

Abstract
Availability of human embryonic stem cells (hESC) has enhanced human neural differentiation research. The derivation of neural progenitor (NP) cells from hESC facilitates the integration of human embryonic development through the generation of neuronal subtypes and supporting glial cells. These cells will likely lead to new and novel drug screening and cell therapy uses. This review will discuss the current status of derivation, maintenance and further differentiation of NP cells with special emphasis on the cellular signaling involved in these processes. The derivation process affects the yield and homogeneity of the NP cells. Then when exposed to the correct environmental signaling cues, NP cells can follow a unique and robust temporal cell differentiation process forming numerous phenotypes.
Keywords: Human embryonic stem cells, neural differentiation, cell signaling, regional specification
INTRODUCTION
Human embryonic stem cells (hESC) are pluripotent self-renewable cells derived from early blastocyst embryos [Thomson et al., 1998]. Being pluripotent, hESC can generate neural cells including functional neurons, glial cells and oligodendrocytes under appropriate culture conditions [Carpenter et al., 2001; Hong et al., 2008; Keirstead et al., 2005; Shin et al., 2006] and has been the topic of recent reviews [Cai and Grabel, 2007; Robertson et al., 2008; Wilson and Stice, 2006; Zhang, 2004]. While differentiation is directed towards the neural lineage, lack of an optimal protocol implies generation of other lineage cells as contaminants. Thus, one major challenge in the field is to generate a homogeneous and renewable, easy to culture, neural progenitor (NP) cell population committed to the neural lineage, capable of serving as an unlimited lineage-restricted cell source for replacement therapy and/or for other studies. Producing NP cell populations from hESC, specific to different regions of nervous system still remains elusive. Therefore, it is critical to optimally differentiate hESC to NP cells, their maintenance as a self-renewing population, and further to generate highly pure population of regionally specified cell types in culture. Cell signaling plays an important role in each process. Recent advances covering these issues of hESC biology for generation of NP cells are the focus of this review.
PRINCIPLE OF NEURAL INDUCTION IN hESC
Maintenance of pluripotent state allows hESC to multiply continually in culture while preventing differentiation to other lineage cells. Any disruption of the pluripotency supporting system (for example, by withdrawing key factor(s) from the medium or forcing the hESC to grow in suspension etc) promotes spontaneous differentiation to several lineages. Due to the stochastic nature of spontaneously differentiating hESC, many other cell types are also generated; therefore, spontaneous differentiation is not an efficient method for generating neural cells. Thus, it is imperative to make use of certain factors that direct hESC differentiation specifically to neural lineage. Addition of instructive factors and removal of preventive factors form the basis of directed differentiation resulting in an improved NP cell yield. Once the hESC differentiate into NP cells, the newly formed NP cells maintain expression of SOX2 and begin expressing other neuroepithelial markers, such as Nestin, SOX1, SOX3, PSA-NCAM, and MUSASHI-1 (Figure 1). Formation of “neural rosettes” is another morphologic marker of hESC differentiation to neural cells. Here, neuroectodermal columnar cells also express markers found in neuroepithelial cells of neural plate and they arrange radially to form a rosette like structure reminiscent of the in vivo structural formation of developing neural tube [O'Rahilly and Muller, 1994]. Cells in neural rosettes are capable of multiplying by symmetrical division over extended period in culture. These features provide researchers the opportunity to manipulate and expand the derived cells.
Figure 1. Directed differentiation of human embryonic stem cells (hESC) by adherent or suspension (embryoid body, EB) culture models.
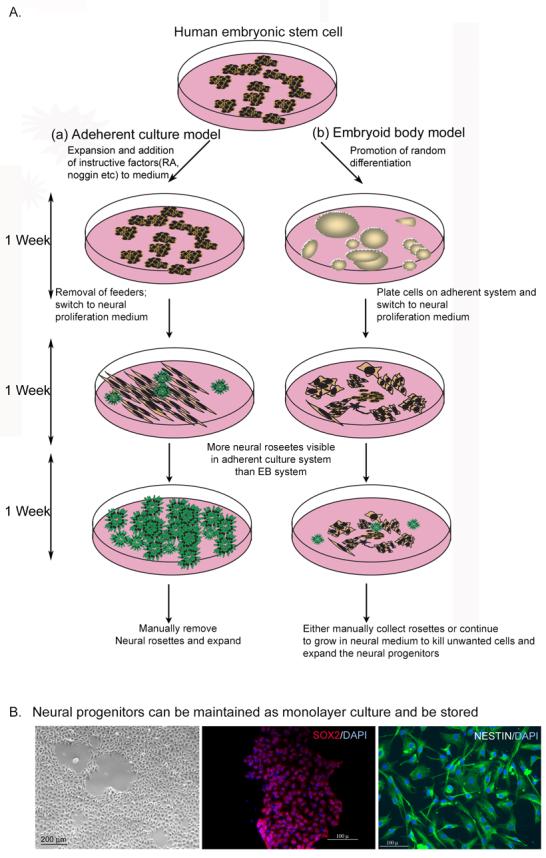
A. In each model cells are differentiated in three steps for a varying time period ranging between 18-21 days.
Panel (a): hESC differentiation on adherent culture. This is done with/or without feeder cells. Usually, factors that promote neural differentiation are added to the medium. Commonly, retinoic acid (RA), BMP-inhibitors (such as, noggin) and other supplements (like N2, B27, ITS) are used in the culture. In a feeder-free system, hESC are plated on a variety of matrices such as laminin, fibronectin, collagen or Matrigel. If feeder cells are present in the culture, they are removed at around 10 days of differentiation. By this time, cells are committed to neural fates showing neural rosettes in culture. In the next step, cells are cultured with neural permissive medium and neural rosettes are passaged with the same medium that contains high amount of basic FGF (FGF2) and sometimes, EGF.
Panel (b): hESc differentiation in suspension culture. cells are grown in suspension to allow EB formation with or without FGF2 for a period varying between 1-2 weeks. During this time, cells randomly differentiate and the process stochastically generates many cell types. During EB formation, neural induction factors are also used. After this, differentiated cells are plated on adherent culture to selectively permit the growth of neural cells, and neural rosettes are manually passaged in neural permissive medium.
B. Neural progenitors generated by either of these methods can be maintained in culture for an unlimited time in the presence of high FGF2 and EGF. In culture they maintain characteristic morphology (bright field image, left panel) and show expression of NP-markers such as SOX2 (red in middle panel) and Nestin (in green, right). DAPI staining in blue indicates nuclei. These images are taken with neural progenitor line EnStem-A (Millipore) derived from WA09 hESC and maintained in neurobasal medium supplemented with FGF2 and B27.
DIRECTED NEURAL DIFFERENTIATION OF hESC
a) Embryoid body (EB) model versus adherent culture
As was pointed out before, spontaneous differentiation is inefficient, making differentiation under conditions favoring pro-neural selection is imperative. This calls for a culture system that will allow differentiation of hESC, and support selective growth and proliferation of neural cells. Conventionally, two types of culture systems are employed for this purpose: adherent and suspension culture. In suspension culture hESC are allowed to grow as a mass of cells, “embryoid bodies” for different lengths of time. With longer duration of differentiation, an embryoid body (EB) is capable of forming a multilayered structure that contains a mixed population of cells including neural cells. In a prolonged culture, these EBs show formation of cavities filled with cystic fluid [Schuldiner et al., 2001]. To increase pro-neural differentiation and improve survival of desired cell types, growth factors or morphogens have been commonly used in culture. However there are several disadvantages associated with EB culture: 1) variability of the EB size (due to different initial cell numbers or duration of differentiation) which affects the yield of NP cells, 2) higher concentration of morphogens in culture required to enable the morphogens to reach the innermost layers of the EBs forming a concentration gradient that leads to the generation of differentiated cells at different developmental stages belonging to tissues of different germ layer origin, 3) in addition being an aggregate of many cells EBs also present difficulties in monitoring cell morphology during differentiation using microscopy. These issues can be overcome when hESC are differentiated in an adherent culture system wherein cells are differentiated as a monolayer culture and exposed uniformly to morphogens [Shin et al., 2006] (Figure 1).
b) Role of media and factors in neural differentiation
Different factors or signaling molecules, supplements/additives and conditioned media have been used to differentiate hESC into NP cells [Carpenter et al., 2001; Dhara et al., 2008; Reubinoff et al., 2001; Shin et al., 2006; Zhang et al., 2001]. Selection of these factors in the context of hESC differentiation is based in part on mouse ES cell neural differentiation. Specific roles for many of the components (like media additives) in promoting differentiation are unknown. Based on information available, we and others have derived NP cells using few-stepwise procedures [Carpenter et al., 2001; Dhara et al., 2008; Reubinoff et al., 2001; Shin et al., 2006; Zhang et al., 2001]. In these steps, FGF2, RA and media supplements like N2 and B27 were used. The following is a brief discussion of the possible roles of different factors/signaling related to neural differentiation.
Stromal-derived inducing activity (SDIA)
Stromal cells are loose connective tissue cells found in number of organs, such as gonads and bone marrow. They provide matrix-support for other cells in the organ to function. To promote neural differentiation, hESC have been co-cultured in the presence of stromal cells [Perrier et al., 2004; Vazin et al., 2008]. These support cells successfully generated terminally differentiated neuronal cells, like dopaminergic neurons in culture [Perrier et al., 2004]. Here, in vitro neural differentiation occurs because of the existence of SDIA produced by the stromal cells. This concept was based on the fact that mesodermal signaling is required for neural induction as demonstrated in both mouse and primate ES cell differentiation [Kawasaki et al., 2000; Kawasaki et al., 2002]. Numerous stromal cell lines (like PA6, MS5, S2) were used for this purpose in different laboratories [Mizuseki et al., 2003; Perrier et al., 2004]. In a recent SDIA study it was shown that cell surface activity of rodent stromal cells line PA6 is required for neural differentiation of hESC [Vazin et al., 2008]. SDIA-mediated differentiation provides an efficient way for neural tissue generation, but this co-culture technique can introduce unknown stromal factors of non-human origin in culture and may obscure the exact mechanisms involved during neural differentiation.
Retinoic acid (RA) signaling
Retinoic acid plays important roles in many aspects of neural development and activity. These include axon regeneration in the adult, neuronal differentiation and patterning of the neural plate and neural tube in the early embryos [Maden, 2007]. In a developing embryo, higher concentrations of RA are found in the posterior hindbrain and spinal cord and the concentration decreases towards the anterior region. This pattern forms an increasing gradient of RA in antero-posterior axis in the neural plate. Additionally, in the absence of RA signaling, the posterior hindbrain is not formed [Maden, 2007] and development of the anterior spinal cord is affected. Thus, RA is implicated in specifically organizing the posterior hindbrain and anterior spinal cord. Similarly, in the dorso-ventral axis of the neural tube RA works synergistically with other molecules such as sonic hedgehog (SHH), FGF and bone morphogenetic protein (BMP), to determine the fates of sensory neurons, inter neurons and motor neurons. RA-mediated neuronal and glial differentiation occurs by activation of different genes that include transcription factors (BRN2, NFkB, SOX1, SOX6 etc), cell signaling molecules or associated cell structures (ceramide, PSEN1, MAP2 etc), and extracellular molecules (WNT signaling members). Activation of these diverse molecules indicates that complex molecular pathways regulate neuronal differentiation. Adopting the protocol from mouse ES cells to hESC neural differentiation, RA was used by every research group with a concentration between 1-10 μM in the medium [Baharvand et al., 2007; Carpenter et al., 2001; Erceg et al., 2008; Levenberg et al., 2003; Park et al., 2004; Reubinoff et al., 2001; Schuldiner et al., 2001; Zhang et al., 2001]. As RA is a morphogen, varying concentration may have been responsible for generating NP cell populations with different differentiation potentials obtained by these groups.
Bone Morphogenetic Protein (BMP) signaling
Bone morphogenetic proteins are members of TGFβ (transforming growth factor) superfamily and they play multiple roles in the development of central and peripheral nervous system. In vertebrates, loss of BMP signaling is implicated in neural induction from ectoderm at or before gastrulation [Harland, 2000]. After neural tube closure, BMP signaling from dorsal epidermal ectoderm generates neural crest cells and dorsal inter neurons but limits ventral interneuron differentiation. Expression of BMP-antagonists like noggin, chordin, follistatin secreted from notochord and floorplate attenuate the effect BMP and are suggested to be involved in dorso-ventral patterning of the spinal cord [Graham et al., 1994; Shimamura and Rubenstein, 1997]. Also, over-expression of BMPs by bead implantation resulted in loss of ventral forebrain markers. However, this over-expression maintained the expression of dorsal forebrain markers [Furuta et al., 1997]. With the aid of SHH signaling, BMPs generates ventral regions of the forebrain, whereas, SHH alone specifies ventral regions of the hindbrain [Dale et al., 1997]. Later in development, BMP signaling promotes generation of neuronal and astro-glial cells but prevents oligo-glial differentiation. Thus, BMPs mainly specify forebrain cells and inhibiting BMPs before or during gastrulation causes neural induction.
To induce neural differentiation, BMP-antagonist noggin was used in hESC culture grown adherently on laminin or matrigel [Baharvand et al., 2007; Gerrard et al., 2005; Pera et al., 2004] or grown in suspension as EBs [Itsykson et al., 2005]. Treatment with noggin up-regulated the expression of neural transcription factors Pax6 and Sox2 and other neural marker such as nestin [Pera et al., 2004]. hESC treated with noggin, that potentially binds to BMP-2,4 and 7, generated neural progenitors and also prevented formation of extra-embryonic endoderm (AFP, GATA6 down-regulation). Noggin has also been shown to up-regulate the expression of phosphorylated Smad1[Pera et al., 2004] and down regulate the BMP-2 target proteins Id2 and Id3. Thus, noggin is likely to directly increase neurogenesis using Smad1-mediated signaling. It can also acts indirectly preventing BMP target proteins (Id2, Id3) that repress the promoter activation induced by neurogenic bHLH factors [Miyazono and Miyazawa, 2002]. However, another BMP-inhibitor follistatin, that has higher binding affinity for activin, did not generate neuroectoderm, though it prevented formation of extra-embryonic-ectoderm cells and differentiated cells had fibroblast like morphology [Gerrard et al., 2005].
Fibroblast Growth Factor (FGF) signaling
Members of the FGF family act through FGF receptors (FGFRs) that activate cascades of kinase signaling including Raf, MEK (mitogen-activated and extracellular signal-regulated kinase), and MAPK (mitogen activated protein kinase) [Nutt et al., 2001]. FGF signaling works in association with other pathways such as TGFβ, WNT, Hedgehog, and Notch [Gerhart, 1999]. FGF signaling have been implicated in the induction and patterning of mesoderm and neural tissues in vertebrate embryos (for a review, see [Doniach, 1995]; [Streit et al., 2000]).
In neural specification during early embryonic development, FGF molecules might play two different roles: 1) they may induce a “pro” neural state at an early stage, and 2) while acting as antagonist to BMP signaling, they may stabilize neural identity [Zhang, 2004]. In view of information on patterning cues, FGF signaling may enrich survival and proliferation of the early forebrain neural precursor cell types. Based on this, basic FGF (or FGF2) was used to selectively promote the proliferation of neural population [Okabe et al., 1996]. We and others routinely use FGF2 to isolate and maintain NP cells [Carpenter et al., 2001; Dhara et al., 2008; Elkabetz et al., 2008; Shin et al., 2006]. See [Mason, 2007] for more on the role of FGF signaling in neural development.
Media supplements and conditioned-media
Often NP cells are derived and maintained in Neurobasal medium (from Gibco) which is a modified form of Dulbecco's Modified Eagle's minimal essential medium- Ham's F12 (DMEM/F12), in which the osmolality and the concentration of several amino acids have been reduced and ferrous sulfate eliminated [Price and Brewer, 2001]. For culturing NP cells, B27 (Gibco) is added to this medium. B27 contains RA (all trans-retinol and retinyl acetate, 0.1 mg/lit each) and this supplement when used as suggested in the published protocol [Shin et al., 2006] yields a NP cell proliferation medium containing over 12 μM RA. This concentration may affect neural specification since 1-10 μM RA was used for hESC differentiation to produce the caudal-specific cells. Also, we showed that B27-supplemented medium-derived NP cells generated motoneurons as characterized by expression of HB9 (a motor neuron specific marker), choline acetyltransferase (ChAT, motoneuronphenotype marker) and TuJ1 (neuronal beta III tubulin) in induction medium with FGF2, Sonic hedgehog and additional RA [Shin et al., 2005].
The other popularly used medium-supplement added to Neurobasal or DMEM/F12 media is N2 (Gibco). N2 has a subset of B27 components that include insulin which may act through insulin-like growth factor receptors to promote differentiation of hESC and proliferation of differentiated cells. We found that NP cell proliferation and survival is superior in B27-supplemented medium than in medium with N2. To obtain improved survival with N2 supplemented medium, higher density seeding was required. N2 supplements are chemically defined, and therefore, it is possible to study the effect of unknown factors without the confounding effects found in serum or serum components, such as serum albumin [Price and Brewer, 2001]. In addition to the factors discussed above, HepG2-conditioned MedII medium was also used to generate neural cells from hESC in adherent [Shin et al., 2006] or suspension [Schulz et al., 2003] culture. However, the mechanism of such differentiation is not clear.
c) Temporal events during neural differentiation
The developing nervous system is controlled by complex signaling events, in part, neuroepithelial cells are specified in the ectoderm under specific FGF and WNT signaling, which also prevents mesoderm differentiation. This neuroectoderm forms the neural plate which folds to generate the neural tube. In the neural tube, rostral or forebrain features are acquired first after which gradually caudalization occurs. In the rostro-caudal patterning process, WNT, BMP, FGF and RA play major roles. This patterning results in the formation of the major segments of central nervous system: forebrain, midbrain, hindbrain and spinal cord. Once rostro-caudal patterning is over, features of dorso-ventral patterning appear in the neural tube, controlled by two opposing signals, dorsally BMP from the roof plate and ventrally SHH from the notochord (Figure 2). Cells located in the unique positions along rostro-caudal and dorso-ventral axial grids respond to specific morphogens and generate specific subtypes of neurons and glial cells [Zhang, 2006].
Figure 2. Embryonic neural development and major signaling events in mammals.
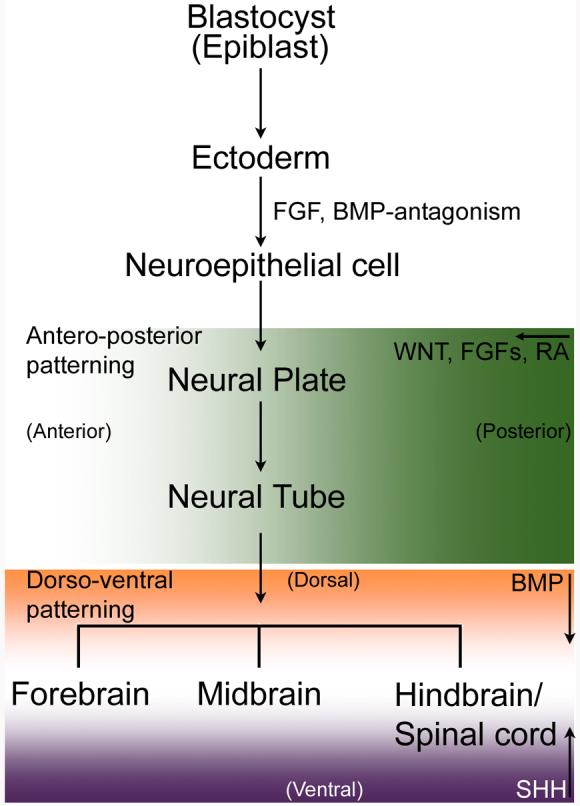
In early blastocyst, of three germ layers (ectoderm, endoderm and mesoderm), ectoderm gives rise to neural tissues. Initial mesodermal signals induce ectodermal cells to become neuroepithelial cells that rapidly multiply to form neural plate.
Neural induction is controlled by an instructive signal from fibroblast growth factor (FGF). FGF can also inhibit BMP (bone morphogenetic protein) that in the presence of WNT molecules converts ectodermal cells into epidermis. In this induction step, some roles of NOTCH signaling are also known [Lowell S et al. (2006) Notch promotes neural lineage entry by pluripotent embryonic stem cells, PLoS Biol. 4: e121 and for a review, Louvi, A and Artavanis-Tsakonas, S. (2006) Notch signalling in vertebrate neural development, Nat Rev Neurosci 7:93-102].
The neural tube forms from the neural plate. WNT, FGFs and Retinoic acid (RA) (forming a gradient shown here in green shade) impart hindbrain character to the neural tube. Signaling molecules (such as Nodal-related proteins that inhibit both WNT and BMP members, and WNT-inhibitors like Cerbeus and Dickkopf) secreted from extra-embryonic tissue prevent the caudalization of the neural tube and thereby promote forebrain formation. FGF8 plays an important role in determining midbrain identity in the neural tube. These signaling molecules (WNT, FGF, RA) are involved in antero-posterior development. Dorso-ventral patterning of the neural tube occurs under two main opposing signals. Dorsal features are determined by BMP and sonic hedgehog (SHH) ventralizes the brain and spinal cord. Both these signals form gradients (BMP in orange and SHH in purple) in opposite direction. In the hindbrain and spinal cord RA generates inter-neurons.
During in vivo development, the neural plate forms and then eventually the neural tube when the human embryo is approximately 3 weeks old [O'Rahilly and Muller, 1994]. hESC are cells derived from inner cell mass of 5-6 days old blastocysts and propagated in tissue culture. In vitro differentiation of hESC to neural lineage is characterized by formation of neural rosette, reminiscent of transverse-section of a neural tube. Even though neural rosettes start appearing approximately by day 10 of hESC differentiation, the majority are observed at about day 18-21 of differentiation [Dhara et al., 2008]. Therefore, it can be argued that in vivo developmental events in terms of spatial and temporal changes are grossly recapitulated during the in vitro formation of neural rosettes.
PROLIFERATION AND RENEWAL OF NEURAL CELLS IN CULTURE
Understanding the active cell signaling in NP cells will help us determine the important factors needed to maintain uniform undifferentiated NP cells in tissue culture. Several studies indicate that many signaling pathways like, EGF, WNT, Hedgehog, TGFβ, leukemia inhibiting factor (LIF) and FGF are important for maintenance of NP cell proliferation. In addition to these pathways, two other aspects need attention for adaptation of any culture method for proper growth and self-renewal of NP cells: 1) to mimic the in vivo niche the system needs to provide an appropriate stem cell micro-environment with proper extracellular matrix (ECM), and 2) to supply nutrients and mitotic factors in the medium. These provisions are aimed at promoting symmetric divisions of undifferentiated NP cells in the culture [Huttner and Kosodo, 2005].
Ventricular zone (VZ) in the developing embryonic CNS (central nervous system) serves as the reservoir of proliferating neural progenitors and/or stem cells. It has been shown that laminin, a heterotrimeric family of ECM molecules is expressed in many different stem cell microenvironments including murine embryonic VZ [Campos et al., 2004]. Laminins are known to interact with β1 integrins receptors expressed on neural stem cell surface. The interaction between the beta integrins and laminin activates MAPK signaling that helps in maintenance of neural stem cells in CNS [Campos et al., 2004]. For this reason, laminin is widely used as substrate to coat tissue culture dishes for growing many types of stem cells including NP cells [Shin et al., 2006]. In addition to this, another ECM molecule, fibronectin, that acts also through β1 integrins are implicated for neural cell migration [Prestoz et al., 2001]. Thus, fibronectin has been used either in coating tissue culture dishes or added to medium to grow the NP cells [Carpenter et al., 2001; Elkabetz et al., 2008]. Collagen IV has been used as a neural cell culture substrate [Erceg et al., 2008]. Matrigel, a proprietary material from BD Bioscience, contains laminin, collagen and several growth factors, was used by several groups to differentiate hESC into neural cells in adherent culture and as well as in cells in scaffold [Baharvand et al., 2007; Levenberg et al., 2003].
To support NP cell proliferation we have supplemented neurobasal medium with L-glutamine, LIF (10 ng/ml), FGF2 (20 ng/ml) and B27 [Dhara et al., 2008; Shin et al., 2006]. Essentially, this is a serum-free formulation that supports prolonged growth and self-renewal of NP cells in culture. In addition, epidermal growth factors (EGF) had been detected in a wide area of rodent forebrain and mid brain [Fallon et al., 1984] and this growth factor plays an important role in the regulation of cell growth, proliferation, and differentiation, and other studies have described the use of EGF in the medium for culturing NP cells [Carpenter et al., 2001; Elkabetz et al., 2008].
REGIONAL SPECIFICATION OF NEURAL CELLS
As described before, nervous system consists of different regions. As with mouse ES cell [Barberi et al., 2003], derivation of region-specific NP cells from hESC will be of great interest for several reasons. Cells that carry their positional identity upon further differentiation, will express regional specific activity/ phenotype. Presumably being more responsive to environmental cues for a specific site, these progenitors are considered to be better suited for transplantation. In addition, they would potentially produce homogeneous cell types/ tissue and be useful to study the fundamental and applied neurobiology in vitro. It will be useful if these cells can be maintained for an extended period in culture, without altering their region-specific differentiation potentials. Here, region specific differentiation can be influenced by temporal factors and the morphogen concentration added to the culture.
Endogenous forebrain-specific NP cells appear first and characteristically express homeodomain transcription factors such as OTX1, OTX2 and BF1. Recently, Studer and his associates have derived and described rosette specific neural stem (R-NSC) cells that express anterior CNS specific marker Forse 1 [Elkabetz et al., 2008]. These cells propagated in the presence of mitogen FGF2 and retained Forse 1 expression, even though FGF2 was considered to caudalize the cell fate as found by another group [Zhang, 2006]. Zhang and colleagues also generated OTX2 and BF1 expressing NP cells from hESC [Li et al., 2005]. Forebrain specific NP cells have been derived from mouse ESC by inhibiting Wnt signaling [Watanabe et al., 2005] and by removing LIF from the culture medium [Ying et al., 2003]. Additional studies are required to determine if WNT and LIF signaling will affect regional specificity in hESC-derived NP cells as they did in mouse ESC studies.
Midbrain specific cells, dopamine neurons are of great interest for therapeutic use in Parkinson's disease. Midbrain specific cells express TH, RAX, NURR1, PTX3 and LMX1a/b, PAX2 and EN-1(also expressed in hindbrain cells). Even though FGF8 signaling enhances midbrain differentiation [Perrier et al., 2004], recently Stojkovic and colleagues demonstrated that FGF2 alone can generate matured dopamine neurons [Erceg et al., 2008]. Therefore, it seems that FGF2 has some role in generating neural cells of all regions along the antero-posterior axis.
Along the antero-posterior axis, hindbrain specific cells are specified by the expression of HOX genes under the influence of FGF2, SHH and RA. RA does not seem to initiate neural induction, rather it caudalizes the existing neuroepithelial cells. Whereas, along the dorso-ventral regions, BMPs are thought to specify cells of the dorsal region and SHH imparts ventral characteristics to the cells. Considering this interplay, protocols have been successfully used for generating region-specific cell types from hESC [Li et al., 2005; Li et al., 2008; Shin et al., 2005]. Along the same line, neural crest cells, peripheral neurons [Lee et al., 2007; Pomp et al., 2005] and oligodendrocyte progenitors [Keirstead et al., 2005] have been generated from hESC. Major factors used in regional subtype cell differentiation have been presented in Table 1. There is still a need for more homogenous types of differentiated cells in order to demonstrate that every cell type found in the human nervous system can be derived from hESC.
Table 1.
Morphogens used for differentiation of ESC-derived neural progenitors to regional subtype cells
| Target population | Morphogens/ Factors used | Reference |
|---|---|---|
| To continue as neural progenitors |
FGF2, ± EGF | [Elkabetz et al., 2008; Shin et al., 2006] |
| Neural crest cells; peripheral neurons |
SDIA co-culture | [Lee et al., 2007; Pomp et al., 2005] |
| GABA neurons | Neurotrophin-4 | *[Barberi et al., 2003] |
| Dopamine neurons | FGF8/Shh followed by ascorbic acid and BDNF |
[Perrier et al., 2004] |
| Serotonin neurons | Same condition as for Dopamine neurons except FGF4 instead of FGF8 |
*[Barberi et al., 2003] |
| Motoneurons | FGF2, RA, SHH | [Li et al., 2005; Shin et al., 2005] |
| Astrocytes | CNTF | *[Barberi et al., 2003] |
| Oligodendrocytes | Triiodothyronine/ NT-3 CNTF/ PDGF |
[Dhara et al., 2008; Keirstead et al., 2005; Shin et al., 2006] |
Neural progenitors (NP) are maintained in neurobasal medium supplemented with high level of mitogen FGF2. Removal of FGF2 from the medium differentiates them into many types of neurons and astro-glial cells. Here is a list of some factors that have directed differentiation of NP cells to different subtype neural cells (in mouse ES cells)
OUR EXPERIENCE WITH NEURAL DIFFERENTIATION
We have derived NP cells from hESC using different methods. In one method we grew hESC on mouse embryonic fibroblast (MEF) in hESC medium for the first 7 days and then in derivation medium for the next 7 days and finally propagated the rosette like cells in proliferation medium (Figure1). This procedure allowed us to generate a morphologically stable population of NP cells which grow in culture over 100 passages. Entire derivation and culture was performed in an adherent culture system [Shin et al., 2006]. We also derived NP cells from hESC without an initial propagation on MEF. This was aimed to minimize non-human components during the NP cell derivation. This differentiation technique produced a high percent of Nestin+ NP cells and this method further allowed us to evaluate the roles of some of the commonly used medium components. We found that the N2-supplement influenced the derivation of NP cells as indicated by up-regulation of NP cell specific marker in culture. Even though FGF2 has been shown to be involved in neural specification and further neural patterning [Lillien and Raphael, 2000], surprisingly we found FGF2 had negligible contribution to improving the derivation of NP cells. However it is an important mitogen in NP cell cultures and when FGF2 was withdrawn from medium, these cells became neurons expressing TuJ1, GABA, and TH [Dhara et al., 2008]. Using RA, FGF2 and SHH, they could be terminally differentiated into HB9 expressing cells with motor neuron phenotype marked by ChAT expression [Shin et al., 2005]. Further these NP cells were capable of producing astrocytes and oligodendrocytes under either basal conditions or with medium supplemented with ciliary neurotrophic factor or platelet-derived growth factor [Dhara et al., 2008; Shin et al., 2006].
In the future, NP cells could be used as a new primary cell source for cell-based assays of new drugs for neurodegenerative diseases like spinal muscular atrophy [Wilson et al., 2007]. Intuitively, better lead compounds for treating neural diseases such as spinal muscular atrophy will be found if they are screened in the cell type that the disease affects. As previously stated, NP cells can form a motor neuron phenotype in culture and should be used to study this devastating disease. We transfected NP cells with SMN1 and SMN2 gene expression and splice variants mini reporter genes [Wilson et al., 2007]. Potentially motor neurons differentiated from these modified NP cells would provide a more representative cell based assay to screen chemical libraries for compounds that could move spinal muscular atrophy drug candidates into animal testing. This is just one of many potential drug screening applications. Since NP cells express many receptors including G-protein coupled receptors (unpublished), ion channels and other drug targets NP cells and their derivatives will potentially serve as physiologically normal cell types for different diseases and drug evaluation.
CONCLUDING REMARKS
In the CNS, antero-posterior and dorso-ventral axes form unique grid like regions that generate specific subpopulation of neuronal and astro-glial cells. As discussed, the generation of glial and neuronal subtypes is dependent on specific morphogen concentration, time and duration of its exposure. It will be a challenge to generate and maintain such regionally committed but self renewing populations for therapeutic and research use. Although encouraging progress toward these goals continues to be made [Elkabetz et al., 2008; Erceg et al., 2008; Keirstead et al., 2005], but there is still more needs to be done in this area. Several approaches might expedite the process. For example, genetic manipulation should be further explored as a means of isolating regionally committed cell populations. One might express drug resistance genes under region-specific gene promoter which would eliminate undesired cell types and allow selective expansion of desired NP subpopulations.
Signaling pathways in the context of hESC differentiation to neural cells will be helpful to better culture the NP cells in vitro as well as to further differentiate them into pure populations of neurons/glial cells. One such example would be to explore the effect of ß-catenin signaling since it was demonstrated earlier that ß-catenin signaling was sufficient to induce and promote neurogenesis in mouse ES cultures even in the absence of RA [Otero et al., 2004].
In addition, a profile of cell surface receptors expressed by NP cells would not only help in finding the better ECM for supporting NP cell growth in culture, but also would be useful in better characterizing the NP subpopulation as well as identifying the potential drug targets. Further, moving to a completely defined medium for growth and differentiation of NP cells would be a great step-forward in the field.
ACKNOWLEDGEMNT
SKD is thankful to all current members of the Stice lab, especially to Dr. Anirban Majumder, Ms. Jennifer L. Mumaw, Dr. Mahesh C Dodla, and to Dr. David W. Machacek of the Aruna Biomedical, Inc for critically reading the manuscript. SKD also acknowledges the help from Mr. Animesh Dhara of the graduate program in neuroscience, University of Georgia, Athens, GA. Published work from this laboratory was supported by funds from Aruna Biomedical, Inc and NIH grants HL074303 and NS053272 to SLS.
Grant information:
Contract grant sponsor: Aruna Biomedical, Inc.; Contract grant number: N/A.
Contract grant sponsor: National Institute of Health; Contract grant number: HL074303.
Contract grant sponsor: National Institute of Health; Contract grant number: NS053272.
REFERENCES
- Baharvand H, Mehrjardi NZ, Hatami M, Kiani S, Rao M, Haghighi MM. Neural differentiation from human embryonic stem cells in a defined adherent culture condition. Int J Dev Biol. 2007;51:371–8. doi: 10.1387/ijdb.72280hb. [DOI] [PubMed] [Google Scholar]
- Barberi T, Klivenyi P, Calingasan NY, Lee H, Kawamata H, Loonam K, Perrier AL, Bruses J, Rubio ME, Topf N, Tabar V, Harrison NL, Beal MF, Moore MA, Studer L. Neural subtype specification of fertilization and nuclear transfer embryonic stem cells and application in parkinsonian mice. Nat Biotechnol. 2003;21:1200–7. doi: 10.1038/nbt870. [DOI] [PubMed] [Google Scholar]
- Cai C, Grabel L. Directing the differentiation of embryonic stem cells to neural stem cells. Dev Dyn. 2007;236:3255–66. doi: 10.1002/dvdy.21306. [DOI] [PubMed] [Google Scholar]
- Campos LS, Leone DP, Relvas JB, Brakebusch C, Fassler R, Suter U, ffrench-Constant C. Beta1 integrins activate a MAPK signalling pathway in neural stem cells that contributes to their maintenance. Development. 2004;131:3433–44. doi: 10.1242/dev.01199. [DOI] [PubMed] [Google Scholar]
- Carpenter MK, Inokuma MS, Denham J, Mujtaba T, Chiu CP, Rao MS. Enrichment of neurons and neural precursors from human embryonic stem cells. Exp Neurol. 2001;172:383–97. doi: 10.1006/exnr.2001.7832. [DOI] [PubMed] [Google Scholar]
- Dale JK, Vesque C, Lints TJ, Sampath TK, Furley A, Dodd J, Placzek M. Cooperation of BMP7 and SHH in the induction of forebrain ventral midline cells by prechordal mesoderm. Cell. 1997;90:257–69. doi: 10.1016/s0092-8674(00)80334-7. [DOI] [PubMed] [Google Scholar]
- Dhara SK, Hasneen K, Machacek DW, Boyd NL, Rao RR, Stice SL. Human neural progenitor cells derived from embryonic stem cells in feeder-free cultures. Differentiation. 2008;76:454–64. doi: 10.1111/j.1432-0436.2007.00256.x. [DOI] [PubMed] [Google Scholar]
- Doniach T. Basic FGF as an inducer of anteroposterior neural pattern. Cell. 1995;83:1067–70. doi: 10.1016/0092-8674(95)90133-7. [DOI] [PubMed] [Google Scholar]
- Elkabetz Y, Panagiotakos G, Al Shamy G, Socci ND, Tabar V, Studer L. Human ES cell-derived neural rosettes reveal a functionally distinct early neural stem cell stage. Genes Dev. 2008;22:152–65. doi: 10.1101/gad.1616208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erceg S, Lainez S, Ronaghi M, Stojkovic P, Perez-Arago MA, Moreno-Manzano V, Moreno-Palanques R, Planells-Cases R, Stojkovic M. Differentiation of human embryonic stem cells to regional specific neural precursors in chemically defined medium conditions. PLoS ONE. 2008;3:e2122. doi: 10.1371/journal.pone.0002122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fallon JH, Seroogy KB, Loughlin SE, Morrison RS, Bradshaw RA, Knaver DJ, Cunningham DD. Epidermal growth factor immunoreactive material in the central nervous system: location and development. Science. 1984;224:1107–9. doi: 10.1126/science.6144184. [DOI] [PubMed] [Google Scholar]
- Furuta Y, Piston DW, Hogan BL. Bone morphogenetic proteins (BMPs) as regulators of dorsal forebrain development. Development. 1997;124:2203–12. doi: 10.1242/dev.124.11.2203. [DOI] [PubMed] [Google Scholar]
- Gerhart J. 1998 Warkany lecture: signaling pathways in development. Teratology. 1999;60:226–39. doi: 10.1002/(SICI)1096-9926(199910)60:4<226::AID-TERA7>3.0.CO;2-W. [DOI] [PubMed] [Google Scholar]
- Gerrard L, Rodgers L, Cui W. Differentiation of human embryonic stem cells to neural lineages in adherent culture by blocking bone morphogenetic protein signaling. Stem Cells. 2005;23:1234–41. doi: 10.1634/stemcells.2005-0110. [DOI] [PubMed] [Google Scholar]
- Graham A, Francis-West P, Brickell P, Lumsden A. The signalling molecule BMP4 mediates apoptosis in the rhombencephalic neural crest. Nature. 1994;372:684–6. doi: 10.1038/372684a0. [DOI] [PubMed] [Google Scholar]
- Harland R. Neural induction. Curr Opin Genet Dev. 2000;10:357–62. doi: 10.1016/s0959-437x(00)00096-4. [DOI] [PubMed] [Google Scholar]
- Hong S, Kang UJ, Isacson O, Kim KS. Neural precursors derived from human embryonic stem cells maintain long-term proliferation without losing the potential to differentiate into all three neural lineages, including dopaminergic neurons. J Neurochem. 2008;104:316–24. doi: 10.1111/j.1471-4159.2007.04952.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huttner WB, Kosodo Y. Symmetric versus asymmetric cell division during neurogenesis in the developing vertebrate central nervous system. Curr Opin Cell Biol. 2005;17:648–57. doi: 10.1016/j.ceb.2005.10.005. [DOI] [PubMed] [Google Scholar]
- Itsykson P, Ilouz N, Turetsky T, Goldstein RS, Pera MF, Fishbein I, Segal M, Reubinoff BE. Derivation of neural precursors from human embryonic stem cells in the presence of noggin. Mol Cell Neurosci. 2005;30:24–36. doi: 10.1016/j.mcn.2005.05.004. [DOI] [PubMed] [Google Scholar]
- Kawasaki H, Mizuseki K, Nishikawa S, Kaneko S, Kuwana Y, Nakanishi S, Nishikawa SI, Sasai Y. Induction of midbrain dopaminergic neurons from ES cells by stromal cell-derived inducing activity. Neuron. 2000;28:31–40. doi: 10.1016/s0896-6273(00)00083-0. [DOI] [PubMed] [Google Scholar]
- Kawasaki H, Suemori H, Mizuseki K, Watanabe K, Urano F, Ichinose H, Haruta M, Takahashi M, Yoshikawa K, Nishikawa S, Nakatsuji N, Sasai Y. Generation of dopaminergic neurons and pigmented epithelia from primate ES cells by stromal cell-derived inducing activity. Proc Natl Acad Sci U S A. 2002;99:1580–5. doi: 10.1073/pnas.032662199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keirstead HS, Nistor G, Bernal G, Totoiu M, Cloutier F, Sharp K, Steward O. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. J Neurosci. 2005;25:4694–705. doi: 10.1523/JNEUROSCI.0311-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee G, Kim H, Elkabetz Y, Al Shamy G, Panagiotakos G, Barberi T, Tabar V, Studer L. Isolation and directed differentiation of neural crest stem cells derived from human embryonic stem cells. Nat Biotechnol. 2007;25:1468–75. doi: 10.1038/nbt1365. [DOI] [PubMed] [Google Scholar]
- Levenberg S, Huang NF, Lavik E, Rogers AB, Itskovitz-Eldor J, Langer R. Differentiation of human embryonic stem cells on three-dimensional polymer scaffolds. Proc Natl Acad Sci U S A. 2003;100:12741–6. doi: 10.1073/pnas.1735463100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li XJ, Du ZW, Zarnowska ED, Pankratz M, Hansen LO, Pearce RA, Zhang SC. Specification of motoneurons from human embryonic stem cells. Nat Biotechnol. 2005;23:215–21. doi: 10.1038/nbt1063. [DOI] [PubMed] [Google Scholar]
- Li XJ, Hu BY, Jones SA, Zhang YS, Lavaute T, Du ZW, Zhang SC. Directed differentiation of ventral spinal progenitors and motor neurons from human embryonic stem cells by small molecules. Stem Cells. 2008;26:886–93. doi: 10.1634/stemcells.2007-0620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lillien L, Raphael H. BMP and FGF regulate the development of EGF-responsive neural progenitor cells. Development. 2000;127:4993–5005. doi: 10.1242/dev.127.22.4993. [DOI] [PubMed] [Google Scholar]
- Maden M. Retinoic acid in the development, regeneration and maintenance of the nervous system. Nat Rev Neurosci. 2007;8:755–65. doi: 10.1038/nrn2212. [DOI] [PubMed] [Google Scholar]
- Mason I. Initiation to end point: the multiple roles of fibroblast growth factors in neural development. Nat Rev Neurosci. 2007;8:583–96. doi: 10.1038/nrn2189. [DOI] [PubMed] [Google Scholar]
- Miyazono K, Miyazawa K. Id: a target of BMP signaling. Sci STKE. 2002;2002:PE40. doi: 10.1126/stke.2002.151.pe40. [DOI] [PubMed] [Google Scholar]
- Mizuseki K, Sakamoto T, Watanabe K, Muguruma K, Ikeya M, Nishiyama A, Arakawa A, Suemori H, Nakatsuji N, Kawasaki H, Murakami F, Sasai Y. Generation of neural crest-derived peripheral neurons and floor plate cells from mouse and primate embryonic stem cells. Proc Natl Acad Sci U S A. 2003;100:5828–33. doi: 10.1073/pnas.1037282100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nutt SL, Dingwell KS, Holt CE, Amaya E. Xenopus Sprouty2 inhibits FGF-mediated gastrulation movements but does not affect mesoderm induction and patterning. Genes Dev. 2001;15:1152–66. doi: 10.1101/gad.191301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Rahilly R, Muller F. Neurulation in the normal human embryo. Ciba Found Symp. 1994;181:70–82. doi: 10.1002/9780470514559.ch5. discussion 82-9. [DOI] [PubMed] [Google Scholar]
- Okabe S, Forsberg-Nilsson K, Spiro AC, Segal M, McKay RD. Development of neuronal precursor cells and functional postmitotic neurons from embryonic stem cells in vitro. Mech Dev. 1996;59:89–102. doi: 10.1016/0925-4773(96)00572-2. [DOI] [PubMed] [Google Scholar]
- Otero JJ, Fu W, Kan L, Cuadra AE, Kessler JA. Beta-catenin signaling is required for neural differentiation of embryonic stem cells. Development. 2004;131:3545–57. doi: 10.1242/dev.01218. [DOI] [PubMed] [Google Scholar]
- Park S, Lee KS, Lee YJ, Shin HA, Cho HY, Wang KC, Kim YS, Lee HT, Chung KS, Kim EY, Lim J. Generation of dopaminergic neurons in vitro from human embryonic stem cells treated with neurotrophic factors. Neurosci Lett. 2004;359:99–103. doi: 10.1016/j.neulet.2004.01.073. [DOI] [PubMed] [Google Scholar]
- Pera MF, Andrade J, Houssami S, Reubinoff B, Trounson A, Stanley EG, Ward-van Oostwaard D, Mummery C. Regulation of human embryonic stem cell differentiation by BMP-2 and its antagonist noggin. J Cell Sci. 2004;117:1269–80. doi: 10.1242/jcs.00970. [DOI] [PubMed] [Google Scholar]
- Perrier AL, Tabar V, Barberi T, Rubio ME, Bruses J, Topf N, Harrison NL, Studer L. Derivation of midbrain dopamine neurons from human embryonic stem cells. Proc Natl Acad Sci U S A. 2004;101:12543–8. doi: 10.1073/pnas.0404700101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pomp O, Brokhman I, Ben-Dor I, Reubinoff B, Goldstein RS. Generation of peripheral sensory and sympathetic neurons and neural crest cells from human embryonic stem cells. Stem Cells. 2005;23:923–30. doi: 10.1634/stemcells.2005-0038. [DOI] [PubMed] [Google Scholar]
- Prestoz L, Relvas JB, Hopkins K, Patel S, Sowinski P, Price J, ffrench-Constant C. Association between integrin-dependent migration capacity of neural stem cells in vitro and anatomical repair following transplantation. Mol Cell Neurosci. 2001;18:473–84. doi: 10.1006/mcne.2001.1037. [DOI] [PubMed] [Google Scholar]
- Price PJ, Brewer GJ. Serum-free media for neural cell culture. In: Fedoroff S, Richardson A, editors. Protocols for Neural Cell Culture. Humana Press; 2001. pp. 255–264. [Google Scholar]
- Reubinoff BE, Itsykson P, Turetsky T, Pera MF, Reinhartz E, Itzik A, Ben-Hur T. Neural progenitors from human embryonic stem cells. Nat Biotechnol. 2001;19:1134–40. doi: 10.1038/nbt1201-1134. [DOI] [PubMed] [Google Scholar]
- Robertson MJ, Gip P, Schaffer DV. Neural stem cell engineering: directed differentiation of adult and embryonic stem cells into neurons. Front Biosci. 2008;13:21–50. doi: 10.2741/2558. [DOI] [PubMed] [Google Scholar]
- Schuldiner M, Eiges R, Eden A, Yanuka O, Itskovitz-Eldor J, Goldstein RS, Benvenisty N. Induced neuronal differentiation of human embryonic stem cells. Brain Res. 2001;913:201–5. doi: 10.1016/s0006-8993(01)02776-7. [DOI] [PubMed] [Google Scholar]
- Schulz TC, Palmarini GM, Noggle SA, Weiler DA, Mitalipova MM, Condie BG. Directed neuronal differentiation of human embryonic stem cells. BMC Neurosci. 2003;4:27. doi: 10.1186/1471-2202-4-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimamura K, Rubenstein JL. Inductive interactions direct early regionalization of the mouse forebrain. Development. 1997;124:2709–18. doi: 10.1242/dev.124.14.2709. [DOI] [PubMed] [Google Scholar]
- Shin S, Dalton S, Stice SL. Human motor neuron differentiation from human embryonic stem cells. Stem Cells Dev. 2005;14:266–9. doi: 10.1089/scd.2005.14.266. [DOI] [PubMed] [Google Scholar]
- Shin S, Mitalipova M, Noggle S, Tibbitts D, Venable A, Rao R, Stice SL. Long-term proliferation of human embryonic stem cell-derived neuroepithelial cells using defined adherent culture conditions. Stem Cells. 2006;24:125–38. doi: 10.1634/stemcells.2004-0150. [DOI] [PubMed] [Google Scholar]
- Streit A, Berliner AJ, Papanayotou C, Sirulnik A, Stern CD. Initiation of neural induction by FGF signalling before gastrulation. Nature. 2000;406:74–8. doi: 10.1038/35017617. [DOI] [PubMed] [Google Scholar]
- Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145–7. doi: 10.1126/science.282.5391.1145. [DOI] [PubMed] [Google Scholar]
- Vazin T, Chen J, Lee CT, Amable R, Freed WJ. Assessment of stromal-derived inducing activity in the generation of dopaminergic neurons from human embryonic stem cells. Stem Cells. 2008;26:1517–25. doi: 10.1634/stemcells.2008-0039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe K, Kamiya D, Nishiyama A, Katayama T, Nozaki S, Kawasaki H, Watanabe Y, Mizuseki K, Sasai Y. Directed differentiation of telencephalic precursors from embryonic stem cells. Nat Neurosci. 2005;8:288–96. doi: 10.1038/nn1402. [DOI] [PubMed] [Google Scholar]
- Wilson PG, Cherry JJ, Schwamberger S, Adams AM, Zhou J, Shin S, Stice SL. An SMA project report: neural cell-based assays derived from human embryonic stem cells. Stem Cells Dev. 2007;16:1027–41. doi: 10.1089/scd.2007.0061. [DOI] [PubMed] [Google Scholar]
- Wilson PG, Stice SS. Development and differentiation of neural rosettes derived from human embryonic stem cells. Stem Cell Rev. 2006;2:67–77. doi: 10.1007/s12015-006-0011-1. [DOI] [PubMed] [Google Scholar]
- Ying QL, Stavridis M, Griffiths D, Li M, Smith A. Conversion of embryonic stem cells into neuroectodermal precursors in adherent monoculture. Nat Biotechnol. 2003;21:183–6. doi: 10.1038/nbt780. [DOI] [PubMed] [Google Scholar]
- Zhang SC. Neural subtype specification from embryonic stem cells. Brain Pathol. 2006;16:132–42. doi: 10.1111/j.1750-3639.2006.00008.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang SC, Wernig M, Duncan ID, Brustle O, Thomson JA. In vitro differentiation of transplantable neural precursors from human embryonic stem cells. Nat Biotechnol. 2001;19:1129–33. doi: 10.1038/nbt1201-1129. [DOI] [PubMed] [Google Scholar]
- Zhang Z-WDaS-C. Neural Differentiation from Embryonic Stem Cells:Which Way? Stem Cells and Development. 2004;13:372–381. doi: 10.1089/scd.2004.13.372. [DOI] [PubMed] [Google Scholar]