

Abstract
Corticotropin releasing factor (CRF) is a neuropeptide expressed in micturition reflex circuitry and different roles in these reflexes have been suggested. These studies examined the expression of CRF/CRF receptors in the urinary bladder during postnatal development in the rat. Urinary bladder was harvested from rats (postnatal (P) day 0-adult) euthanized by isoflurane (4%) and thoracotomy. CRF protein expression significantly (p ≤ 0.01) decreased in the urothelium with increasing postnatal age. In contrast, CRF-immunoreactivity (IR) was increased in nerve fibers in the suburothelial plexus during the second-third postnatal week. Total CRF protein from urinary bladder significantly increased during the second-third postnatal weeks as determined with ELISAs. CRF receptor 2 (CRFR2) transcript was expressed in urinary bladder of all postnatal ages examined whereas no CRFR1 transcript was expressed at any postnatal age examined. We also demonstrated changes in urinary bladder mRNA expression for the neuropeptides, galanin, substance P, vasoactive intestinal polypeptide and pituitary adenylate cyclase activating polypeptide during postnatal development. These studies demonstrate changes in the CRF expression in urinary bladder, specifically in the urothelium and nerve fibers of the suburothelial plexus during postnatal development. Changes in CRF expression and neuropeptide expression in general in the urinary bladder may contribute to the emergence of mature voiding reflexes.
Keywords: urothelium, ELISA, suburothelial plexus, neuropeptides, development
Introduction
Corticotropin releasing factor (CRF) is prominently expressed in the descending pathway from Barrington’s nucleus to the sacral parasympathetic nucleus (SPN) in the lumbosacral spinal cord and CRF-immunoreactivity (IR) is present in the SPN of adult rats (Valentino et al., 1995; Valentino et al., 1996; Puder and Papka, 2001). Opposing roles for CRF in mature bladder function have been proposed (Pavcovich et al., 1998; Klausner and Steers, 2004; Million et al., 2007) but there is strong evidence that CRF in micturition reflexes is inhibitory. Intrathecal CRF inhibits bladder contraction elicited by chemical stimulation of Barrington’s nucleus (Pavcovich and Valentino, 1995). Conversely, CRF antagonists increase the amplitude of bladder contractions (Pavcovich and Valentino, 1995).
Recent studies have demonstrated a developmental (Studeny and Vizzard, 2005) as well as inflammatory-induced plasticity (LaBerge et al., 2006) of CRF-IR in micturition spinal reflex pathways and supraspinal locations (Mineta et al., 2004). We have demonstrated an age-dependent upregulation of CRF-IR in association with preganglionic parasympathetic neurons (Studeny and Vizzard, 2005) in the SPN. We suggested that upregulation of CRF-IR in bulbospinal projections to the SPN may contribute to mature voiding reflexes (Studeny and Vizzard, 2005) and play a role in continence. Cyclophosphamide (CYP)-induced cystitis increased CRF expression in the region of the SPN in the lumbosacral spinal cord and in the urinary bladder (LaBerge et al., 2006). In control urinary bladder, CRF receptor 2 (CRFR2) is expressed (LaBerge et al., 2006) and gene-profiling studies have demonstrated upregulation of CRF receptor in the urinary bladder with cystitis (Saban et al., 2002).
During the course of postnatal maturation, primitive reflex pathways organized at the spinal level are replaced by a spinobulbospinal reflex leading to emergence of voluntary voiding (de Groat and Araki, 1999). Postnatal maturation of voiding function may involve prominent reorganization of synaptic connections in bladder reflex pathways in the brain, spinal cord, and periphery. This reorganization leads to down-regulation of primitive spinal mechanisms and upregulation of mature supraspinal pathways (de Groat and Araki, 1999). Previous studies have suggested the importance of neuroactive compounds in the process of maturation of the micturition reflexes during prenatal and early postnatal development (Ekstrom et al., 1994; Iuchi et al., 1994; Sann et al., 1997). These changes in micturition reflexes during postnatal development may be mediated, in part, by changes in the neurochemical properties of central micturition pathways as well as by similar changes in the urinary bladder. In the present study, we have examined postnatal CRF expression in the urinary bladder. In postnatal and adult rats, we determined: (1) CRF protein expression in the whole urinary bladder by enzyme-linked immunoassays (ELISAs); (2) tissue expression of CRF in urinary bladder with an emphasis on urothelial cell and nerve fiber expression; (3) CRFR1 and CRFR2 transcript expression in urinary bladder during postnatal development; and (4) neuropeptide transcript expression in postnatal urinary bladder.
Materials and Methods
Adult female (3–4 months; 150 – 200 g) and postnatal (P) day 0 (birth)-36 Wistar rats were used in these studies (Charles River Canada; St. Constant, Canada). Pups of the same postnatal age were used from six separate litters. Postnatal rat pups of both genders were used in these studies. No differences in CRF expression in bladder between genders were demonstrated; therefore, data from both genders has been pooled for data analysis and presentation.
Tissue Harvesting, sample preparation and assay
Procedures for tissue processing and ELISAs were identical to those described in detail previously (LaBerge et al., 2006). Adult and postnatal rats were euthanized with isoflurane anesthesia (4%) and thoracotomy and the bladder (n = 6–8 for each age group) was rapidly dissected and weighed. Individual bladders were solubilized in Tissue Protein Extraction Reagent (T-PER; Pierce, Rockford, IL; 1 g tissue/20 ml) with Complete (protease inhibitors cocktail tablets; Roche Diagnostics GmbH, Germany). Bladder tissue was disrupted with a Polytron homogenizer and then centrifuged (10,000 rpm for 5 min). Total protein was determined by the Coomassie Plus Protein Assay Reagent Kit (Pierce). The supernatant was used for CRF quantification using commercially available rat specific CRF ELISA kits (Peninsula Laboratories, member of the Bachem group, San Carlos, CA) in accordance with the manufacturer’s instructions.
Rat CRF ELISA
Rabbit anti-rat CRF antibody was adsorbed to microtiter (R&D Systems, Inc., Minneapolis, MN) plates as described (LaBerge et al., 2006). After addition of the sample or standard solution, the second antibody (detection antibody) was applied. Sample and standard solutions were run in duplicate. This antibody complex was detected with a horseradish peroxidase-labeled immunoglobulin. Enzyme activity was quantified by the change in optical density, using tetramethyl benzidine as substrate. The CRF standard provided with this system generated linear standard curves from 0–10 ng/ml (R2 = 0.998, p ≤ 0.001). The absorbance values of standards and samples were corrected by subtraction of the background value (absorbance due to nonspecific binding). Samples were used undiluted to bring the absorbance values onto the linear portion of the standard curve. No samples fell below the minimum detection limits of the assay. Curve fitting of standards and evaluation of CRF content of samples was performed using a least squares fit.
Immunohistochemistry
Urinary bladder Adult and postnatal rats were euthanized as above and the bladder (n = 6 for each) was rapidly dissected and weighed. Urinary bladders were postfixed in 4% paraformaldehyde, placed in ascending concentrations of sucrose (10–30%), frozen, sectioned (25 µm) and mounted on gelled (0.5%) microscope slides for on-slide processing (Vizzard, 1997). Briefly, all sections were incubated overnight at room temperature with rabbit anti-CRF (1:3000; Phoenix Pharmaceuticals, Inc., Belmont, CA) in 1% goat serum and 0.1 M phosphate buffered saline (PBS), and then washed (3 × 10 min) with 0.1 M PBS, pH 7.4. The tissues were then incubated a Cy3-conjugated goat anti-rabbit secondary antibody in PBGT (1:500; Jackson Immunoresearch Laboratories, West Grove, PA) for 2 hr at room temperature. Following washing (3 × 10 min with PBS), the slides were coverslipped with Citifluor (Citifluor Ltd., London). Control sections incubated in the absence of primary or secondary antibody were also processed and evaluated for specificity or background staining levels. Some bladder sections were also stained with YOYO-1 (Molecular Probes, Eugene, Oregon; 1:10,000) to stain RNA and visualize cell nuclei. In the absence of primary antibody, no positive immunostaining was observed. Absorption of CRF-IR in bladder sections was also performed with CRF peptide (Phoenix Pharmaceuticals Inc.; 10 µg/ml). CRF-IR in tissue sections with immunoabsorption was significantly reduced and was not above background staining. Previous studies with the same CRF antibody demonstrated CRF-IR in the dorsal horn and autonomic nucleus in the lumbosacral spinal cord (Studeny and Vizzard, 2005) consistent with previous reports of CRF distribution (Vincent and Satoh, 1984; Puder and Papka, 2001).
Whole Mount Bladder Preparation
To determine if CRF-IR was present in the suburothelial nerve plexus, whole mount preparations of urinary bladder were prepared. Previous studies have demonstrated greater ability to see the suburothelial nerve plexus in wholemount preparations in comparison to sections of the urinary bladder (Zvarova and Vizzard, 2005a; LaBerge et al., 2006; Corrow and Vizzard, 2007). Adult and postnatal rats (n = 6 for each) were euthanized as above. The urinary bladder was dissected and placed in Krebs solution. The bladder was cut open along the midline and pinned to a sylgard-coated dish. The bladder was incubated for 1.5 hr at room temperature in cold fixative (2% paraformaldehyde + 0.2% picric acid) and urothelium removed as previously described (Zvarova and Vizzard, 2005a; LaBerge et al., 2006; Yuridullah et al., 2006; Corrow and Vizzard, 2007). While pinned, the bladder neck on one side was notched for orientation purposes and regional analyses of immunoreactivity. Using fine tips forceps and iris scissors, the urothelium was dissected from the underlying detrusor smooth muscle with the aid of a dissecting microscope. The urothelium has suburothelial structures associated with it; the term urothelium in this paper refers to both urothelial and suburothelial structures. Urothelium and bladder musculature were processed for CRF-IR as described above. As a comparison, some wholemount preparations were stained with the pan neuronal marker, protein gene product 9.5 (Abcam, Cambridge, MA; 1:15) to visualize all nerve fibers in the suburothelial plexus.
Assessment of Positively Stained Urinary Bladder Regions
Staining observed in experimental tissue was compared to that observed from experiment-matched negative controls. Urinary bladder sections exhibiting immunoreactivity that was greater than the background level observed in experiment-matched negative controls were considered positively stained. No autofluorescence was observed in any bladder section from any postnatal age examined.
Visualization and quantitative analysis of CRF–IR in urinary bladder sections
Six to 10 urinary bladder sections per urinary bladder were examined using an Olympus fluorescence photomicroscope (Optical Analysis Corp., Nashua, NH) with a multiband filter set for visualization of the Cy3 fluorophore. Cy3 was visualized with a filter with an excitation range of 560–596 nm and an emission range from 610–655 nm. YOYO-1 was visualized with a filter with an excitation range of 470–490 and an emission range from 510–530. Grayscale images acquired in tiff format were imported into Meta Morph image analysis software (version 4.5r4; Universal Imaging, Downingtown, PA) as previously described (LaBerge et al., 2006; Yuridullah et al., 2006; Corrow and Vizzard, 2007). The free hand drawing tool was selected and the urothelium was drawn (green outline) and measured in total pixels area (Fig. 1). A threshold encompassing an intensity range of 100–250 grayscale values was applied to the region of interest (Fig. 1) in the least brightly stained condition first. The same threshold was subsequently used for all images (yellow shading). Average intensity was calculated within the outlined area (Fig. 1, green outlines). The average intensity represents the average value of all of the pixels above the threshold value (Fig. 1, yellow shadings). Percent CRF expression above threshold in the total area selected was then calculated and averaged for all samples from adult (n = 6) and P0–P36 rats (n = 6 each). Percentage CRF expression in P0–P36 rats was expressed as a percentage of adult CRF expression.
Figure 1.

Semi-quantitative analysis of CRF-immunoreactivity (IR) in the urothelium during postnatal development. The urothelium was outlined (green) and measured in total pixels area (A–F). A threshold encompassing an intensity range of 100–250 grayscale values was applied to the region of interest (A–F). The same threshold was subsequently used for all images. Percent CRF expression above threshold in the total area selected was then calculated. Grayscale versions of CRF-IR in urinary bladder at birth, postnatal (P) day 0 (A), P7 (B), P14 (C), P21 (D), P28 (E) and adult (F) with the urothelium (U) outlined in green are demonstrated. Images are thresholded and little CRF (absence of yellow within the outlined region) is above threshold in adult bladder compared to significant CRF-IR that is above threshold at younger ages (A–E, presence of yellow within U). Calibration bar represents 50 µm.
RNA Extraction, Reverse Transcription and Polymerase Chain Reaction (PCR)
Urinary bladders were dissected from rats and total RNA from the different tissues was extracted using the STAT-60 total RNA/mRNA isolation reagent (Tel-Test “B”, Friendswood, TX) as previously described (Braas et al., 2006). Spinal cord (L6) was also dissected from rats and total RNA extracted as described for inclusion as a positive control for some PCR and quantitative PCR reactions (see below). The quality and quantity of the resulting RNA was assessed with spectrometry at 260/280 nm and 2 µg of total RNA from each sample was used to synthesize first strand cDNA using SuperScript II reverse transcriptase with the SuperScript II Preamplification System (Invitrogen, Carlsbad, CA) in a 20 µl final reaction volume. Following digestion with RNase H to remove residual RNA, the cDNA was amplified with 200 µM dNTP, 0.2 M primers and 1.25 U AmpliTaq Gold DNA Polymerase with the following parameters; initial denaturation, 94 °C, 5 min, denaturation 94 °C for 45 sec; annealing, primer-specific annealing temperature for 30 sec; extension 72 °C for 45 sec (30 – 35 cycles); final extension, 72 °C for 5 min. The oligonucleotide primers for CRF receptors (CRFR1 and CRFR2) were previously described (Wu et al., 2007). The amplified products were resolved on 2% agarose gels, stained with ethidium bromide, and visualized under UV illumination. Complementary DNA synthesis in the absence of RNA template or reverse transcriptase, or amplification without template, primers or DNA polymerase were used as controls.
Real-time quantitative reverse transcription-polymerase chain reaction (Q RT-PCR)
Urinary bladders from adult and postnatal rats were dissected under RNase-free conditions and total RNA was extracted using the STAT-60 total RNA/mRNA isolation reagent (Tel-Test ‘B’, Friendswood, TX, USA). Complementary DNA was synthesized using SuperScript II reverse transcriptase and random hexamer primers with the SuperScript II Preamplification System (Invitrogen, Carlsbad, CA, USA) in a 20 µl final reaction volume. The quantitative PCR standards for vasoactive intestinal polypeptide (VIP), substance P (sub P), galanin, pituitary adenylate cyclase activating polypeptide (PACAP) and 18S transcripts were prepared with the amplified cDNA products ligated directly into pCR2.1 TOPO vector using the TOPO TA cloning kit (Invitrogen). The nucleotide sequences of the inserts were verified by automated fluorescent dideoxy dye terminator sequencing (Vermont Cancer Center DNA Analysis Facility). To estimate the relative expression of the receptor transcripts, 10-fold serial dilutions of stock plasmids were prepared as quantitative standards. The range of standard concentrations was determined empirically. Real-time quantitative PCR was performed using SYBR Green I detection as previously described (Girard et al., 2002; Girard et al., 2006). Complementary DNA templates, diluted 5-fold to minimize the inhibitory effects of the reverse transcription reaction components, were assayed using SYBR Green I JumpStart™. Taq ReadyMix™ (Sigma, St. Louis, MO, USA) containing 5 mM MgCl2, 200 mM dATP, dGTP, dCTP and dTTP, 0.64 U Taq DNA polymerase and 300 nM of each primer in a final 25 µl reaction volume. The real-time quantitative PCR was performed on an Applied Biosystems 7500 Fast real-time PCR system (Applied Biosystems, Foster City, CA, USA) using the following standard conditions: (i) initial heating 94 °C for 2 min; (ii) amplification over 40 cycles at 94 °C for 15 seconds (s) and 65 °C or 64 °C for 30 s.
The amplified product from these amplification parameters was subjected to SYBR Green I melting analysis by ramping the temperature of the reaction samples from 60 °C to 95 °C. A single DNA melting prof ile was observed under these dissociation assay conditions demonstrating amplification of a single unique product, free of primer dimers or other anomalous products. Oligonucleotide primer sequences for VIP, galanin, and PACAP and the housekeeping gene, 18S, used in these studies were previously described (Girard et al., 2002). Selected forward (F) and reverse (R) primers for sub P were F: 5’-AGAGGAAATCGGTGCCAAC-3’, R: 5’- TGCCCATTAATCCAAAGAACTG-3’.
For data analyses, a standard curve was constructed by amplification of serially diluted plasmids containing the target sequence. Data were analyzed at the termination of each assay using the Sequence Detection Software version 1.3.1 (Applied Biosystems, Norwalk, CT). In standard assays, default baseline settings were selected. The increase in SYBR Green I fluorescence intensity (ΔRn) was plotted as a function of cycle number and the threshold cycle was determined by the software as the amplification cycle at which the ΔRn first intersects the established baseline. All data are expressed as the relative quantity of the gene of interest normalized to the relative quantity of the housekeeping gene 18S.
Statistics
All values are means ± S.E.M. Comparisons of CRF protein concentration in urinary bladder samples were made using analysis of variance. Percentage data from image analysis were arcsin transformed to meet the requirements of this statistical test. Animals, processed and analyzed on the same day, were tested as a block in the analysis of variance. Two variables were being tested in the analysis: (1) age and (2) the effect of day (i.e., tissue from different postnatal or adult groups of animals were processed on different days). When F ratios exceeded the critical value (p ≤ 0.05), the Newman-Keul’s post-hoc test was used to compare the experimental means.
Figure Preparation
Digital images were obtained using a CCD camera (MagnaFire SP; Optronics; Optical Analysis Corp., Nashua, NH) and LG-3 frame grabber attached to an Olympus microscope (Optical Analysis Corp.). Exposure times were held constant when acquiring images from animals processed and analyzed on the same day. Images were imported into Adobe Photoshop 7.0 (Adobe Systems Incorporated, San Jose, CA) where groups of images were assembled and labeled. To obtain images of CRF-IR structures in whole-mount preparations, whole-mounts were examined and optical sections were acquired using a Zeiss LSM 510 confocal scanning system (Carl Zeiss MicroImaging, Inc., Thornwood, NY) attached to a Zeiss LSM 510 microscope using a plan Fluor 40x oil objective and an excitation wavelength of 543 nm. Images from confocal scanning were assembled as described above.
Results
Corticotropin Releasing Factor (CRF) Immunoreactivity (IR) in Urothelium During Postnatal Development
The expression of CRF-IR was greatest in urothelium from P0–P7 urinary bladder sections. CRF-IR was present in all urothelial cell layers (Fig. 2, Fig. 3). An age-dependent decrease (Fig. 2, Fig. 4; R2= 0.978; p ≤ 0.001) in CRF expression was observed in the urothelium with adult urothelium exhibiting the least CRF expression. Depending on the degree of bladder distention, the urothelial cell layers may be more or less compact (Fig. 3). CRF-IR in individual urothelial cells is more easily observed from urinary bladders with less distention. However, even in urinary bladders with greater distention, CRF-IR in urothelial cells is apparent. CRF-IR is urothelial cells is also observed in wholemount preparations (see below). CRF-IR in detrusor smooth muscle was also observed but not quantified due to patchy expression in all sections examined at any age. Weak and patchy CRF-IR was present in the suburothelium during postnatal development and in adult urinary bladder and this expression did not appear to change with development (Fig. 2).
Figure 2.

CRF expression in urothelium in postnatal rat bladder. Fluorescence images of CRF expression in urinary bladder sections of P0 (A), P7 (B), P14 (C), P21 (D), P28 (E) and adult (F) rats. For all images, exposure times were held constant and all tissues were processed simultaneously. In adult and P28 rats, little CRF expression was visible in the urothelium (U; E, F). In younger rats (P0–P21; A–D) greater expression of CRF in the urothelium in all layers (apical, intermediate and basal) of the U was observed. In the youngest rats examined (P0–P7; A, B), some diffuse CRF expression was present in the suburothelium (SU) region and in some regions of smooth muscle (sm) but this staining was non-homogeneous. Calibration bar represents 50 µm.
Figure 3.

CRF-immunoreactivity (IR) in urothelial cells from urinary bladder with less (A–E) or more distention (F–H). A. CRF-IR is present in urothelial cells (arrows) throughout the thickness of the urothelium in a cryostat section of urinary bladder from postnatal day (P)14. B. Same section of urinary bladder stained with YOYO-1 to visualize cell nuclei. C. Merged image of A and B. The area enclosed by the rectangle (white lines) is magnified in panels D and E. D, E. Higher power magnification of urothelial cells exhibiting CRF-IR in the cytoplasm with visible nuclei (YOYO-1; arrows). F. CRF-IR in urothelial cells (arrows) from a urinary bladder with greater distention and flatter urothelial cells as a result of the distention. G. Same section of urinary bladder stained with YOYO-1 to visualize cell nuclei. H. Merged image of F and G with CRF-IR urothelial cells indicated (arrows). I. Absorption of CRF antiserum with CRF peptide (10 µg/ml) significantly reduced staining to background levels. U, urothelium; L, lumen. Calibration bar represents 80 µm in A–H, I 90 µm in D and 100 µm in E.
Figure 4.

Age-dependent reduction in urothelial CRF expression. Summary histogram of CRF expression in the urothelium during postnatal development expressed as a percentage of adult CRF expression. Histogram represents the average of n = 6 samples per postnatal age. An age-dependent reduction in urothelial CRF expression was evident (R2= 0.978; p ≤ 0.001). *, p ≤ 0.01 compared to P14–P36; **, p ≤ 0.01 compared to P14–P36; #, p ≤ 0.01 compared to P36.
Corticotropin Releasing Factor (CRF) Protein Expression in Urinary Bladder with Postnatal Development
CRF protein expression in whole urinary bladders as determined by ELISAs was greatest (p ≤ 0.01) in P14–P21 urinary bladder (Fig. 5). CRF expression in P14 urinary bladder was significantly (p ≤ 0.01) greater than that observed in P0–P4, P7, P28, P36 and adult bladder (Fig. 5). CRF expression in P21 urinary bladder was significantly (p ≤ 0.01) greater than that observed in P0-4, P28, P36 and adult bladder (Fig. 5). CRF protein expression in adult bladder was significantly (p ≤ 0.01) less than that observed in P0–P21 rat bladder (Fig. 5).
Figure 5.

Postnatal expression of corticotropin releasing factor (CRF) in urinary bladder. Changes in total urinary bladder CRF as detected with immunoassays during postnatal (P) development (P0-Adult). Significant increases in total urinary bladder CRF were detected at P0–P4 (##, p ≤ 0.01) compared to P36 and adult as well as at P7 (#, p ≤ 0.01) compared to P28-Adult. At P14, total urinary bladder CRF significantly (*, p ≤ 0.01) increased compared to P0–P7 and P28-Adult. At P21, total urinary bladder CRF significantly (**, p ≤ 0.01) increased compared to P0–P4 and P28-Adult. Histogram represents the average for n = 6 samples per group.
Corticotropin Releasing Factor (CRF) expression in whole-mounts of urothelium/suburothelium and detrusor smooth muscle
Our previous studies have demonstrated CRF-IR and PGP-positive nerve fibers in adult urinary bladder whole-mounts (LaBerge et al., 2006). Whole mount preparations of urinary bladder were prepared to aid in the visualization of the CRF-IR nerve plexus during postnatal development in the present study. As previously described (LaBerge et al., 2006), very few CRF-immunoreactive nerve fibers were observed in the detrusor whole mount preparation of postnatal and adult rats (not shown). In contrast, CRF-immunoreactive nerve fibers in the suburothelial plexus (Fig. 6) in the urothelium/suburothelium whole mount preparation were more abundant in trigone and neck regions but abundance varied with postnatal age. CRF-IR nerve fibers of the suburothelial plexus in the trigone and neck regions of the urinary bladder were most abundant in P14–P21 rat bladder (Fig. 6C, D). In P0–P7 urinary bladder wholemounts, CRF-IR is not observed in nerve fibers of the suburothelial plexus but CRF-IR is observed in urothelial cells (Fig. 6A, B)
Figure 6.

CRF-immunoreactive nerve fibers in suburothelial plexus of urinary bladder. Fluorescence photographs of CRF-immunoreactive nerve fibers in nerve fibers in the suburothelial plexus in whole mount preparations of the urothelium/suburothelium from postnatal rats (P0, A; P7, B; P14, C; P21, D; P36, E; Adult, F). CRF-immunoreactive nerve fibers in the suburothelial plexus in the urothelium/suburothelium whole mount preparation were abundant during the second-third postnatal weeks (C, D; arrows) compared to an absence of CRF expression in nerve fibers from P0–P7. Although CRF expression in nerve fibers was not evident at P0–P7, CRF expression in urothelial cells was present (A, B; arrows). With increasing age (P36-Adult; arrows) CRF expression in nerve fibers appeared to decrease in density. G. As a comparison, the suburothelial plexus was stained with the pan neuronal antibody, protein gene product 9.5, that labels all nerve fibers. All images (A–G) are from the bladder neck region that together with the trigone region exhibited the greatest density of CRF staining from P14-adult. Calibration bar represents 80 µm.
CRF receptor (CRFR1, CRFR2) transcript expression in urinary bladder during postnatal development
Our previous studies (LaBerge et al., 2006) demonstrated CRFR2 mRNA and protein expression in adult urinary bladder. In the present study from reverse transcription-PCR analyses, urinary bladder from postnatal rats also demonstrates CRFR2 transcript expression at all ages examined (Fig. 7). In contrast to adult urinary bladder expression of CRFR2 mRNA and protein (LaBerge et al., 2006), no CRFR1 mRNA or protein was demonstrated in adult rat urinary bladder. Similarly in the present study, postnatal rat urinary bladder did not express CRFR1 transcript in any postnatal age examined (Fig. 7). In contrast, L6 spinal cord does express CRFR1 transcript.
Figure 7.

CRF receptor (CRFR2) expression in postnatal urinary bladder. Urinary bladder from all postnatal ages examined express CRFR2 transcript (B). In contrast, CRFR1 is not detected in urinary bladder under the specified conditions despite the robust expression of CRFR1 in lumbar (L) 6 spinal cord (A).
Neuropeptide transcript expression in urinary bladder during postnatal development
A number of previous studies have demonstrated changes in neuropeptide expression in urinary bladder during postnatal development. In this study, we examined VIP, sub P, galanin and PACAP mRNA expression in postnatal (P7, P14) and adult urinary bladder. Consistent with previous studies, some neuropeptide transcripts exhibited a significant (p ≤ 0.05)(Fig. 8) reduction with increasing postnatal age.
Figure 8.
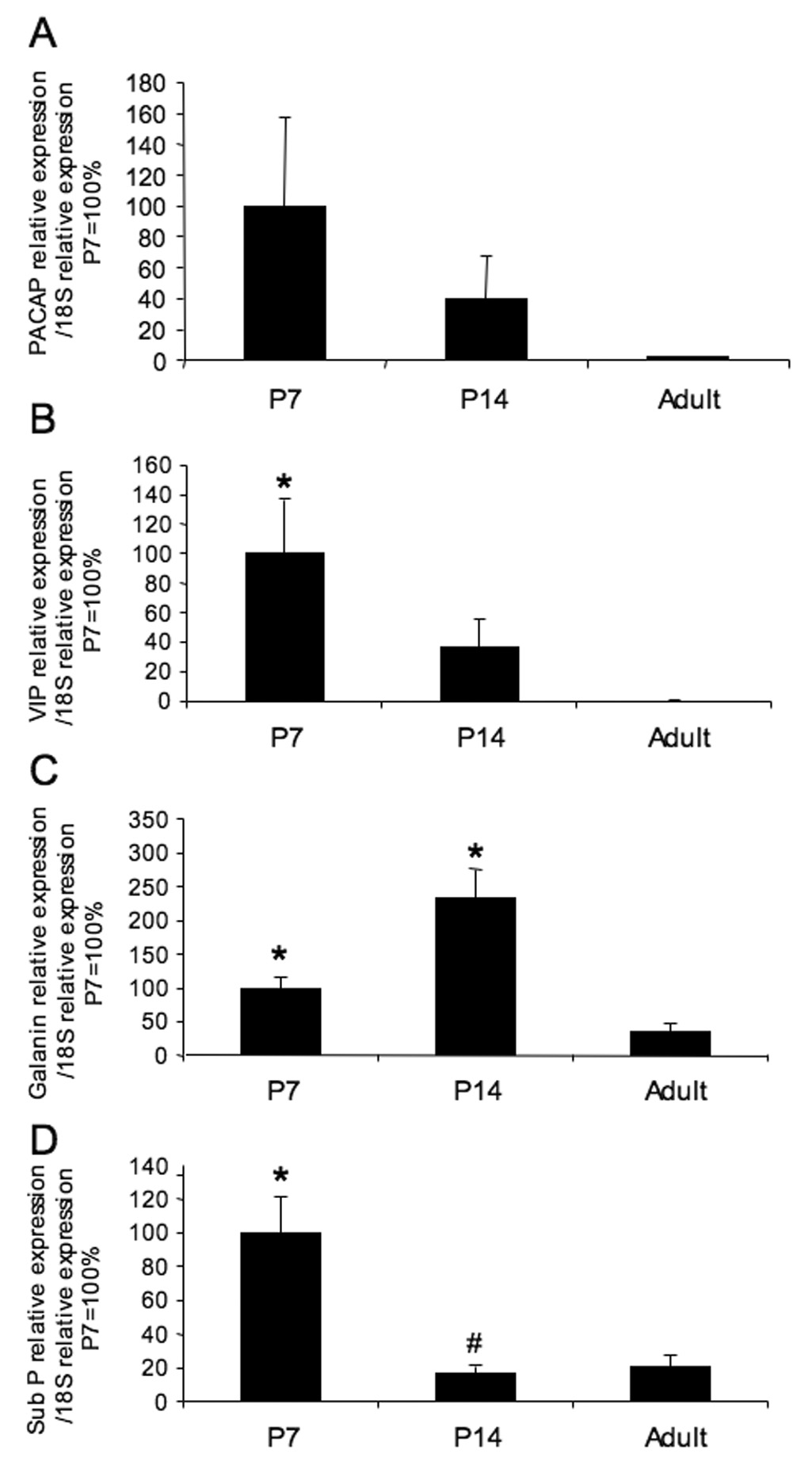
Postnatal plasticity in neuropeptide mRNA expression in urinary bladder. For each postnatal age, total RNA from individual urinary bladder was reverse transcribed, and the cDNA templates synthesized from urinary bladder were analyzed by real-time quantitative PCR using primers specific for rat pituitary adenylate cyclase activating polypeptide (PACAP), vasoactive intestinal polypeptide (VIP), galanin and substance P (sub P). Transcript levels are normalized to the housekeeping gene, 18S, and expressed as relative change compared to P7 (100%). For all neuropeptides examined, adult transcript expression in urinary bladder was less than that observed in younger urinary bladder. n = 4 for Adult; n = 8 each for P7, P14. *, p ≤ 0.01 versus adult; #, p ≤ 0.001 versus P7.
Discussion
We demonstrate several novel findings with respect to corticotropin releasing factor (CRF) expression in the urinary bladder during postnatal development. We have focused our analysis of postnatal CRF bladder expression on CRF expression in the urothelium and nerve fibers of the suburothelial plexus. Whereas age-dependent reduction in CRF expression was observed in postnatal urinary bladder, CRF expression in nerve fibers increased in density during the second-third postnatal weeks. Consistent with our previous studies of CRF receptor (CRFR) expression in the urinary bladder, urinary bladder from all postnatal ages examined expressed CRFR2. In contrast, CRFR1 expression was not observed. In addition, we demonstrate postnatal plasticity in mRNA expression of other neuropeptides including galanin, VIP, sub P and PACAP in urinary bladder.
Previous studies have demonstrated developmental plasticity in neuropeptide expression in the urinary bladder (Maggi et al., 1988; Iuchi et al., 1994; Sann et al., 1997; Mohammed et al., 2002; Zvarova and Vizzard, 2005a). We believe that this study is the first to demonstrate altered CRF in the urinary bladder and specifically in the urothelium and bladder nerve fibers during postnatal development. Although we were able to quantify an age-dependent reduction in CRF expression in urothelial cells, quantification of CRF-immunoreactive nerve fibers was confounded by the presence of the CRF-immunoreactive urothelial cells in the whole-mount preparations. Our qualitative assessments of CRF-immunoreactive nerve fibers in the suburothelial plexus revealed an increase in density at P14–P21 that corresponded to an increase in CRF expression in P14–P21 whole urinary bladder determined by ELISAs. Although the detrusor smooth muscle and suburothelial structures also expressed CRF-IR, expression was not homogenous and much less robust than that observed for the urothelium. The age-dependent reduction in CRF expression in the postnatal urothelium is in contrast to our previous demonstration of bladder inflammation induction of CRF in urothelium (LaBerge et al., 2006). Previous studies of sub P-immunoreactive nerve fibers in the urinary bladder also revealed a delayed expression until the second postnatal week (Maggi et al., 1988; Sann et al., 1997). The present study also demonstrated postnatal plasticity in mRNA expression in urinary bladder in the neuropeptides, galanin, sub P, PACAP and VIP. Future studies will be directed at determining the cellular distribution of these neuropeptides (galanin, sub P, PACAP, VIP) in the postnatal urinary bladder.
Urothelial cells share a number of similarities with sensory neurons and the urothelium has been suggested to have ‘neuronal-like’ properties (Birder et al., 2001; Birder, 2005). The present study demonstrates robust CRF-IR in the urothelium that decreases with increasing postnatal age and adds CRF to a growing list of neurotransmitters, neuropeptides and receptor systems that exist in both sensory neurons and urothelium (Birder, 2005; Birder, 2006; Hanna-Mitchell et al., 2007). The exact role of CRF expression in the urinary bladder is not known but with CRF expression in the urothelium and sensory nerves, release from both elements may be possible under certain conditions.
CRF is expressed in a bulbospinal pathway from Barrington’s nucleus to the sacral parasympathetic nucleus in the lumbosacral spinal cord (Merchenthaler et al., 1982; Merchenthaler, 1984; Vincent and Satoh, 1984; Suzuki et al., 1990). Historically, Barrington’s nucleus has been viewed as the supraspinal-switching center that regulates storage and elimination of urine (Satoh et al., 1978; Loewy et al., 1979; Noto et al., 1989). We have demonstrated an age-dependent upregulation of CRF-IR in the lumbosacral spinal cord in the region of preganglionic parasympathetic neurons (Studeny and Vizzard, 2005) as well as bladder inflammation induced upregulation of CRF in urinary bladder and spinal cord (LaBerge et al., 2006). We suggested that upregulation of CRF-IR in bulbospinal projections to the SPN may contribute to mature voiding reflexes (Studeny and Vizzard, 2005) and may act as a local, proinflammatory agent (Kawahito et al., 1995; LaBerge et al., 2006). Although identified as a neurotransmitter in micturition reflex pathways, opposing functional roles for CRF in mature bladder function have been described (Pavcovich et al., 1998; Klausner and Steers, 2004; Klausner et al., 2005). Intrathecal administration of CRF decreases the amplitude of bladder contractions induced by electrical stimulation of Barrington’s nucleus (Pavcovich et al., 1998), suggesting that CRF in descending projections from Barrington’s nucleus to the SPN is inhibitory. Similarly, intrathecal or systemic administration of the CRFR1 antagonists, antalarmin and NBI-30775, increase urinary frequency and decrease bladder capacity in conscious rats (Kiddoo et al., 2006). However, other studies demonstrate that intrathecal or systemic CRF induces bladder overactivity (Klausner and Steers, 2004; Klausner et al., 2005). Different putative roles for CRF in these studies may be related to the use of anesthesia (Pavcovich et al., 1998) or its absence (Klausner and Steers, 2004; Klausner et al., 2005) for functional studies.
The mechanism(s) underlying the changes in CRF expression in the neonatal rat bladder in the present study and the development of mature voiding reflexes is not known. Previous studies have suggested that maturation of peptidergic afferent pathways (Maggi et al., 1988; Iuchi et al., 1994; Sann et al., 1997; Mohammed et al., 2002; Zvarova and Vizzard, 2005a) and purinergic receptor systems (Studeny et al., 2005b) in the urinary bladder of the neonatal rat may be involved in these processes. It has also been suggested therefore that a certain afferent innervation density of the urinary bladder is a prerequisite for the development of the mature voiding reflex (Sann et al., 1997) and our demonstration of an increase in CRF innervation density of the urinary bladder at the second-third postnatal week supports this suggestion. The reduction in CRF expression in nerve fibers after this time (2nd-3rd postnatal week) was similarly observed for another peptide (cocaine-and-amphetamine regulated transcript peptide, CARTp) in the postnatal bladder (Zvarova and Vizzard, 2005a). In addition, we have demonstrated significant changes in urinary bladder mRNA for other neuropeptides including galanin, sub P, PACAP and VIP during early postnatal development consistent with a number of studies demonstrating neuropeptide protein content and immunoreactivity changes during early postnatal development (Maggi et al., 1988; Iuchi et al., 1994; Sann et al., 1997; Mohammed et al., 2002; Zvarova and Vizzard, 2005a).
This study also extends our findings with respect to CRF receptor (CRFR) expression in the urinary bladder. We have previously demonstrated CRFR2 mRNA and protein expression in the adult urinary bladder and increased transcript expression with bladder inflammation (LaBerge et al., 2006). In contrast, no CRFR1 transcript or protein expression was evident in adult urinary bladder (LaBerge et al., 2006). In the present study, we examined CRFR2 and CRFR1 transcript expression in postnatal urinary bladder to determine if CRFR1 transcript expression, in particular, was present during early postnatal development. Consistent with our findings in adult urinary bladder, no CRFR1 transcript expression was amplified from any postnatal urinary bladder. In contrast, CRFR2 transcript expression was present from the earliest postnatal age examined. It is important to note that CRF may not be the natural ligand for CRFR2. Numerous studies suggest that the natural ligand(s) for CRFR2 are the urocortins (urocortin 2 and 3) (Kageyama et al., 2000; Gravanis and Margioris, 2005; Nijsen et al., 2005). Bimodal effects on a number of physiological responses including visceral nociception (Nijsen et al., 2005; Million et al., 2006), gastrointestinal motility (Porcher et al., 2005; Million et al., 2007) and stress (Valdez et al., 2002; Nijsen et al., 2005; Million et al., 2007) have been demonstrated for CRFR1 and CRFR2 activation. Whereas CRF may not be the natural ligand for CRFR2 (Kageyama et al., 2000), recent studies suggest that CART protein may function through CRF receptors. Recent studies (Zvarova and Vizzard, 2005a) have demonstrated CART expression in intramural neurons and nerve fibers of the urinary bladder during development and in adult rats. Furthermore, CART peptide also facilitates detrusor contractility (Zvarova and Vizzard, 2005b). Whether CART functions through CRF receptors in the urinary bladder remains to be determined.
In conclusion, these studies have demonstrated significant changes in CRF expression in the urinary bladder during postnatal development. An age-dependent reduction in CRF expression in the urothelium was demonstrated whereas increases in CRF-immunoreactive nerve fibers increased in density during the second-third postnatal week. This increase in CRF-immunoreactive nerve fiber staining corresponded to an increase in total CRF expression in the urinary bladder demonstrated with ELISAs during the second-third postnatal weeks. CRFR2 transcript expression was present in urinary bladder of all ages examined in contrast to the absence of CRFR1 transcript expression. Current studies are directed at determining the contribution of neuropeptide/receptor systems to the maturation of bladder reflexes.
Acknowledgments
J. LaBerge was supported, in part, by a Neuroscience Summer Undergraduate Research Fellowship from the University of Vermont, Departments of Anatomy and Neurobiology and Neurology. Gratitude is expressed to the UVM Microscopy Imaging Center for expert guidance and support with semi-quantitative analysis of CRF expression.
Grants
This work was funded by NIH grants DK051369, DK060481, DK065989.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Benarroch EE, Schmeiche IAM. Depletion of corticotrophin-releasing factor neurons in the pontine micturition area in multiple system atrophy. Annals Neurol. 2001;50(5):640–645. doi: 10.1002/ana.1258. [DOI] [PubMed] [Google Scholar]
- Birder LA. More than just a barrier: urothelium as a drug target for urinary bladder pain. Am J Physiol Renal Physiol. 2005;289(3):F489–F495. doi: 10.1152/ajprenal.00467.2004. [DOI] [PubMed] [Google Scholar]
- Birder LA. Urinary bladder urothelium: molecular sensors of chemical/thermal/mechanical stimuli. Vascul Pharmacol. 2006;45(4):221–226. doi: 10.1016/j.vph.2005.08.027. [DOI] [PubMed] [Google Scholar]
- Birder LA, Kanai AJ, deGroat WC, Kiss S, Nealen ML, Burke NE, Dineley KE, Watkins S, Reynolds IJ, Caterina MJ. Vanilloid receptor expression suggests a sensory role for urinary bladder epithelial cells. Proc Natl Acad Sci (USA) 2001;98(23):13396–13401. doi: 10.1073/pnas.231243698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braas KM, May V, Zvara P, Nausch B, Kliment J, Dunleavy JD, Nelson MT, Vizzard MA. Role for pituitary adenylate cyclase activating polypeptide in cystitis-induced plasticity of micturition reflexes. Am J Physiol Regul Integr Comp Physiol. 2006;290(4):R951–R962. doi: 10.1152/ajpregu.00734.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bregman BS. Development of serotonin immunoreactivity in the rat spinal cord and its plasticity after neonatal spinal cord lesions. Dev Brain Res. 1987;34:245–263. doi: 10.1016/0165-3806(87)90213-6. [DOI] [PubMed] [Google Scholar]
- Corrow KA, Vizzard MA. Phosphorylation of extracellular signal-regulated kinases in urinary bladder in rats with cyclophosphamide-induced cystitis. Am J Physiol Regul Integr Comp Physiol. 2007;293(1):R125–R134. doi: 10.1152/ajpregu.00857.2006. [DOI] [PubMed] [Google Scholar]
- de Groat WC, Araki I. Maturation of bladder reflex pathways during postnatal development. Adv Exp Med Biol. 1999;462:253–263. doi: 10.1007/978-1-4615-4737-2_19. discussion 311–220. [DOI] [PubMed] [Google Scholar]
- Ekstrom J, Ekman R, Hakanson R. Ontogeny of neuropeptides in the rat urinary bladder. Regul Pept. 1994;50(1):23–28. doi: 10.1016/0167-0115(94)90187-2. [DOI] [PubMed] [Google Scholar]
- Fitzgerald M. Development of pain mechanisms. Br Med Bull. 1991;47:667–675. doi: 10.1093/oxfordjournals.bmb.a072499. [DOI] [PubMed] [Google Scholar]
- Girard BA, Lelievre V, Braas KM, Razinia T, Vizzard MA, Ioffe Y, El Meskini R, Ronnett GV, Waschek JA, May V. Noncompensation in peptide/receptor gene expression and distinct behavioral phenotypes in VIP- and PACAP-deficient mice. J Neurochem. 2006;99(2):499–513. doi: 10.1111/j.1471-4159.2006.04112.x. [DOI] [PubMed] [Google Scholar]
- Girard BM, May V, Bora SH, Fina F, Braas KM. Regulation of neurotrophic peptide expression in sympathetic neurons: quantitative analysis using radioimmunoassay and real-time quantitative polymerase chain reaction. Regul Pept. 2002;109(1–3):89–101. doi: 10.1016/s0167-0115(02)00191-x. [DOI] [PubMed] [Google Scholar]
- Gravanis A, Margioris AN. The corticotropin-releasing factor (CRF) family of neuropeptides in inflammation: potential therapeutic applications. Curr Med Chem. 2005;12(13):1503–1512. doi: 10.2174/0929867054039008. [DOI] [PubMed] [Google Scholar]
- Hanna-Mitchell AT, Beckel JM, Barbadora S, Kanai AJ, de Groat WC, Birder LA. Non-neuronal acetylcholine and urinary bladder urothelium. Life Sci. 2007;80(24–25):2298–2302. doi: 10.1016/j.lfs.2007.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iuchi H, Satoh Y, Ono K. Postnatal development of neuropeptide Y-and calcitonin gene-related peptide-immunoreactive nerves in the rat urinary bladder. Anat Embryol (Berl) 1994;189(4):361–373. doi: 10.1007/BF00190591. [DOI] [PubMed] [Google Scholar]
- Johnson JM, Skau KA, Gerald MC, Wallace LJ. Regional noradrenergic and cholinergic neurochemistry in the rat urinary bladder: effect of age. J Urol. 1988;139:611–615. doi: 10.1016/s0022-5347(17)42543-2. [DOI] [PubMed] [Google Scholar]
- Kageyama K, Gaudriault GE, Bradbury MJ, Vale WW. Regulation of corticotropin-releasing factor receptor type 2 beta messenger ribonucleic acid in the rat cardiovascular system by urocortin, glucocorticoids, and cytokines. Endocrinology. 2000;141(7):2285–2293. doi: 10.1210/endo.141.7.7572. [DOI] [PubMed] [Google Scholar]
- Kawahito Y, Sano H, Mukai S, Asai K, Kimura S, Yamamura Y, Kato H, Chrousos GP, Wilder RL, Kondo M. Corticotropin releasing hormone in colonic mucosa in patients with ulcerative colitis. Gut. 1995;37(4):544–551. doi: 10.1136/gut.37.4.544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiddoo DA, Valentino RJ, Zderic S, Ganesh A, Leiser SC, Hale L, Grigoriadis DE. Impact of state of arousal and stress neuropeptides on urodynamic function in freely moving rats. Am J Physiol Regul Integr Comp Physiol. 2006;290(6):R1697–R1706. doi: 10.1152/ajpregu.00742.2005. [DOI] [PubMed] [Google Scholar]
- Klausner AP, Steers WD. Corticotropin releasing factor: a mediator of emotional influences on bladder function. J Urol. 2004;172(6 Pt 2):2570–2573. doi: 10.1097/01.ju.0000144142.26242.f3. [DOI] [PubMed] [Google Scholar]
- Klausner AP, Streng T, Na YG, Raju J, Batts TW, Tuttle JB, Andersson KE, Steers WD. The role of corticotropin releasing factor and its antagonist, astressin, on micturition in the rat. Auton Neurosci. 2005;123(1–2):26–35. doi: 10.1016/j.autneu.2005.08.003. [DOI] [PubMed] [Google Scholar]
- Kruse MN, Tanowitz M, Cheng C, de Groat WC. Postnatal development of the sensory innervation of the urinary bladder in the rat. Soc Neurosci Abstr. 1990;16:1064. [Google Scholar]
- LaBerge J, Malley SE, Zvarova K, Vizzard MA. Expression of corticotropin-releasing factor and CRF receptors in micturition pathways after cyclophosphamide-induced cystitis. Am J Physiol Regul Integr Comp Physiol. 2006;291(3):R692–R703. doi: 10.1152/ajpregu.00086.2006. [DOI] [PubMed] [Google Scholar]
- Loewy AD, Saper CB, Baker RP. Descending projections from the pontine micturition center. Brain Res. 1979;172:533–538. doi: 10.1016/0006-8993(79)90584-5. [DOI] [PubMed] [Google Scholar]
- Maggi CA, Santicioli P, Geppeti S, Frilli MG, Spillantini MG, Nediani C, Hunt SP, Meli A. Biochemical, anatomical and functional correlates of postnatal developments of the capsaicin-sensitive innervation of the rat urinary bladder. Dev Brain Res. 1988;43:183–190. doi: 10.1016/0165-3806(88)90098-3. [DOI] [PubMed] [Google Scholar]
- Marti E, Gibson SJ, Polak JM, Facer P, Springall DR, Van Aswegen G, Aithison M, Koltzenburg M. Ontogeny of peptide and amine containing neurones in motor, sensory, and autonomic regions of rat and human spinal cord, dorsal root ganglia and rat skin. J Comp Neurol. 1987;266:332–359. doi: 10.1002/cne.902660304. [DOI] [PubMed] [Google Scholar]
- Merchenthaler I. Corticotropin releasing factor (CRF)-like immunoreactivity in the rat central nervous system. Extrahypothalamic distribution. Peptides. 1984;5:53–69. doi: 10.1016/0196-9781(84)90265-1. [DOI] [PubMed] [Google Scholar]
- Merchenthaler I, Vigh S, Petrusz P, Schally AV. Immunocytochemical localization of corticotropin releasing factor (CRF) in the rat brain. Am J Anat. 1982;165:385–896. doi: 10.1002/aja.1001650404. [DOI] [PubMed] [Google Scholar]
- Million M, Wang L, Stenzel-Poore MP, Coste SC, Yuan PQ, Lamy C, Rivier J, Buffington T, Tache Y. Enhanced pelvic responses to stressors in female CRF-overexpressing mice. Am J Physiol Regul Integr Comp Physiol. 2007;292(4):R1429–R1438. doi: 10.1152/ajpregu.00626.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Million M, Wang L, Wang Y, Adelson DW, Yuan PQ, Maillot C, Coutinho SV, McRoberts JA, Bayati A, Mattsson H, Wu V, Wei JY, Rivier J, Vale W, Mayer EA, Tache Y. CRF2 receptor activation prevents colorectal distension induced visceral pain and spinal ERK1/2 phosphorylation in rats. Gut. 2006;55(2):172–181. doi: 10.1136/gut.2004.051391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mineta K, Nomura M, Terado M, Fujimoto N, Sasaguri T, Ueta Y, Matsumoto T. Upregulation of corticotropin-releasing hormone gene expression in the paraventricular nucleus of cyclophosphamide-induced cystitis in male rats. Brain Res. 2004;1018(2):193–200. doi: 10.1016/j.brainres.2004.05.063. [DOI] [PubMed] [Google Scholar]
- Mohammed H, Hannibal J, Fahrenkrug J, Santer R. Distribution and regional variation of pituitary adenylate cyclase activating polypeptide and other neuropeptides in the rat urinary bladder and ureter: effects of age. Urol Res. 2002;30(4):248–255. doi: 10.1007/s00240-002-0261-6. [DOI] [PubMed] [Google Scholar]
- Nijsen M, Ongenae N, Meulemans A, Coulie B. Divergent role for CRF1 and CRF2 receptors in the modulation of visceral pain. Neurogastroenterol Motil. 2005;17(3):423–432. doi: 10.1111/j.1365-2982.2005.00644.x. [DOI] [PubMed] [Google Scholar]
- Noto H, Roppolo JR, Steers WD, de Groat WC. Electrophysiological analysis of the ascending and descending components of the micturition reflex pathway in the rat. Brain Res. 1989;549:95–105. doi: 10.1016/0006-8993(91)90604-t. [DOI] [PubMed] [Google Scholar]
- Pavcovich LA, Valentino RJ. Central regulation of micturition in the rat by corticotropin-releasing hormone from Barrington's nucleus. Neurosci Lett. 1995;196:185–188. doi: 10.1016/0304-3940(95)11873-u. [DOI] [PubMed] [Google Scholar]
- Pavcovich LA, Yang M, Miselis RR, Valentino RJ. Novel role for the pontine micturition center, Barrington's nucleus: evidence for coordination of colonic and forebrain activity. Brain Res. 1998;784(1–2):355–361. doi: 10.1016/s0006-8993(97)01178-5. [DOI] [PubMed] [Google Scholar]
- Porcher C, Juhem A, Peinnequin A, Sinniger V, Bonaz B. Expression and effects of metabotropic CRF1 and CRF2 receptors in rat small intestine. Am J Physiol Gastrointest Liver Physiol. 2005;288(5):G1091–G1103. doi: 10.1152/ajpgi.00302.2004. [DOI] [PubMed] [Google Scholar]
- Puder BA, Papka RE. Distribution and origin of corticotropin-releasing factor-immunoreactive axons in the female rat lumbosacral spinal cord. J Neurosci Res. 2001;66(6):1217–1225. doi: 10.1002/jnr.10033. [DOI] [PubMed] [Google Scholar]
- Saban MR, Nguyen NB, Hammond TG, Saban R. Gene expression profiling of mouse bladder inflammatory responses to LPS, substance P, and antigen-stimulation. Am J Path. 2002;160(6):2095–2110. doi: 10.1016/S0002-9440(10)61159-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sann H, Walb G, Pierau FK. Postnatal development of the autonomic and sensory innervation of the musculature in the rat urinary bladder. Neurosci Lett. 1997;236(1):29–32. doi: 10.1016/s0304-3940(97)00752-0. [DOI] [PubMed] [Google Scholar]
- Satoh K, Shimizu N, Tohyama M, Maeda T. Localization of the micturition reflex center at dorsolateral pontine tegmentum of the rat. Neurosci Lett. 1978;8:27–33. doi: 10.1016/0304-3940(78)90092-7. [DOI] [PubMed] [Google Scholar]
- Studeny S, Torabi A, Vizzard MA. P2X2 and P2X3 Receptor Expression in Postnatal Rat Urinary Bladder and Lumbosacral Spinal Cord: A Developmental Study. Am J Physiol Regul Integr Comp Physiol. 2005;289(4):R1155–R1168. doi: 10.1152/ajpregu.00234.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Studeny S, Vizzard MA. Corticotropin-releasing factor (CRF) expression in postnatal and adult rat sacral parasympathetic nucleus (SPN) Cell Tissue Res. 2005;322(3):339–352. doi: 10.1007/s00441-005-0014-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugaya K, de Groat WC. Micturition reflexes in the in vitro neonatal rat brainstem-spinal cord-bladder preparation. Am J Physiol Regul Integr Comp Physiol. 1994;266(35):R658–R667. doi: 10.1152/ajpregu.1994.266.3.R658. [DOI] [PubMed] [Google Scholar]
- Sugaya K, deGroat WC. Influence of temperature on activity of the isolated whole bladder preparation of neonatal and adult rats. Am J Physiol Regul Integr Comp Physiol. 2000;278(1):R238–R246. doi: 10.1152/ajpregu.2000.278.1.R238. [DOI] [PubMed] [Google Scholar]
- Suzuki T, Kawatani M, Erdman S, de Groat WC. Role of CRF and 5-HT in central pathways controlling micturition in the rat. Soc Neurosci Abstr. 1990;16:1064. [Google Scholar]
- Szell EA, Somogyi GT, de Groat WC, Szigeti GP. Developmental changes in spontaneous smooth muscle activity in the neonatal rat urinary bladder. Am J Physiol Regul Integr Comp Physiol. 2003;285(4):R809–R816. doi: 10.1152/ajpregu.00641.2002. [DOI] [PubMed] [Google Scholar]
- Valdez GR, Inoue K, Koob GF, Rivier J, Vale W, Zorrilla EP. Human urocortin II: mild locomotor suppressive and delayed anxiolytic-like effects of a novel corticotropin-releasing factor related peptide. Brain Res. 2002;943(1):142–150. doi: 10.1016/s0006-8993(02)02707-5. [DOI] [PubMed] [Google Scholar]
- Valentino RJ, Chen S, Zhu Y, Aston-Jones G. Evidence for divergent projections to the brain noradrenergic system and the spinal parasympahetic system from Barrington's nucleus. Brain Res. 1996;732(1–2):1–15. doi: 10.1016/0006-8993(96)00482-9. [DOI] [PubMed] [Google Scholar]
- Valentino RJ, Pavcovich LA, Hirata H. Evidence for corticotrophin-releasing hormone projections from Barrington's nucleus to the periaqueductal gray and dorsal motor nucleus of the vagus in the rat. J Comp Neurol. 1995;363(3):402–422. doi: 10.1002/cne.903630306. [DOI] [PubMed] [Google Scholar]
- Vincent SR, Satoh K. Corticotropin-releasing factor (CRF) immunoreactivity in the dorsolateral pontine tegmentum: further studies on the micturition reflex system. Brain Res. 1984;308:387–391. doi: 10.1016/0006-8993(84)91085-0. [DOI] [PubMed] [Google Scholar]
- Vizzard MA. Increased expression of neuronal nitric oxide synthase in bladder afferent and spinal neurons following spinal cord injury. Dev Neurosci. 1997;19(3):232–246. doi: 10.1159/000111212. [DOI] [PubMed] [Google Scholar]
- Vizzard MA, Erdman SL, Forstermann U, de Groat WC. Ontogeny of nitric oxide synthase in the lumbosacral spinal cord of the neonatal rat. Dev Brain Res. 1994;81:201–217. doi: 10.1016/0165-3806(94)90307-7. [DOI] [PubMed] [Google Scholar]
- Vizzard MA, Wu KH, Jewett IT. Developmental expression of urinary bladder neurotrophic factor mRNA and protein in the neonatal rat. Brain Res Dev Brain Res. 2000;119(2):217–224. doi: 10.1016/s0165-3806(99)00174-1. [DOI] [PubMed] [Google Scholar]
- Wu SV, Yuan PQ, Wang L, Peng YL, Chen CY, Tache Y. Identification and characterization of multiple corticotropin-releasing factor type 2 receptor isoforms in the rat esophagus. Endocrinology. 2007;148(4):1675–1687. doi: 10.1210/en.2006-0565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuridullah R, Corrow KA, Malley SE, Vizzard MA. Expression of fractalkine and fractalkine receptor in urinary bladder after cyclophosphamide (CYP)-induced cystitis. Auton Neurosci. 2006:126–127. 380–389. doi: 10.1016/j.autneu.2006.02.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zvarova K, Vizzard MA. Distribution and fate of cocaine- and amphetamine-regulated transcript peptide (CARTp)-expressing cells in rat urinary bladder: a developmental study. J Comp Neurol. 2005a;489(4):501–517. doi: 10.1002/cne.20657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zvarova K, Vizzard MA. Ontogeny of cocaine-and amphetamine-regulated transcript peptide (CARTp) in urinary bladder and lumbosacral spinal cord of neonatal rat. Urothelial Cell Physiology in Normal and Disease States. 2005b:58. [Google Scholar]