

Abstract
We previously reported large-scale reorganization of visual processing (i.e., activation of “foveal” cortex by peripheral stimuli) in two individuals with loss of foveal input from macular degeneration (Baker, Peli, Knouf & Kanwisher, 2005). Here, we replicate this result in three new individuals. Further we test the hypothesis that this reorganization is dependent on complete loss of foveal input. In two other individuals with extensive retinal lesions but some foveal sparing we found no evidence for reorganization. We conclude that large-scale reorganization of visual processing in MD occurs only in the complete absence of functional foveal vision.
Keywords: V1, cortical reorganization, fMRI, cortical plasticity, deprivation
1. Introduction
We previously reported large-scale reorganization of visual processing in two people with loss of foveal vision due to macular degeneration (MD): stimuli presented in the periphery activated regions of retinotopic cortex that would normally only be responsive to central visual stimuli (Baker et al., 2005). Although another recent study reported reorganization following loss of visual input as a result of optic radiation damage (Dilks, Serences, Rosenau, Yantis & McCloskey, 2007), and reorganization has been reported in individuals with congenital visual deficits (Baseler, Brewer, Sharpe, Morland, Jagle & Wandell, 2002), other groups have failed to find convincing and consistent evidence for reorganization in MD (Cheung, Schuchard, He, Tai, Legge & Hu, 2005, Masuda, Dumoulin, Nakadomari & Wandell, 2008, Masuda, Nakadomari, Asakawa, Dumoulin, Liu, Cheung, Ben-Schachar, Kitahara & Wandell, 2006, Sunness, Liu & Yantis, 2004). In reviewing the other studies of individuals with MD, we noted that in some the retinal lesion did not completely encompass the fovea (participant JMD4 in Masuda et al., 2008, Sunness et al., 2004), and in others the participants were older, with later onset of the disease than in our study (Cheung et al., 2005). We hypothesized that activation of foveal retinotopic cortex by peripheral stimuli might arise only in people with complete bilateral loss of foveal vision. Here, we scanned five new individuals with MD to determine whether our original findings could be reproduced, and to further establish what factors (e.g., presence of foveal sparing, age of onset of MD, type of MD) determine whether such reorganization will occur.
Extensive topographic reorganization of primary visual cortex (V1) has also been reported in adult cats and monkeys following discrete retinal lesions (Chino, Kaas, Smith, Langston & Cheng, 1992, Chino, Smith, Kaas, Sasaki & Cheng, 1995, Gilbert & Wiesel, 1992, Heinen & Skavenski, 1991, Kaas, Krubitzer, Chino, Langston, Polley & Blair, 1990). Neurons in the deprived region of V1 (i.e., the region that previously responded only to stimuli falling on the subsequently lesioned part of the retina) became responsive to stimuli falling on parts of the retina adjacent to the lesioned area. However, one prominent result failed to find reorganization in adult primates following retinal lesions (Smirnakis, Brewer, Schmid, Tolias, Schuz, Augath, Inhoffen, Wandell & Logothetis, 2005), spawning considerable controversy (Calford, Chino, Das, Eysel, Gilbert, Heinen, Kaas & Ullman, 2005, Giannikopoulos & Eysel, 2006, Horton & Hocking, 1998, Smirnakis, Schmid, Brewer, Tolias, Schuz, Augath, Inhoffen, Wandell & Logothetis, 2005). Thus, in both adult humans and animals a consensus no longer exists on the existence or degree of adult cortical reorganization following visual deprivation.
Here, we used fMRI to test for reorganization of visual processing in five individuals with extensive retinal damage from MD with different etiologies and ages of onset. Further, to test whether reorganization depends upon complete loss of foveal input or whether instead the cortex receiving foveal input is recruited by peripheral stimuli after any extensive retinal damage, we compared individuals with and without foveal sparing. Consistent with our earlier report, we find clear activation of the cortical region that typically receives only foveal input (i.e. the “foveal confluence” at the occipital pole) (Dougherty, Koch, Brewer, Fischer, Modersitzki & Wandell, 2003) by peripheral visual stimuli in all three individuals with complete bilateral loss of foveal function. However, we found no evidence for such reorganization in the two individuals with some spared foveal vision. Thus, we conclude that large-scale reorganization of visual processing (i.e., activation of the foveal confluence by peripheral visual stimuli) is only observed when functional input to foveal cortex is completely lost.
2. Methods
2.1 Participants
We tested five individuals with macular degeneration (Table 1) with large central bilateral scotomata. To distinguish these individuals from those we reported previously, termed MD1 and MD2 (Baker et al., 2005), we refer to these individuals as MD3-MD7. For each MD individual we also tested a control participant (with full field vision) with visual stimuli presented to the same retinal locations as for their matched MD participant. MD6 and MD7 had similar scotomata and a “Preferred Retinal Locus” or PRL (new retinal locus for fixation) in very similar retinal positions, and so only one matched control was run for these 2 participants.
Table 1.
Participant details for all seven individuals with macular degeneration we have tested including two (MD1 and MD2) whose data we reported previously (Baker et al., 2005). MDs 1-5 all had no measurable foveal function whereas MDs 6-7 both had some degree of spared foveal function. JMD is Juvenile Macular Degeneration. AMD is Age-related Macular Degeneration. An asterisk denotes the tested eye (no asterisk indicates that the participant was tested binocularly).
| Participant | Gender | Age of onset (years) |
Time since onset (years) |
Visual Acuity | Foveal sparing? |
Diagnosis | Fixation Stability | Stimulus Size (degrees: w×h) |
Evidence for reorganization |
||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OD | OS | ||||||||||||
| OD | OS | 2° (%) |
4° (%) |
2° (%) |
4° (%) |
||||||||
| MD1 | M | 37 | 19 | 20/330 | 20/330 | N | Cone-rod dystrophy |
88 | 99 | 50 | 95 | 12×5 | Yes |
| MD2 | M | 11 | 39 | 20/600 | 20/350* | N | JMD | 5 | 31 | 21 | 63 | 8×10 | Yes |
| MD3 | M | 9 | 19 | 20/80 | 20/160* | N | JMD | 58 | 94 | 25 | 60 | 3×3 | Yes |
| MD4 | M | 9 | 46 | 20/300 | 20/400 | N | JMD with peripheral cone involveme nt |
58 | 89 | 71 | 100 | 10×4 | Yes |
| MD5 | F | 58 | 19 | 20/80 | 20/80 | N | Atrophic AMD |
68 | 97 | 58 | 99 | 3×4 | Yes |
| MD6 | F | 36 | 21 | 20/350 central |
20/80* central |
Y | JMD | 28 | 67 | 23 | 79 | 20×7 | No |
| MD7 | F | 46 | 18 | 20/350 central |
20/30* central |
Y | JMD | 21 | 61 | 7 | 26 | 20×7 | No |
All MD participants were carefully tested behaviorally to determine 1) visual field loss, including testing for any residual macular function, 2) location of the subject's PRL (Timberlake, Mainster, Peli, Augliere, Essock & Arend, 1986), and 3) fixation stability. Participants were only selected for this study if they had a single, stable PRL and they could maintain fixation using this PRL.
We recruited two groups of participants. In the first group (MD3-MD5), participants had large bilateral central scotomata with complete loss of foveal function as measured behaviorally. In the second group (MD6-MD7), participants had large bilateral central scotomata, but with some preserved foveal function due to sparing at the center of their lesions. These participants could fixate either with PRL or with fovea.
The precise experimental details were tailored to each individual MD participant depending on the shape and size of the scotoma and location of the PRL. Below we will first describe experimental procedures common to all participants before describing the specific details for each MD participant.
2.2 Stimuli
Stimuli were gray-scale photographs of objects (e.g., airplane, chair, watch) or words presented at either the fovea (or the location of the former fovea) or at a peripheral retinal location (i.e., the MD participant's PRL, or the control's matched peripheral location). For a given MD participant, the images were the same size for all locations and were scaled for each MD participant according to 1) the size of the scotoma (so that the images would be entirely within the scotoma when presented at the fovea) and 2) the distance of the PRL from the former fovea (so that the objects presented at PRL were large enough to be discriminated by the participant). The appropriate size of the stimuli was determined behaviorally before scanning by adjusting the size of the stimuli at the PRL location until the participant could reliably discriminate between individual objects or letters. We further adjusted the aspect ratio of the stimuli, depending on the location of each MD participant's PRL and the shape of the scotoma, to maximize the amount of visual stimulation while ensuring that the stimuli would still fit within the scotoma. For example, in MD4, the PRL was located above the scotoma, which extended much more horizontally than vertically, therefore we presented stimuli that were also elongated horizontally. For the specific stimulus sizes used for each participant, see section 2.4 below and Table 1.
2.3 Behavioral testing/Vision testing
Retinal imaging and perimetry
A Nidek MP-1 retinal microperimeter (Nidek Technologies, Vigonza, Italy) was used to map the locations of the PRL and fovea, and to measure the stability of fixation at the PRL for all MD participants. Stability of fixation was also measured at the fovea for those MD participants with some spared foveal vision. The retinal image tracker of the MP-1 recorded the subject's eye movement during 30s fixation trials (fixation task), and during longer static perimetry tasks (Figure 1 and Supplementary Figure 1).
Figure 1.
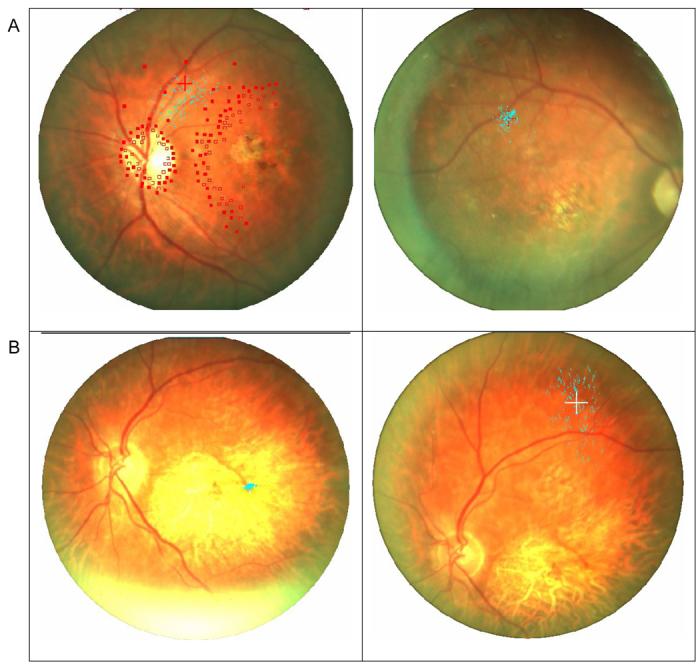
Samples of fixation stability measured with the Nidek MP-1. The blue dots represent 900 samples of the position of the fixation cross on the retina. A (Left). MD3's PRL fixation stability taken during an extended session of static perimetry (open squares represent invisible targets and filled square visible ones). Even during this extended and attention requiring task, fixation was stable (45% of the samples within 4 degrees), and the fixation cross was never closer than 10 degrees from the fovea. A (Right). MD3's PRL fixation stability during the fixation task (shown with the right eye to illustrate the correspondence of the PRLs between the eyes) was more stable (94% of the samples within 4 degrees). B (Left). MD7's central fixation stability with the residual left fovea demonstrating normal foveal fixation stability where 100% of the samples were within 4 degrees. B (Right). MD7's PRL fixation stability for the same subject was much less stable (26% of the samples within 4 degrees), yet it is obvious that the fixation cross was never closer than 10 degrees from the fovea.
Foveal location was determined using the Nidek fixation image and data file. An experimenter manually marked with a cursor a series of points along the optic disk margin. A custom Matlab program was used to fit an ellipse to the set of points (Fitzgibbon, Pilu & Fisher, 1999) and find the center of the optic disc. The position of the fovea was computed to be at 15.3° temporally and 1.5° below the center of the optic disk. The latter values were averages of the values determined for normally sighted observers (Rohrschneider, Springer, Bultmann & Volcker, 2005, Schuchard & Fletcher, 1994, Timberlake, Sharma, Grose, Gobert, Gauch & Maino, 2005) (Supplementary Figure 2).
The fixation task provided a cluster of 750 samples of the location of the fixation cross on the retina (25 samples per second over the 30 seconds of fixation). The same fixation records collected during the perimetry provide an estimate of the fixation stability during longer tasks that require attention similar to that needed during the fMRI recording. This procedure is more extensive and accurate than the scanning laser ophthalmoscope (SLO) PRL evaluation of fixation stability conducted manually in our previous study (Baker et al., 2005) due to the continuous sampling of fixation position and the automatic tracking of the eye in the Nidek. The distance from the computed foveal position to the average position of the fixation points was computed to derive the eccentricity of the PRL. All MD participants had developed a clear, single stable PRL (Mean = 73% of sample fixations were within 4 degrees, SE = 14, see Table 1). MD7 exhibited the least stable PRL fixation, which is understandable given she reported still using the bit of spared fovea for some daily tasks. Additionally, fixation with the spared fovea was stable for MD6 and MD7 (100% of sample fixations were within 4 degrees for each participant). For MD7 we also had auto-fluorescent images of the retina obtained with Heidelberg SLO (Courtesy of FC Delori) that clearly delineated the residual functioning retina in the middle of the lesion and confirmed the reason for the central fixation for this patient. For MD5 and MD7, we also obtained PRL locations and microperimetery using a Rodenstock SLO. The PRL locations and apparent stability of fixation were very similar with both techniques. Note that even if subjects did not fixate as instructed, no pattern of eye movements can explain the observed activation of the foveal confluence to perpherial stimuli in MD3-MD5 because the foveal part of the retina is destroyed in these subjects.
Visual field plotting
To document visual field loss, measurements were conducted using a custom computerized central perimetry system. A rear projection screen was used to present a uniform background of (luminance 97 cd/m2 and square target stimuli luminance 0.28 cd/m2 (Minolta LS-110 spot photometer)). Each eye was tested separately. Participants were instructed to maintain fixation with their PRL (or in cases with foveal sparing, with the fovea) on a fixation cross at the center of the screen while a 19mm (∼1 deg) target was moved across the screen using a mouse. In an exploration phase (aided by the MP-1 measurements) subjects were asked to report whenever the target disappeared. When the scotomatous areas were located, the target was placed inside the scotoma and moved from non-seeing to seeing regions in different directions (dynamic perimetry). The point of first seeing the target as reported by the participants was marked as the edge of the scotoma. Once the scotoma was mapped, targets were presented in random positions within the scotoma in search for any residual central vision. In all but MD6 static (seen/unseen) perimetry was also conducted in the MP-1, and it confirmed in all cases the location of the scotoma found with the custom perimeter described here.
2IFC testing
In those participants where visual field plotting showed that the scotomata covered the fovea, we further tested for residual macular function by using a 2-interval forced choice (2IFC) psychophysical test. MD participants fixated (with their PRL) on a fixation target. Each trial contained two temporal intervals, and a 1 cycle/degree horizontal Gabor patch (1 degree in size, 90% contrast) was presented to part of the retina in one of those intervals. The participant indicated the interval with the Gabor patch, guessing when they were unsure. To avoid light scatter to functioning portions of the retina, the average luminance of the test patches was identical to the background. Participants performed 50 trials with stimuli at each testing position (fovea and PRL).
2.4. fMRI
Stimulus presentation
MD and control participants completed 4 to 8 runs of simple blocked-design experiments. Participants viewed blocks (16 blocks of 15 or 16 seconds each) of images presented at either the fovea or a peripheral retinal location (the MD participant's PRL, or the control's matched peripheral location), interleaved with blocks (5 blocks of 15 or 16 seconds) of only a fixation cross (referred to as fixation baseline). In each stimulus block, 20 images were presented (MD5, MD6, and MD7 – 550 ms stimulus, 200 ms inter-stimulus interval; MD3 and MD4 – 500 ms stimulus, 30 ms inter-stimulus interval). Unless otherwise indicated, participants performed a one-back task, responding via a button box every time they saw a consecutive repetition of the same image.
Participants with similar scotomata in each eye viewed the stimuli with both eyes. However, participants with scotomata that were not consistent between the two eyes viewed the stimuli with one eye only (the preferred eye - the other eye was patched). Control participants completed exactly the same runs as their matched MD participant and viewed the stimuli with the same eye(s). MD participants fixated on a cross at the PRL location, whereas matched control participants fixated on a fixation cross at the foveal location. All other experimental parameters were identical between MD participants and controls.
MD3, MD4, and MD5
Photographs of objects (MD3 – 3 degrees of visual angle wide by 3 degrees high; MD4 – 10 degrees wide by 4 degrees high; MD5 – 3 degrees wide by 4 degrees high) were presented at the fovea and PRL in interleaved blocks. MD3 viewed stimuli with his left eye only (right eye patched), whereas MD4 and MD5 viewed the stimuli binocularly. MD3 and MD4 completed 4 runs yielding 32 object blocks per condition, whereas MD5 completed 6 runs yielding 48 object blocks per condition.
MD6 and MD7
Due to the limited size of the visual display and the distance of the PRL from the former fovea (making it impossible to present stimuli on the screen at both PRL and foveal locations in an interleaved fashion), photographs of objects or words (both 20 degrees wide by 7 degrees high) were presented on separate runs at either the fovea or the PRL. MD6 and M7 viewed the stimuli with their left eye only (right eye patched). On fovea runs, they fixated with their fovea, and on PRL runs with their PRL. To facilitate comparison with the other MD participants, data are reported here only for photographs of objects. However, exactly the same pattern of results was observed with the word stimuli. MD6 completed 8 runs (4 foveal and 4 PRL) yielding 32 object blocks per condition, whereas MD7 completed 6 runs (3 foveal and 3 PRL) yielding 24 object blocks per condition.
Functional Imaging
MD3 and MD4 were scanned on a 3.0 T Siemens Trio scanner at the A. A. Martinos Imaging Center at the McGovern Institute, MIT, Cambridge, MA. Functional images were acquired with a Siemens 12-channel phased-array head-coil and gradient echo single shot echo planar imaging sequence (22 slices, 2 × 2 × 2 mm, 0.2 mm inter-slice gap, TR = 2 s, TE = 30 ms).
MD5, MD6, and MD7 were scanned on a 3.0 T Siemens Trio scanner at the Martinos Center for Biomedical Imaging in Charlestown, MA. Functional images were acquired with a Siemens 8-channel phased-array head-coil and gradient echo single shot echo planar imaging sequence (18-24 slices, 1.4 × 1.4 × 2 mm, 0.4 mm inter-slice gap, TR = 3 s, TE = 33 ms).
Control participants were scanned on the same scanner and with the same parameters as their matched MD participants, except for the control for MD3 and MD4 who were scanned at the Martinos Center for Biomedical Imaging in Charlestown, MA with the same scanning parameters as for MD5, 6, and 7.
For all scans, slices were oriented approximately perpendicular to the calcarine sulcus. High-resolution anatomical images were also acquired for each participant for reconstruction of the cortical surface.
Data Analysis
Data were analyzed using Freesurfer and FS-FAST software (http://surfer.nmr.mgh.harvard.edu/). Prior to statistical analysis, images were motion-corrected (Cox & Jesmanowicz, 1999) and smoothed (3 mm FWHM Gaussian kernel). Activations (stimulus conditions > baseline) were visualized on the inflated and flattened cortical surface (Dale, Fischl & Sereno, 1999, Fischl, Sereno & Dale, 1999) (Figure 2).
Figure 2.

Cortical flattening and the occipital cortical patch. Examples of cortical flattening in two control participants (top row, control for MD3; bottom row, control for MD5) showing the relationship between functional activation for foveal stimuli and the anatomically defined occipital pole ROI (white outline on flattened patch). The overlaid activation maps show areas with significantly greater activation for visual objects (top row, 3 × 3 degrees; bottom row, 3 × 4 degrees) presented at the fovea compared with a fixation baseline. The flattened representations of the occipital lobe were produced by first inflating the cortex, unfolding the sulci and gyri. A cut was then made along the fundus of the calcarine sulcus and the posterior cortex (including the occipital lobe) separated from the rest of the brain. Next, the occipital lobe patch was then flattened to produce the representations on which we displayed the activation data. The foveal confluence lies at the posterior end of the calcarine sulcus, near the occipital pole and is at the apex of the cut on the flattened cortical patch. The anatomically defined occipital pole ROI clearly overlaps the activation produced by foveal stimuli. The fundus of the calcarine sulcus separates the cortical representations of the upper and lower visual field. The representation of the lower visual field is dorsal to the fundus (above the foveal confluence on the patch), while the representation of the upper visual field is ventral to the fundus (below the foveal confluence on the patch). The eccentricity of the cortical representation increases as you move anteriorly from the posterior end of the calcarine sulcus (moving away from the foveal confluence on the patch in a direction parallel to the calcarine sulcus cut).
To measure the magnitude of response in cortical regions corresponding to the representation of the fovea (i.e., the foveal confluence), a region of interest (ROI) was defined for both hemispheres of all participants based on anatomical criteria (occipital pole ROI) (Figure 2). ROIs were drawn at the posterior end of the calcarine sulcus with a surface area in each hemisphere of ∼500 mm2 (range 482-535 mm2, 176-351 voxels)(Figures 3 and 4, and Supplementary Figure 3). Since the ROIs were defined based on the individual anatomy, there was some variation in the precise shape and size of the individual ROIs. These analyses were also performed on smaller ROIs (surface area of ∼200 mm2) producing nearly identical results. Activations in occipital pole ROIs to stimuli in different locations were compared with planned one-tailed t-tests.
Figure 3.
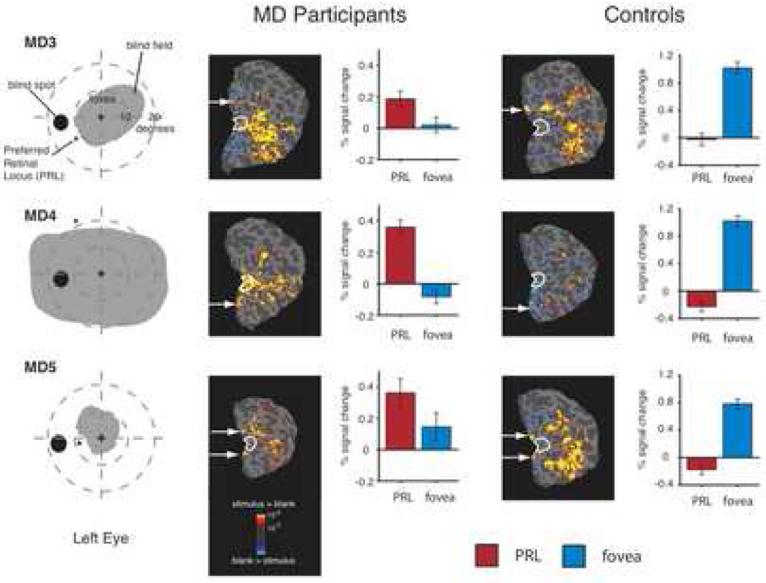
Large-scale reorganization of visual processing in three MD participants with extensive retinal lesions covering the fovea and matched control participants. Column 1. Schematic of visual fields in the left eye of each MD participant showing the large extent of the blind field (scotoma). MD3 and MD5 were tested with the right eye patched. MD4 was tested binocularly and the field loss in the right eye was very similar to that shown for the left eye. Column 2. Statistical parametric maps on the flattened cortex showing activation at the occipital pole (white outlines show the anatomically defined occipital pole ROI) for MD participants. The activation maps are displayed on the flattened cortex and show activation in response to visual objects presented at the PRL compared with the fixation baseline. In all three participants, the PRL was located in the left visual field, and data are shown for the right hemisphere only. In each case, activation was observed not only in parts of cortex corresponding to the retinal location of the PRL (white arrows), but also in the foveal confluence. Column 3. Bar charts showing percent signal change in the independently defined occipital pole ROI (white outlines). Stimuli presented at the PRL (red bars) elicited strong responses while stimuli presented at the fovea (blue bars) elicited little or no response. Column 4. Unlike the MD participants, in the control participants the flat maps show no activation at the occipital pole for stimuli presented to peripheral retina (corresponding to the matched MD participant's PRL). Column 5. Percent signal change in the occipital pole ROI for control participants shows a strong response to foveal stimuli but no response to peripheral stimuli.
Figure 4.
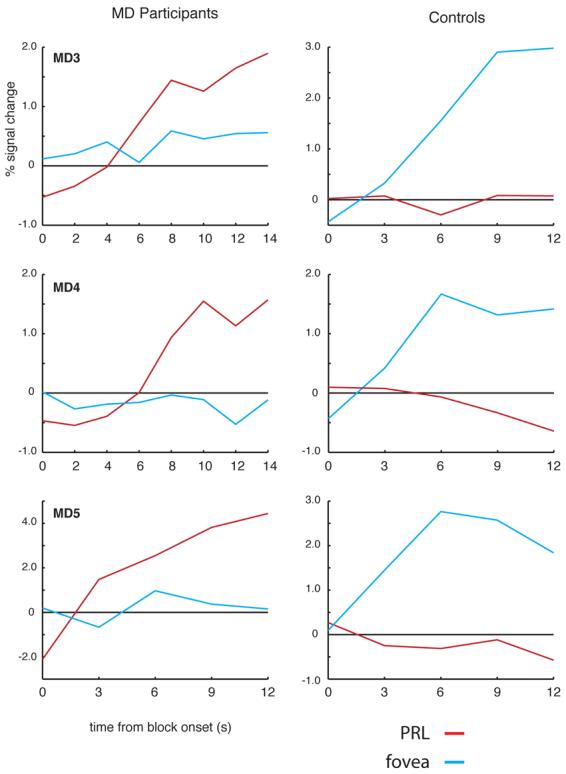
Time course of activation at the occipital pole ROI for MD3, MD4, MD5, and matched controls. Average time course of visually active voxels in the occipital pole ROI for MD participants (left column) and matched controls (right column). In each participant, voxels within the occipital pole ROI responsive to either foveal or peripheral stimuli (relative to fixation baseline) were selected using half of the total data collected. Independent time courses were plotted by averaging data from those voxels using the other half of the data. In each MD participant, there is a strong increase in activation relative to the fixation baseline for stimuli presented at the PRL (red lines) over the course of the blocks. In contrast stimuli presented at the fovea (blue lines) elicited little or no change in the activation over time. The opposite pattern was observed in the matched control participants: large increases in activation over the course of the blocks for foveal stimuli but no or even small decreases in activation over time for stimuli presented in the periphery.
To measure the time course of response to stimuli presented at the PRL and fovea within the occipital pole ROIs we used a split half analysis to first identify the relevant sub-regions of the anatomical ROI that were responsive to either foveal or PRL stimulation (p < 0.01 compared with fixation baseline), and then to extract the time course of response in those sub-regions in the other half of the data. Time courses were first detrended and then averaged across blocks according to the location of the visual stimuli (PRL or fovea) within that block. The resulting time courses show the change in signal over the course of a block.
3. Results
3.1 Complete loss of foveal function (MD3, MD4, and MD5)
3.1.1 Behavioral
MD3, MD4 and MD5 all had extensive central scotomata and complete loss of foveal function as measured by perimetry (Figure 3, Column 1). The 2IFC testing confirmed the perimetry findings, and showed that participants were at chance for stimuli presented in the center of the scotoma (former fovea), but with near perfect performance for stimuli presented at the PRL. The distance of the PRL from the former fovea ranged from approximately 8 degrees in MD5 to 22 degrees in MD4. While MD3 and MD4 were both diagnosed with juvenile macular degeneration (JMD) with onset in early adolescence, MD5 has age-related macular degeneration (AMD) which started around age 58 years (Table 1).
3.1.2 fMRI
In MD3, MD4 and MD5, the PRLs were located in the left visual field and the stimuli presented at the PRL were largely confined to the left visual field. Thus, the results presented below focus on responses in the hemisphere contralateral to the visual field location of the PRL (i.e., the right hemisphere); responses in the ipsilateral hemisphere are shown in Supplementary Figure 4).
We performed three separate analyses on the fMRI data. First we produced statistical parametric maps of the visual activation to peripheral and foveal stimuli relative to the fixation baseline in striate and extrastriate cortex. Second, we defined an ROI at the occipital pole based on anatomical criteria and examined the mean responses across this ROI to stimuli presented at both the fovea and the PRL location. Finally, we extracted the time courses for visually active voxels within the anatomically defined occipital pole ROIs to look at the change in activity over time. We describe the results of each of these analyses in turn below.
Consistent with the behavioral profile, statistical parametric mapping throughout striate and extrastriate cortex revealed that presentation of stimuli at the fovea produced no significant activation in any of MD3, MD4 or MD5, confirming the total loss of foveal function. However, visual stimulation at the PRL compared with the fixation baseline produced significant activation at the occipital pole, corresponding to the foveal confluence (white outlines in Figure 3, Column 2). As expected, significant visual activation was also observed in regions of cortex corresponding to the location of the PRL (arrows in Figure 3) and in object-selective cortex. In contrast, in control participants, significant activation to peripheral stimuli was observed in peripheral retinotopic cortex and object-selective regions, but crucially, no significant activation was observed at the occipital pole (Figure 3, Column 4).
MD3
Consistent with the PRL location in the lower half of the left visual field, significant activation corresponding to stimulation of the PRL was observed in the upper bank of the calcarine sulcus in the right hemisphere (white arrow) in both MD3 and the matched control. But in MD3 significant activation for peripheral stimuli compared with the fixation baseline was also observed at the occipital pole. This activation was not contiguous with activation corresponding to stimulation of the PRL.
MD4
Consistent with the PRL location in the upper half of the left visual field, significant activation corresponding to stimulation of the PRL was observed in the lower bank of the calcarine sulcus in the right hemisphere (white arrow) in both MD4 and the matched control. But in MD4, significant activation extended from regions of cortex representing peripheral retina into the occipital pole. In contrast to MD3, a continuous swath of significant activation was observed from the occipital pole to the regions of cortex corresponding to the PRL location.
MD5
Unlike MD3 and MD4, MD5's PRL was located close to the horizontal meridian with stimuli extending into both the upper and lower visual fields. Consistent with this PRL location, significant activity was observed in both the upper and lower banks of the calcarine sulcus (white arrows) in both MD5 and the matched control. However, in MD5 only, there was also significant activation for peripheral stimuli at the occipital pole. Similar to MD3 the observed activation at the occipital pole was not contiguous with the activation observed in regions of cortex corresponding to the PRL location.
To measure the magnitude of activation in the foveal confluence, the average activation relative to the fixation baseline within an independently defined ROI was calculated for each MD and control participant. The ROI was defined based on anatomical considerations alone without reference to the patterns of activation observed (see Methods) . In all three MD participants there was significant activation in this occipital pole ROI to stimuli presented at the PRL compared with the fixation baseline (all p < 0.001), but no significant response above baseline to stimuli presented at the fovea, as expected given the scotomata (all p > 0.1) (Figure 3, Column 3). Further, in all three of the above MD participants activation to stimuli presented at the PRL was significantly greater than activation to stimuli presented at the fovea (all p < 0.05) (see Supplementary Figure 4 for data from the hemisphere ipsilateral to the visual field location of the PRL). In matched control subjects, the opposite pattern of activation was observed (Figure 3, Column 5): No significant response above baseline in the foveal confluence in response to stimuli presented to peripheral retina (in fact, the response was less than baseline), but a significant response to stimuli presented at the fovea (all p < 0.001) compared with the fixation baseline.
The same pattern of results described above is clear in the time courses of visually active voxels within the occipital pole ROIs (Figure 4). The number of visually active voxels selected in the occipital pole ROI of each participant was 53, 114, and 16 in MD3, MD4, and MD5, respectively. In the control participants, the average number of visually active voxels was 227. In all three MD participants, stimuli presented at the fovea produced little or no change in visual activation compared with the fixation baseline over time. In contrast, presentation of stimuli at the PRL produced strong increases in activity over the course of the blocks. Again, the opposite pattern was observed in the matched control participants: little or no change in activation relative to the fixation baseline for peripheral stimuli but strong increases in activity for stimuli presented at the fovea.
Thus, all three analyses described above show significant activation for peripheral stimuli relative to the fixation baseline at the occipital pole of all MD participants with complete loss of functional foveal vision. Following the loss of foveal input, the deprived region of cortex that would normally be responsive only to foveal visual stimuli responds to peripheral visual stimuli, replicating our earlier finding of reorganization of visual processing in MD (Baker et al., 2005). We refer to this reorganization as “large-scale” because the activation at the occipital pole by peripheral stimuli is far in the cortex from the standard locus of activation for peripheral stimuli (Baker et al., 2005).
3.2. Residual foveal function (MD6 and MD7)
3.2.1 Behavioral
Both MD6 and MD7 had extensive central scotomata, similar to those of MD3-MD5 but with some sparing of central retina and residual foveal function measured by perimetry (Figure 5, Column 1). Since both MD6 and MD7 had residual foveal function measurable with standard field plotting techniques, 2IFC testing was not performed on these participants. The distance of the PRL from the fovea was approximately 18 degrees in MD6 and 20 degrees in MD7. Both MD6 and MD7 were diagnosed with JMD with onset after 30 years of age (Table 1).
Figure 5.

Lack of large-scale reorganization in two MD participants with foveal sparing and a matched control participant. Column 1. Schematic of visual fields in the left eye of MD6 and MD7. Visual fields in the other eye for both patients were similar, both showing some residual central foveal vision. The right eye of each participant was patched. Column 2. Statistical parametric maps on the flattened cortex showing activation in both hemispheres in response to visual objects presented at the PRL compared with the fixation baseline. In both participants, the PRL was located on the vertical mid-line and ROI data are shown for both hemispheres. In both participants, while activation was observed in parts of cortex corresponding to the PRL location (white arrows), no activation was observed at the occipital pole. Column 3. Bar charts showing percent signal change in the occipital pole ROI in the left (light bars) and right (dark bars) hemispheres. Stimuli presented at the fovea (blue bars) elicited small but significant responses while stimuli presented at the PRL (red bars) elicited little or no response. Column 4. Statistical parametric map showing activation elicited by peripheral stimuli compared with the fixation baseline for matched control participant. Similar to MD6 and MD7, the flat maps show no activation at the occipital pole for stimuli presented to peripheral retina. Column 5. Percent signal change in the occipital pole ROI shows a strong response to foveal stimuli but no response to peripheral stimuli. Color scale for statistical parametric maps is the same as in Figure 3.
3.2.2 fMRI
In MD6 and MD7, the PRLs were located close to or on the vertical midline in the visual field and the stimuli presented at the PRL extended into both the left and right visual fields. Thus, the results presented below focus on responses in both hemispheres.
Consistent with the residual foveal function, statistical parametric mapping showed that stimulation of the foveal retina in MD6 and MD7 produced significant activation above fixation baseline at the occipital pole. Such activation was quite weak, reflecting the fact that even though there is some foveal sparing, the fovea is still severely compromised in these participants. But in contrast to subjects MD3-MD5, who did not show such foveal sparing, visual stimulation at the PRL compared with the fixation baseline produced no significant activation at the occipital pole in MD6 and MD7 (Figure 5, Column 2). This pattern of results was qualitatively similar to that seen in the control participant: Significant activation above fixation baseline for foveal stimuli but no significant response to stimuli presented to peripheral retina (Figure 5, Column 4).
In both MD6 and MD7, the PRL was located close to the vertical meridian in the lower visual field. Thus, significant activation corresponding to stimulation of the PRL was observed in the upper bank of the calcarine sulcus in both hemispheres for MD6 and MD7 and the matched control. However, while significant activation consistent with the peripheral stimulation is clearly visible, there was no significant activation above fixation baseline at the occipital pole, and in MD7 there was actually less activity for peripheral stimulation compared to fixation baseline.
Within the anatomically defined occipital pole ROI, there was no significant activation elicited by stimuli presented at the PRL (all p > 0.35) in contrast to MD3, MD4 and MD5. However, activation above baseline was observed for stimuli presented at the fovea (Figure 5). In MD7, activation by foveal stimuli was significantly greater than baseline in both hemispheres (both p < 0.03), and significantly greater than activation produced by stimuli at the PRL (both p < 0.002). In MD6, the greater activation by foveal stimuli compared to baseline was marginally significant (right hemisphere, p < 0.06; left hemisphere, p< 0.06), although the difference between foveal and PRL stimuli only approached significance in the right hemisphere (right hemisphere, p < 0.07; left hemisphere, p > 0.15). The weak activation of the occipital pole by foveal stimuli in MD6 and MD7 reflects the fact that even though there is some foveal sparing, the foveal retina is still severely compromised.
The same pattern of response is also evident in the time course of visually active voxels (Figure 6). The number of visually active voxels selected in the occipital pole ROI of each participant was 26 and 55 for the right hemisphere and 22 and 45 for the left hemisphere in MD6 and MD7, respectively. In the control participant, the number of visually active voxels was 242 and 206 in the right and left hemispheres, respectively. While there are only weak increases in activation to foveal stimuli, critically, in contrast to MD3, MD4 and MD5, there is no increase in activation above baseline over time for stimuli presented at the PRL and the activation for peripheral stimuli is less than for foveal stimuli.
Figure 6.
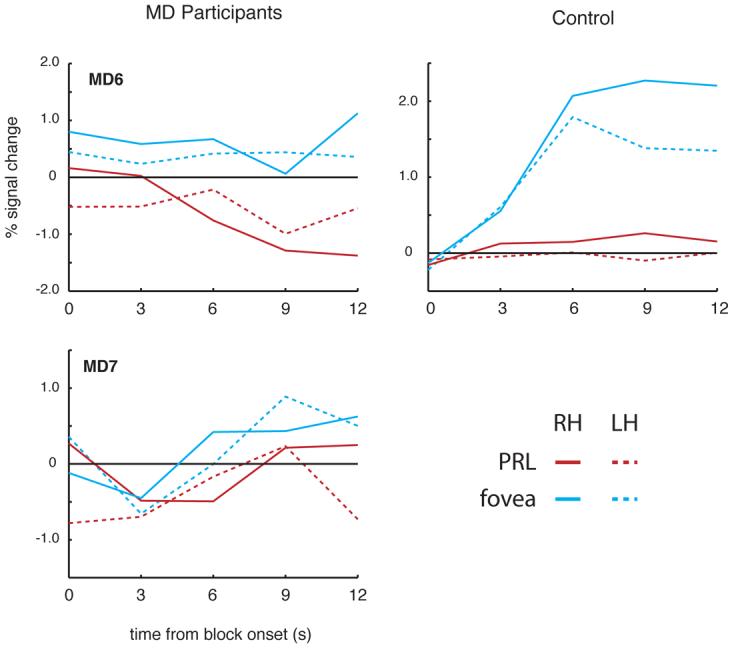
Time course of activation at the occipital pole for MD6 and MD7 and matched control. In both MD participants (left column), stimuli presented at the fovea (blue lines) elicited weak increases in activation relative to the fixation baseline in both the right (solid lines) and left hemisphere (dotted lines). In contrast to MD3, MD4 and MD5, however, stimuli presented at the PRL (red lines) elicited weak decreases or little change in activation relative to fixation over the course of the blocks, and the activation for foveal stimuli was greater than for peripheral stimuli. In the matched control participant, while the change in activation for foveal stimuli was much stronger than in MD6 and MD7, since there was no damage to the fovea, the same qualitative pattern was observed.
In the matched control participant exactly the same qualitative pattern of activation was observed as in MD6 and MD7: Response within the occipital pole for foveal stimuli in both hemispheres (both p < 0.0001), but not for peripheral stimuli (both p > 0.09) with significantly stronger activation by foveal than peripheral stimuli (both p < 0.0001) (Figure 5, Column 5; Figure 6).
Although there were some differences in the testing procedure for MD6 and MD7 compared with the other MD participants (e.g., stimulus locations interleaved across runs rather than within runs), these differences cannot account for the differences in activation we observed across participants. In particular, MD4 was scanned a second time using the same scanner and protocol as for MD6 and MD7, and again significant activation was observed at the occipital pole for stimuli presented at the PRL (Supplementary Figure 5). Further, the strong activation in peripheral retinotopic cortex for stimuli presented at the PRL in MD6 and MD7 (Figure 5, column 2) shows that there is sufficient power to detect visual activations in retinotopic cortex. Therefore the absence of activation at the occipital pole in participants with some foveal sparing is not the result of differences in testing procedure.
Thus, in participants with some sparing of foveal vision despite extensive retinal lesions, we find no evidence for significant activation of the occipital pole (corresponding to the foveal confluence) by peripheral stimuli. These data suggest that complete loss of foveal input is necessary for large-scale reorganization to occur.
4. Discussion
In three individuals with complete loss of foveal vision from macular degeneration we report activation of the occipital pole (corresponding to the foveal confluence) by peripheral visual stimuli, replicating our earlier report of large-scale reorganization of visual processing (Baker et al., 2005). Further we show that such reorganization is only observed when foveal function is completely lost.
Taking our two studies together, we have now found evidence for large-scale reorganization of visual processing in five individuals with macular degeneration. The age of onset of macular degeneration, time since onset and specific diagnosis vary widely across these participants, and thus do not seem to be the main factor that determines whether such reorganization occurs. MD2, MD3 and MD4 all have JMD with an onset in early adolescence. In contrast, MD5 has AMD with onset age 58, and MD1 has cone-rod dystrophy with onset age 37. What is consistent across these individuals is the complete loss of input to the foveal confluence. In contrast, in two individuals (MD6 and MD7) with some spared foveal function, we found no evidence for large-scale reorganization of visual processing. Thus, activation of the foveal confluence by peripheral stimuli does not simply arise because of macular degeneration, but is specifically caused by the complete loss of visual input to this region. This raises the possibility that the previously published failure to find any evidence for reorganization in a single person with AMD arose because of spared foveal input (see figure 2 in Sunness et al., 2004), although as noted previously (Baker et al., 2005), the strength of this null result is questionable because of the sparsity of data and ambiguity of the imaging results. Similarly, the absence of any activation at the occipital pole for peripheral stimuli in one MD participant in a recent study (JMD4 in Masuda et al., 2008) may also reflect some degree of foveal sparing. While we cannot rule out the importance of factors such as age of onset or type of macular degeneration, our results suggest that the severity of foveal loss is a critical factor. Here, we focus on what we have previously termed large-scale reorganization of visual processing (Baker et al., 2005); that is, activation of foveal confluence by peripheral stimuli following central retinal damage. We use the term “reorganization” to refer to the profound changes in the retinotopic maps, without implicating any particular underlying mechanism (see also below), similar to previous authors (Baseler et al., 2002, Calford & Tweedale, 1988, Merzenich, Kaas, Wall, Nelson, Sur & Felleman, 1983, Merzenich, Nelson, Stryker, Cynader, Schoppmann & Zook, 1984). While we find no evidence for such large-scale reorganization in MD6 and MD7 we cannot rule out the possibility of more subtle reorganization close to the representation of the scotoma border. The cortical representation of the scotoma is very difficult to localize, and the precise representation of the border impossible to identify reliably based on the location of visual field loss. Thus, while reorganization of visual processing is not large enough to produce activation of the foveal confluence by stimuli presented at the PRL (far away from the fovea), some local reorganization on the representation of the scotoma border may occur.
Our finding of large-scale reorganization of visual processing in adult human visual cortex dovetails with converging neural and behavioral evidence for cortical reorganization of V1 in a stroke patient (BL) with optic radiation damage (Dilks et al., 2007) and with evidence for reorganization of cortical maps in rod monochromats (Baseler et al., 2002). However, rod monochromacy is congenital and reorganization in these individuals might reflect only developmental plasticity. Although we can't determine the cortical regions (i.e., V1-V4) showing reorganization of visual processing in MDs, because the foveal representations of all visual areas come together at the occipital pole (Dougherty et al., 2003), the data from BL (Dilks et al., 2007) suggests that V1 might be involved.
How does the reorganization of visual processing arise? One possible mechanism is disinhibition of pre-existing long-range horizontal connections within V1 (Darian-Smith & Gilbert, 1994; Das & Gilbert, 1995). To explain the extent of activation observed in our MD patients, however, horizontal connections would have to spread activation farther than the length of typical horizontal connections in primate V1 (i.e., 6-8 mm) (Angelucci, Levitt, Walton, Hupe, Bullier & Lund, 2002, Gilbert, Das, Ito, Kapadia & Westheimer, 1996), and a poly-synaptic chain of connections would be required. A second potential mechanism for reorganization involves the growth of new horizontal connections (Darian-Smith & Gilbert, 1994), rather than the unmasking of existing connections. Finally, a third possible source of reorganization could be the unmasking of intracortical feedback to V1 from higher visual areas. At this time our data cannot distinguish between these three alternatives, and no other experiments in humans or animals have definitively pinpointed the respective contributions of horizontal and feedback connections in cortical reorganization.
It was recently reported that activation of the foveal confluence by peripheral stimuli was observed only when participants were performing a task(Masuda et al., 2008). This observation was taken to indicate that activation of the foveal cortex by peripheral stimuli reflects unmasking of feedback connections from extrastriate visual cortex (Mechanism 3 above). However, modulation of activity by a task does not provide strong support for a solely top-down mechanism of reorganization, because it is well established that bottom-up visual responses can be modulated by attention. Importantly, we have observed foveal confluence activity in both MD1 and MD2 during passive viewing of flickering checkerboards (Supplementary Figure 6) and in MD1 during passive viewing of visual words (Experiment 2 in Baker et al., 2005). These activations appear to be weaker than for complex visual stimuli with a task, consistent with numerous other reports that attention modulates responses in visual cortex including V1. Modulation by task and/or stimuli provides evidence for a top-down component, but it is not clear if this top-down influence differs from the standard attentional modulation of bottom-up visual responses observed in participants with full-field vision. Thus, the mechanism of reorganization remains an open question.
In conclusion, we have demonstrated again clear evidence for large-scale reorganization of visual processing in adult humans following loss of cortical input from macular degeneration. Furthermore, we have shown that such large-scale reorganization (activation of foveal confluence by peripheral stimuli) requires complete loss of functional foveal vision. Future work should explore the mechanisms underlying this reorganization.
Supplementary Material
Acknowledgements
We express our sincere thanks to the participants with macular degeneration. We would also like to thank the Athinoula A. Martinos Imaging Center at the McGovern Institute for Brain Research, MIT, Cambridge, MA, and the Martinos Center for Biomedical Imaging, Charlestown, MA. This work was supported by NIH grants EY016559 (NK), EY005957 (EP), a Kirschstein-NRSA EY017507 (DDD), and by Dr. and Mrs. Joseph Byrne.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contribution statement
CIB, EP, and NK designed the experiments. CIB and DDD collected and analyzed functional imaging data. EP selected MD participants and collected behavioral data. CIB and DDD wrote the paper, with contributions from EP and NK.
References
- Angelucci A, Levitt JB, Walton EJ, Hupe JM, Bullier J, Lund JS. Circuits for local and global signal integration in primary visual cortex. J Neurosci. 2002;22(19):8633–8646. doi: 10.1523/JNEUROSCI.22-19-08633.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker CI, Peli E, Knouf N, Kanwisher NG. Reorganization of visual processing in macular degeneration. J Neurosci. 2005;25(3):614–618. doi: 10.1523/JNEUROSCI.3476-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baseler HA, Brewer AA, Sharpe LT, Morland AB, Jagle H, Wandell BA. Reorganization of human cortical maps caused by inherited photoreceptor abnormalities. Nat Neurosci. 2002;5(4):364–370. doi: 10.1038/nn817. [DOI] [PubMed] [Google Scholar]
- Calford MB, Chino YM, Das A, Eysel UT, Gilbert CD, Heinen SJ, Kaas JH, Ullman S. Neuroscience: rewiring the adult brain. Nature. 2005;438(7065):E3. doi: 10.1038/nature04359. discussion E3-4. [DOI] [PubMed] [Google Scholar]
- Calford MB, Tweedale R. Immediate and chronic changes in responses of somatosensory cortex in adult flying-fox after digit amputation. Nature. 1988;332(6163):446–448. doi: 10.1038/332446a0. [DOI] [PubMed] [Google Scholar]
- Cheung S-H, Schuchard RA, He S, Tai Y, Legge GE, Hu XP. Limited retinotopic reorganization in age-related macular degeneration. Journal of Vision. 2005;5(8):299–299. [Google Scholar]
- Chino YM, Kaas JH, Smith EL, 3rd, Langston AL, Cheng H. Rapid reorganization of cortical maps in adult cats following restricted deafferentation in retina. Vision Res. 1992;32(5):789–796. doi: 10.1016/0042-6989(92)90021-a. [DOI] [PubMed] [Google Scholar]
- Chino YM, Smith EL, 3rd, Kaas JH, Sasaki Y, Cheng H. Receptive-field properties of deafferentated visual cortical neurons after topographic map reorganization in adult cats. J Neurosci. 1995;15(3 Pt 2):2417–2433. doi: 10.1523/JNEUROSCI.15-03-02417.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox R, Jesmanowicz A. Real-time 3D image registration for functional MRI. Magnetic Resonance in Medicine. 1999;42:1014–1018. doi: 10.1002/(sici)1522-2594(199912)42:6<1014::aid-mrm4>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage. 1999;9(2):179–194. doi: 10.1006/nimg.1998.0395. [DOI] [PubMed] [Google Scholar]
- Dilks DD, Serences JT, Rosenau BJ, Yantis S, McCloskey M. Human adult cortical reorganization and consequent visual distortion. J Neurosci. 2007;27(36):9585–9594. doi: 10.1523/JNEUROSCI.2650-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dougherty RF, Koch VM, Brewer AA, Fischer B, Modersitzki J, Wandell BA. Visual field representations and locations of visual areas V1/2/3 in human visual cortex. J Vis. 2003;3(10):586–598. doi: 10.1167/3.10.1. [DOI] [PubMed] [Google Scholar]
- Fischl B, Sereno M, Dale A. Cortical surface-based analysis II: Inflation, flattening, and a surface-based coordinate system. NeuroImage. 1999;9(2):195–207. doi: 10.1006/nimg.1998.0396. [DOI] [PubMed] [Google Scholar]
- Fitzgibbon AW, Pilu M, Fisher RB. Direct least-squares fitting of ellipses. IEEE Transaction of Pattern Analysis and Machine Intelligence. 1999;21(5):476–480. [Google Scholar]
- Giannikopoulos DV, Eysel UT. Dynamics and specificity of cortical map reorganization after retinal lesions. Proc Natl Acad Sci U S A. 2006;103(28):10805–10810. doi: 10.1073/pnas.0604539103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert CD, Das A, Ito M, Kapadia M, Westheimer G. Spatial integration and cortical dynamics. Proc Natl Acad Sci U S A. 1996;93(2):615–622. doi: 10.1073/pnas.93.2.615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert CD, Wiesel TN. Receptive field dynamics in adult primary visual cortex. Nature. 1992;356(6365):150–152. doi: 10.1038/356150a0. [DOI] [PubMed] [Google Scholar]
- Heinen SJ, Skavenski AA. Recovery of visual responses in foveal V1 neurons following bilateral foveal lesions in adult monkey. Exp Brain Res. 1991;83(3):670–674. doi: 10.1007/BF00229845. [DOI] [PubMed] [Google Scholar]
- Horton JC, Hocking DR. Monocular core zones and binocular border strips in primate striate cortex revealed by the contrasting effects of enucleation, eyelid suture, and retinal laser lesions on cytochrome oxidase activity. J Neurosci. 1998;18(14):5433–5455. doi: 10.1523/JNEUROSCI.18-14-05433.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaas JH, Krubitzer LA, Chino YM, Langston AL, Polley EH, Blair N. Reorganization of retinotopic cortical maps in adult mammals after lesions of the retina. Science. 1990;248(4952):229–231. doi: 10.1126/science.2326637. [DOI] [PubMed] [Google Scholar]
- Masuda Y, Dumoulin SO, Nakadomari S, Wandell BA. V1 Projection Zone Signals in Human Macular Degeneration Depend on Task, not Stimulus. Cereb Cortex. 2008 doi: 10.1093/cercor/bhm256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masuda Y, Nakadomari N, Asakawa K, Dumoulin SO, Liu J, Cheung S, Ben-Schachar M, Kitahara K, Wandell BA. Society for Neuroscience. Atlanta, GA: 2006. Lack of cortical reorganization in macular degeneration patients. [Google Scholar]
- Merzenich MM, Kaas JH, Wall J, Nelson RJ, Sur M, Felleman D. Topographic reorganization of somatosensory cortical areas 3b and 1 in adult monkeys following restricted deafferentation. Neuroscience. 1983;8(1):33–55. doi: 10.1016/0306-4522(83)90024-6. [DOI] [PubMed] [Google Scholar]
- Merzenich MM, Nelson RJ, Stryker MP, Cynader MS, Schoppmann A, Zook JM. Somatosensory cortical map changes following digit amputation in adult monkeys. J Comp Neurol. 1984;224(4):591–605. doi: 10.1002/cne.902240408. [DOI] [PubMed] [Google Scholar]
- Rohrschneider K, Springer C, Bultmann S, Volcker HE. Microperimetry--comparison between the micro perimeter 1 and scanning laser ophthalmoscope--fundus perimetry. Am J Ophthalmol. 2005;139(1):125–134. doi: 10.1016/j.ajo.2004.08.060. [DOI] [PubMed] [Google Scholar]
- Schuchard RA, Fletcher DC. Preferred retinal locus - a review with applications in low vision rehabilitation. In: Colenbrader A, Fletcher DC, editors. Low Vision and Vision Rehabilitation. W. B. Saunders Co.; 1994. pp. 243–256. [Google Scholar]
- Smirnakis SM, Brewer AA, Schmid MC, Tolias AS, Schuz A, Augath M, Inhoffen W, Wandell BA, Logothetis NK. Lack of long-term cortical reorganization after macaque retinal lesions. Nature. 2005;435(7040):300–307. doi: 10.1038/nature03495. [DOI] [PubMed] [Google Scholar]
- Smirnakis SM, Schmid MC, Brewer AA, Tolias AS, Schuz A, Augath M, Inhoffen W, Wandell BA, Logothetis NK. Neuroscience - Rewiring the adult brain - Reply. Nature. 2005;438(7065):E3–E4. doi: 10.1038/nature04359. [DOI] [PubMed] [Google Scholar]
- Sunness J, Liu T, Yantis S. Retinotopic Mapping of the Visual Cortex Using Functional Magnetic Resonance Imaging in a Patient with Central Scotomas from Atrophic Macular Degeneration. Ophthalmology. 2004;111:1595–1598. doi: 10.1016/j.ophtha.2003.12.050. [DOI] [PubMed] [Google Scholar]
- Timberlake GT, Mainster MA, Peli E, Augliere RA, Essock EA, Arend LE. Reading with a macular scotoma. I. Retinal location of scotoma and fixation area. Invest Ophthalmol Vis Sci. 1986;27(7):1137–1147. [PubMed] [Google Scholar]
- Timberlake GT, Sharma MK, Grose SA, Gobert DV, Gauch JM, Maino JH. Retinal location of the preferred retinal locus relative to the fovea in scanning laser ophthalmoscope images. Optom Vis Sci. 2005;82(3):177–185. doi: 10.1097/01.opx.0000156311.49058.c8. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.