

Abstract
Recombinant human factor VIII (rFVIII), a multidomain glycoprotein is used in replacement therapy for treatment of hemophilia A. Unfortunately, 15%–30% of the treated patients develop inhibitory antibodies. The pathogenesis of antibody development is not completely understood. The presence of aggregated protein in formulations is generally believed to enhance the immune response. rFVIII has a tendency to aggregate but the effect of such aggregation on the immunogenicity of rFVIII is not known. We have, therefore, characterized aggregated rFVIII produced by thermal stress and evaluated its effect on the immunogenicity of rFVIII in hemophilia A mice. Aggregated rFVIII alone and mixtures of rFVIII with aggregated rFVIII were less immunogenic than native rFVIII. In vitro Th-cell proliferation studies and cytokine analyses conducted on splenocytes obtained from immunized animals suggest that aggregated rFVIII behaves as a unique antigen compared to native monomeric rFVIII. The antigenic properties of the aggregated and native rFVIII were compared using ELISAs (epitope availability) and cathepsin-B (an antigen processing enzyme) digestion. The data suggest significant differences in the antigenic properties of rFVIII and aggregated rFVIII. Overall it appears that aggregated rFVIII does not enhance the immunogenicity (inhibitor development) of rFVIII in hemophilia A mice but rather acts as a distinct antigen.
Keywords: circular dichroism, fluorescence spectroscopy, hemophilia A, inhibitor development, protein aggregation, protein structure, recombinant human Factor VIII, immunogenicity, immunology
INTRODUCTION
Factor VIII (FVIII) or antihemophilic factor is a multidomain glycoprotein, which is an essential cofactor in the coagulation cascade. FVIII is comprised of six domains, NH2-A1-A2-B-A3-C1-C2-COOH.1,2 The protein is synthesized as a single polypeptide chain which is 2351 amino acids long. The polypeptide is further proteolytically processed to form the heavy chain (A1-A2-B) and the light chain (A3-C1-C2).3 Finally, the heavy chain is further cleaved by proteases at several sites between the A2 and B domains generating polypeptides with molecular masses ranging from 90 to 220 kDa.3 The functional FVIII, is a heterodimeric molecule which is comprised of the heavy chain and the light chain held together by a divalent calcium ion.4,5
The deficiency or dysfunction of FVIII activity causes hemophilia A, a life-threatening bleeding disorder. Replacement therapy with plasma derived or recombinant FVIII preparations is the treatment of choice for controlling bleeding episodes in hemophilia A patients.6 Administration of exogenous FVIII, however, leads to the development of inhibitory antibodies (inhibitors) in 15%–30% of hemophilia A patients.7,8 The pathogenesis of inhibitor development is not completely understood.9 Both genetic as well as environmental factors related to product type, dosing regimen, etc., are suspected of triggering the development of inhibitors.10
The presence of antibodies against a therapeutic protein is often detrimental to its therapeutic utility in the management of the disease.11 Factors related to a protein product that can influence its immunogenicity include protein aggregation, protein sequence, frequency of administration and the particular excipients used in the formulation.12–15 Presence of aggregates in a protein product can not only compromise the efficacy of the product due to loss of activity15,16 but also enhance immunogenicity of protein preparations.17 Problems of immunogenicity for proteins such as human growth hormone18 and interferon-α15 have been attributed to the presence of aggregates in the respective protein preparations. Since rFVIII has a tendency to undergo aggregation19–21 the presence of trace amounts of aggregates formed during manufacture or handling of rFVIII products may be a factor contributing to its immunogenicity. No systematic study has yet been undertaken to investigate the impact of rFVIII aggregates on antibody development. In this study, we have evaluated the effect of aggregation on the immunogenicity of rFVIII in a murine model of hemophilia A. Our data suggests that aggregated rFVIII differs significantly as an antigen from the monomeric form of rFVIII.
MATERIALS AND METHODS
Materials
rFVIII and monoclonal antibody 8860 were obtained from Baxter Healthcare Corporation (Carlsbad, CA). Monoclonal antibodies ESH4, ESH5, and ESH8 were obtained from American Diagnostica, Inc., (Greenwich, CT). Monoclonal antibodies OBT-0037 and N77210M were purchased from Oxford Biotechnology (Oxfordshire, UK) and Biodesign International (Saco, ME), respectively. Sterile, pyrogen free water for injection was obtained from Henry Schein, Inc. (Melville, NY). Goat antimouse-Ig and antirat-Ig, alkaline phosphatase conjugates were obtained from Southern Biotechnology Associates, Inc. (Birmingham, AL). Normal coagulation control plasma and FVIII deficient plasma for the activity assays were purchased from Trinity Biotech (Co. Wicklow, Ireland). Platelin L reagent used in aPTT and Bethesda assays was purchased from BioMerieux (Durham, NC). The aPTT and Bethesda assay were performed using a COAG-A-MATE coagulation analyzer from Organon Teknika Corporation, (Durham, NC). Bovine cathepsin B (10 U/mg protein), dithiothreitol, disodium-EDTA, Ig-free bovine serum albumin (BSA), sodium pyruvate and diethanolamine were obtained from Sigma, (St. Louis, MO). p-nitro-phenyl phosphate was purchased from Pierce (Rockford, IL). RPMI-1640 culture medium, penicillin, streptomycin, L-glutamine, 2-mercaptoethanol, and polymyxin-B were all obtained from Invitrogen Corp., (Carlsbad, CA). 3H-thymidine was obtained from Perkin Elmer, Inc. (Boston, MA). All buffer salts and solvents were purchased from Fisher Scientific, (Fair Lawn, NJ) and used without further purification.
Circular Dichroism Spectroscopy
Far UV circular dichroism (CD) spectroscopy was used to determine potential secondary structure changes in aggregated rFVIII relative to the native protein. CD spectra were acquired on a JASCO-715 spectropolarimeter calibrated with d-10 camphor sulfonic acid. The spectra were obtained from 250 to 205 nm in a 0.1 cm quartz cuvette. rFVIII and aggregated rFVIII concentrations typically used for all experiments were 25 μg/100 μL in tris buffer (25 mM Tris, 5 mM CaCl2 and 300 mM NaCl, pH 7.0). The CD spectra were corrected by subtracting the baseline of the tris buffer.
Size Exclusion Chromatography and SDS–PAGE
SEC was performed with a Biosep-SEC4000S, 300 × 4.6 mm2 (Phenomenex, Torrance, CA) column with an exclusion limit of 2,000 kDa. Prior to use, the column was calibrated using a standard protein mixture of known molecular weights from Biorad (Hercules, CA). The chromatograph system consisted of an autoinjector with sample cooler (maintained at 4°C), column oven (maintained at 20°C), fluorescence detector, integrator from Shimadzu (Shimadzu, Braintree, MA) and a Waters 510 isocratic pump (Waters, Milford, MA). Tris buffer was used as the mobile phase at a flow rate of 0.4 mL/min and eluent was monitored by the fluorescence detector at an excitation of 285 nm and emission of 335 nm. Fifty microliter sample injection volumes were used for all samples. rFVIII or Aggregated rFVIII in tris buffer at a total protein concentration of 20 μg/mL were analyzed by SEC.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS–PAGE) was performed according to the procedure of Laemmli (Biorad, Hercules, CA) under reducing and nonreducing conditions. Samples (30 μL) containing 6 μg of total protein were loaded onto 4%–15% Tris-HCL precast acrylamide gels (Biorad, Hercules, CA) and subjected to electrophoresis under a constant voltage of 120 V for 65 min. The gel was stained with coomasie blue and densitometric scans were obtained using Kodak Image Station 2000MM (Kodak, Rochester, NY).
Steady State Fluorescence Anisotropy Studies
Changes in the steady-state emission anisotropy (r) of rFVIII upon aggregation were measured by a PTI-Quantamaster fluorescence spectrophotometer, equipped with a peltier unit and motorized Glan–Thompson polarizing prisms. The concentration of the protein (rFVIII or aggregated rFVIII) used was 5 μg/mL and a variable path length cuvette was used to minimize the inner filter effect. Samples were excited at 280 nm and the emission was monitored at 335 nm. Excitation and emission slit widths were set at 4 nm.
Anisotropy was calculated using the following equation:22
| Eq. 1 |
where, III = emission intensity with the emission polarizer oriented parallel to the excitation polarizer, I⊥ = emission intensity with the emission polarizer oriented perpendicular to the excitation polarizer, and G = instrument specific G-factor (G = III/I⊥).
Animals
A colony of C57BL/6J mice, with a deletion of exon 16 of the FVIII gene (hemophilia A mice) was established with breeding pairs from the original colony.23 Since the sex of the animal appears to have no detectable impact on the immune response, equal numbers of adult male and female mice (8–12 weeks), were used for these studies.24 Hemophilia A mice represent a nontolerized (to human rFVIII) animal model which upon immunization with rFVIII produces a qualitatively similar immune response to that observed in humans25 and is, therefore, valuable for studying relative immunogenicity and mechanisms of immune responses against rFVIII.26,27
Preparation of Antigens for Immunization
Native rFVIII (rFVIII) was dissolved in tris buffer for administration in hemophilia A mice. Tris buffer is a pharmaceutically relevant buffer and is a component of a currently marketed formulation.28 Aggregated rFVIII (Agg) was prepared by incubating rFVIII in tris buffer at 80°C for 2 min. Thermal stress was used to prepare aggregates since thermal stress has been used to mimic pharmaceutical stress conditions29 and to evaluate excipients for protein formulations.30 SEC profiles of samples confirmed the completion of aggregation (Fig. 1b). rFVIII containing 5% of Agg (Agg5) was prepared by mixing 95% rFVIII with 5% Agg. rFVIII containing 20% of Agg (Agg20) was prepared by mixing 80% rFVIII with 20% Agg. For both Agg5 and Agg20 the Agg was prepared separately and subsequently mixed with the corresponding amount of rFVIII. SEC profiles of Agg5 and Agg20 confirmed the presence of the predicted amount of both aggregated and native rFVIII in these samples (data not shown). For all the preparations, sterile, pyrogen free, tris buffer was used. All samples were handled under aseptic conditions.
Figure 1.

(a) The far UV-CD spectrum of rFVIII and aggregated rFVIII in tris buffer (300 mM NaCl, 25 mM Tris and 5 mM CaCl2, pH = 7.0). The concentration of the protein was ~25 μg/100 μL and the path length of the quartz cuvette was 0.1 cm. (b) Representative size exclusion chromatography (SEC) profiles of rFVIII, aggregated rFVIII (Agg) and mixture of rFVIII + Agg. Elution time is shown in minutes. (c) Steady state fluorescence anisotropy of rFVIII and aggregated rFVIII. The protein concentration was ~5 μg/mL. *Independent t-test, p value <0.05. (d) Densitometric scans of the polypeptide bands obtained in SDS–PAGE of rFVIII and aggregated rFVIII under reducing and non-reducing conditions.
Immunization and Sampling
Equal numbers of male (n = 6) and female (n = 6) mice of ages 8–12 weeks were immunized with four subcutaneous (s.c.) injections 1 week apart of each antigen. Each antigen dose consisted of 2 μg of total protein in 100 μL of tris buffer. The s.c. route of administration was chosen to amplify the immune response since administration by the i.v. route resulted in very low titers, which were inadequate for any meaningful statistical analysis of comparison between groups.31 Furthermore, studies by Reipert et al., have demonstrated similar IgG subtype levels after s.c. and i.v. administration in hemophilia A mice, suggesting an identical mechanism of immune response for the s.c. and i.v. routes.25 The mice were sacrificed 6 weeks after the first antigen dose and blood samples were obtained in acid citrate dextrose buffer by cardiac puncture. Previous studies have demonstrated that 6 weeks is an appropriate time for comparison of antibody levels between various treatment groups.31 Plasma samples were stored at −80°C until analyses. All studies were performed in accordance with the guidelines of Institutional Animal Care and Use Committees (IACUC) at the University at Buffalo.
Measurement of Total rFVIII Antibody Titers
Standard antibody-capture, ELISA was used to determine total anti-rFVIII antibody titers as previously described.31 Breifly, Nunc-Maxisorb 96-well plates were coated with rFVIII (2.5 μg/mL in carbonate buffer) and subsequently blocked with 1% bovine serum albumin (blocking buffer). Fifty microliter per well of various dilutions of the sample (1:100–1:40000) in blocking buffer and standard concentrations (12.5–150 μg/mL) of ESH8 antibody were incubated at 37°C for 1 h. The plate wells were washed and incubated with 50 μL of a 1:1000 dilution in blocking buffer of goat antimouse-Ig (IgG + IgM + H + L)-alkaline phosphatase conjugate (SouthernBiotech, Birmingham, AL) at room temperature for 1 h. Plates were washed and incubated with 100 μL of 1 mg/mL p-nitrophenyl phosphate solution in diethanolamine buffer (consisting of 1 M diethanolamine and 0.5 mM MgCl2) for 30 min at room temperature for color development. Hundred microliter of 3N NaOH was added to quench the reaction. Optical density at 405 nm was measured using a plate reader. Samples were analyzed in triplicate.
Data Analysis
To account for the plate to plate variability a plate specific parameter (PSP) was calculated for each plate to normalize the data. The PSP for each plate was obtained from the standard ESH8 antibody used in each experiment as follows: The concentrations of ESH8 (within the linear portion of the sigmoidal standard curve) and the corresponding optical densities were plotted on a linear scale and a linear regression analysis was performed. The regression analysis was used to calculate the maximum and minimum predicted absorbance. PSP was calculated as the half of the difference between the maximum and minimum predicted optical density. The optical densities of the various dilutions (1:100–1:40000) of the samples were plotted against the corresponding log of dilution and a linear regression analysis was performed on the linear part of the curve. The regression analysis was used to calculate the dilution that gave an optical density equal to the PSP. The dilution so obtained was the antibody titer for the sample.
Measurement of Inhibitory rFVIII Titers
Nijmegen modification of the Bethesda (NMB) assay32 was used to determine the inhibitory antibody titers as previously described.31 The assay is a modification of the one stage aPTT assay and measures the ability of rFVIII antibodies to inhibit the biological activity of rFVIII. Briefly, serial dilutions (1:2–1:32000) of the mouse plasma samples obtained from immunized animals were prepared using FVIII deficient plasma as the diluent (Trinity Biotech, CA). Equal volumes (100 μL) of the diluted mouse plasma samples and normal human plasma were mixed and incubated at 37°C for 2 h. At the end of the incubation residual activity was determined using aPTT assay. The residual activity was plotted against the corresponding log of the dilution and a linear regression analysis was performed on the linear part of the curve. The regression analysis was used to calculate the inhibitory titer for the sample which is the dilution causing a 50% reduction in biological activity. Inhibitory titers were expressed in Bethesda Units (BU).
Detection of Th (CD4+) Cell Responses
Hemophilia A mice (n = 3), aged 8–12 weeks were immunized with two subcutaneous (s.c.) injections of rFVIII or Agg (2 μg) at weekly intervals. Mice immunized with tris buffer alone served as controls. Animals were sacrificed 3 days after the second injection and their spleens were isolated. A unicellular suspension of splenocytes was prepared from the individual spleens and used as a source for the CD4+ cells. The splenocytes were depleted of CD8+ cells by using magnetic beads (Dynal Biotech, Oslo, Norway) coated with a rat antimouse monoclonal antibody for the Lyt 2 membrane antigen. Cells 2 × 105/200 μL/well were cultured in a 96-well flat bottom plates with rFVIII or Agg (1000 ng of protein antigen/well) in complete RPMI-1640 culture medium containing 10000 U/mL penicillin, 10 mg/mL streptomycin, 2.5 mM sodium pyruvate, 4 mM L-Glutamine, 0.05 mM 2-mercaptoethanol, 2 mg/mL Polymyxin B, and 0.5% heat inactivated hemophilic mouse serum. After 72 h of culture at 37°C, 1 μCi of 3H-thymidine (6.7 Ci/mmol)/well, was added and incubated for an additional 16 h. At the end of the incubation the cells were harvested using a Micromate Harvester (Packard, Meriden, CT) and 3H-thymidine incorporation was measured using a TopCount™ microplate scintillation and luminescence counter (Packard Instrument Company, Meriden, CT). The cells from individual mice were cultured in quadruplicates and stimulation indexes (SI) were obtained for individual mice. Representation of the results as SI allows us to normalize the data for comparison between experiments conducted at different times and with different animals. SI is the ratio of the average counts per minute (cpm) of cells incubated with rFVIII to the average counts per minute (cpm) of the cells incubated without the antigen.
Cytokine Analysis
The supernatant media obtained from cells incubated for 72 h under conditions identical to those described above were used for detection of secreted IL-10 or IFN-γ. IFN-γ was employed as a representative cytokine to detect Th1 responses and IL-10 was used to detect Th2 responses. The supernatants were analyzed for the respective cytokines using a standard antibody capture ELISA kit (R&D systems, Minneapolis, MN).
Detection of Known rFVIII Epitopes in Aggregated rFVIII
The presence of known rFVIII epitopes in Agg was detected using a series of sandwich ELISAs. These ELISAs utilized an immobilized monoclonal, plate adsorbed, capture antibody against a specific epitope of rFVIII. A rat polyclonal antibody against rFVIII was used as the probe antibody. Ability of the rat polyclonal to bind to Agg was determined independently (data not shown). The sandwich ELISA format was preferred over simple direct antibody capture ELISA because the antigen species are in solution rather than bound to surface of the ELISA plate. Agg may have different binding characteristics to the plate compared to rFVIII and hence the amount of protein antigen localized on the plate can be a confounding variable in the antibody capture ELISA format. In addition, the binding of protein antigen to the ELISA plate may obscure conformational epitopes, leading to false negatives.
A series of monoclonal antibodies (Tab. 1) against known epitopes of rFVIII were used as the capture antibodies. The general conditions used for the ELISA were as follows: Nunc-Maxisorb 96-well plates were coated with the monoclonal antibody by incubating with 100 μL of an antibody solution of appropriate concentration (Tab. 1) at 4°C, overnight. Plates were subsequently blocked with 1% fish gelatin solution (blocking buffer). Hundred microliter well of dilutions of rFVIII (0.5 μg/mL) or Agg (0.5 and 5 μg/mL) in blocking buffer were incubated at 37°C for 1 h. The concentration of rFVIII chosen was confirmed to be within the linear range of the ELISA for a given set of antibodies (Tab. 1). The plates were washed and incubated with 100 μL of a 1:500 dilution of rat polyclonal antibody containing a 1:1000 dilution of goat antirat-Ig-alkaline phosphatase conjugate in blocking buffer at room temperature for 1 h. Plates were washed and incubated with 200 μL of a 1 mg/mL p-nitrophenyl phosphate solution in diethanolamine buffer (consisting of 1 M diethanolamine, and 0.5 mM MgCl2) for 30 min at room temperature for color development. Hundred microliter of 3N NaOH was added to quench the reaction. The optical density at 405 nm was determined using a plate reader. Samples were analyzed in quadruple.
Analysis of the Proteolytic Pattern of rFVIII and Agg upon Digestion with Cathepsin B
Forty-five microliters of rFVIII solution in sodium acetate buffer (40 mM sodium acetate, 1 mM EDTA and 1 mM dithiothreitol, pH 5.0) at a concentration of 100 μg/mL was incubated with 0.05 U (5 μL) of cathepsin B at 37°C. Samples were obtained at 15, 30, and 60 min. Fifteen microliter of sample containing ~1.5 μg of total protein was mixed with 15 μL of the Biorad Laemli buffer (Biorad, Hercules, CA) and heated for 5 min at 90°C. Samples (30 μL) were loaded onto 4%–15% Tris-HCl precast acrylamide gels (Biorad, Hercules, CA) and subjected to electrophoresis under a constant voltage of 120 V for 65min. The gel was stained with coomasie blue and densitometric scans were obtained using Kodak Image Station 2000MM (Kodak, Rochester, NY).
Statistical Analysis
All statistical analysis was conducted using Minitab Statistical Software, Minitab Release 14 (Minitab, Inc., State College, PA).
RESULTS
Characterization of Aggregated rFVIII
Far-UV circular dichroism spectroscopy was used to monitor any changes in the secondary structure of rFVIII upon aggregation. In its native state (20°C), the CD spectrum of the rFVIII shows a broad negative band at 215 nm (Fig. 1a), which suggests that the protein is comprised mainly of β-sheets.20 The far-UV CD spectrum of aggregated rFVIII differs significantly from that of rFVIII. The far UV-CD spectrum of aggregated rFVIII shows a positive band at 210 nm and the broad negative band at 215 nm undergoes a bathochromic shift. Large changes in rFVIII have also been seen in FTIR studies,20 which suggest that aggregated rFVIII is stabilized by intermolecular β-strands. It is appropriate to mention here that all our studies were carried out in tris buffer. Tris buffer has a high temperature coefficient, which can lead to pH shifts at elevated temperatures. Hence, the aggregation produced at elevated temperatures in tris buffer could be due to combination of temperature and pH changes. However, similar studies conducted previously in MOPS buffer, which has a lower temperature coefficient (minimal pH fluctuations) have shown that the aggregation behavior and structure of rFVIII aggregates is similar to that observed in tris buffer.19,33
Size exclusion chromatography was utilized to further characterize aggregated rFVIII. The SEC profile of native rFVIII appears as a single broad peak with a retention time between ~6.5 and 7.2 min (Fig. 1b). The SEC profile of completely aggregated rFVIII has a single peak at ~5.1–5.2 min and is excluded from the gel matrix due to its high molecular weight (Fig. 1b). The above observations indicate that the column can resolve the native and aggregated forms of rFVIII. Furthermore, since the exclusion limit of the SEC column is approximately 2000 kDa, the individual aggregated protein appears to consist of at least six or more individual rFVIII molecules. The increase in size of the molecule upon aggregation observed in SEC was also confirmed using steady state fluorescence anisotropy measurements. The increase in molecular size upon aggregation is expected to cause an increase in the anisotropy values due to the lower tumbling rates of the aggregated protein relative to the native rFVIII. As shown in Figure 1c, the anisotropy values of the aggregated protein is significantly higher compared to that of the monomeric protein (Independent t-test, p value <0.05).
Monomeric and Aggregated rFVIII were also analyzed by SDS–PAGE under reducing and nonreducing conditions to characterize the chemical nature of the aggregates. Monomeric rFVIII showed prominent bands corresponding to the heavy chain (210–90 kDa) and light chain (73 kDa). In addition to the heavy and light chain bands, the aggregated rFVIII under nonreducing conditions displayed higher molecular weight material (>250 kDa) (Fig. 1d). In order to confirm whether the higher molecular weight material was composed of disulfide linkages, SDS–PAGE was carried out under reducing conditions. The existence of high molecular weight material even under reducing conditions suggests the involvement of nonreducible bonds in the aggregation of rFVIII. The heavy and light chain bands seen in the aggregated samples are probably generated from the aggregates due to the disruption of the noncovalent bonds by the conditions of SDS–PAGE. This suggests the involvement of both noncovalent and covalent bonds in the formation of rFVIII aggregates. Additionally the involvement of strong noncovalent interactions, which are not totally disrupted under the conditions employed for the SDS–PAGE cannot be ruled out.16 In summary, the above data confirms that aggregated rFVIII differs structurally from native rFVIII and may be stabilized by covalent and noncovalent interactions.
Impact of Aggregation on the Immunogenicity of rFVIII
The impact of rFVIII aggregation on immunogenicity of rFVIII was evaluated in hemophilia A mice. The immunogenicity of rFVIII aggregates generated by thermal stress (Agg) was compared to native rFVIII. Antibodies formed against rFVIII in immunized animals were detected by ELISA and NMB assay as described in Materials and Methods. For mice immunized with native rFVIII, at the end of 6 weeks the median total antibody titer was 10937 (n = 15) while the median inhibitory antibody titer was 679 (n = 13) (Fig. 2a). For animals given Agg, however, the median total (3019, n = 12) and inhibitory (116, n = 10) antibody titers were significantly lower (Mann–Whitney, nonparametric test) than that observed for native rFVIII (Fig. 2a and b). Protein mixtures are known to have unique immune responses due to phenomenon such as antigenic competition, relative to that observed after administration of individual proteins.34,35 Hence, the immunogenicity of native rFVIII mixed with various amounts of Agg (Agg5, 5% aggregate and Agg20, 20% aggregate) was also evaluated. Antibody titers of animals immunized with Agg5 or Agg20 were comparable to animals immunized with Agg and lower than that observed for native rFVIII (Fig. 2a and b). This indicates that Agg mixed with rFVIII influences the overall immune response against monomeric rFVIII. The mechanism by which Agg influences the overall immune response against rFVIII in mixtures of Agg and rFVIII (Agg5 and Agg20) is not clear. Furthermore, no differences were observed between the antibody titers (total or inhibitory) for Agg, Agg5, or Agg20 (Kruskal–Wallis, One way nonparametric ANOVA). The above observations suggest that Agg prepared by thermal stress does not appear to enhance the immunogenicity of rFVIII.
Figure 2.

(a) Total median (bars) and individual (open symbols) antibody titers against rFVIII at 6 weeks in hemophilia-A mice immunized with rFVIII, Agg, Agg5 or Agg20. (b) Median (bars) and individual (open symbols) inhibitory titers against rFVIII at 6 weeks in hemophilia A mice immunized with rFVIII, Agg, Agg5 or Agg20. (Statistical analysis of total and inhibitory antibody titers conducted using the Kruskal–Wallis, One way nonparametric ANOVA. (c) T-cell proliferation response of hemophilic mice immunized with rFVIII or Agg. Animals immunized with rFVIII were restimulated in vitro with rFVIII (denoted as rFVIII). Animals immunized with Agg were restimulated in vitro with rFVIII (denoted as Agg-rFVIII) or Agg (denoted as Agg-Agg). Mean (bars) and individual (open symbols) stimulation indexes (d) IL-10 secretion by Th cells isolated from animals immunized with rFVIII or Agg and restimulated in vitro with relevant antigen (rFVIII, Agg-rFVIII or Agg-Agg). The open symbols represent values from individual animals and the bar represents the mean IL-10 levels secreted into the culture medium.
Detection of Th (CD4+) Cell and Corresponding Cytokine Responses
To detect the stimulation of rFVIII specific T-cells in vivo following immunization with rFVIII or Agg, in vitro T-cell proliferation studies were conducted with splenocytes isolated from immunized animals. We expected rFVIII and Agg to differ significantly in their antigenic properties. Hence, cells from animals immunized with Agg were restimulated in vitro by rFVIII and Agg independently, to detect potential Agg specific responses. rFVIII specific Th cellular responses (SI > 1.5) were detected for all animals immunized with rFVIII (Fig. 2c). Among the animals immunized with Agg, only two of the three animals showed specific Th cell responses against rFVIII (restimulation with rFVIII) as well as Agg (restimulation with Agg). The above data suggests that both rFVIII and Agg stimulate antigen specific Th responses.
We also analyzed the production of IL-10 (produced by Th2 cells) and IFN-γ (produced by Th1 cells) following in vitro restimulation. It has been shown that, IL-10 secreted by the rFVIII specific Th2 cells may be essential for production of rFVIII antibodies.36 Th cells isolated from animals immunized with rFVIII produced significant amounts of IL-10 (Fig. 2d) and negligible amounts of IFN-γ (data not shown) upon in vitro restimulation with rFVIII. Th cells from animals immunized with Agg and restimulated in vitro with rFVIII produced no IL-10 and were considered to have zero pg/mL of IL-10 (Fig. 2d). Upon restimulation in vitro with Agg, however, substantial amounts of IL-10 were detected (Fig. 2d). A negligible amount of IFN-γ was produced by the Th cells obtained from Agg immunized animals, independent of the restimulating antigen (rFVIII or Agg) (data not shown).
Detection of Known rFVIII Epitopes in Agg
The presence and availability of known specific epitopes of FVIII in aggregated rFVIII was tested using a sandwich ELISA. The location of the epitopes in the rFVIII molecule, recognized by the monoclonal antibodies utilized in the ELISA is known (Tab. 1). The ELISA was standardized to determine the linear range of response for each individual antibody and maintain consistency in the concentration of rFVIII or Agg used in the ELISA (Tab. 1). As can be seen in Figure 3a, independent of the concentration (0.5 or 5 μg/mL) used in the ELISA, Agg did not demonstrate any detectable binding to antibodies ESH4, ESH5, and ESH8. Although it is not clear whether these antibodies recognize linear or conformational epitopes, the above observations suggest that the epitopes recognized by these antibodies are probably absent or inaccessible in Agg. In contrast, antibodies 8860, N77210M, and OBT-0037 did demonstrate binding to Agg at higher concentrations, suggesting their epitopes are present and, at least, partially accessible in Agg. The sandwich ELISA was also conducted employing polyclonal human inhibitory antibodies. Plasma from two hemophilia A patients (George King Biomedical, Inc., Overland Park, KS) containing inhibitory antibodies was used as the source of the polyclonal antibodies. Patient-1 had an inhibitory titer of 301 and patient-2 an inhibitory titer of 56 BU/mL. As can be seen in Figure 3b, independent of the concentration used in the ELISA, Agg did not appear to bind to these human polyclonal antibodies. This further confirms differences in antigenic epitopes between native rFVIII and aggregated rFVIII.
Figure 3.

(a) Presence of specific epitopes recognized by ESH4 (2303–2332, C2-domain), ESH8 (2248–2285, C2-domain), ESH5 (a1, acidic region), 8860 (A2-domain), OBT-0037 (Heavy chain, A1 or A2-domain), and N77210M (A2 domain) in Agg, as determined in a sandwich ELISA. (b) Binding of rFVIII and Agg to polyclonal antibodies from hemophilia A patients as detected by the sandwich ELISA.
Proteolysis of rFVIII and Agg upon Digestion with Cathepsin B
The polypeptides formed upon incubation of rFVIII or Agg with cathepsin B were compared by SDS–PAGE as described in the Materials and Methods. The densitometric scans of SDS–PAGE gels of rFVIII and Agg (Fig. 4) show prominent bands at 214, 119, 99.0 (corresponding to the heavy chain), and 73 kDa (corresponding to the light chain). Incubation of rFVIII with cathepsin B for 60 min results in the formation of polypeptides of 76, 72, 45, and 30 kDa molecular weight. None of the peptide bands corresponding to the original intact heavy or light chain of rFVIII remain. This suggests that both the heavy and light chains are cleaved by cathepsin B. Incubation of Agg with cathepsin B for 60 min results in the formation of polypeptides of 60, 47, 41, and 29 kDa. Additionally a substantial amount of intact light chain at 73 kDa could still be detected. The polypeptides of molecular weight 76 (probably derived from heavy chain), 72, and 30 observed when monomeric rFVIII was incubated with cathepsin B were not observed upon digestion of Agg. The above data shows differences in the proteolytic digestion of rFVIII and Agg upon incubation with cathepsin B, suggesting the possibility of altered processing of Agg by endosomal proteases.
Figure 4.
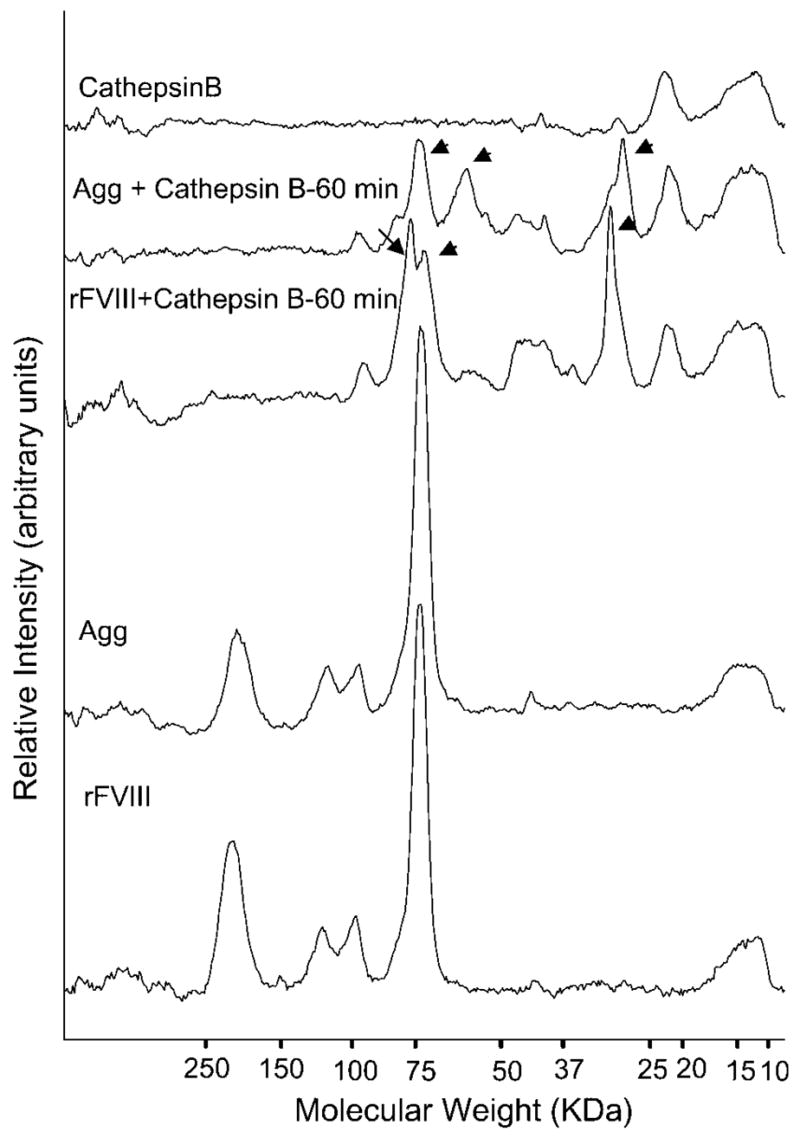
Densitometric scans of the polypeptide bands obtained in SDS–PAGE of rFVIII, Agg, rFVIII + cathepsin B, Agg + cathepsin B and cathepsin B. rFVIII and Agg were incubated with 0.05 U of cathepsin B at 37°C for 60 min. One unit will hydrolyze 1 μmole of Z-lysine p-nitrophenyl ester per min at pH 5.0 at 25°C.
DISCUSSION
Aggregates of proteins have been shown to be more immunogenic than their native counterparts.15,18 The lower immune response observed for aggregated rFVIII may be attributed to changes in the antigenic properties of rFVIII upon aggregation. We have shown that Agg differ structurally from rFVIII (Fig. 1). Differences in antigen structure can cause diversity in the mechanisms of immune response.37–39 The humoral immune response against rFVIII in hemophilia A mice and in humans has been shown to be T-cell dependant involving both Th1 and Th2 subtypes.40,41 Th2 driven responses, however, lead to higher inhibitor titers relative to Th1 induced effects.41 The in vitro studies here, conducted with splenocytes obtained from immunized animals indicates a predominantly Th2 response as suggested by the substantial IL-10 production upon restimulation. Furthermore, immunization with Agg does not lead to skewing of the Th1/Th2 balance toward Th1, since negligible amounts of IFN-γ was detected upon in vitro restimulation. It is important to note that Th cells from animals immunized with Agg were able to produce IL-10 only when restimulated with Agg, suggesting an Agg specific Th2 cellular response. This implies that the uptake and intracellular processing of Agg by antigen presenting cells may vary from that of native rFVIII and is discussed below.
Protein aggregates are generally expected to enhance antibody production because of their multivalent nature42 and presence of specific epitopes of the native protein with the highly associated polypeptide. We compared the binding of rFVIII and Agg to monoclonal and polyclonal antibodies to test for differences in reaction to specific epitopes. The results clearly indicate that there may be marked differences in the antibody epitopes between rFVIII and Agg (Fig. 3). These differences could be due to the shielding or obliteration of specific epitopes of rFVIII during aggregation. The presence or absence of individual epitopes can alter the interaction of an antigen with specific B-cell receptors essential for uptake and ultimately dictate the repertoire of antibodies produced.
Processing of endocytosed antigens by endosomal proteases into peptides is a critical step for the presentation of exogenous antigens by the MHC-II complex. The endosomal proteases cathepsins B, D, L, and S have been proposed to be involved in exogenous antigen processing.43 Involvement of specific proteases in the processing of rFVIII as an exogenous antigen has not been investigated. Nevertheless, we compared the proteolytic patterns of rFVIII and Agg generated upon digestion with cathepsin B to investigate the possibility of distinctive processing within antigen presenting cells of the two different forms of the protein. The differences in proteolytic patterns observed (Fig. 4) can again be explained on the basis of structural differences between rFVIII and Agg, which could result in altered accessibility of the enzyme to the peptide backbone for cleavage. Of particular significance is the observation that the light chain is resistant to proteolysis in Agg. This suggests that the light chain may be involved in the region of the molecule, which stabilizes the Agg thus making it inaccessible to the protease. This observation lends indirect support to our recent report that a conformational change localized in the C2 domain initiates rFVIII aggregation.19 The precise immunological consequences of the distinct proteolytic digestion patterns of rFVIII and Agg cannot be ascertained. The fact that there exist differences in proteolysis, however, implies that processing of rFVIII and Agg may also differ leading to generation of distinct immunogenic peptides, which in turn can lead to disparate immune responses.
We have identified differences in the antigenic properties of rFVIII and Agg by demonstrating, (1) Agg specific IL-10 secretion by Th cells; (2) differences in epitopes recognized by antibodies, and (3) differences in proteolysis by cathepsin B. These differences may partly explain our observations as to why Agg does not enhance the antibody development against rFVIII. The immunotoxicological consequences of the administration of Agg on the cellular immune responses are currently unknown and, therefore, require further investigation. Aggregates in general are also known to break immune tolerance to self proteins.15 Since the above immune response studies were conducted in a nontolerized (to human rFVIII) strain of mice, the ability of Agg to break immune tolerance could not be tested and will need to be evaluated. Furthermore, the above studies were conducted in a murine model to assess the relative immunogenicity of artificially produced Agg and rFVIII against a common genetic background. It is difficult to predict whether aggregates found in marketed products are structurally similar to the aggregates used in this study and whether they would produce similar responses in humans.
Antigen characteristics are critical for their uptake by the relevant antigen presenting cells or B-lymphocytes and the subsequent intracellular processing by proteases to generate peptides for MHC Class II presentation.44,45 The type of antigen will also dictate the nature of the T-helper cell response produced.46 As we have demonstrated here, aggregation of a protein molecule can induce structural changes in a protein molecule and such changes in an antigen can influence the eventual immune response.38 For antigens like ovalbumin,47,48 ProDer P 149 and influenza vaccines50 it has been demonstrated that aggregated forms of the antigen can produce a T-helper cell response distinct from that observed for the native monomeric antigen. The exact mechanism by which aggregated antigens produce a different immune response compared to the monomeric antigen is not known. We conjecture, however, based on findings of the current study and the above cited literature that differences probably arise due to the alterations in antigenic properties brought about by structural changes and epitope observation occurring upon protein association. Structural changes can lead to: (1) altered epitopes relative to the native protein essential for uptake by specific B-cells, (2) altered intracellular signaling post antigen capture essential for subsequent processing, and (3) altered susceptibility to endosomal proteases which process the antigen into peptides for subsequent association with the MHC-complex. Our observation that the aggregated protein can differ in its antigenic properties from the parent protein is significant since it calls for a more comprehensive examination of the immunotoxicological consequences of the presence of aggregated protein in protein therapeutics during the preclinical development phase.
Overall our results suggest that aggregated rFVIII differs significantly in its antigenic properties relative to native rFVIII and in contrast to what might be expected, does not enhance the immunogenicity of rFVIII in hemophilia A mice.
Table 1.
Experimental Conditions for Determining Presence of Specific Epitopes in Agg
| Stationary Antibody | Epitope of Stationary Antibody | Stationary Antibody Coating Concentration | Antigen Concentration | Probe Antibody | Probe Antibody Dilution | Linear Range |
|---|---|---|---|---|---|---|
| ESH4 | C2 domain (2303–2332) | 5.00 μg/mL | rFVIII = 0.5 μg/mL
Agg = 0.5 and 5 μg/mL |
Rat polyclonal | 1:500 | 62.5–750.0 ng/mL |
| ESH8 | C2 domain (2248–2285) | 5.00 μg/mL | rFVIII = 0.5 μg/mL
Agg = 0.5 and 5 μg/mL |
Rat polyclonal | 1:500 | 62.5–750.0 ng/mL |
| 8860 | A2 domain | 2.80 μg/mL | rFVIII = 0.5 μg/mL
Agg = 0.5 and 5 μg/mL |
Rat polyclonal | 1:500 | 62.5–750.0 ng/mL |
| N77210M | A2 domain | 12.5 μg/mL | rFVIII = 0.5 μg/mL
Agg = 0.5 and 5 μg/mL |
Rat polyclonal | 1:500 | 62.5–750.0 ng/mL |
| ESH5 | Acidic region | 5.00 μg/mL | rFVIII = 0.5 μg/mL
Agg = 0.5 and 5 μg/mL |
Rat polyclonal | 1:500 | 62.5–1000 ng/mL |
| OBT-0037 | A1 or A2 domain | 5.00 μg/mL | rFVIII = 0.5 μg/mL
Agg = 0.5 and 5 μg/mL |
Rat polyclonal | 1:500 | 62.5–1000 ng/mL |
| Patient 1 | Polyclonal | 1:200 dilution | rFVIII = 2.0 μg/mL
Agg = 2.0 and 20 mg/mL |
Rat polyclonal | 1:500 | 0.5–20 μg/mL |
| Patient 2 | Polyclonal | 1:100 dilution | rFVIII = 2.0 μg/mL
Agg = 2.0 and 20 μg/mL |
Rat polyclonal | 1:500 | 0.5–10 μg/mL |
Acknowledgments
The authors thank the Pharmaceutical Sciences Instrumentation Facility, University at Buffalo, for the use of Circular Dichroism spectropolarimeter and spectrofluorometer. This work was supported by NHLBI, National Institute of Health grant R01 HL-70227 to SVB.
Abbreviations used
- ACD
acid citrate dextrose
- aPTT
activated partial thromboplastin time
- ELISA
enzyme-linked immunosorbent assay
- FVIII
factor VIII
- IgG
immunoglobulin G
- i.v
intravenous
- NMB
Nijmegen modification of the Bethesda assay
- rFVIII
recombinant human Factor VIII
- s.c
subcutaneous
- SEC
size exclusion chromatography
References
- 1.Toole JJ, Knopf JL, Wozney JM, Sultzman LA, Buecker JL, Pittman DD, Kaufman RJ, Brown E, Shoemaker C, Orr EC, Amphlett GW, Foster BW, Coe ML, Knutson GJ, Fass DN, Hewick RN. Molecular cloning of a cDNA encoding human antihaemophilic factor. Nature. 1984;312:342–347. doi: 10.1038/312342a0. [DOI] [PubMed] [Google Scholar]
- 2.Vehar GA, Keyt B, Eaton D, Rodriguez H, O’Brien DP, Rotblat F, Oppermann H, Keck R, Wood WI, Harkins RN, Tuddenham EGD, Lawn RD, Capon DJ. Structure of human factor VIII. Nature. 1984;312:337–342. doi: 10.1038/312337a0. [DOI] [PubMed] [Google Scholar]
- 3.Kaufman RJ, Wasley LC, Dorner AJ. Synthesis, processing, and secretion of recombinant human factor VIII expressed in mammalian cells. J Biol Chem. 1988;263:6352–6362. [PubMed] [Google Scholar]
- 4.Foster PA, Zimmerman TS. Factor VIII structure and function. Blood Reviews. 1989;3:180–191. doi: 10.1016/0268-960x(89)90015-5. [DOI] [PubMed] [Google Scholar]
- 5.Fay PJ. Reconstitution of human factor VIII from isolated subunits. Arch Biochem Biophys. 1988;262:525–531. doi: 10.1016/0003-9861(88)90404-3. [DOI] [PubMed] [Google Scholar]
- 6.VanAken WG. The potential impact of recombinant factor VIII on hemophilia care and the demand for blood and blood products. Transfus Med Rev. 1997;11:6–14. doi: 10.1016/s0887-7963(97)80005-3. [DOI] [PubMed] [Google Scholar]
- 7.Jacquemin MG, Saint-Remy JM. Factor VIII immunogenicity. Haemophilia. 1998;4:552–557. doi: 10.1046/j.1365-2516.1998.440552.x. [DOI] [PubMed] [Google Scholar]
- 8.Lollar P, Healey JF, Barrow RT, Parker ET. Factor VIII inhibitors. Adv Exp Med Biol. 2001;489:65–73. doi: 10.1007/978-1-4615-1277-6_6. [DOI] [PubMed] [Google Scholar]
- 9.Saint-Remy JM, Lacroix-Desmazes S, Oldenburg J. Inhibitors in haemophilia: pathophysiology. Haemophilia. 2004;10:146–151. doi: 10.1111/j.1365-2516.2004.01009.x. [DOI] [PubMed] [Google Scholar]
- 10.Oldenburg J, Schroder J, Hermann Brackmann H, Muller-Reible C, Schwaab R, Tuddenham E. Environmental and genetic factors influencing inhibitor development. Semin Hematol. 2004;41:82–88. doi: 10.1053/j.seminhematol.2003.11.016. [DOI] [PubMed] [Google Scholar]
- 11.Wills RJ, Ferraiolo BL. The role of pharma-cokinetics in the development of biotechnologically derived agents. Clin Pharmacokinet. 1992;23:406–414. doi: 10.2165/00003088-199223060-00002. [DOI] [PubMed] [Google Scholar]
- 12.Schellekens H. Immunogenicity of therapeutic proteins: clinical implications and future prospects. Clin Ther. 2002;24:1720–1740. doi: 10.1016/s0149-2918(02)80075-3. discussion 1719. [DOI] [PubMed] [Google Scholar]
- 13.Schellekens H. Bioequivalence and the immunogenicity of biopharmaceuticals. Nat Rev Drug Discov. 2002;1:457–462. doi: 10.1038/nrd818. [DOI] [PubMed] [Google Scholar]
- 14.Ryff JC, Schellekens H. Immunogenicity of rDNA-derived pharmaceuticals. Trends Pharmacol Sci. 2002;23:254–256. doi: 10.1016/s0165-6147(02)02024-2. [DOI] [PubMed] [Google Scholar]
- 15.Braun A, Kwee L, Labow MA, Alsenz J. Protein aggregates seem to play a key role among the parameters influencing the antigenicity of interferon alpha (IFN-alpha) in normal and transgenic mice. Pharm Res. 1997;14:1472–1478. doi: 10.1023/a:1012193326789. [DOI] [PubMed] [Google Scholar]
- 16.Wang W, Kelner DN. Correlation of rFVIII inactivation with aggregation in solution. Pharm Res. 2003;20:693–700. doi: 10.1023/a:1023271405005. [DOI] [PubMed] [Google Scholar]
- 17.Patten PA, Schellekens H. The immunogenicity of biopharmaceuticals. Lessons learned and consequences for protein drug development. Dev Biol (Basel) 2003;112:81–97. [PubMed] [Google Scholar]
- 18.Moore WV, Leppert P. Role of aggregated human growth hormone (hGH) in development of antibodies to hGH. J Clin Endocrinol Metab. 1980;51:691–697. doi: 10.1210/jcem-51-4-691. [DOI] [PubMed] [Google Scholar]
- 19.Ramani K, Purohit VS, Miclea RD, Middaugh CR, Balasubramanian SV. Lipid binding region (2303–2332) is involved in aggregation of recombinant human FVIII (rFVIII) J Pharm Sci. 2005;94:1288–1299. doi: 10.1002/jps.20340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Grillo AO, Edwards KL, Kashi RS, Shipley KM, Hu L, Besman MJ, Middaugh CR. Conformational origin of the aggregation of recombinant human factor VIII. Biochemistry. 2001;40:586–595. doi: 10.1021/bi001547t. [DOI] [PubMed] [Google Scholar]
- 21.Cho Y, Garanchon C, Kashi R, Wong C, Besman MJ. Characterization of aggregates of recombinant human factor VIII by size-exclusion chromatography and immunoassay. Biotechnol Appl Biochem. 1996;24:55–59. [PubMed] [Google Scholar]
- 22.Lakowicz JR. Principles of fluorescence spectroscopy. 2. New York: Kluwer Academic/Plenum Publishers; 1999. [Google Scholar]
- 23.Bi L, Lawler AM, Antonarakis SE, High KA, Gearhart JD, Kazazian HH., Jr Targeted disruption of the mouse factor VIII gene produces a model of haemophilia A. Nat Genet. 1995;10:119–121. doi: 10.1038/ng0595-119. [DOI] [PubMed] [Google Scholar]
- 24.Qian J, Borovok M, Bi L, Kazazian HH, Jr, Hoyer LW. Inhibitor antibody development and T cell response to human factor VIII in murine hemophilia A. Thromb Haemost. 1999;81:240–244. [PubMed] [Google Scholar]
- 25.Reipert BM, Ahmad RU, Turecek PL, Schwarz HP. Characterization of antibodies induced by human factor VIII in a murine knockout model of hemophilia A. Thromb Haemost. 2000;84:826–832. [PubMed] [Google Scholar]
- 26.Behrmann M, Pasi J, Saint-Remy JM, Kotitschke R, Kloft M. Von Willebrand factor modulates factor VIII immunogenicity: comparative study of different factor VIII concentrates in a haemophilia A mouse model. Thromb Haemost. 2002;88:221–229. [PubMed] [Google Scholar]
- 27.Pratt KP, Qian J, Ellaban E, Okita DK, Diethelm-Okita BM, Conti-Fine B, Scott DW. Immunodominant T-cell epitopes in the factor VIII C2 domain are located within an inhibitory antibody binding site. Thromb Haemost. 2004;92:522–528. doi: 10.1160/TH03-12-0755. [DOI] [PubMed] [Google Scholar]
- 28.http://www.baxter.com/products/biopharmaceuticals/downloads/Advate_PI.pdf.
- 29.Remmele RL, Jr, Gombotz WR. Differential scanning calorimetry: a practical tool for elucidating stability of liquid pharmaceuticals. BioPharm. 2000;13:36–46. [Google Scholar]
- 30.Tsai PK, Volkin DB, Dabora JM, Thompson KC, Bruner MW, Gress JO, Matuszewska B, Keogan M, Bondi JV, Middaugh CR. Formulation design of acidic fibroblast growth factor. Pharm Res. 1993;10:649–659. doi: 10.1023/a:1018939228201. [DOI] [PubMed] [Google Scholar]
- 31.Purohit VS, Ramani K, Sarkar R, Kazazian HH, Jr, Balasubramanian SV. Lower inhibitor development in hemophilia A mice following administration of recombinant factor VIII-O-phospho-L-serine complex. J Biol Chem. 2005;280:17593–17600. doi: 10.1074/jbc.M500163200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Verbruggen B, Novakova I, Wessels H, Boezeman J, van den Berg M, Mauser-Bunschoten E. The Nijmegen modification of the Bethesda assay for factor VIII:C inhibitors: improved specificity and reliability. Thromb Haemost. 1995;73:247–251. [PubMed] [Google Scholar]
- 33.Ramani K, Purohit V, Middaugh CR, Balasubramanian SV. Aggregation kinetics of recombinant human FVIII (rFVIII) J Pharm Sci. 2005;94:2023–2029. doi: 10.1002/jps.20432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Reipert BM, Schoppmann A, Schwarz HP. A caution on the use of murine hemophilia models for comparative immunogenicity studies of FVIII products with different protein compositions. Thromb Haemost. 2003;89:1110–1112. author reply 1113. [PubMed] [Google Scholar]
- 35.Hunt JD, Jackson DC, Wood PR, Stewart DJ, Brown LE. Immunological parameters associated with antigenic competition in a multivalent footrot vaccine. Vaccine. 1995;13:1649–1657. doi: 10.1016/0264-410x(95)00145-q. [DOI] [PubMed] [Google Scholar]
- 36.Wu H, Reding M, Qian J, Okita DK, Parker E, Lollar P, Hoyer LW, Conti-Fine BM. Mechanism of the immune response to human factor VIII in murine hemophilia A. Thromb Haemost. 2001;85:125–133. [PubMed] [Google Scholar]
- 37.Thai R, Moine G, Desmadril M, Servent D, Tarride JL, Menez A, Leonetti M. Antigen stability controls antigen presentation. J Biol Chem. 2004;279:50257–50266. doi: 10.1074/jbc.M405738200. [DOI] [PubMed] [Google Scholar]
- 38.Khalili-Shirazi A, Quaratino S, Londei M, Summers L, Tayebi M, Clarke AR, Hawke SH, Jackson GS, Collinge J. Protein conformation significantly influences immune responses to prion protein. J Immunol. 2005;174:3256–3263. doi: 10.4049/jimmunol.174.6.3256. [DOI] [PubMed] [Google Scholar]
- 39.Grundner C, Pancera M, Kang JM, Koch M, Sodroski J, Wyatt R. Factors limiting the immunogenicity of HIV-1 gp120 envelope glycoproteins. Virology. 2004;330:233–248. doi: 10.1016/j.virol.2004.08.037. [DOI] [PubMed] [Google Scholar]
- 40.Sasgary M, Ahmad RU, Schwarz HP, Turecek PL, Reipert BM. Single cell analysis of factor VIII-specific T cells in hemophilic mice after treatment with human factor VIII. Thromb Haemost. 2002;87:266–272. [PubMed] [Google Scholar]
- 41.Reding MT, Lei S, Lei H, Green D, Gill J, Conti-Fine BM. Distribution of Th1- and Th2-induced anti-factor VIII IgG subclasses in congenital and acquired hemophilia patients. Thromb Haemost. 2002;88:568–575. [PubMed] [Google Scholar]
- 42.Charles JA, Travers P, Walport M, Capra JD. Immunobiology: the immune system in health and disease. 4. New York: Elsevier Science Ltd/Garland Publishing; 1999. The induction and detection of immune responses; pp. 36–37. [Google Scholar]
- 43.Katunuma N, Matsunaga Y, Himeno K, Hayashi Y. Insights into the roles of cathepsins in antigen processing and presentation revealed by specific inhibitors. Biol Chem. 2003;384:883–890. doi: 10.1515/BC.2003.099. [DOI] [PubMed] [Google Scholar]
- 44.Watts C. Capture and processing of exogenous antigens for presentation on MHC molecules. Annu Rev Immunol. 1997;15:821–850. doi: 10.1146/annurev.immunol.15.1.821. [DOI] [PubMed] [Google Scholar]
- 45.Parker DC. T cell-dependent B cell activation. Annu Rev Immunol. 1993;11:331–360. doi: 10.1146/annurev.iy.11.040193.001555. [DOI] [PubMed] [Google Scholar]
- 46.Constant SL, Bottomly K. Induction of Th1 and Th2 CD4+ T cell responses: the alternative approaches. Annu Rev Immunol. 1997;15:297–322. doi: 10.1146/annurev.immunol.15.1.297. [DOI] [PubMed] [Google Scholar]
- 47.Speidel K, Osen W, Faath S, Hilgert I, Obst R, Braspenning J, Momburg F, Hammerling GJ, Rammensee HG. Priming of cytotoxic T lymphocytes by five heat-aggregated antigens in vivo: conditions, efficiency, and relation to antibody responses. Eur J Immunol. 1997;27:2391–2399. doi: 10.1002/eji.1830270938. [DOI] [PubMed] [Google Scholar]
- 48.Koch C, Jensen SS, Oster A, Houen G. A comparison of the immunogenicity of the native and denatured forms of a protein. APMIS. 1996;104:115–125. doi: 10.1111/j.1699-0463.1996.tb00696.x. [DOI] [PubMed] [Google Scholar]
- 49.Magi M, Garcia L, Vandenbranden M, Palmantier R, Jacquet A. Heat denaturation affects the ProDer p 1 IgE reactivity and downregulates the development of the specific allergic response. J Allergy Clin Immunol. 2004;114:545–552. doi: 10.1016/j.jaci.2004.04.014. [DOI] [PubMed] [Google Scholar]
- 50.Babiuk S, Skowronski DM, De Serres G, HayGlass K, Brunham RC, Babiuk L. Aggregate content influences the Th1/Th2 immune response to influenza vaccine: evidence from a mouse model. J Med Virol. 2004;72:138–142. doi: 10.1002/jmv.10540. [DOI] [PubMed] [Google Scholar]