

Abstract
Mutations in the progranulin gene (GRN) are a major cause of frontotemporal lobar degeneration with ubiquitin-positive, tau-negative inclusions (FTLD-U) but the distinguishing clinical and anatomical features of this subgroup remain unclear. In a large UK cohort we found five different frameshift and premature termination mutations likely to be causative of FTLD in 25 affected family members. A previously described 4-bp insertion mutation in GRN exon 2 comprised the majority of cases in our cohort (20/25), with four novel mutations being identified in the other five affected members. Additional novel missense changes were discovered, of uncertain pathogenicity, but deletion of the entire gene was not detected. The patient collection was investigated by a single tertiary referral centre and is enriched for familial early onset FTLD with a high proportion of patients undergoing neuropsychological testing, MRI and eventual neuropathological diagnosis. Age at onset was variable, but four mutation carriers presented in their 40s and when analysed as a group, the mean age at onset of disease in GRN mutation carriers was later than tau gene (MAPT) mutation carriers and duration of disease was shorter when compared with both MAPTand FTLD-U without mutation. The most common clinical presentation seen in GRN mutation carriers was behavioural variant FTLD with apathy as the dominant feature. However, many patients had language output impairment that was either a progressive non-fluent aphasia or decreased speech output consistent with a dynamic aphasia. Neurological and neuropsychological examination also suggests that parietal lobe dysfunction is a characteristic feature of GRN mutation and differentiates this group from other patients with FTLD. MR imaging showed evidence of strikingly asymmetrical atrophy with the frontal, temporal and parietal lobes all affected. Both right- and left-sided predominant atrophy was seen even within the same family. As a group, the GRN carriers showed more asymmetry than in other FTLD groups. All pathologically investigated cases showed extensive type 3 TDP-43-positive pathology, including frequent neuronal cytoplasmic inclusions, dystrophic neurites in both grey and white matter and also neuronal intranuclear inclusions. Finally, we confirmed a modifying effect of APOE-E4 genotype on clinical phenotype with a later onset in the GRN carriers suggesting that this gene has distinct phenotypic effects in different neurodegenerative diseases.
Keywords: frontotemporal lobar degeneration, frontotemporal dementia, progranulin, progressive aphasia
Introduction
Frontotemporal lobar degeneration (FTLD) comprises a clinically heterogeneous group of disorders that primarily affect either personality or language. Together, these conditions account for a large proportion of early-onset dementia. Three clinical syndromes are described by consensus criteria (Neary et al., 1998): frontotemporal dementia (FTD or behavioural variant FTLD, bvFTLD), semantic dementia (SD) and progressive non-fluent aphasia (PNFA). bvFTLD is the most common clinical presentation with early behavioural and personality changes. SD is characterized by loss of semantic knowledge and a progressive fluent aphasia whereas PNFA presents with difficulties in speech production. A number of related neurodegenerative diseases with a prominent motor component overlap with FTLD, namely the corticobasal syndrome (CBS), progressive supranuclear palsy (PSP) and motoneuron disease (MND) (Kertesz et al., 1999; Lomen-Hoerth et al., 2002).
The neuropathological substrate of FTLD is the degeneration of the frontal and temporal lobes (Brun et al., 1994). There are two major immunocytochemical subdivisions of FTLD: tauopathies with an accumulation of intraneuronal hyperphosphorylated tau, and FTLD-U with intraneuronal inclusions immunoreactive to ubiquitin (Cairns et al., 2007). Recently the majority of conditions associated with FTLD-U have been linked by the identification of TDP-43 as the major ubiquitinated protein component of the inclusions (Neumann et al., 2006) and four TDP-43 subtypes are now described (Cairns et al., 2007). The clinical categories of FTLD may predict neuropathology to some extent: CBS and PSP are normally tauopathies, whereas SD and FTD associated with MND (FTD-MND) typically have TDP-43 neuropathology (Hodges et al., 2004; Snowden et al., 2007).
The last decade has seen major developments in the understanding of the well-known genetic predisposition to FTLD in 25–50% of cases (Stevens et al., 1998; Bird et al., 2003). Mutations were first discovered in the microtubule-associated protein tau gene (MAPT), which account for 5–10% of FTLD (Hutton et al., 1998, Poorkaj et al., 1998; Spillantini et al., 1998). Mutations in the CHMP2B (Skibinski et al., 2005) and VCP genes (Watts et al., 2004) were later discovered to be rare causes of FTLD. More recently, mutations have been described in the progranulin gene on chromosome 17 (GRN), finally explaining the conundrum of genetic linkage of families with FTLD-U to a region including MAPT (Baker et al., 2006; Cruts et al., 2006). GRN has 12 coding exons, translating to a 593 amino acid secreted glycoprotein of 69 kDa containing tandem repeats of a granulin motif. Cleavage of these repeats produces 6 kDa active granulin peptides that are widely distributed. Granulin peptides have roles in normal development, wound healing and tumourigenesis amongst other effects (Eriksen and Mackenzie, 2007).
A number of previously described FTLD families linked to chromosome 17 have now been shown to have GRN mutations (Lendon et al., 1998; Rosso et al., 2001; Rademakers et al., 2002; Baker et al., 2006; Cruts et al., 2006; Mackenzie et al., 2006; Mukherjee et al., 2006; van der Zee et al., 2006; Behrens et al., 2007; Bronner et al., 2007) and various groups have described GRN mutation series or case reports (Gass et al., 2006; Masellis et al., 2006; Davion et al., 2007; Kelley et al., 2007; Le Ber et al., 2007; Leverenz et al., 2007; Mesulam et al., 2007). In the large referral series, 5–10% of patients had GRN mutation (13–25% in cases with a family history) although this proportion decreased in an unbiased FTLD population. Over 40 different pathogenic mutations have now been described (www.molgen.ua.ac.be/FTDmutations) with all definitely pathogenic mutations having a null effect: either by frameshift, premature termination or disruption of a splice site. The pathological features of GRN mutation cases have been studied in detail with all showing ubiquitin-positive, TDP-43-positive neuronal intracytoplasmic inclusions (NCI), neurites and neuronal intranuclear inclusions (NII), i.e. type 3 TDP-43 pathology according to recently revised consensus criteria (Cairns et al., 2007). Some groups have reported co-existent tau, amyloid or alpha-synuclein pathology in a few cases (Behrens et al., 2007; Leverenz et al., 2007). Age at onset appears to be variable with the youngest GRN mutation case thus far reported being 35 (Leverenz et al., 2007) and the oldest case being 83 years old at onset (Gass et al., 2006) with wide variability even within the same family. Some studies have shown evidence of non-penetrance which may be age related (Gass et al., 2006). Clinically, patients tend to present with either bvFTLD or PNFA although CBS has also been described (Benussi et al., 2006, Masellis et al., 2006; Spina et al., 2007). However, detailed clinical, neuropsychological and radiological studies of GRN mutations are lacking. Here we describe the clinical, neuropsychological and MRI features and genetic modification in a UK series of GRN mutations from a large cohort of FTLD investigated at a specialist referral centre.
Material and Methods
The cohort of patients assessed in this study consisted of 212 patients with a clinical diagnosis of FTLD, CBS or PSP according to consensus criteria (Litvan et al., 1996; Neary et al., 1998; Boeve et al., 2003) and 35 patients with familial MND. The FTLD cohort had either been seen as patients at the National Hospital for Neurology and Neurosurgery, London, UK or were members of a longitudinal study in FTLD at the Dementia Research Centre, Institute of Neurology, London, UK. This study was approved by the local ethics committee. The clinical diagnoses in these patients were as follows: 120 bvFTLD (including two with FTD-MND), 23 SD, 36 PNFA (including five with a progressive mixed aphasia with features of both PNFA and SD), 23 CBS (including eight with a PNFA/CBS overlap) and 10 PSP.
Clinical assessment
All of the FTLD patients had been assessed by an experienced cognitive neurologist (M.N.R., N.C.F. or J.D.W.). Assessment included a standard clinical interview with both the patient and separately with an informant (usually a relative of the patient) as well as a formal neurological examination including assessment of limb and orofacial apraxia. Behavioural symptoms were ascertained by discussion with the patient's informant and also by observation of the patient during the clinical interview.
Neuropsychological assessment
Neuropsychological assessment consisted of a battery of cognitive tests. Verbal and visual episodic memory were tested using the Recognition Memory Tests for Words and Faces, respectively (Warrington, 1984; Warrington, 1996). Executive function was assessed using a Modified Card Sorting test or the Weigl test (Weigl, 1948; Nelson, 1976). Either the National Adult Reading Test or the Schonell reading test was used to test reading skills (Nelson and McKenna, 1975; Nelson, 1982). Naming was assessed with the Graded Naming Test or the Oldfield naming test (Oldfield and Wingfield, 1965; McKenna and Warrington, 1980) whilst single-word comprehension was tested using the Warrington Synonyms Test (Warrington et al., 1998). Other cognitive domains assessed were spelling (Graded Difficulty Spelling Test, Baxter and Warrington, 1994), calculation (Graded Difficulty Arithmetic Test, Jackson and Warrington, 1986) and visuospatial and visuoperceptual skills [subtests of the Visual Object and Spatial Perception (VOSP) battery: Warrington and James, 1991, or the Block Design subtest of the WAIS-R, Wechsler, 1981]. Patients were said to have a deficit in a particular cognitive domain if they scored below the 5th percentile on the test and their score was thought to represent a true deficit in that domain rather than being secondary to other factors such as concentration or attention.
Analysis of family history
All patients in the FTLD cohort were given a ‘Goldman score’ between 1 and 4 as per Goldman et al. (2005) where 1 is an autosomal dominant family history of FTLD, MND, CBS or PSP, 2 is familial aggregation of three of more family members with dementia, 3 is one other first degree relative with dementia (modified to give a score of 3 only if there is a history of young-onset dementia within the family i.e. <65, and 3.5 if onset above 65) and 4 is no or unknown family history. Thirty-eight cases were known to be members of families with MAPT mutations (nine families in total) whilst of the MAPT-negative cases there were 61 cases with a Goldman score of 1, 2 or 3 (from 36 separate families); 20 cases with a score of 3.5 and 93 cases with a score of 4.
MR analysis
Eight patients with definite GRN mutations had volumetric MRI scans. These scans underwent blind visual assessment by an experienced neuroradiologist. The volumetric images were also compared to a group of nine patients with MAPT mutations and a group of eight patients with GRN-negative ubiquitin-positive pathology who had a clinical diagnosis of SD. Image analysis was performed using the MIDAS software package (Freeborough et al., 1997). Scans were outlined using a rapid semi-automated technique, which involves interactive selection of thresholds, followed by a series of erosions and dilations. This yields a brain region which is separated from surrounding cerebrospinal fluid (CSF), skull and dura. Scans and their associated brain regions were then transformed into standard space by registration to the Montreal Neurological Institute (MNI) Template (Mazziotta et al., 1995). A brain region was defined on the MNI template and was subsequently divided into left and right hemispheres and anterior and posterior hemisections. An intersection of each individual's brain region and the regions defined on the MNI template were generated to provide a measure of brain volume in the following regions: left hemisphere, right hemisphere, left anterior, left posterior, right anterior and right posterior.
DNA sequencing
All 13 exons of GRN were assessed by direct sequencing from genomic DNA. Polymerase chain reaction (PCR) amplicons were generated using primers at 500 μM in MegaMix Blue PCR cocktail (Microzone). Details of primers and PCR conditions are available on request. Amplicons cleaned with Microclean (Microzone) were sequenced using Applied Biosytems BigDye v1.1 cycle sequence chemistry with 1 μl BigDye, 5 μl BetterBuffer (Microzone), 0.75 μl sequencing primer at 5 pmol/μl, 2.5 ng of cleaned amplicon and ddH2O to a final volume of 15 μl. Following 25 thermal cycles, sequencing products were precipitated and cleaned used standard EtOH/EDTA methodology and electrophoresed on an ABI 3130xl automated sequencer. Data was analysed using ABI Seqscape software v2.5.
Microsatellite haplotyping
PCR amplicons were generated using fluorescently end-labelled primers at 500 μM for microsatellite markers D17S951(FAM), D17S950(HEX), MAPT-1(FAM) and D17S791(FAM) in MegaMix Blue PCR cocktail (Microzone). A loading mix of 1 μl amplicon diluted 1:50 in ddH2O, 9.25 μl HiDi formamide (ABI) and 0.25 μl 500LIZ size standard (ABI) was prepared and DNA products were electrophoresed on an ABI 3130xl automated sequencer. Data was analysed using ABI GeneMapper software v4.0.
APOE genotyping
APOE genotyping was performed by Cfo1 restriction endonuclease digestion of PCR amplicon and size differentiation by agarose gel electrophoresis.
Copy number analysis
Quantification of GRN copy number was performed using the 5′ nuclease assay in conjunction with fluorescent MGB probes (ABI). Probes were designed to detect GRN and APP genes by hybridization to exon 1 and exon 5, respectively. An ABI Prism 7000 Sequence Detection System was used for relative quantification of GRN against APP using the ΔΔCt method. Normal healthy control and Down's syndrome control DNA were assessed to establish ΔCt values representative of GRN:APP ratios of 1:1 and 0.67:1, respectively, thereby demonstrating required sensitivity to detect the predicted ratio of 0.5 : 1 in potential GRN gene deletions. Standard curves (data not shown) showed that both probes had equal efficiencies, satisfying criteria for the comparative Ct method of quantification. Total reaction volume was 20 μl using 200 nmol probes and 900 nmol primers, Quantitect mastermix (Qiagen) and amplified using standard methods.
Neuropathology
Tissue blocks were selected from representative brain areas and 7 μm tissue sections were stained with haematoxylin and eosin and used for immunohistochemistry with antibodies to TDP-43 (Abnova, 1:800), p62 (BD Transduction Laboratories, 1 : 100) and ubiquitin (Dako, 1 : 200). Immunohistochemical staining was performed using a standard avidin–biotin method.
Statistical analysis
Statistical analysis was performed with SPSS (SPSS Inc.) or Excel 2007 (Microsoft Corp.). Means are quoted±95% confidence intervals calculated from 1.96 × SEM.
Results
Mutation detection
From the entire clinical series of 247 individuals (FTLD n = 212, MND n = 35), we sequenced 36 probands with a Goldman score of 1, 2 or 3, 20 patients with a Goldman score of 3.5, 93 patients with a Goldman score of 4 and 17 probands with familial MND. All exons and splice sites were sequenced in at least one direction in each individual. Five pathogenic mutations were detected, one caused premature termination (Q300X) and four were small insertions or deletions causing frameshift (Fig. 1). These mutations were found in eight probands with a Goldman score of 1, 2 or 3 and one patient (7651_3) with a Goldman score of 4 whose father had died at 78 with no formal diagnosis but with ‘memory and behavioural problems’ for several years before death. Co-segregation was shown in a further five family members (from families DRC32, DRC240, DRC255—Table 1). The entire series of 212 patients contained 16 other members from the eight families with GRN mutations and clinical data were available for each of these patients as well as the one ‘sporadic’ case (7651_3) i.e. 25 in total (Fig. 2). This group represents 12% of the total FTLD clinical series and 20% of the familial cases (Goldman score 1, 2, 3 or 3.5). Unaffected family members were not tested and no mutations were found in the MND cohort. Notably, we found no patients with the Arg493X mutation, which is said to be the most common GRN mutation with likely origins in a UK ancestor (Rademakers et al., 2007).
Fig. 1.
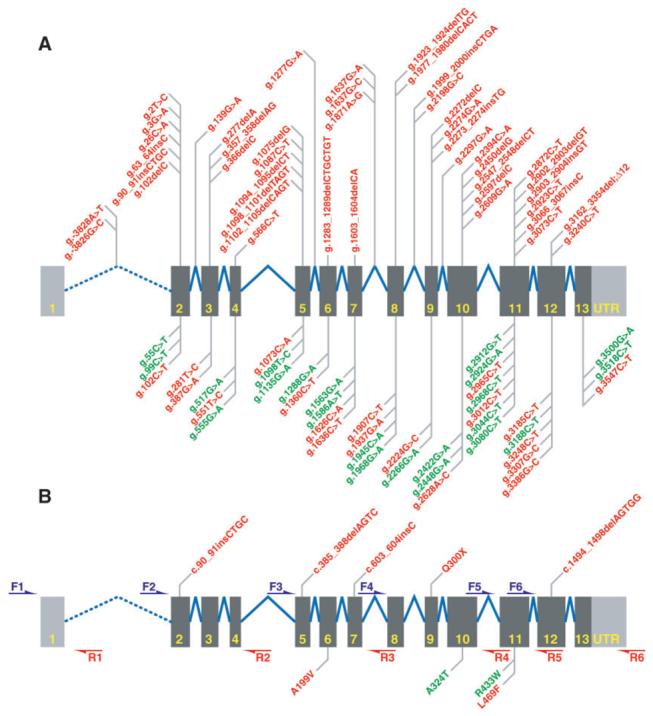
Schematic of GRN showing (A) exons (dark grey bars), UTR (light grey bars) and introns (not to scale, blue lines). Probable null mutations and mutations with unclear pathogenicity are shown above or below the introns/exons respectively (red). Known polymorphisms (green). (B) Screening strategy with six PCR primer pairs (F = forward primer; R = reverse primer); pathogenic and possibly pathogenic mutations (red) are shown above or below the introns/exons respecetively. Also shown are known polymorphisms (green) detected in our screen.
Table 1.
GRN mutations
|
GRN mutation |
Location | HGV nomenclature |
APOE | ||
|---|---|---|---|---|---|
| DRC32_4 | 4-bp insertion | Exon 2 | c.90_91insCTGC | 3 | 3 |
| DRC32_5 | 4-bp insertion | Exon 2 | c.90_91insCTGC | 3 | 3 |
| DRC39_3 | 4-bp insertion | Exon 2 | c.90_91insCTGC | 3 | 3 |
| DRC169_2 | Q300X | Exon 9 | c.898C>T | 3 | 3 |
| DRC240_4 | 4-bp insertion | Exon 2 | c.90_91insCTGC | 3 | 3 |
| DRC240_5 | 4-bp insertion | Exon 2 | c.90_91insCTGC | 3 | 3 |
| DRC255_6 | 4-bp insertion | Exon 2 | c.90_91insCTGC | 3 | 3 |
| DRC255_7 | 4-bp insertion | Exon 2 | c.90_91insCTGC | 2 | 3 |
| DRC255_9 | 4-bp insertion | Exon 2 | c.90_91insCTGC | 3 | 3 |
| DRC255_10 | 4-bp insertion | Exon 2 | c.90_91insCTGC | 3 | 3 |
| DRC425_2 | 4-bp deletion | Exon 5 | c.385_388delAGTC | 3 | 3 |
| DRC430_2 | 5-bp deletion | Exon 12 | c.1494_1498delAGTGG | 3 | 4 |
| DRC431_3 | 1-bp insertion | Exon 7 | c. 603_604insC | 3 | 3 |
| 7651_3 | 4-bp deletion | Exon 5 | c.385_388delAGTC | 3 | 4 |
| DRC429_2 | A199V | Exon 6 | c.596C>T | 3 | 3 |
| 5440_3 | L469F | Exon 11 | c.1407G>T | 3 | 4 |
Fig. 2.

Genealogies of pathogenic and possibly pathogenic mutation carriers. Individuals are identified by the family number (above genealogy)_individual number (adjacent to symbol). Filled symbols refer to definitely affected individuals. Half-filled symbols refer to possibly affected individuals who we were unable to clinically confirm.‘?’ refers to individuals who died aged <70 and may have transmitted the mutation without presentation with FTD.
We also found two sequence variants of a possibly pathogenic nature. A199V was found in a patient with familial CBS (DRC429_2) of Bangladeshi origin (Goldman score 3) but samples were not available from other family members to test for segregation. A199V was not found in 90 samples from healthy control Gujaratis, which represented the most appropriate control population available. L469F was found in a proband (5440_3) with sporadic FTLD (Goldman score 4) whose mother died at 92 but whose father died at 56 of heart disease. Samples were not available from other family members to test for segregation but L469F was not found in over 100 samples from the healthy young UK population.
Non-pathogenic changes
We found six other sequence variants (Tables 2 and 3). Although A324T was found in two patients and has not been described in healthy control individuals, one of our patients was also a carrier of the pathogenic Q300X mutation suggesting that the additional presence of A324T is of uncertain pathogenicity. R433W was found in two patients and has been described in healthy control individuals by other laboratories. Four common non-coding exonic and intronic variants are shown in Table 3.
Table 2.
GRN missense variants
| GRN variation | Location | HGV nomenclature | |
|---|---|---|---|
| DRC169_2a | A324T | Exon 10 | c.970C>A |
| 1498 | A324T | Exon 10 | c.970C>A |
| 17343 | R433W | Exon 11 | c.1297C>G |
| 12424 | R433W | Exon 11 | c.1297C>G |
Individual with Q300X.
Table 3.
GRN polymorphisms
| GRN polymorphisms | Allele frequency of cases (%) |
Genotype |
||
|---|---|---|---|---|
| +/+ | ± | −/− | ||
| IVS4-50_-49insGTCA, c.350-50_-49insGTCA | 0.22 | 7 | 30 | 63 |
| 50′-111delC | 0.28 | 7 | 27 | 39 |
| IVS4+24G>A, c.462+24G>A | 0.17 | 5 | 14 | 50 |
| IVS7+7G>A | 0.11 | 0 | 15 | 59 |
Copy number alteration
Our screening strategy for GRN deletion was to test the relative amplification of a GRN exon 1 qPCR against amplification of APP exon 5. An anonymous sample from a Down's syndrome patient was used as a control, for which the expected allelic ratios GRN:APP are 2:3 (healthy population ratio 1:1). In the case of GRN deletion, we would expect an allelic ratio GRN:APP of 1:2. None of the FTLD/MND cohort samples exceeded the allelic ratio of the Down's syndrome control (n = 171). We thus found no evidence of GRN deletion in our UK series.
c.90_91insCTGC mutation
The DRC255 mutation (4 bp, exon 2, c.90_91insCTGC) was found in four independently ascertained British families accounting for 20 out of 25 (80%) of our GRN mutation clinical cohort. The same mutation has been previously described in a Canadian family of English descent known as UBC17 (Baker et al., 2006; Mackenzie et al., 2006). Haplotype analysis with four microsatellite markers at the GRN locus supports a shared common ancestor of UBC17 and the British families (Table 4). One of the four families shared mutation-linked alleles at two microsatellites adjacent to GRN, suggesting recombination at both 5′ and 3′ ends of the disease haplotype, 0.6 and 2.5 MB from GRN. Although consistent with a common ancestor of all families, as the shared alleles are rare in the healthy population, a small mutation-linked haplotype supports a more distant ancestral relationship between family DRC39 and others with the same mutation.
Table 4.
Microsatellite haplotype data for mutations occurring in independently ascertained families
| c.90_91insCTGC |
c.385_388delAGTC |
||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DRC32_5 |
DRC240_4 |
DRC240_5 |
DRC255_10 |
DRC255_7 |
DRC39_3 |
7651_3 |
DRC425_2 |
||||||||||||
| Allele Primer # |
Allele Label |
Allele Name |
Allele Location on chromosome 17 |
Allele 1 | Allele 2 | Allele 1 | Allele 2 | Allele 1 | Allele 2 | Allele 1 | Allele 2 | Allele 1 | Allele 2 | Allele 1 | Allele 2 | Allele 1 | Allele 2 | Allele 1 | Allele 2 |
| 1 581/582 | 6-FAM | D17S951 | 39175619–39175792 | 176 | 170 | 176 | 176 | 176 | 168 | 176 | 198 | 176 | 168 | 170 | 168 | 166 | 168 | 166 | 174 |
| – | – | GRN | 39778015–39786006 | ||||||||||||||||
| 2 583/584 | 5′ HEX | D17S950 | 40615954–40616143 | 180 | 176 | 180 | 184 | 180 | 192 | 180 | 174 | 180 | 192 | 180 | 180 | 180 | 178 | 180 | 174 |
| 4 587/588 | 6-FAM | MAPT-1 | 41327364–41461548 | 358 | 360 | 358 | 362 | 358 | 376 | 358 | 376 | 358 | 360 | 358 | 360 | 358 | 346 | 358 | 346 |
| 3 585/586 | 6-FAM | D17S791 | 42211537–42211691 | 183 | 185 | 183 | 161 | 189 | 163 | 183 | 181 | 183 | 161 | 187 | 189 | 183 | 161 | 183 | 161 |
The likely mutation linked alleles are shown in bold. Possibly recombinant loci are shown with a black background.
Age at onset and disease duration of GRN mutation compared with MAPTand FTLD-U
Age at onset or death were available for 25 GRN mutation carriers and compared with 26 MAPT mutation carriers and 15 cases with FTLD-U pathology at autopsy (four familial and 11 sporadic) but no GRN mutation (Figs 3 and 4). Mean age at onset for GRN was 57±2.5 years, 8 years later than for MAPT at 49±2.4 years (t-test, two-tailed, P = 8.3 × 10−5). However, age at onset was similar between GRN and FTLD-U with no GRN mutation (GRN-negative age at onset 56±3.4 years, t-test, two-tailed, P = 0.73). GRN carriers had a much shorter clinical duration than other subtypes of FTLD. Mean clinical duration for GRN was 5±1.0 years compared with MAPT 12± 2.0 years (t-test, two-tailed, P = 7.8 × 10−6) and FTLD-U with no GRN mutation 9±1.7 years (t-test, two-tailed, P = 0.002). Across these three aetiological subgroups, a later age at onset was associated with a shorter clinical duration (Pearson r = −0.35, P = 0.006). We fitted a linear regression model to question the proportion of the variation in clinical duration accounted for by aetiological subgroup and/or age at onset. The model with minimal residual that best accounted for clinical duration incorporated aetiological subgroup and age at onset (P = 6.5 × 10−6), this model accounted for 34% of the variance of clinical duration.
Fig. 3.
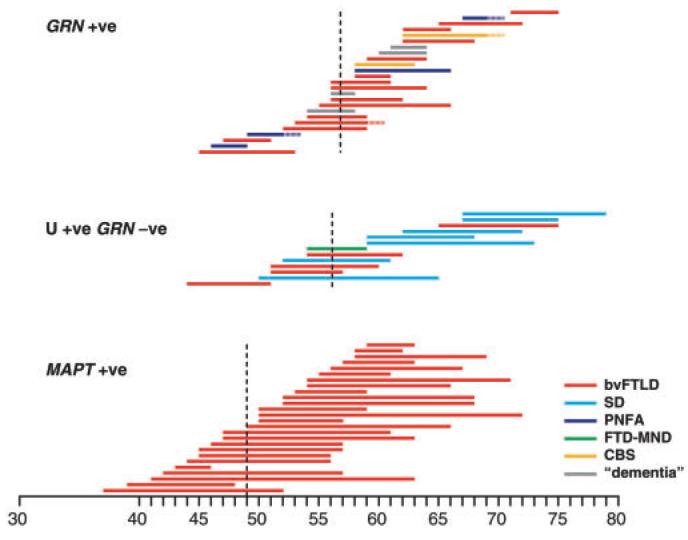
Clinical duration and clinical category of FTD is shown as a coloured bar in GRN mutation carriers (GRN-positive), MAPT mutation carriers (MAPT-positive) and wild-type GRN FTLD-U. See text for statistical analysis. Dotted vertical line indicates the mean age at clinical onset for each group.
Fig. 4.
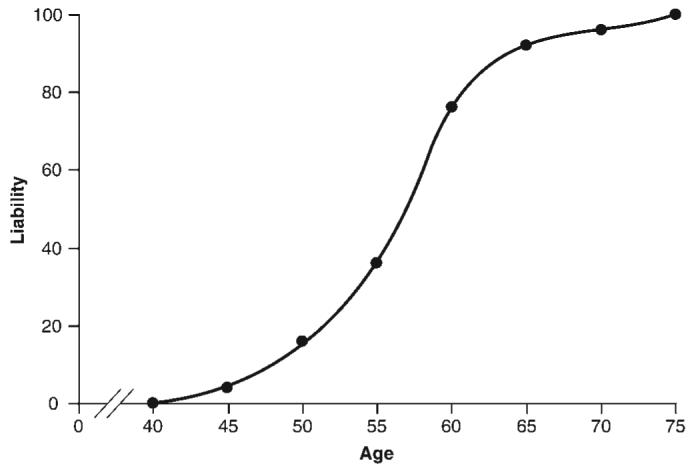
Liability curve derived from 25 GRN mutation carriers. These data are not a complete assessment of penetrance as we did not screen unaffected family members.
Modification of phenotype by APOE genotype or wild-type GRN alleles
APOE genotyping was performed on definite and possibly pathogenic GRN mutation carriers (n = 16, Table 1) to test a hypothesis generated by Gass and colleagues (Gass et al., 2006). In their international series the presence of at least one E4 allele was associated with a 6-year delayed age at onset compared with the E3E3 genotype. In our patient collection, age at onset was similarly delayed by 6 years: age at onset with at least one E4 allele was 62±2 years as opposed to 56±4 years without an E4 allele (t-test, two-tailed, heteroscedastic, P = 0.005).
Clinical features
Detailed clinical assessments were available on 17 patients with definite mutations (Table 5). The most common presentation was of the behavioural variant of FTLD (12/17). The other definite mutation cases presented with speech production impairment, all with features of a PNFA (5/17). Amongst those presenting with behavioural symptoms, the most common initial feature was apathy. Other common behavioural features included abnormal eating behaviour (generally a sweet tooth) and inappropriate social behaviour with lack of insight. Some patients developed disinhibition later in the illness but this was rarely a presenting feature. Less common symptoms included loss of empathy, aggression, obsessive behaviour (including hoarding), impulsivity and hypersomnolence. One patient had tactile hallucinations of insects crawling over his skin with delusions that there were animals present in his bed to the extent that he refused to sleep in his bedroom. In this group of 12 patients, nine had decreased quantity of speech without evidence of speech errors, agrammatism or articulatory impairment. Six of these patients had become mute when last seen. This language impairment would be consistent with dynamic aphasia rather than either the PNFA or SD subtype of FTLD. Of the five patients with speech production impairment, each had features of PNFA i.e. hesitant or stuttering speech, anomia, phonemic paraphasias and poor polysyllabic word repetition. Two of these patients also had CBS with severe limb apraxia in both and in one of the cases asymmetrical parkinsonian and pyramidal signs with myoclonus. In total, 8 of the 17 patients had limb apraxia. One patient had features of both PNFA and SD being profoundly anomic with both phonemic and semantic errors in speech. None of the patients seen by us had evidence of MND although Patient 169_3 (the brother of 169_2) was said to have died of MND in his mid-50s. Forty-one per cent (7/17) of the patients had features of parkinsonism, which is a slightly lower frequency than found in other case series and may represent the fact that many of the patients are still living and may later develop parkinsonism (Josephs et al., 2007; Rademakers et al., 2007; Kelley et al., 2007). Clinical information was also available for the two cases with possible pathogenic mutations: one had a diagnosis of PNFA and the other had CBS with features of behavioural change as well including apathy, decreased speech, hyper-religiosity and also visual hallucinations.
Table 5.
Clinical features
| First symptoms | Apathy | Abnormal eating behaviour |
Loss of empathy |
Inappropriate social behaviour |
Disinhibition | Decreased quantity of speech |
Hesitant or stuttering speech |
Anomia | Phonemic paraphasias |
Mutism | Parkinsonism | Other features |
Diagnosis | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DRC32_1 | Inappropriate social behaviour Apathy |
+ | − | − | + | − | + | − | − | − | − | − | Aggression Echolalia | bvFTLD |
| DRC32_2 | Apathy | + | − | − | − | − | + | − | − | − | + | + | Echolalia Child-like/ fatuous |
bvFTLD |
| DRC32_4 | Apathy Impulsivity |
+ | − | − | + | − | − | − | − | − | − | − | Inflexibility Episodic memory impairment | bvFTLD |
| DRC32_5 | Inappropriate social behaviour Impulsivity |
− | + | − | + | + | + | − | − | − | − | − | Delusions Tactile hallucinations Hoarding Child-like/ fatuous |
bvFTLD |
| DRC39_2 | Apathy | + | − | − | − | − | − | − | + | − | − | − | Episodic memory impairment | bvFTLD |
| DRC39_3 | Inappropriate social behaviour | + | + | − | + | − | + | − | − | − | + | + | Hypersomnolence Episodic memory impairment ‘Dressing apraxia’ | bvFTLD |
| DRC169_2 | Apathy Poor concentration |
+ | − | − | − | − | + | − | − | − | + | + | Episodic memory impairment Dyscalculia | bvFTLD |
| DRC240_4 | Speech production impairment Dyscalculia Dysgraphia | + | − | − | + | − | + | − | − | + | − | − | PNFA | |
| DRC255_6 | Apathy Decreased speech Loss of empathy |
+ | + | + | + | + | + | − | − | − | − | − | Hypersomnolence | bvFTLD |
| DRC255_7 | Apathy Decreased speech Aggression Hypersomnolence |
+ | − | − | + | − | + | − | − | − | + | + | bvFTLD | |
| DRC255_8 | Apathy Decreased speech Aggression |
+ | + | + | + | + | + | − | − | − | + | + | Echolalia | bvFTLD |
| DRC255_9 | Limb apraxia Dyscalculia Speech production impairment Apathy |
+ | + | − | − | − | + | + | + | − | + | + | Asymmetrical pyramidal signs and myoclonus L>R | PNFA/CBS |
| DRC255_10 | Word-finding difficulty Apathy | + | − | − | − | − | + | − | + | + | − | − | PNFA | |
| DRC425_2 | Apathy Decreased speech Obsessive |
+ | + | + | + | − | + | − | + | − | + | + | Child-like/fatuous Episodic memory impairment | bvFTLD |
| DRC430_2 | Speech production impairment | − | + | − | − | − | + | + | + | + | + | − | PNFA/CBS | |
| DRC 431_3 | Word-finding difficulty Apathy Dysgraphia Dyslexia |
+ | − | − | − | − | − | − | + | + | − | − | Semantic errors Phonological dyslexia | Mixed PNFA/SD |
| 7651_3 | Loss of empathy Disinhibition | + | + | + | + | + | − | − | − | − | − | − | Aggression Hypersomnolence Impulsivity Inflexibility |
bvFTLD |
| Possible pathogenic mutations | ||||||||||||||
| DRC429_2 | Apathy Decreased speech |
+ | + | − | + | + | + | − | + | − | + | + | Hyperreligiosity Visual hallucinations Asymmetrical pyramidal signs and myoclonus L>R |
CBS |
| 5440_3 | Speech production impairment Word-finding difficulty | − | − | − | − | − | + | + | + | + | − | − | PNFA | |
+=present; −=absent; NK=not known; L=left; R=right.
The clinical categories of FTLD were very differently represented between GRN mutation carriers, MAPT mutation carriers and FTLD-U without GRN mutation. GRN mutation carriers were mixed with five patients diagnosed with a pure PNFA syndrome or overlap with either CBS or SD. These categories were not found in MAPT or FTLD-U without GRN mutation. Similarly, SD was over-represented in FTLD-U without GRN mutation (with all cases being sporadic), and bvFTLD was over-represented in MAPT mutation carriers (although bvFTLD is seen in all genetic and pathological subgroups). Overall, the differences were highly significant by chi-squared test (GRN versus FTLD-U without GRN mutation, 4 df, P = 0.001, GRN versus MAPT, 2 df, P = 0.006).
Neuropsychological features
Thirteen patients with definite mutations and two with possible pathogenic mutations underwent neuropsychological testing (Table 6). Of the definite mutation cases, the most common features were of executive dysfunction (8/10 tested), dyscalculia (9/12) and episodic memory impairment (9/13). Executive dysfunction is commonly seen in patients with FTLD but impairment of episodic memory is less frequently described, particularly early in the disease. Parietal lobe features are also less commonly found in patients with FTLD but in our GRN cohort all of the 13 definite mutation cases had evidence of parietal lobe dysfunction with at least one of the following: dyscalculia, limb apraxia, dysgraphia, visuospatial or visuoperceptual impairments.
Table 6.
Neuropsychological features
| Years since onset at testing |
Verbal memory impairment |
Visual memory impairment |
Executive dysfunction |
Dyslexia | Anomia | Single-word comprehension impairment |
Orofacial apraxia |
Limb apraxia |
Spelling impairment |
Dyscalculia | Visuo-spatial impairment |
Visuo-perceptual impairment |
|
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DRC32_4 | 2 | − | + | + | − | NT | − | − | − | NT | + | + | NT |
| DRC32_5 | 2 | − | − | + | − | − | − | − | + | NT | − | − | − |
| DRC39_3 | 2 | + | NT | NT | + | NT | NT | NT | NT | NT | + | + | NT |
| DRC169_2 | 3 | + | + | NT | − | NT | NT | NT | NT | NT | + | + | NT |
| DRC240_4 | 4 | − | + | + | − | − | NT | + | + | − | + | NT | − |
| DRC255_6 | 2 | + | + | + | − | − | − | − | + | − | + | + | + |
| DRC255_8 | 2 | + | + | + | + | − | + | − | + | NT | + | NT | − |
| DRC255_9 | 1 | − | + | + | + | + | − | NT | + | + | + | NT | + |
| DRC255_10 | 3 | − | − | − | − | + | − | + | + | − | − | − | − |
| DRC425_2 | 3 | + | + | + | − | + | − | − | − | − | + | − | − |
| DRC430_2 | 5 | + | + | NT | + | + | + | + | + | NT | NT | NT | + |
| DRC431_3 | 1 | − | − | − | + | + | − | − | − | + | + | − | − |
| 7651_3 | 3 | − | − | + | − | − | − | − | + | NT | − | − | − |
| Possible pathogenic mutations | |||||||||||||
| DRC429_2 | 5 | + | + | + | NT | + | − | + | + | NT | − | + | + |
| 5440_3 | 2 | + | − | + | − | − | − | − | − | NT | − | NT | − |
+=present i.e. scores at or below the 5th percentile on neuropsychological testing; −=absent; NT=not tested.
We have described the presence of parietal lobe deficits in the DRC255 GRN mutation family previously (Rohrer et al., in press). Clinical and neuropsychological examination of this larger group of GRN mutation patients confirms that the parietal lobe does seem to be involved in GRN-associated FTLD early in the disease (average years from onset of symptoms to time of testing was 2.5 with a range of 1–5). To investigate this further we compared the presence of parietal lobe deficits in this group with the rest of the cohort. Neuropsychological data was available for 121 GRN-negative patients from the cohort. We assessed for the presence of dyscalculia, limb apraxia, spelling problems and visuospatial or visuoperceptual impairments. Whilst in the GRN group 100% of the patients with definite mutations had at least one deficit attributable to parietal lobe dysfunction, in the MAPT group only 10% had deficits (1/10), whilst in the GRN-negative FTLD-U group there were 25% (4/16) (chi-squared test GRN versus MAPT, 1 d.f., P = 6.8×10−6; GRN versus FTLD-U, 1 d.f., P = 7.0×10−6). Looking at the 121 GRN-negative patients by clinical diagnosis the percentage of cases with parietal lobe deficits was 100% in CBS (15/15) with all having limb apraxia (of note this group contained patients with Goldman scores 3, 3.5 and 4 only), 32% in the bvFTLD group (18/56), 36% in SD (8/22) and 43% in PNFA (12/28). This emphasizes that there is greater parietal lobe involvement in GRN-associated FTLD than in other FTLD groups with the exception of CBS, a disorder known to involve frontal and parietal lobes.
MRI findings
Eight patients with definite GRN mutations had MR scans (Table 7). The most striking feature of the scans was the asymmetrical nature of the atrophy with either side being affected even within the same family: five had left greater than right atrophy and three had left greater than right. The other common feature between patients was the involvement of the parietal lobe consistent with the clinical and neuropsychological features of parietal lobe dysfunction described above. Figure 5 shows representative images from two of the patients. Two patients with possible pathogenic mutations also had MR scans, each showing asymmetrical fronto-temporo-parietal atrophy also, one with right greater than left and the other with the opposite pattern.
Table 7.
MR imaging features
| Handedness | Years from onset |
Asymmetry | A-P gradient | Frontal | Temporal | Parietal | Occipital | |
|---|---|---|---|---|---|---|---|---|
| DRC32_5 | R | 3 | R>L anteriorly | − | +++ | + | ++ | − |
| DRC240_4 | R | 5 | L>R | − | ++ | + | ++ | − |
| DRC255_6 | R | 2 | R>>L | − | ++ | ++ | ++ | − |
| DRC255_10 | R | 1 | L>>R | − | + | +++ | + | − |
| DRC425_2 | L | 3 | L>R | + | +++ | +++ | ++ | − |
| DRC430_2 | R | 6 | L>>R | − | ++ | ++ | ++ | − |
| DRC431_3 | R | 2 | L>>R | − | + | ++ | ++ | − |
| 7651_3 | R | 4 | R>>L | − | +++ | +++ | +++ | − |
| Possible pathogenic mutations | ||||||||
| DRC429_2 | NK | 5 | R>>L | − | +++ | +++ | +++ | − |
| 5440_3 | R | 4 | L>R | − | ++ | + | + | − |
For the individual lobes atrophy is rated as −=not present; +=mild; ++=moderate; +++=severe, and refers to the most affected side; NK=not known, A-P=antero-posterior, R=right, L=left.
Fig. 5.
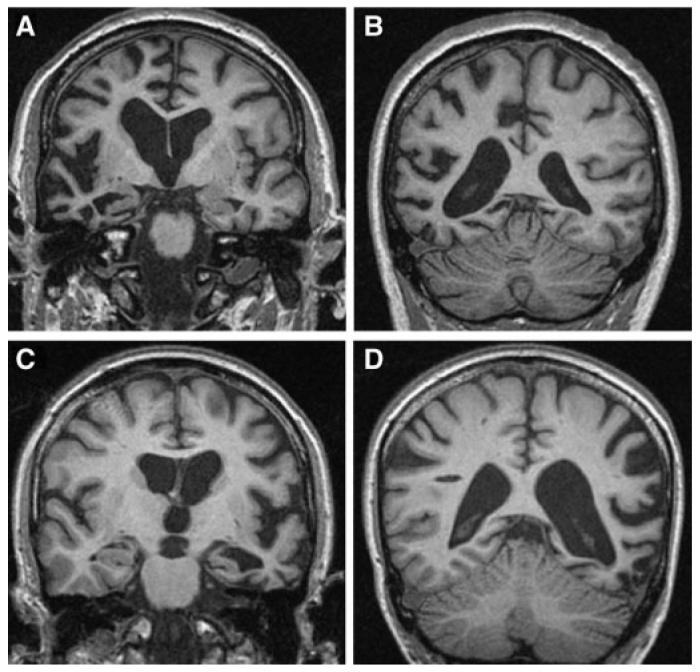
Representative brain imaging from patients with GRN mutations. (A) and (B) Coronal T1-weighted MR images showing asymmetrical right-sided fronto-temporo-parietal atrophy (Patient 7651_3)—4 years from symptom onset; (C) and (D) Coronal T1-weighted MR images showing asymmetrical left-sided fronto-temporo-parietal atrophy (Patient DRC431_3)—2 years from symptom onset.
The degree of hemispheric asymmetry was assessed by comparing the difference between the two hemisphere volumes in the GRN group with three other groups (MAPT mutation cases, GRN-negative, ubiquitin-positive cases with SD and a cognitively-normal control group). The absolute hemispheric difference was significantly greater for the GRN patients than any of the other groups (P<0.01). This difference was also greater for the ubiquitin-positive SD group compared to MAPT (P = 0.01). In fact, in the MAPT patients there was no significant left-right asymmetry (P = 0.25). All SD patients had a smaller left than right hemisphere compared to the GRN group where the predominantly atrophied hemisphere was variable. Anterior and posterior quadrant volumes were also assessed in the two hemispheres. In the SD group, the anterior/posterior (A/P) ratio was significantly different from controls on the left-hand side (P = 0.009) but not the right (P = 0.50). In the MAPT group, the A/P ratio was also significantly different from the controls on both the left (P = 0.006) and the right (P = 0.03) side. In the GRN group, the A/P ratio was not significantly different to controls on either side (left, P = 0.06; right, P = 0.67). In summary, volumetric analysis confirms the striking asymmetry of GRN cases even when compared to the canonically asymmetrical disease of SD. The analysis also shows that unlike the antero-posterior gradient seen in both SD and MAPT-associated FTLD there is no significant antero-posterior gradient in the GRN group consistent with atrophy extending more posteriorly into the parietal lobe in this group.
Other investigation findings
Eight patients with definite mutations and two with possible pathogenic mutations underwent EEG testing between 1 and 5 years from symptom onset. Alpha rhythm was present in all but one of the patients: DRC429_2 (one of the possible pathogenic mutations) had absent alpha rhythm at 5 years from symptom onset. DRC255_6 however had only traces of alpha rhythm present at 2 years from symptom onset. Other findings were non-specific with seven of the patients showing occasional theta activity.
Five patients underwent CSF analysis each showing normal constituents with matched oligoclonal bands in one patient and no bands in the other four. Two patients (DRC431_3 and DRC255_6) had samples sent for further analysis: tau levels were normal in DRC431_3 (207) but raised in DRC255_6 (943). Both had negative 14-3-3 protein.
Pathological findings
Eight patients have come to post-mortem from four different families (DRC 39_2, DRC39_3, DRC240_4, DRC240_5, DRC255_6, DRC255_7, DRC255_8 and DRC425_2) with all showing similar features. In cases where macroscopic data was available, there was involvement of the frontal, temporal and parietal lobes with sparing of the occipital lobes. There was a variable degree of spongiosis of the neuropil, neuronal loss and gliosis in the frontal and temporal cortices. In the most severely affected areas, there was also white matter pallor and gliosis. TDP-43 immunohistochemistry demonstrated numerous neuropil threads, NCI and scattered NII in affected cortical areas. In cases in which the dentate fascia was available for assessment, TDP-43 positive NCI were found in variable numbers of granule cells. The overall TDP-43 pattern corresponded to type 3 ubiquitin-positive, TDP-43-positive pathology (as per the revised consensus pathological criteria described in Cairns et al., 2007).
Discussion
We describe the clinical, neuropsychological and radiological phenotype of a GRN mutation cohort representing 12% of a large UK FTLD cohort and 20% of those FTLD subjects with a family history. 80% of these patients in four separate families had the DRC255 mutation (c.90_91 insCTGC insertion) but further analysis suggests these families have a common ancestor shared with the UBC17 family previously described (Baker et al., 2006; Mackenzie et al., 2006). Similar to the experience of other neurodegenerative diseases (Mead, 2006), it seems likely that GRN mutation frequency will vary with geography and ethnicity due to a balance of genetic drift, loss and new mutation. We found four novel pathogenic null-mutations: three frameshift indel mutations and one premature stop codon mutation. Two further missense changes were detected in patients and not in controls although families were too small to demonstrate segregation. The alanine to valine change at codon 199 is located between conserved granulin motifs. The leucine to phenylalanine change at codon 469 is located in granulin D but this residue is not conserved between motifs, indeed, the phenylalanine is found at the homologous residue in granulin motifs A and G. The location and nature of these missense changes do not support pathogenicity, but no effect on protein structure is predictable with certainty. Nonetheless, it is interesting that the phenotype of these possible mutations was similar to that seen in the definite pathogenic mutations. We did not identify splice mutations commonly found in other screening studies, or large-scale deletion of GRN involving exon 1.
GRN patients presented with bvFTLD, PNFA or CBS. No patient with a classic SD syndrome or PSP was found to have mutations. Considering the clinical presentation in more detail, GRN patients who present with behavioural symptoms commonly have apathy as the initial presenting feature although most of the common behavioural symptoms of bvFTLD are also seen during the disease. Previous case series have described language output impairment as a feature of GRN mutations (Cruts et al., 2006; Gass et al., 2006; Mesulam et al., 2007). In our study, different language phenotypes were seen within the cohort: many patients presenting with bvFTLD had decreased spontaneous speech in the absence of speech errors, consistent with dynamic aphasia, often becoming mute as the disease progressed (Snowden et al., 2006). Other patients presented with PNFA, a primary language syndrome known to be heterogeneous in its clinical presentation (Rohrer et al., 2007): some patients had hesitant, effortful speech consistent with apraxia of speech whilst others had agrammatism and phonemic errors without articulatory impairment. Some of these cases also had evidence of semantic errors and early single-word comprehension impairment, consistent with a progressive mixed aphasia (Grossman and Ash, 2004).
Seventy per cent of those undergoing neuropsychometry showed deficits on tests of episodic memory. Although patients can fail psychometric tests of memory for a number of reasons, many of these patients also complained of amnestic symptoms (Table 5) suggesting true episodic memory impairment. This feature has been found in other series of GRN patients with some patients even receiving an initial diagnosis of Alzheimer's disease (Kelley et al., 2007, Rademakers et al., 2007). This is an important aspect that should be borne in mind in the differential diagnosis of patients presenting with dementia.
A number of studies have now reported GRN mutations in association with CBS (Benussi et al., 2006; Masellis et al., 2006; Spina et al., 2007). We describe two patients with pathogenic mutations who had a CBS although both also had features of PNFA. This is a well-described syndrome overlap (Graham et al., 2003) although the underlying pathology in many previously reported cases has been tau-positive corticobasal degeneration (CBD) pathology (Knopman et al., 2005). CBS is classically described as involving the frontal and parietal lobes both clinically and radiologically, differing from other FTLD syndromes with its more posterior cerebral involvement. In fact, despite heterogeneity by established clinical criteria, all of the patients in our series had evidence of early parietal lobe dysfunction. This has been noted by us and other groups previously (Le Ber et al., 2007; Rademakers et al., 2007; Spina et al., 2007; Rohrer et al., in press) and study of this large FTLD cohort confirms the involvement of the parietal lobes early in GRN-associated FTLD with much lesser involvement in other genetic and clinical subgroups (except in cases of CBS).
Consistent with previous reports, all of the patients who have come to post-mortem have shown TDP-43/ubiquitin-positive type 3 pathology with the presence of neuronal intranuclear inclusions. Four other patients in the FTLD series (from two families DRC306 and DRC321) were known to have the same pathological findings but were found to be GRN negative. One recent study suggested that only about 60% of type 3 pathology cases were GRN-positive with the other 40% being mainly autosomal dominant familial cases without a known genetic mutation (Josephs et al., 2007). One member from each of our families had volumetric MR brain imaging and it is interesting to note that both scans showed asymmetrical left-sided predominant atrophy with parietal lobe involvement. This intriguing finding suggests that the GRN phenotype we have described may be a feature of type 3 TDP-43/ubiquitin-positive pathology rather than GRN mutations per se.
The radiological findings support our clinical and neuropsychological findings of parietal lobe involvement with all of the patients showing fronto-temporo-parietal atrophy. Other studies have also shown early parietal lobe atrophy in some patients (Kelley et al., 2007, Spina et al., 2007) and a more detailed imaging study using voxel-based morphometry showed greater parietal lobe atrophy in a GRN mutation group compared to a ubiquitin-positive, GRN-negative group (Whitwell et al., 2007). The further feature of GRN cases that distinguishes them from other FTLD cases is the striking asymmetrical atrophy (and something that will not necessarily be picked up with voxel-based morphometry if there is variable laterality within the group) (Kelley et al., 2007). Volumetric analysis showed that the GRN cases as a group were even more asymmetrical than the canonically asymmetrical subtype of FTLD, SD (Chan et al., 2001). The mechanism for this asymmetry is unclear but suggests focal rather than diffuse onset: involvement of fronto-parietal pathways that are preferentially affected on one side first before affecting the opposite side.
The duration of disease appears to be shorter for patients with GRN mutations (average 5 years) compared to other FTLD patients studied here, although a weakness of this study is the lack of FTD-MND cases, the particular subgroup of FTLD known to have short duration [a mean of ∼3 years (Hodges et al., 2003)]. One study found that GRN patients have more generalized atrophy and smaller brains at post-mortem compared to a group of GRN-negative patients and therefore suggested that GRN results in a more rapid, ‘malignant’ form of FTLD (Josephs et al., 2007). This finding of smaller brains at post-mortem could also result from the fact that with early involvement of the parietal lobe more of the brain is involved in GRN disease compared with other FTLD groups where atrophy is confined to a large extent to the frontal and temporal lobes. Longitudinal studies of the rate of atrophy in GRN cases are needed to help to answer the question of whether the short duration of disease is associated with a higher rate of atrophy compared to other FTLD subgroups.
Although there was a suggestion of MND in an unexamined GRN mutation relative, this was not a feature of the well-studied cases and no GRN mutations were found by screening a UK cohort of non-SOD1 familial MND. This is consistent with other studies suggesting that GRN is not a common cause of a MND phenotype (Gass et al., 2006; Schymick et al., 2007).
In concordance with other reports (et al., 2006) we found evidence that APOE-E4 genotype delays age of clinical onset by around 6 years. Our sample was small so this result is not statistically robust, and a large study of the Arg493X mutation did not find such an association (Rademakers et al., 2007). APOE is thought to have widespread effects on brain metabolism (Scarmeas et al., 2005) and neuroanatomy (Shaw et al., 2006), and as our understanding of the pathogenesis of GRN mutation is at an early stage, it would be premature to speculate on a mechanism.
The distinct GRN phenotype of our series does not necessarily imply an effective management strategy for genetic testing. The most statistically sensitive but costly approach would be to sequence GRN in all patients with FTLD. Indeed, technical improvements in high-throughput sequencing may soon translate into diagnostic genetic testing at a much reduced cost. Although a positive family history, parietal lobe signs and asymmetrical MRI brain scan were found in the majority of GRN cases, these features were not necessarily found at presentation when a decision about genetic testing is usually made. Our series was not large or detailed enough to define the benefits of using these characteristics to filter cases.
In conclusion, patients with GRN mutations: have a shorter disease duration compared to other FTLD groups (with the exception of FTD-MND); present with bvFTLD (commonly apathy initially), language output impairment (either a PNFA or decreased speech output consistent with a dynamic aphasia) or with CBS; have parietal lobe dysfunction seen particularly on neuropsychological testing; and commonly have evidence of asymmetrical atrophy on MR imaging with the frontal, temporal and parietal lobes all affected.
Acknowledgements
This work was funded by the Alzheimer Research Trust and the Medical Research Council UK and by an EC Contract awarded to the APOPIS Consortium. J.D.R. is supported by a Wellcome Trust Research Training Fellowship. J.D.W. is supported by a Wellcome Trust Intermediate Clinical Fellowship. This work was undertaken at UCLH/UCL who received a proportion of funding from the Department of Health's NIHR Biomedical Research Centres funding scheme. We are grateful to the MRC London Neurodegenerative Brain Bank at the Institute of Psychiatry for providing brain tissue for analysis. We are very grateful also for the generous contribution of the patients and their relatives. Funding to pay the Open Access publication charges for this article was provided by MRC.
Abbreviations
- bvFTLD
behavioural variant FTLD
- CBD
corticobasal degeneration
- CBS
corticobasal syndrome
- CSF
cerebrospinal fluid
- FTD
frontotemporal dementia
- FTLD
frontotemporal lobar degeneration
- MND
motoneuron disease
- NII
neuronal intranuclear inclusion
- NCI
neuronal intracytoplasmic inclusion
- PCR
polymerase chain reaction
- PNFA
progressive non-fluent aphasia
- PSP
progressive supranuclear palsy
- SD
semantic dementia
References
- Baker M, Mackenzie IR, Pickering-Brown SM, Gass J, Rademakers R, Lindholm C, et al. Mutations in progranulin cause tau-negative frontotemporal dementia linked to chromosome 17. Nature. 2006;442:916–9. doi: 10.1038/nature05016. Epub 2006 Jul 16. [DOI] [PubMed] [Google Scholar]
- Baxter DM, Warrington EK. Measuring dysgraphia: a graded-difficulty spelling test. Behav Neurol. 1994;7:107–16. doi: 10.3233/BEN-1994-73-401. Epub 2006 Jul 16. [DOI] [PubMed] [Google Scholar]
- Behrens MI, Mukherjee O, Tu PH, Liscic RM, Grinberg LT, Carter D, et al. Neuropathologic heterogeneity in HDDD1: a familial frontotemporal lobar degeneration with ubiquitin-positive inclusions and progranulin mutation. Alzheimer Dis Assoc Disord. 2007;21:1–7. doi: 10.1097/WAD.0b013e31803083f2. [DOI] [PubMed] [Google Scholar]
- Benussi L, Binetti G, Sina E, Gigola L, Bettecken T, Meitinger T, et al. A novel deletion in progranulin gene is associated with FTDP-17 and CBS. Neurobiol Aging. 2006 doi: 10.1016/j.neurobiolaging.2006.10.028. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Bird T, Knopman D, VanSwieten J, Rosso S, Feldman H, Tanabe H, et al. Epidemiology and genetics of frontotemporal dementia/Pick's disease. Ann Neurol. 2003;54:S29–31. doi: 10.1002/ana.10572. [DOI] [PubMed] [Google Scholar]
- Boeve BF, Lang AE, Litvan I. Corticobasal degeneration and its relationship to progressive supranuclear palsy and frontotemporal dementia. Ann Neurol. 2003;54:S15–19. doi: 10.1002/ana.10570. [DOI] [PubMed] [Google Scholar]
- Bronner IF, Rizzu P, Seelaar H, van Mil SE, Anar B, Azmani A, et al. Progranulin mutations in Dutch familial frontotemporal lobar degeneration. Eur J Hum Genet. 2007;15:369–74. doi: 10.1038/sj.ejhg.5201772. [DOI] [PubMed] [Google Scholar]
- Brun A, Englund B, Gustafson L, Passant U, Mann DMA, Neary D, et al. Clinical and neuropathological criteria for frontotemporal dementia. J Neurol Neurosurg Psychiatry. 1994;57:416–8. doi: 10.1136/jnnp.57.4.416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cairns NJ, Bigio EH, Mackenzie IR, Neumann M, Lee VM, Hatanpaa KJ, et al. Neuropathologic diagnostic and nosologic criteria for frontotemporal lobar degeneration: consensus of the Consortium for Frontotemporal Lobar Degeneration. Acta Neuropathol (Berl) 2007;114:5–22. doi: 10.1007/s00401-007-0237-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan D, Fox NC, Scahill RI, Crum WR, Whitwell JL, Leschziner G, et al. Patterns of temporal lobe atrophy in semantic dementia and Alzheimer's disease. Ann Neurol. 2001;49:433–42. [PubMed] [Google Scholar]
- Cruts M, Gijselinck I, van der Zee J, Engelborghs S, Wils H, Pirici D, et al. Null mutations in progranulin cause ubiquitin-positive frontotemporal dementia linked to chromosome 17q21. Nature. 2006;442:920–4. doi: 10.1038/nature05017. [DOI] [PubMed] [Google Scholar]
- Davion S, Johnson N, Weintraub S, Mesulam M, Cairns N, Engberg A, et al. Clinicopathologic correlation in progranulin mutations. Mod Pathol. 2007;20:297A. doi: 10.1212/01.wnl.0000267701.58488.69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksen JL, Mackenzie IR. Progranulin: normal function and role in neurodegeneration. J Neurochem. 2007 doi: 10.1111/j.1471-4159.2007.04968.x. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Freeborough PA, Fox NC, Kitney RI. Interactive algorithms for the segmentation and quantitation of 3-D MRI brain scans. Comput MethodsProg Biomed. 1997;53:15–25. doi: 10.1016/s0169-2607(97)01803-8. [DOI] [PubMed] [Google Scholar]
- Gass J, Cannon A, Mackenzie IR, Boeve B, Baker M, Adamson J, et al. Mutations in progranulin are a major cause of ubiquitin-positive frontotemporal lobar degeneration. Hum Mol Genet. 2006;15:2988–3001. doi: 10.1093/hmg/ddl241. [DOI] [PubMed] [Google Scholar]
- Goldman JS, Farmer JM, Wood EM, Johnson JK, Boxer A, Neuhaus J, et al. Comparison of family histories in FTLD subtypes and related tauopathies. Neurology. 2005;65:1817–19. doi: 10.1212/01.wnl.0000187068.92184.63. [DOI] [PubMed] [Google Scholar]
- Graham NL, Bak T, Patterson K, Hodges JR. Language function and dysfunction in corticobasal degeneration. Neurology. 2003;61:493–9. doi: 10.1212/01.wnl.0000081230.09863.ed. [DOI] [PubMed] [Google Scholar]
- Grossman M, Ash S. Primary progressive aphasia: a review. Neurocase. 2004;10:3–18. doi: 10.1080/13554790490960440. [DOI] [PubMed] [Google Scholar]
- Hodges JR, Davies RR, Xuereb JH, Casey B, Broe M, Bak TH, Kril JJ, Halliday GM. Clinicopathological correlates in frontotemporal dementia. Ann Neurol. 2004;56:399–406. doi: 10.1002/ana.20203. [DOI] [PubMed] [Google Scholar]
- Hodges JR, Davies R, Xuereb J, Kril J, Halliday G. Survival in frontotemporal dementia. Neurology. 2003;61:349–54. doi: 10.1212/01.wnl.0000078928.20107.52. [DOI] [PubMed] [Google Scholar]
- Hutton M, Lendon CL, Rizzu P, Baker M, Froelich S, Houlden H, et al. Association of missense and 5′-splice-site mutations in tau with the inherited dementia FTDP-17. Nature. 1998;393:702–5. doi: 10.1038/31508. [DOI] [PubMed] [Google Scholar]
- Jackson M, Warrington EK. Arithmetic skills in patients with unilateral cerebral lesions. Cortex. 1986;22:611–20. doi: 10.1016/s0010-9452(86)80020-x. [DOI] [PubMed] [Google Scholar]
- Josephs KA, Ahmed Z, Katsuse O, Parisi JF, Boeve BF, Knopman DS, et al. Neuropathologic features of frontotemporal lobar degeneration with ubiquitin-positive inclusions with progranulin gene (PGRN) mutations. J Neuropathol Exp Neurol. 2007;66:142–51. doi: 10.1097/nen.0b013e31803020cf. [DOI] [PubMed] [Google Scholar]
- Kelley BJ, Haidar W, Boeve BF, Baker M, Graff-Radford NR, Krefft T, et al. Prominent phenotypic variability associated with mutations in Progranulin. Neurobiol Aging. 2007 doi: 10.1016/j.neurobiolaging.2007.08.022. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kertesz A, Davidson W, Munoz DG. Clinical and pathological overlap between frontotemporal dementia, primary progressive aphasia and corticobasal degeneration: the Pick complex. Dement Geriatr Cogn Disord. 1999;10:46–9. doi: 10.1159/000051212. [DOI] [PubMed] [Google Scholar]
- Knopman DS, Boeve BF, Parisi JE, Dickson DW, Smith GE, Ivnik RJ, et al. Antemortem diagnosis of frontotemporal lobar degeneration. Ann Neurol. 2005;57:480–8. doi: 10.1002/ana.20425. [DOI] [PubMed] [Google Scholar]
- Le Ber I, Camuzat A, van der Zee J, Hannequin D, Pasquier F, Gijselinck I, et al. PGRN gene mutations in both sporadic and familial frontotemporal dementia. Neurology. 2007;68:A351. doi: 10.1002/humu.20520. [DOI] [PubMed] [Google Scholar]
- Lendon CL, Lynch T, Norton J, Mckeel DW, Busfield F, Craddock N, et al. Hereditary dysphasic disinhibition dementia - a frontotemporal dementia linked to 17q21-22. Neurology. 1998;50:1546–55. doi: 10.1212/wnl.50.6.1546. [DOI] [PubMed] [Google Scholar]
- Leverenz JB, Yu CE, Montine TJ, Steinbart E, Bekris LM, Zabetian C, et al. A novel progranulin mutation associated with variable clinical presentation and tau, TDP43 and alpha-synuclein pathology. Brain. 2007;130:1360–74. doi: 10.1093/brain/awm069. [DOI] [PubMed] [Google Scholar]
- Litvan I, Agid Y, Calne D, Campbell G, Dubois B, Duvoisin RC, et al. Clinical research criteria for the diagnosis of progressive supranuclear palsy (Steele–Richardson–Olszewski syndrome): report of the NINDS-SPSP International Workshop. Neurology. 1996;47:1–9. doi: 10.1212/wnl.47.1.1. [DOI] [PubMed] [Google Scholar]
- Lomen-Hoerth C, Anderson T, Miller B. The overlap of amyotrophic lateral sclerosis and frontotemporal dementia. Neurology. 2002;59:1077–9. doi: 10.1212/wnl.59.7.1077. [DOI] [PubMed] [Google Scholar]
- Mackenzie IR, Baker M, West G, Woulfe J, Qadi N, Gass J, et al. A family with tau-negative frontotemporal dementia and neuronal intranuclear inclusions linked to chromosome 17. Brain. 2006;129:853–67. doi: 10.1093/brain/awh724. [DOI] [PubMed] [Google Scholar]
- Masellis M, Momeni P, Meschino W, Heffner R, Elder J, Sato C, et al. Novel splicing mutation in the progranulin gene causing familial corticobasal syndrome. Brain. 2006;129:3115–23. doi: 10.1093/brain/awl276. [DOI] [PubMed] [Google Scholar]
- Mazziotta JC, Toga AW, Evans A, Fox P, Lancaster J. A Probabilistic atlas of the human brain - theory and rationale for its development. Neuroimage. 1995;2:89–101. doi: 10.1006/nimg.1995.1012. [DOI] [PubMed] [Google Scholar]
- McKenna P, Warrington EK. Testing for nominal dysphasia. J Neurol Neurosurg Psychiatry. 1980;43:781–8. doi: 10.1136/jnnp.43.9.781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mead S. Prion disease genetics. Eur J Hum Genet. 2006;14:273–81. doi: 10.1038/sj.ejhg.5201544. [DOI] [PubMed] [Google Scholar]
- Mesulam M, Johnson N, Krefft TA, Gass JM, Cannon AD, Adamson JL, et al. Progranulin mutations in primary progressive aphasia - the PPA1 and PPA3 families. Arch Neurol. 2007;64:43–7. doi: 10.1001/archneur.64.1.43. [DOI] [PubMed] [Google Scholar]
- Mukherjee O, Pastor P, Cairns NJ, Chakraverty S, Kauwe JSK, Shears S, et al. HDDD2 is a familial frontotemporal lobar degeneration with ubiquitin-positive, tau-negative inclusions caused by a missense mutation in the signal peptide of progranulin. Ann Neurol. 2006;60:314–322. doi: 10.1002/ana.20963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neary D, Snowden JS, Gustafson L, Passant U, Stuss D, Black S, et al. Frontotemporal lobar degeneration - a consensus on clinical diagnostic criteria. Neurology. 1998;51:1546–54. doi: 10.1212/wnl.51.6.1546. [DOI] [PubMed] [Google Scholar]
- Nelson HE. A modified card sorting test sensitive to frontal lobe defects. Cortex. 1976;12:313–24. doi: 10.1016/s0010-9452(76)80035-4. [DOI] [PubMed] [Google Scholar]
- Nelson HE. National Adult Reading Test (NART): test manual. UK: NFER-NELSON; Windsor: 1982. [Google Scholar]
- Nelson HE, McKenna P. The use of current reading ability in the assessment of dementia. Br J Soc Clin Psychol. 1975;14:259–67. doi: 10.1111/j.2044-8260.1975.tb00178.x. [DOI] [PubMed] [Google Scholar]
- Neumann M, Sampathu DM, Kwong LK, Truax AC, Micsenyi MC, Chou TT, et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006;314:130–3. doi: 10.1126/science.1134108. [DOI] [PubMed] [Google Scholar]
- Oldfield RC, Wingfield A. Response latencies in naming objects. Q J Exp Psychol (series A) 1965;17:273–81. doi: 10.1080/17470216508416445. [DOI] [PubMed] [Google Scholar]
- Poorkaj P, Bird TD, Wijsman E, Nemens E, Garruto RM, Anderson L, et al. Tau is a candidate gene for chromosome 17 frontotemporal dementia. Ann Neurol. 1998;43:815–25. doi: 10.1002/ana.410430617. Erratum in: Ann Neurol 1998; 44:428. [DOI] [PubMed] [Google Scholar]
- Rademakers R, Cruts M, Dermaut B, Sleegers K, Rosso SM, Van Den BM, et al. Tau negative frontal lobe dementia at 17q21: significant fine-mapping of the candidate region to a 4.8 cM interval. Mol Psychiatry. 2002;7:1064–74. doi: 10.1038/sj.mp.4001198. [DOI] [PubMed] [Google Scholar]
- Rademakers R, Baker M, Gass J, Adamson J, Huey ED, Momeni P, et al. Phenotypic variability associated with progranulin haploinsufficiency in patients with the common 1477C–>T (Arg493X) mutation: an international initiative. Lancet Neurol. 2007;6:857–68. doi: 10.1016/S1474-4422(07)70221-1. [DOI] [PubMed] [Google Scholar]
- Rohrer JD, Knight WD, Warren JE, Fox NC, Rossor MN, Warren JD. Word-finding difficulty: a clinical analysis of the progressive aphasias. Brain. 2008;131(Pt 1):8–38. doi: 10.1093/brain/awm251. Epub 2007 Oct 18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rohrer JD, Warren JD, Omar R, Mead S, Beck J, Revesz T, et al. Parietal lobe deficits are a feature of frontotemporal lobar degeneration caused by a mutation in the progranulin gene. Arch Neurol. doi: 10.1001/archneur.65.4.506. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosso SM, Kamphorst W, De Graaf B, Willemsen R, Ravid R, Niermeijer MF, et al. Familial frontotemporal dementia with ubiquitin-positive inclusions is linked to chromosome 17q2l-22. Brain. 2001;124:1948–57. doi: 10.1093/brain/124.10.1948. [DOI] [PubMed] [Google Scholar]
- Scarmeas N, Habeck CG, Hilton J, Anderson KE, Flynn J, Park A, et al. APOE related alterations in cerebral activation even at college age. J Neurol Neurosurg Psychiatry. 2005;76:1440–44. doi: 10.1136/jnnp.2004.053645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schymick JC, Yang Y, Andersen PM, Vonsattel JP, Greenway M, Momeni P, et al. Progranulin mutations and amyotrophic lateral sclerosis or amyotrophic lateral sclerosis-frontotemporal dementia phenotypes. J Neurol Neurosurg Psychiatry. 2007;78:754–6. doi: 10.1136/jnnp.2006.109553. Epub 2007 Mar 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw P, Lerch J, Taylor K, Seal J, Evans A, Rapoport J, et al. Cognitively intact children carrying the apolipoprotein E epsilon (E)4 allele have a thinner mesial temporal cortex. Am J Med Genet B-Neuropsychiatr Genet. 2006;141B:768. [Google Scholar]
- Skibinski G, Parkinson N, Brown J, Chakrabarti L, Lloyd S, Hummerich H, et al. Mutations in endosomal ESCRTlll-complex subunit CHMP2B in frontotemporal dementia. Nat Genet. 2005;17:806–8. doi: 10.1038/ng1609. [DOI] [PubMed] [Google Scholar]
- Snowden JS, Pickering-Brown SM, Mackenzie IR, Richardson AM, Varma A, Neary D, et al. Progranulin gene mutations associated with frontotemporal dementia and progressive non-fluent aphasia. Brain. 2006;129(Pt 11):3091–102. doi: 10.1093/brain/awl267. Epub 2006 Sep 26. [DOI] [PubMed] [Google Scholar]
- Snowden J, Neary D, Mann D. Frontotemporal lobar degeneration: clinical and pathological relationships. Acta Neuropathol (Berl) 2007;114:31–8. doi: 10.1007/s00401-007-0236-3. Epub 2007 Jun 14. [DOI] [PubMed] [Google Scholar]
- Spillantini MG, Murrell JR, Goedert M, Farlow MR, Klug A, Ghetti B. Mutation in the tau gene in familial multiple system tauopathy with presenile dementia. Proc Natl Acad Sci USA. 1998;95:7737–41. doi: 10.1073/pnas.95.13.7737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spina S, Murrell JR, Huey ED, Wassermann EM, Pietrini P, Grafman J, et al. Corticobasal syndrome associated with the A9D Progranulin mutation. J Neuropathol Exp Neurol. 2007;66:892–900. doi: 10.1097/nen.0b013e3181567873. [DOI] [PubMed] [Google Scholar]
- Stevens M, van Duijn CM, Kamphorst W, de Knijff P, Heutink P, van Gool WA, et al. Familial aggregation in frontotemporal dementia. Neurology. 1998;50:1541–5. doi: 10.1212/wnl.50.6.1541. [DOI] [PubMed] [Google Scholar]
- van der Zee J, Rademakers R, Engelborghs S, Gijselinck I, Bogaerts V, Vandenberghe R, et al. A Belgian ancestral haplotype harbours a highly prevalent mutation for 17q21-linked tau-negative FTLD. Brain. 2006;129:841–52. doi: 10.1093/brain/awl029. [DOI] [PubMed] [Google Scholar]
- Warrington EK. Manual for the recognition memory test for words and faces. NFER-Nelson; Windsor, UK: 1984. [Google Scholar]
- Warrington EK. The Camden memory tests manual. Psychology Press; Hove, UK: 1996. [Google Scholar]
- Warrington EK, James M. The visual object and space perception battery. Thames Valley Test Company; Bury St Edmunds, UK: 1991. [Google Scholar]
- Warrington EK, McKenna P, Orpwood L. Single word comprehension: a concrete and abstract word synonym test. Neuropsychol Rehabil. 1998;8:143–54. [Google Scholar]
- Watts GD, Wymer J, Kovach MJ, Mehta SG, Mumm S, Darvish D, et al. Inclusion body myopathy associated with Paget disease of bone and frontotemporal dementia is caused by mutant valosin-containing protein. Nat Genet. 2004;36:377–81. doi: 10.1038/ng1332. [DOI] [PubMed] [Google Scholar]
- Wechsler D. Manual for the Wechsler Adult Intelligence Scale. Psychological Corporation; New York: 1981. Revised. [Google Scholar]
- Weigl E. On the psychology of the so called process of abstraction. J Abnorm Soc Psychol. 1948;36:3–33. [Google Scholar]
- Whitwell JL, Jack CR, Baker M, Rademakers R, Adamson J, Boeve BF, et al. Voxel-based morphometry in frontotemporal lobar degeneration with ubiquitin-positive inclusions with and without progranulin mutations. Arch Neurol. 2007;64:371–6. doi: 10.1001/archneur.64.3.371. [DOI] [PMC free article] [PubMed] [Google Scholar]