but slows the transport of a subset of proteins (Schimmöller et al., 1995). Based on these facts, Emp24p was proposed to function in selective cargo export from the ER.
An independent approach also suggested the involvement of Emp24p in cargo selection. EMP24 is identical to BST2, which was identified as one of the three mutations that negated the lethal phenotype of the disruption of SEC13, a gene encoding a component of the COP II coat. In the bst2 mutant, the secretion of a cargo protein (invertase) was delayed, whereas ER resident proteins (Kar2p and Pdi1p) were missecreted (Elrod-Erickson and Kaiser, 1996). Such a phenotype implies that Emp24p plays a role not only in the export of cargo but also in the retention of ER proteins.
Erv25p is also a protein found to be enriched in COP II-coated vesicles isolated from yeast cells (Belden and Barlowe, 1996). Disruption of the gene ERV25 showed a similar selective transport defect as that of EMP24. Emp24p and Erv25p showed significant sequence homologies but were not functionally interchangeable. Emp24p and Erv25p were efficiently incorporated into COP II-coated vesicles only when both of them were present. A cross-linking experiment suggested that they interact physically. From these results, Emp24p and Erv25p were proposed to function as a complex (Belden and Barlowe, 1996).
Searches of DNA sequence databases revealed many other proteins to be similar in sequence to Emp24p; these are now collectively called the “p24 family” of proteins (Fiedler et al., 1996). Among these are CHOp24 and p23, which are enriched in COP I-coated vesicles isolated from mammalian cells (Blum et al., 1996; Sohn et al., 1996; Stamnes et al., 1995). Both CHOp24 and p23 have double-lysine and double-phenylalanine signals that can directly bind to COP I coatomer and have been suggested to be actively packaged into COP I-coated vesicles (Fiedler et al., 1996; Sohn et al., 1996).
To understand the precise role of Emp24p in cargo selection, the mode of Emp24p transport between the ER and the Golgi should first be clarified. Although Emp24p is enriched in COP II-coated vesicles, the majority of the protein exists in the ER (Schimmöller et al., 1995). Thus, Emp24p must somehow be transported back to the ER, after the transfer of cargo, which is in the same transport vesicle to the cis-Golgi. Interestingly, sequence similarities between yeast Emp24p and mammalian CHOp24 and yeast Erv25p and mammalian p23 suggest that these are putative functional homologues. If so, they may be packaged in both COP I- and COP II-coated vesicles and actively cycle between the ER and the Golgi.
For Emp24p to be actively recycled between the ER and the Golgi we reasoned that the protein must contain one or more, as yet unidentified, signals, and we sought to characterize such signals. When we started our work, several retrograde transport signals (ER retrieval signals) were already identified for transmembrane proteins. First was the carboxyl-terminal HDEL tetrapeptide signal in the luminal domain of Sec20p (Sweet and Pelham, 1992), which drives the Erd2p-dependent retrieval of the protein (Lewis et al., 1990). Second was the cytoplasmic double-lysine signal (Jackson et al., 1993; Gaynor et al., 1994). This signal directly binds to COP I coatomer for its retrieval (Cosson and Letourneur, 1994; Letourneur et al., 1994). Third was the cytoplasmic double-arginine signal (Schutze et al., 1994). Finally there was the TMDs of Sec12p and Sed4p (Sato et al., 1995; Sato et al., 1996), which drive Rer1p-dependent retrieval of the proteins.
While our work was in progress, the existence of anterograde transport signals emerged. These included the double-phenylalanine signal of the p24 family proteins (Fiedler et al., 1996), the similar double-phenylalanine signal at the carboxyl-terminus of ERGIC-53 (Kappeler et al., 1997), and the diacidic signal of the vesicular stomatitis virus glycoprotein and lysosomal acid phosphatase (Nishimura and Balch, 1997). Meanwhile, it was shown that Sec71p and Sec63p are also localized in the ER by an Rer1p-dependent pathway (Sato et al., 1997). It was also shown that Ufe1p is localized in the ER in a TMD-dependent but Rer1p-independent manner (Rayner and Pelham, 1997).
With these examples in mind, we focused our attention on the TMD and the cytoplasmic domain of the Emp24p with a hope of finding transport signal(s) that support the recycling of Emp24p between the ER and the Golgi. Here we report the identification of the elements (transport signals) that affect the transport rate of reporter proteins and presumably regulate the transport of Emp24p.
MATERIALS AND METHODS
Antibodies
Anti-invertase, α1,6-mannose and α1,3-mannose antibodies were described previously (Nishikawa and Nakano, 1993; Yamazaki et al., 1997).
Yeast Strain and Media
Yeast strain, SEY6210 (MATα leu2-3, 112 ura3-52 his3Δ200 trp1-Δ901 lys2-801 suc2-Δ9 GAL+) was kindly provided by Dr. Scott Emr (University of California, San Diego, CA). Cells were grown in YPD or SD (Kaiser et al., 1994) supplemented with 0.5% casamino acid (Difco Laboratories, Detroit, MI), 0.01% adenine sulfate, and 0.05% tryptophan (SDC-Ura−) for Ura+ screening. Supplemented minimal medium (Kaiser et al., 1994), from which methionine and uracil were depleted (SMM-Ura−-Met−), was used for [35S]methionine labeling.
Plasmid Construction
The construction of the invertase-Wbp1p chimera was described previously (Yamazaki et al., 1997). The DNA fragment encoding the carboxyl-terminal 84 amino acids of Emp24p was obtained from the yeast genome by PCR using an appropriate pair of oligonucleotide primers. The DNA fragments encoding chimeric proteins with Emp24p and their mutants were constructed by PCR using appropriately synthesized pairs of oligonucleotides with EcoRI sites at both ends and cloned into the EcoRI site of yeast multicopy expression vector pFo (Yamazaki et al., 1997). The sequences of all the constructs made by PCR were confirmed. Plasmids were introduced to SEY6210, and the transformants were selected by Ura+ expression.
Pulse–Chase Experiments
Cells were cultured in 2 ml of SDC-Ura− medium overnight at 30°C, diluted with 4 ml fresh medium, and further cultured for 2 h. Cells were then collected by centrifugation (300 × g for 2 min), washed once with 2.5 ml of SMM-Ura−-Met−, and incubated with 5 ml of SMM-Ura−-Met− for 1 h. Cells were then pulse labeled by adding 25 μl (9.25 MBq) of [35S]methionine and chased by adding 75 μl of chase solution (0.4% methionine and 0.3% cysteine in distilled water) at 30°C with continuous shaking. Aliquots (1 ml) were taken at appropriate time intervals and immediately mixed with 150 μl alkali-lysis solution (7.2% NaOH and 7.4% β-mercaptoethanol). Total proteins were precipitated with 80 μl of 100% trichloroacetic acid, washed once with acetone, and extracted with 400 μl of 1% SDS in TEN (50 mM Tris-HCl, pH 8.0, 5 mM EDTA, 150 mM NaCl) with brief sonication to dislodge the pellet and 5 min incubation at 95°C. Samples were diluted with 1 ml of 2% Triton X-100 in TEN containing 50 μg/ml α2-macroglobulin, 0.1% trasyrol, and 1 mM PMSF and centrifuged at 10,000 × g for 5 min to remove insoluble material. Anti-invertase antibodies (∼2 μl) and a 10-μl bed volume of protein A-Sepharose beads (Pharmacia Biotech, Uppsala, Sweden) were added and incubated for 2 h at room temperature. Beads were washed once each with immunoprecipitation (IP) buffer (0.2% SDS, 1% Triton X-100 in TEN), urea buffer (2 M urea in IP buffer), high-salt buffer (500 mM NaCl in IP buffer), and TEN sequentially. Proteins were eluted in 1% SDS in TEN (100 μl) with 95°C, 5-min incubation, split into aliquots, and immunoprecipitated again with specific antibodies (anti-invertase, anti-α1,6, or anti-α1,3 mannose antibodies) by the same procedure described above. Proteins were again eluted in 20 μl of denaturing buffer (0.5% SDS, 1% β-mercaptoethanol) with 95°C, 5-min incubation, added with 2 μl of 0.5 M sodium citrate (pH 5.5) and 0.5 μl of endoglycosidase H (Endo Hf; New England Biolabs, Beverley, MA), and incubated at 37°C for 1 h. Samples were analyzed by SDS-PAGE and autoradiographed with the BioImage BAS2000 analyzer (Fuji Photo Film, Tokyo, Japan).
Calculation of Transport Rate Constants
For the transport to the vacuole, samples were analyzed after 0 and 30 min of chase, and the 72-kDa band (full length form, S) and the 62-kDa band (processed form, P) were densitometrically quantified from autoradiograms. For the transport from the ER to the cis-Golgi, samples were analyzed after 0 and 5 min of chase, and the full-length form precipitated by anti-invertase antibody (total proteins, S + P) and that by anti-α1,6 antibody (P) were densitometrically quantified from autoradiograms.
The degree of transport (T) was calculated by the following equation:
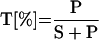 |
1 |
Rate constants (kv) were calculated by the following equation derived from Eq. 4 in RESULTS:
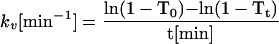 |
2 |
T0 and Tt are T at 0 and t min.
RESULTS
Quantitative Analysis of Protein Transport to the Vacuole
In this work, we used an invertase-Wbp1p chimera as a reporter molecule to measure the transport rate from the ER to the Golgi and that through the Golgi to the vacuole. This chimeric protein is proved to be useful for monitoring the transit of a cargo protein through distinct Golgi compartments (Gaynor et al., 1994; Yamazaki et al., 1997).
The ER to cis-Golgi transport is monitored by α1,6-mannosylation of core oligosaccharides, which is mediated by Och1p (Nakayama et al., 1992). Passage through the medial- and the trans-Golgi is monitored by further elongation of α1,6-mannose residues (Gaynor et al., 1994) and Mnn1p-mediated α1,3 mannosylation (Graham and Emr, 1991), respectively. Finally, arrival at the vacuole is monitored by proteolytic processing of the reporter protein, which is mediated by Pep4p-dependent protease (Gaynor et al., 1994).
A reporter protein that has a double-lysine, ER retrieval signal arrives at the cis-Golgi compartment at a rate similar to that of a reporter protein in which the retrieval signal was disrupted. But it is efficiently retrieved back to the ER before arriving at the medial-Golgi compartment and arrives at the vacuole very slowly (Gaynor et al., 1994).
To see whether Emp24p has a similar retrieval signal, we constructed reporter molecules based on the invertase-Wbp1p chimera by replacing parts of the carboxyl-terminal region with corresponding parts of Emp24p (Figure 1) and analyzed the overall transport rate to the vacuole.
Figure 1.

Schematic representation of the chimeras. (A) All of the constructs were generated based on the invertase-Wbp1p chimera. Open boxes, hatched boxes, and shaded boxes indicate invertase, Wbp1p, and Emp24p sequences, respectively. Characters in circles indicate the key residues and their mutations: K, KK; S, SS; N, TN; F, FF; A, AA; and L, LV. (B) Amino acid sequences of the transmembrane and the cytoplasmic domains of the constructs. Mutations in key residues are indicated by small letters.
Yeast cells expressing a reporter protein were pulse labeled with [35S]methionine for 10 min and then chased for 60 min. Cell extracts were prepared at several time points and immunoprecipitated with anti-invertase antibodies. To observe the proteolytic processing of the core protein moiety, N-linked oligosaccaride side chains were removed by treatment with endoglycosidase H (Endo H). As shown before, the full-length ER-Golgi form (∼72 kDa) first appeared and was then gradually processed to yield the vacuolar form (∼62 kDa) with a different rate for each reporter (Figure 2A).
Figure 2.

Transport to the vacuole is accelerated by the presence of the cytoplasmic domain of Emp24p. (A) Autoradiograms of the pulse–chase experiments. Reporter proteins (indicated on the right; see Figure 1) were expressed in yeast cells, pulse labeled for 10 min with [35S]methionine, added with excess methionine, and chased up to 60 min. Cells were lysed at indicated time points during the chase, and the labeled reporter proteins were immunoprecipitated by anti-invertase antiserum. Samples were Endo H treated and analyzed by autoradiography after SDS-PAGE (see MATERIALS AND METHODS for details). Labeled full-length forms (arrows) and processed forms (arrowheads) of the reporter proteins were indicated. (B) Quantitation of the transport to the vacuole. For each sample, the amount of a full-length form and a processed form were densitometrically quantified from an autoradiogram. The degree of transport (T) was calculated as described in MATERIALS AND METHODS and plotted against chase time for each reporter. (C) Logarithmic plots of the degree of remaining substrate for transport (1−T). All the reporters gave linear profiles up to 60 min of chase. (D) Rate constants of transport to the vacuole. The rate constants (kv) were calculated as described in MATERIALS AND METHODS. Bars, mean values; error bars, SD. The numbers of experiments are shown on top of the bars.
A reporter, which has the original transmembrane and cytoplasmic domains of Wbp1p (WWW), was efficiently retrieved back to the ER before arriving at the vacuole and accumulated as the full-length ER-Golgi form (Figure 2A). In contrast, when the ER retrieval signal was disrupted by replacing double-lysine residues with serine residues (WWS), the reporter protein was not retrieved but transported to the vacuole and converted to and accumulated as a vacuolar form (Figure 2A).
Amounts of the two forms of the reporter protein (72 and 62 kDa) in each sample were densitometrically quantified. The degree of transport to the vacuole (T) was expressed as the ratio of the processed form (62 kDa; P) to the total proteins (72 + 62 kDa; S + P) to minimize the effect of variation in the recovery of the proteins by immunoprecipitation. Based on the reduction of molecular weight by vacuolar processing, 1∼3 of 13 methionine and cysteine residues are predicted to be removed. Therefore the reduction of the radioactivity by the processing is 77∼92%, and this was negligible for the following analyses. The plots of the degree of transport against chase time gave simple saturation curves for all reporters except WWW (Figure 2B). A nonspecific protein band appeared at the 62-kDa position, and this gave ∼20% background.
Because the logarithmic plots of the degree of remaining substrates for transport (1 − T) gave linear profiles (Figure 2C), the following equations of a first-order reaction were applicable:
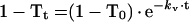 |
3 |
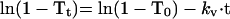 |
4 |
This allowed us to calculate a transport rate constant (kv) for each reporter (Figure 2D). The logarithmic plots usually gave a linear profile up to 30 min of chase. Therefore, for practical reasons, we only measured the degree of the transport (T) at chase 0 and 30 min to calculate the rate constants (kv) for the rest of experiments.
When the transmembrane and cytoplasmic domains were replaced with those of Emp24p (WEE), the reporter was transported at a similar but slightly reduced transport rate in comparison with the control, WWS (Figure 2D). However, when the TMD and the cytoplasmic domain were replaced separately, they showed clear and opposing effects. The cytoplasmic domain (WWE) increased the transport rate by 100%, whereas the TMD (WES) decreased it by 50%. These results suggested that Emp24p does not use a strong ER retrieval signal like the double-lysine signal. The TMD may function as an ER retrieval signal, although the efficiency was rather low (discussed later).
Interestingly, the cytoplasmic domain of Emp24p accelerated the overall transport to the vacuole. There was a possibility that the cytoplasmic domain of Wbp1p, in which the double-lysine signal was mutated to serine (WWS), still had a signal to slow down the transport. So the effect of the Emp24p cytoplasmic domain could merely be the result of the removal of this signal. However, this did not appear to be the case, because the reporter, in which the entire cytoplasmic domain was removed (WW−), was transported with a rate similar to that of WWS (Figure 2D). Therefore, the cytoplasmic domain of Emp24p was thought to contain an anterograde transport signal, although the possibility that the cytoplasmic domain of Emp24p somehow reduced the retrograde transport of the chimeric protein could not be excluded.
Quantitative Analysis of ER to cis-Golgi Transport
We next analyzed the rate of ER to cis-Golgi transport by monitoring α1,6-mannose modification. Shorter pulse (2 min) and chase (5 min) experiments were performed. Cell extracts were prepared, and the reporter protein was immunoprecipitated first with anti-invertase antibodies. Then, bound proteins were eluted, split into two equal aliquots, and reimmunoprecipitated with either anti-invertase or anti-α1,6-mannose antibodies. Bound proteins were eluted, Endo H treated, and analyzed as above (Figure 3A).
Figure 3.

ER to cis-Golgi transport is accelerated by the cytoplasmic domain of Emp24p. (A) Autoradiograms of the pulse–chase experiments. The cells expressing the indicated reporters were pulse labeled with [35S]methionine for 2 min and then chased for 5 min. Extracts were prepared at 0 and 5 min after the chase and immunoprecipitated with anti-invertase antibodies. The bound proteins were eluted, separated into two equal aliquots, and immunoprecipitated again with anti-invertase antibodies (Inv) or with anti-α1,6-mannose antibodies (α1,6). Proteins were eluted, treated with Endo H, and analyzed by SDS-PAGE followed by autoradiography. Arrows on the left indicate the full-length form. (B) Rate constants of transport from the ER to the cis-Golgi. The rate constants (kv) were calculated as described in MATERIALS AND METHODS. Bars, mean values; error bars, SD. The numbers of experiments are shown on top of the bars.
A quantitative analysis, similar to the case of the transport to the vacuole, was introduced. The amount of the 72-kDa protein precipitated by anti-α1,6 antibody (P) and anti-invertase antibody (S + P) was densitometrically quantified. The degree of ER to cis-Golgi transport (T) was expressed as the ratio of the former to the latter. The logarithmic plots of the degree of remaining substrates for transport (1 − T) against chase time gave linear profiles up to 5 min of chase (our unpublished results), indicating that the equations of a first-order reaction (Eqs. 3 and 4) were also applicable to this transport step. This again allowed us to calculate the transport rate constants (kv), which are presented in Figure 3B.
In accordance with the report by Gaynor et al. (1994), the rate constants of ER to cis-Golgi transport were almost the same for WWW and WWS (Figure 3B), thus confirming their conclusion that the reporter proteins were exported from the ER and arrived at the cis-Golgi at similar rates irrespective of the presence or absence of the double-lysine, ER retrieval signal. Moreover, the reporter with the signal was retrieved after arriving at the cis-Golgi.
Strikingly, the cytoplasmic domain of Emp24p increased the rate of ER to cis-Golgi transport by ∼120% (Figure 3B, compare WWE with WWS). Again, this is not the effect of the removal of the Wbp1p cytoplasmic domain, because the complete removal of the cytoplasmic domain (WW-) did not alter the transport rate significantly.
Retrieval of the protein before arriving at the cis-Golgi and receiving α1,6-mannose modification is unlikely, because such effective retrieval of the protein by the double-lysine signal would have affected the rate of α1,6-mannose modification (compare WWW with WWS). Therefore the accelerated ER to cis-Golgi transport must be caused by the accelerated anterograde transport from the ER to the cis-Golgi.
In contrast, the TMD of Emp24p (WES) significantly decreased the rate of ER to cis-Golgi transport (35% on average). Again, because the accelerated retrieval of the protein is unlikely, the TMD must reduce the rate of anterograde transport from the ER to the cis-Golgi.
The existence of the TMD of Emp24p strongly counteracted the cytoplasmic domain and no acceleration was seen with WEE (21% decrease). Therefore the effect of the TMD is dominant in the ER to cis-Golgi transport.
Identification of the Anterograde Transport Signal in the Cytoplasmic Domain
Mutational analyses were performed to identify the anterograde transport signal in the cytoplasmic domain of Emp24p (see Figure 1).
First, double-phenylalanine residues, which were reported to bind COP I-coatomer and have been implicated in anterograde transport (Fiedler et al., 1996), were changed to alanine residues (Figure 4, A and B, compare WWE with WWA). Unexpectedly, the mutation increased the transport rate (1.7-fold in overall vacuolar transport, 1.8-fold in ER to cis-Golgi transport). This clearly showed that the double-phenylalanine residues were not the anterograde transport signal. Instead, they were counteracting the anterograde transport signal (discussed later).
Figure 4.

Carboxyl-terminal two residues function as an anterograde transport signal. Mutants were made as indicated in Figure 1. Pulse–chase experiments were done, and transport rate constants were calculated as described for Figures 2 and 3. (A) Rate constants of transport to the vacuole. (B) Rate constants of ER to cis-Golgi transport. Bars, mean values; error bars, SD. The numbers of experiments are shown on top of the bars.
Second, the carboxyl-terminal two hydrophobic residues (leucine and valine, LV) were changed to the corresponding carboxyl-terminal hydrophilic residues from Wbp1p (threonine and asparagine, TN). The potential importance of these residues was noticed because their hydrophobicity of these residues is the only evident difference from the control, WWS (see Figure 1B). Also, the hydrophobic character of the two carboxyl-terminal residues is somewhat conserved among the p24 family members (Table 1). As expected, the conversion of the two carboxyl-terminal residues (LV) to hydrophilic residues (TN) abolished the acceleration of the transport (Figure 4, A and B, compare WWE and WWN; WWA and WWAN). This clearly showed that the two carboxyl-terminal residues (LV) were necessary to accelerate the anterograde transport.
Table 1.
Sequence comparison of the cytoplasmic domain of the yeast Emp24p family proteins
| Protein | Sequencea | Accession no. |
|---|---|---|
| Emp24p | RRFFEVTSLV | X67317 |
| yp24b | QFLFTGRQKNYV | L22015 |
| yp24c | KNFFVKQKVV | U00059 |
| yp24d | GKFFVKQKIL | L22015 |
| yp24e | QFFFTSRQKNYV | X87331 |
| yp24f | EFIFRESRKHNV | T36996 |
| Erv25p | KNYFKTKHII | Z49810 |
| yp24h | RSFFVKQKVL | Z48432 |
Conserved carboxyl terminal hydrophobic residues are indicated as bold letters.
Finally, the two carboxyl-terminal residues of Emp24p (LV) were transplanted onto a control reporter (WWS; see Figure 1) to confirm their function (Figure 4, A and B, WWL). As expected, this mutation increased the transport rate to a similar level with WWA, indicating that these two residues are enough to accelerate the anterograde transport. Mutation of the two carboxyl-terminal residues to serines (WWSS) showed no effect, confirming that the removal of TN was not responsible for the acceleration. Therefore we conclude that the two carboxyl-terminal residues (LV) are indeed the anterograde transport signal.
All of the mutations showed similar effects on both overall transport to the vacuole and ER to cis-Golgi transport (discussed later).
Characterization of the Anterograde Transport Signal
Further mutational analyses were performed to determine the sequence specificity of the two carboxyl-terminal residues (LV) for the acceleration of anterograde transport. To quantify the effects of the mutations, relative acceleration of transport was calculated by setting the transport rate constant of WWL (LV) as 100% and that of WWSS (SS) as 0% (Figure 5, values on top of the bars).
Figure 5.

Importance of the leucine and valine residues for the anterograde transport signal. The terminal two residues of the control reporter (WWS) were mutated as indicated at the bottom by the single-letter amino acid code. LV, WWL; SS, WWSS. Pulse–chase experiments were done, and transport rate constants were calculated as described for Figures 2 and 3. (A) Rate constants of transport to the vacuole. (B) Rate constants of ER to cis-Golgi transport. Bars, mean values of three experiments; error bars, SD. The values on top of the bars indicate the relative acceleration of the transport (%).
Mutation of either of the terminal LV residues to serine diminished the acceleration of transport (Figure 5, A and B, compare LS and SV with LV). The mutation of valine (LS) had a more severe effect; it showed only a marginal increase in the transport rates (8% acceleration in overall transport to the vacuole and 10% in ER to cis-Golgi transport). On the other hand, the mutation of leucine (SV) had less severe effects (40% acceleration in overall transport to the vacuole and 33% in ER to cis-Golgi transport).
Reversing the order of the leucine and the valine (VL) also diminished the accelerating effect (20% acceleration in overall transport to the vacuole and 12% in ER to cis-Golgi transport), confirming the importance of the terminal valine to enhance anterograde transport.
The result of the VL mutation implied that the hydrophobic nature was not enough to form an anterograde transport signal. In accordance with this notion, alanines, leucines, isoleucines, and phenylalanines (AA, LL, II, and FF, respectively) did not increase the transport rate as highly as LV (12, 27, 24, and 12% acceleration, respectively, in overall transport to the vacuole and 9, 12, 19, and 0%, respectively, in ER to cis-Golgi transport). Only two valines supported the efficient increase in the transport rate (VV; 52% acceleration in overall transport to the vacuole and 37% in ER to the cis-Golgi transport).
These results strongly suggested that both the leucine and valine residues were required for the efficient acceleration of the anterograde transport and the terminal valine was more critically required.
Importance of the Glutamine Residues in the TMD
Fiedler and Rothman (1997) noticed that the primary amino acid sequence of the TMD was significantly conserved among the p24 family proteins. They showed that mutations of the conserved charged or polar residues (glutamic acid and glutamine) in the transmembrane domain of CHOp24 influenced the transport rate (Fiedler and Rothman, 1997). Therefore, we next tried to find the significance of these residues in Emp24p transport. It was noted that the first position (glutamic acid) was not well conserved in p24 family members, and Emp24p has a glutamine residue at that position.
We mutated two glutamine residues in the TMD of the WES constructs and analyzed the transport rates (Figure 6). The existence of either of the glutamine residues supported the reduction of both the overall transport rate to the vacuole and the ER to cis-Golgi transport rate (Fig. 6, A and B; compare QQ, AQ, and QA with WWS). However, mutation of both glutamine residues to alanine increased the transport rate close to the control level (compare AA with WWS). These results indicated that either of the two glutamine residues was required for reducing the transport rate.
Figure 6.

Importance of the glutamine residues in the TMD. (A) Glutamine residues in the TMD of WES construct (QQ) were mutated to alanines. The positions of the mutated residues are indicated (A) underneath the original Emp24p transmembrane sequence. The names of the constructs are indicated on the left. (B) Rate constants of transport to the vacuole. (C) Rate constants of ER to cis-Golgi transport. Pulse–chase experiments were done, and transport rate constants were calculated as described for Figures 2 and 3. Bars, mean values; error bars, SD. The numbers of experiments are shown on top of the bars.
Single mutation of the second glutamine (compare QA with QQ) clearly reduced the overall transport rate to the vacuole (Figure 6A), although it did not affect the rate of ER to cis-Golgi transport (Figure 6B). This result suggested that the mutation of the second glutamine reduced the transport rate in a step after arriving at the cis-Golgi. In contrast, single mutation of the first glutamine (compare AQ with QQ) did not show any significant effects on both the overall transport to the vacuole (Figure 6A) and the ER to cis-Golgi transport (Figure 6B).
Taken together, these results strongly suggested that the glutamine residues do indeed play important roles in reducing the transport rate.
DISCUSSION
As an initial step to understand the mode of Emp24p transport between the ER and the Golgi, we sought to identify transport signal(s) on Emp24p. By constructing chimeric reporter proteins and measuring their transport rates, we could identify several elements that potentially regulate the transport of Emp24p.
A Novel Anterograde Transport Signal
The carboxyl-terminal two amino acid residues, leucine and valine (LV), form a novel anterograde transport signal. This was confirmed by three experiments. First, the transport was accelerated when the cytoplasmic domain of the control reporter (WWS) was replaced by that of Emp24p (Figure 2 and 3, WWE). Second, the acceleration was abolished when the terminal residues were mutated to the corresponding residues of the control reporter, threonine and asparagine (TN) (Figure 4, A and B, WWN). Finally, the accelerating effect was successfully transplanted to the control reporter (WWS) just by placing the two terminal residues (LV) (Figure 4, A and B, WWL).
Both leucine and valine were necessary for the full acceleration of the anterograde transport, and the terminal valine appeared to be the more important. The acceleration of the transport was suggested to be sequence specific because other hydrophobic amino acid residues could not support the full acceleration of the anterograde transport. It is interesting to note that the terminal valine was conserved in five of eight members of the yeast p24 family proteins (Table 1).
The ER to the cis-Golgi transport was remarkably accelerated by the presence of this anterograde transport signal (Figures 3, 4B, and 5B). This must have resulted from the accelerated anterograde transport. An alternative explanation would be the reduced retrieval of the protein before arriving at the cis-Golgi. However, no evidence for the existence of such a retrieval system has so far been reported. Our results also support the absence of such a retrieval system. The ER to cis-Golgi transport was not at all affected even in the presence of the effective double-lysine ER-retrieval signal (Figure 3, compare WWW with WWS).
Concerning the function of Emp24p in cargo selection at the ER, the acceleration of the anterograde transport is likely to be caused by accelerated export from the ER, i.e., active incorporation of the protein into budding vesicles. However, the possibility that the transport of budded vesicles to the cis-Golgi is accelerated, for example, by enhanced vesicle transfer or fusion to target membrane, cannot be ignored.
Kappeler et al. (1997) reported that the carboxyl-terminal double-phenylalanine residues promote COP II-binding and enhance the ER exit of ERGIC53. However, the carboxyl terminal double-phenylalanine residues did not enhance the ER exit of the reporter protein in our experimental system (Figure 5B). On the other hand, Nishimura and Balch (1997) reported that a diacidic signal (DXE; aspartic acid and glutamic acid residues separated by one amino acid residue) accelerates the anterograde transport and may serve as a COP II packaging signal. The reporters we used in this study do not have a diacidic signal, and thus the anterograde sorting signal we found here seems to be independent of the diacidic signal. Therefore, we conclude that the carboxyl-terminal LV residues are a novel anterograde transport signal.
Interestingly, the levels of the increase in the transport rates by various mutations were quite similar in overall transport to the vacuole and ER to cis-Golgi transport (Figures 3–5). The rates of overall transport to vacuole were one order slower than that of ER to cis-Golgi transport, and the kinetic profile of the overall transport to vacuole fitted well with a first-order reaction. This result implies the existence of a rate-limiting transport step that was independently but similarly controlled by the anterograde transport signal somewhere between the cis-Golgi and the vacuole. However, the possibility that the transport rate from the ER to the cis-Golgi somehow determines the overall transport rate to the vacuole without affecting the rate of later transport steps cannot be excluded. Because the overall transport to the vacuole is a complex process that is composed of multiple and bidirectional intercompartmental transport steps, a simple kinetic modeling may not be entirely applicable.
The Function of the Double-Phenylalanine Residues
Fiedler et al. (1996) reported that the double-phe-nylalanine residues are recognized by COP I components and function as an anterograde transport signal. Recently, Dominguez et al. (1998) reported that the double-phenylalanine residues can interact with Sec23p, a COP II component, and this may promote the exit of the protein from the ER. In either case, we would expect that the replacement of the double-phenylalanine residues with alanine residues would decrease the rate of ER to cis-Golgi transport. However, the replacement increased the transport rate in the presence of the terminal anterograde transport signal (Figure 4, compare WWE and WWA). The double-phenylalanine residues did not show any effects when the terminal anterograde transport signal was disrupted (compare WWN and WWAN). These results suggest that the double-phenylalanine residues modulate the function of the terminal anterograde transport signal but do not simply function as a transport signal themselves.
The recent report by Fiedler and Rothman (1997) on the effect of the double-phenylalanine residues is puzzling. When the double-phenylalanine residues were replaced by alanine in the presence of the wild-type TMD of CHOp24, the transport was slowed down. On the other hand, the transport was accelerated by the same replacement in the absence of the conserved glutamic acid and glutamine residues in the TMD. The latter result is consistent with ours, although the former result is not.
The discrepancy between our results and others may reflect the difference between yeast and mammalian transport systems.
The Function of the TMD of Emp24p
The TMD of the Emp24p significantly reduced the rate of ER to cis-Golgi transport (∼35%) by itself and restricted the action of the cytoplasmic anterograde transport signal (Figure 3). This must be caused by the reduced anterograde transport, because the accelerated retrieval of reporter proteins before arriving at the cis-Golgi was unlikely. It is also unlikely that the TMD directly reduces the transfer or fusion of the budded vesicles to the cis-Golgi. Therefore the TMD must function as a weak static ER retention signal and mask the function of the cytoplasmic anterograde transport signal before or during the budding.
The TMD of Emp24p reduced the overall transport rate to the vacuole (50%) more effectively than the rate of ER to cis-Golgi transport (Figure 2). Therefore the TMD may reduce the transport rate also in later transport steps. The mutation of the second conserved glutamine consistently reduced the overall transport rate to the vacuole, with no significant change in the ER to cis-Golgi transport rate (Figure 6, compare QQ with QA). This indicates a decrease in the transport rate after arriving at the cis-Golgi. This may be caused either by increased retrograde transport or by decreased anterograde transport of the protein in the cis-Golgi or following compartment(s).
Fiedler and Rothman (1997) reported that the conserved glutamic acid appeared to be a key determinant for the retention of the protein in the ER. The glutamine residue in the TMD and the double-phenylalanine residues in the cytoplasmic domain counteract the glutamic acid residue to enable the export of the protein from the ER. They also found that the transport of the reporter molecule was slowed down when the TMD alone was placed. Our results were basically consistent with this. The first glutamine residue effectively reduced the overall transport rate to the vacuole (Figure 6A, compare QA with AA), and the second glutamine residue counteracted this (compare QA with QQ).
Nickel et al. (1997) reported that p23 is recycled back to the ER by the double-lysine- and double-phenylalanine–dependent mechanism, and the TMD does not have such a significant effect on the transport. However, p23 has serine at the position where Emp24p has glutamine and CHOp24 has glutamic acid. Therefore it is consistent with the idea that the glutamine or glutamic acid at this position plays an important role for the function of the TMD.
The Regulation of the Transport of Emp24p
The existence of the anterograde transport signal on the cytoplasmic domain of Emp24p put forward the possibility that Emp24p actively cycles between the ER and the Golgi. However, this did not answer the question of how Emp24p is localized in the ER. It is obvious that Emp24p should be retrieved back to the ER if it is actively exported from the ER.
Because there was no retrieval signal in the cytoplasmic domain of Emp24p, the TMD or the luminal domain could serve for the retrieval. Although the TMD did not appear to be efficient enough to keep the protein in the ER, the luminal domain can help the function of the TMD. As discussed above, the TMD has a potential to reduce the transport after arriving at the cis-Golgi, and this can be caused by the retrieval of the protein. Therefore it is possible that the TMD takes part in the retrieval.
It was reported that Emp24p interacts with Erv25p, and Erv25p has double-lysine residues similar to the ER retrieval signal (Belden and Barlowe, 1996). Therefore, there is a good chance that the interaction with Erv25p through the luminal domain and/or the TMD promotes the retrieval of Emp24p to the ER. Alternatively, the TMD may interact with Rer1p for retrieval. From this point of view, the effects of the mutations of the conserved glutamine residues in the TMD could be explained by altered interaction of the TMD of Emp24p with the TMD of another transmembrane protein, namely Erv25p or Rer1p.
Why does Emp24p have several counteracting elements that accelerate (the anterograde transport signal, LV) or slow down (the double-phenylalanine residues and the TMD) the transport? Does the cytoplasmic anterograde transport signal really function in the authentic Emp24p molecule? Because the anterograde transport effect was so clearly seen in reporter constructs, we believe that it somehow functions in a regulating manner. These elements may be devices for regulating transport of the protein. For example, the elements may be presented differently in the ER and the Golgi by interactions with other molecules such as cargo. In other words, the binding of cargo to Emp24p (directly or indirectly) may accelerate the export of the protein from the ER, and their dissociation in the Golgi may facilitate the retrieval of the protein.
ACKNOWLEDGMENTS
We thank Dr. Scott Emr (University of California, San Diego, CA) for the yeast strains, Dr. Naomi Hachiya (Japan Science and Technology Corporation), Dr. Graham Warren and colleagues (Imperial Cancer Research Fund), Dr. Tommy Nilsson and colleagues (European Molecular Biology Laboratory), and all the members of Mihara lab for their helpful comments and discussions, Dr. Toru Komiya and Dr. Norman Hui for critical reading of the manuscript, and Chiaki Matsunaga for her technical assistance. Special thanks go to Yumi Kosai for excellent secretarial work. This work was partly supported by a Grant-in-Aid from the Ministry of Education, Science and Culture of Japan and a grant for the Core Research for Science and Technology from Japan Science and Technology Corporation. S.Y. was supported by a research fellowship from the Japan Society for the Promotion of Science.
Abbreviations used:
- COP
coatomer protein
- Endo H
endoglycosidase H
- ER
endoplasmic reticulum
- TMD
transmembrane domain
REFERENCES
- Aridor M, Bannkyh SI, Rowe T, Balch WE. Sequential coupling between Cop II and Cop I vesicle coats in endoplasmic reticulum to Golgi transport. J Cell Biol. 1995;131:875–893. doi: 10.1083/jcb.131.4.875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barlowe C, Orci L, Yeung T, Hosobuchi M, Hamamoto S, Salama N, Rexach MF, Ravazzola M, Amherdt M, Schekman R. COP II: a membrane coat formed by sec proteins that drive vesicle budding from the endoplasmic reticulum. Cell. 1994;77:895–907. doi: 10.1016/0092-8674(94)90138-4. [DOI] [PubMed] [Google Scholar]
- Bednarek SY, Ravazzola M, Hosobuchi M, Amherdt M, Perrelet A, Scheckman R, Orci L. COP I- and COP II-coated vesicles bud directly from the endoplasmic reticulum in yeast. Cell. 1995;83:1183–1196. doi: 10.1016/0092-8674(95)90144-2. [DOI] [PubMed] [Google Scholar]
- Belden WJ, Barlowe C. Erv25p, a component of COP II-coated vesicles, forms a complex with Emp24p that is required for efficient endoplasmic reticulum to Golgi transport. J Biol Chem. 1996;271:26939–26946. doi: 10.1074/jbc.271.43.26939. [DOI] [PubMed] [Google Scholar]
- Blum R, Feick P, Puype M, Vanderckhove J, Klengel R, Nastainczyk W, Schulz I. Tmp21 and p24A, two type I proteins enriched in pancreatic microsomal membranes, are members of a protein family involved in vesicular trafficking. J Biol Chem. 1996;271:17183–17189. doi: 10.1074/jbc.271.29.17183. [DOI] [PubMed] [Google Scholar]
- Cosson P, Letourneur F. Coatmer interaction with di-lysine endoplasmic reticulum retention motifs. Science. 1994;263:1629–1631. doi: 10.1126/science.8128252. [DOI] [PubMed] [Google Scholar]
- Dominguez M, Dejgaard K, Füllekrug J, Dahan S, Fazel A, Paccaud J-P, Thomas DY, Bergeron JJM, Nilsson T. gp25L/emp24/p24 protein family members of the cis-Golgi network bind both COP I and II coatomer. J Cell Biol. 1998;140:751–765. doi: 10.1083/jcb.140.4.751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elrod-Erickson MJ, Kaiser CA. Genes that control the fidelity of endoplasmic reticulum to Golgi transport identified as suppressors of vesicle budding mutations. Mol Biol Cell. 1996;7:1043–1058. doi: 10.1091/mbc.7.7.1043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiedler K, Rothman JE. Sorting determinants in the transmembrane domain of p24 proteins. J Biol Chem. 1997;272:24739–24742. doi: 10.1074/jbc.272.40.24739. [DOI] [PubMed] [Google Scholar]
- Fiedler K, Veit M, Stamnes MA, Rothman JE. Bimodal interaction of coatomer with the p24 family of putative cargo receptors. Science. 1996;273:1396–1399. doi: 10.1126/science.273.5280.1396. [DOI] [PubMed] [Google Scholar]
- Gaynor EC, te Heesen S, Graham TR, Aebi M, Emr SD. Signal-mediated retrieval of membrane protein from the Golgi to the ER in yeast. J Cell Biol. 1994;127:653–665. doi: 10.1083/jcb.127.3.653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham TR, Emr SD. Compartmental organization of Golgi-specific protein modification and vacuolar protein sorting events defined in a yeast sec18 (NSF) mutant. J Cell Biol. 1991;114:207–218. doi: 10.1083/jcb.114.2.207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson M, Nilsson T, Peterson PA. Retrieval of transmembrane proteins to the endoplasmic reticulum. J Cell Biol. 1993;121:317–333. doi: 10.1083/jcb.121.2.317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaiser C, Michaelis S, Mitchell A. Methods in Yeast Genetics. Plainview, NY: Cold Spring Harbor Laboratory Press; 1994. [Google Scholar]
- Kappeler F, Klopfenstein DRC, Fouguet M, Paccaud J-P, Hauri H-P. The recycling of ERGIC-53 in the early secretory pathway. J Biol Chem. 1997;272:31801–31808. doi: 10.1074/jbc.272.50.31801. [DOI] [PubMed] [Google Scholar]
- Kuehn MJ, Schekman R. COP II and secretory cargo capture into transport vesicles. Curr Opin Cell Biol. 1997;9:477–483. doi: 10.1016/s0955-0674(97)80022-1. [DOI] [PubMed] [Google Scholar]
- Letourneur F, Gaynor E, Hennecke S, Démollière C, Duden R, Emr S, Riezman H, Cosson P. Coatomer is essential for retrieval of dilysine-tagged proteins to the endoplasmic reticulum. Cell. 1994;79:1199–1207. doi: 10.1016/0092-8674(94)90011-6. [DOI] [PubMed] [Google Scholar]
- Lewis MJ, Sweet DJ, Pelham HRB. The ERD2 gene determines the specificity of the luminal ER protein retention system. Cell. 1990;61:1359–1363. doi: 10.1016/0092-8674(90)90699-f. [DOI] [PubMed] [Google Scholar]
- Nakayama K, Nagasu T, Shimma Y, Kuromitsu J, Jigami Y. OCH1 encodes a novel membrane bound mannosyltransferase: outer chain elongation of asparagine-linked oligosaccharides. EMBO J. 1992;11:2511–2519. doi: 10.1002/j.1460-2075.1992.tb05316.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nickel W, Sohn K, Böning C, Wieland F. p23, a major COP I-vesicle membrane protein, constitutively cycles through the early secretory pathway. Proc Natl Acad Sci USA. 1997;94:11393–11398. doi: 10.1073/pnas.94.21.11393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishikawa S, Nakano A. Identification of a gene required for membrane protein retention in the early secretory pathway. Proc Natl Acad Sci USA. 1993;90:8179–8183. doi: 10.1073/pnas.90.17.8179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishimura N, Balch WE. A di-acidic signal required for selective export from the endoplamic reticulum. Science. 1997;277:556–558. doi: 10.1126/science.277.5325.556. [DOI] [PubMed] [Google Scholar]
- Rayner JC, Pelham HRB. Transmembrane domain-dependent sorting of proteins to the ER and plasma membrane in yeast. EMBO J. 1997;16:1832–1841. doi: 10.1093/emboj/16.8.1832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato K, Nishikawa S, Nakano A. Membrane protein retrieval from the Golgi apparatus to the endoplasmic reticulum (ER): characterization of the RER1 gene products as a component involved in ER localization of Sec12p. Mol Biol Cell. 1995;6:1459–1477. doi: 10.1091/mbc.6.11.1459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato K, Sato M, Nakano A. Rer1p as common machinery for the endoplasmic reticulum localization of membrane proteins. Proc Natl Acad Sci USA. 1997;94:9693–9698. doi: 10.1073/pnas.94.18.9693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato M, Sato K, Nakano A. Endoplasmic reticulum localization of Sec12p is achieved by two mechanisms: Rer1p-dependent retrieval that requires the transmembrane domain and Rer1p-independent retention that involves the cytoplasmic domain. J Cell Biol. 1996;134:279–293. doi: 10.1083/jcb.134.2.279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schekman R, Orci L. Coat proteins and vesicle budding. Science. 1996;271:1526–1533. doi: 10.1126/science.271.5255.1526. [DOI] [PubMed] [Google Scholar]
- Schimmöller F, Singer-Krüger B, Schröder S, Krüger U, Barlowe C, Riezman H. The absence of Emp24p, a component of ER-derived COP II-coated vesicles, causes a defect in transport of selected proteins to the Golgi. EMBO J. 1995;14:1329–1339. doi: 10.1002/j.1460-2075.1995.tb07119.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schutze M-P, Peterson PA, Jackson MR. An N-terminal double-arginine motif maintains type II membrane proteins in endoplasmic reticulum. EMBO J. 1994;13:1696–1705. doi: 10.1002/j.1460-2075.1994.tb06434.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sohn K, Orci L, Ravazzola M, Amherdt M, Bremser M, Lottspeich F, Fiedler K, Helms JB, Wieland FT. A major transmembrane protein of Golgi-derived COP I-coated vesicles involved in coatomer binding. J Cell Biol. 1996;135:1239–1248. doi: 10.1083/jcb.135.5.1239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stamnes MA, Craighead MW, Hoe MH, Lampen N, Geromanos S, Tempst P, Rothman JE. An integral membrane component of coatomer-coated transport vesicles defines a family of proteins involved in budding. Proc Natl Acad Sci USA. 1995;92:8011–8015. doi: 10.1073/pnas.92.17.8011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweet DJ, Pelham HRB. The Saccharomyces cerevisiae SEC20 gene encodes a membrane glycoprotein which is sorted by the HDEL retrieval system. EMBO J. 1992;11:423–432. doi: 10.1002/j.1460-2075.1992.tb05071.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamazaki S, Harashima S, Sakaguchi M, Mihara K. Identification and functional characterization of yeast ζ-COP. J Biochem. 1997;121:8–14. doi: 10.1093/oxfordjournals.jbchem.a021574. [DOI] [PubMed] [Google Scholar]